NII-Electronic Library Service τhc∫
ロ
P砌ε5¢Jeurnai of PSJ伽加 n寉‘‘s[ien‘e 2013、
VoL 32,
No,
1,
101−
119講演
論 文
精
神
疾患
の
中間表現 型
と
し
て の
未 成熟歯 状
回
* ’昌
子
浩
孝
a’
b
*2 ・萩
原
英
雄
a’
b ・高
雄
啓
三 a’
b’
c・
Noah
M
.
WALTONC
・松
本
光
之
c ・宮
川
岡
u
a,
b,
c a 藤田保健 衛生大学 総合医科学研究所シ ステム医科 学 研 究 部 門,
b科 学 技術振興 機構 戦 略的創造研究推進事業 (CREST )
,
C 生理学研究所 行 動・
代 謝 分 子解 析セ ン ター
行 動 様 式 解 析 室, dCNS
,
AstellasResearch
lnstitute
of America LL(:Immature
dentate
gyrus
*1
A
potential
new endophenotype ofpsychiatric
disorders
Hirotaka
SHoJia
・
b
*2,
H
江deo
HAGIHARAa
・
b
,
Keizo
TAKAoa
・
b・
c ,N
・ahM
肱 LT・Nc,
Mit
・u四ki
M
… uM … G,・ndT
・uy ・ ・hi
・MlyAKAwAa
,
以 ‘aDi ・跏 嚇 ・t・m・M 眦 ・1 s伽 ・e
,
・ln・鰍 ゆ ・C
・mp ・・h
・n・i・eM ・di
・al s・i・鵬 F卿 H・肅 σ厩 ・吻 bCREST,
Japan
S‘ience and Technology Agenc7,
CSe
伽 n ofBehavior
Patterns
,伽 剛br
G8
”eti‘A鰕 ケ∫f5〔ゾBehavior.
Natio砌1
lnstitutゆr恥 5ゴological S‘勧 ‘es.
d
CN5
,
Astetlas Research lnstitute(ifAmerica
LLCMaturation and integration ofneurons and grial cells in the hippocampus is considered to
be
essentialfor
regu−
lating
endocrine,
affective,
and congnitive functions,
and the disruptiQn of such process maycause mental illness
.
Previousl矜 we have reported that mice heterozygous for a nul mutation inα
一
CaMKII,
which has a key role in a synaptic plasticity,
show abnormal behaviors related to psychiatri⊂disorders,
such as sChizophrenia andbipolar
disorder
.
ln these rnutant mice,
almost all neurons in thedentate
gyrus
are at apseudo−
immature properties,
whihc we referred to as“
immature dentategyrus
(iDG ),
”
ユb date
,
the iDGphenotype
havebeen
found in mustant strainsincluding Schnurri
−
2 kneckout,
SNAP−
25 mutant,
and forebrain−
spec盗¢ calcineurinknockout
mice which showsimilar
behavioral
phenotアpes,
In
addition,
both
chronicfluoxetine
treatment and pilocarpine−
induced seizures canreverse the maturation state of the mature neurons
,
resulting in the iDG phenotype in wild−
type mice,
Such iDG−
likephenomenon was observed in the
brains
from
patients
with schizQphrenia 〆bipolar disordeL Based on the findings,
we
proposed
that
the iDG is a potential new endophenotype of neuropsychiatricdisorders
,
This review summarizesthe
behavioral
abnor 皿alities,
iDGphenotype
,
and the implications in the pathophysiology of neuropsychiatricdisorders
,
Key word8 :genetically engineered mice
,
behavioral test battery,
working memory,
dentategyrus
,
immaturegranule cell
,
neural plasticity1
。
は じ め に*lThis article can be d正stributed under the Creative Com
−
mons Attribution License (CC BY 3
.
0),
which permitsunrestricted use
,
distribution,
and reproduGtion in anymedium
,
provided the original werk is properly cited.
*2Corresponding
author Division of Systems Medical Sci
−
ence
,
Institute fer Comprehens正ve Medical Science,
FujitaHealth University
,
1−
98 Dengakugakubo,
Kutsukake−
cho,
Toアoake , Aichi 470
−
1192,Japan
.
E−
ma 重1
:hshoji
@fu
,ita−
hu
・
ac・
jp
近 年
,
我が国 に お け る精神 疾 患の患者数は320万 人に上る と推 定さ れ (厚 生 労 働 省 「平 成23年 患 者 調 査」)
,
その生 涯 有 病 率は
,
統 合 失 調 症で約1%,
双極 性 障害で 0.
3〜
1.
4% と なっ てい る (Goldner,
Hsu,
Waraich,
& Somers,
2002;Goodwin &
Jamison
,
2007;Kendler,
Gallagher,
Abelson,
& Kessler
,
1996; McGrath,
Saha,
Chant,
& Welham,
2008)。
精 神 疾 患は
,
決 して珍 しい病 気で は な く,
日常生活に障害を も た ら し
,
自殺の原 因とな る ほか,
労 働 力の低 下 や102 基 礎 心理学 研 究 第32巻 第1号 医 療 費の増 加な どに よ る社会的損失を招く結果となるた め, その根 底にある こ ころの問 題の琿解と適 切な予 防
・
治 療に向け た取 り組みが急 務の課 題となっ て い る。 しか し な が ら,
精 神 疾 患 が な ぜ,
どの よ う に して発症す るの か,
また精 神 疾 患 を抱 える人の脳の中で は何が起こっ て い るの か,
につ い てすら未だ よくわかっ てい ないの が現 状である。 今 日,
精 神 疾 患の分 類と診 断は,
主に国 際 疾 病 分 類で あ るICD−
10や米国精 神医学 会が刊 行 す るDSM−
IVの基 準に従っ ており.
患者が訴え た症状が その分類の症 状 項 目とどの程度一
致し,
どの く らい持 続して い る の か に基 づい て行われ る。
例 え ば,
統 合 失 調 症は,
陽 性 症 状 (幻 覚,
妄想,
解体し た行動,
精 神 運 動 興 奮など)や陰 性 症 状 (感 情の平 板 化,
思 考の貧 困,
意 欲の欠 如,
社 会 的機 能の低 下など), 認 知 障 害 (記 憶・
注 意 機 能の低 下 ) な どの症 状を特徴 とし,
双極 性 障 害は うつ 状 態と躁 状 態 (気 分の高 揚,
注 意 散 漫,
睡 眠の低 下,
幻 覚・
妄 想な ど) が繰 り返 し経 験さ れるこ とを主症状と して診 断が行わ れ る。
こ の ように,
精 神 疾 患は症 状に基づく分 類と診 断が 行わ れ て き た が, 最 近の大 規 模な人 類 遺 伝 学 的 研 究か ら,
統 合 失 調 症と双極 性 障 害に は共 通の遺 伝因 子 が ある こ と が示 唆さ れて お り (Lichtenstein et al.
,
2009),
生物 学 的 な観 点から見て,
両 疾 患 を別 々の疾 患として扱 うこ と が 果 た して妥当なのかとい う 問 題 が生じて き た。
今 後,
精 神 疾 患の分 類・
診 断 を行 うた め に は,
発 症 原 因や客 観 的 な 基 準となる生 物 学 的 指 標に基づい て精 神 疾 患 を捉え 直すこと が必 要で あ る と考え られ る。
そ の た め に は, 精 神 疾 患の発 症 原因 を追 求し,
脳の 中で何が生じ てい る の か を明らか にする こ と が重要で ある。 我 々の研 究 室で は, 過 去十 年ほどにわ たっ て,
国 内外 の 100以 上の 研 究 室との共 同 研 究に より,
160系 統 以 上 の遺 伝了政 変マ ウスや薬 物 投 与マ ウス の行動を解 析し た 結果,
統 合失調症や双極性 障害を は じ めとす る精 神・
神 経 疾 患 患 者の症 状に類 似した行 動 異 常の パ ター
ンを示 す マ ウス系統を多数見出して き た。 その中で も特に顕 著な 行動異常を示 す複数 系統のマ ウス の脳を調べ たと ころ,
成 体の脳で あ るに も関わ らず 海 馬の歯 状回 と い う部 位の 神 経 細 胞の ほ と んどが擬 似 的に未 成 熟な状 態にある とい う驚くべ き現 象 (「未 成熟 歯状回」)を発見し た。 ま た,
正常なマ ウス で も抗 うつ 薬 (Kobayashi et aL,
2010) やて ん か ん症 状を誘 発 する ヒ゜
ロ カ ル ピン (Shin et al.
,
2013) を投 与す るこ と に よっ て未 成 熟 歯 状同 に酷 似し た 現象を 誘 導す るこ と が口∫能で あ るこ と も わ かっ て き た 。 さ ら に,
こ の未 成熟歯状回が生じてい るこ とを示 唆 する分 了 レ ベ ル の変 化が,
マ ウス の み な らず,
統 合 失 調 症 患 者や 双極 性 障 害 患 者の死 後 脳で も確 認 されつ つ ある (Walton et al.
,
2012)。
我々 は,
未 成 熟 歯 状同が精 神 疾 患の中 間 表 現 型[ と して,
今後,
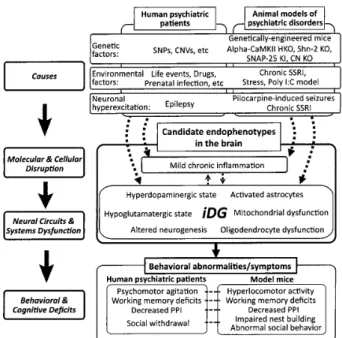
診 断法や治療法を開発す る上で重 要な研 究 対 象になる の で は ない か と考えて い る。
本稿で は,
未成 熟歯状回とい う現象を発見し た経 緯とその背 景 にあると推 測 されるメカニ ズムや 脳機 能につ いて紹介す る。 な お,
本稿は,
オー
プン ァクセス ジャー
ナル 「NeuralPlasticity」(vQL 2013
,
Article ID 318596,
doi
:1 ユ155120131318596)に掲 載さ れ たHagihara
,
Takao,
Walton,
Matsumoto,
&Miyakawa (2013)の総 説 論 文 「lrnmature Dentate Gyrus:
An Endophenetype ofNeuropsychiatric Disorders」 (Creative
Commons attribution 3
.
unported license)を翻 訳 し,
改変 した もの である。 2
.
「未成熟歯状
回」の発見
精 神 疾 患の 病 因・
病 態 を解 明し,
適 切 な 治 療 法 を開 発 するた めには,
実験動 物を用い た基 礎 的研究が不 可欠で ある。 従来か ら,
精神疾 患の研究領 域で は, 様々 な薬 物 やス ト レ スな ど をラ ッ トやマ ウス に与え,
精神疾患のモ デル動 物 を 開 発 する試みが続けられて きた。 近 年で は,
ヒ トの遺 伝 学 的 解 析か ら同 定さ れ た精 神 疾 患の関連 遺 伝 子と相 同な遺伝 子を欠損し たマ ウスや変異 型の遺 伝 子を 導 入 したマ ウスが作 製 されるよ うにな り,
それ らを利用 し て精 神 疾 患の病因・
病 態の解 明が進め ら れてい る。
そ の一
方で,
1990年代に は特定の遺伝 子を操作す ることが 可 能 な遺 伝 子工学 技 術や分 子生物 学的于法が発 展し たこ と に より, 遺 伝 子 改 変マ ウス を用いた心 理 学 的,
神 経 科 学 的 研 究が盛ん に行わ れ る よ う に なっ た。
我々 の研 究 室 で は,
様々 な遺 伝了改変マ ウスを対 象に,
何ら かの仮説 に基づい て特 定の行 動 表 現 型に つ い て解 析 す る だ けで は な く 「網 羅 的 行 動テス トバ ッテ リー
」2を用いて様々 な種 類の行動 表現型を解 析し,
脳で発現 す る遺 伝子の機 能を 明ら か に して き た。 さ ら に,
遺伝子と行動・
精神疾患の 関 係 を明 らか にする た め に,
行 動 異 常を示し た遺 伝 子 改 変マ ウス の脳につ い て分 了・
細 胞・
回路レペ ル で の多 様 な研究を行っ て き た。 これ まで に数 多くのマ ウス系統の ] 遺伝的に規定さ れ る要因の大きい生物学 的特徴を指 し,
遺 伝 子と表 現 型の問に位 置づ けられる。
統 合 失 調症の 中間表現型として, 作業 記憶やプレパル ス 抑 制などの障 害が知ら れてい る。
2 感 覚・
知 覚,
運 動,
情 動,
睡 眠・
リ ズム,
注 意,
学 習・
記 憶, 社 会 的 行 動などの行 動 を測 定 する た め の 代 表 的な テス ト を組み合わ せ た も の (脳 科 学 事 典 「遺 伝子改 変マ ウス の 行 動テス トバ ッ テ リー
」 (http
:〃bsd.
neuroinf.
jp
!wiki1 遺 伝 子 改 変マ ウス の行 動 テス トバ ッテリー
)参照)。NII-Electronic Library Service
昌 子
・
萩 原・
高 雄・
WALTON・
松 本・
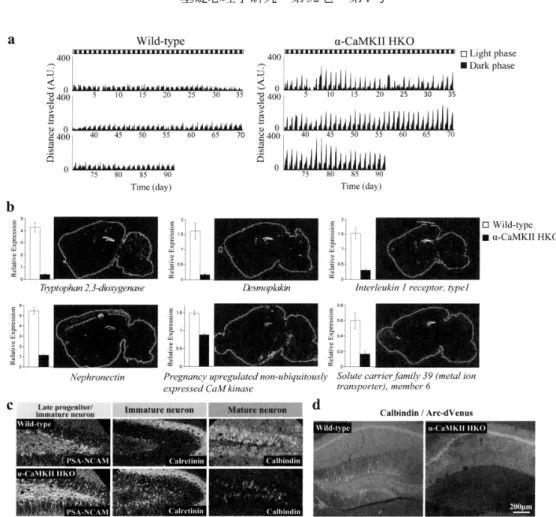
宮 川:精 神 疾 患の中 間 表 現 型として の未 成 熟 歯 状回 103BodyweighI Recml tempera加re
GrLP Streng【h
Anxiety
−
tikebehaviQrLoc。m。tOT activity
SeCial inveraction(訊ovel cage) SOCLal interaction〔home cage) Mαorcoordma に10n Gait Pain s¢nsitivi 【y Prepulse inhlbition Latent inhibitien DepreSSLon
−
hk已behavior Working皿emery Reference nlemory 口Increased 園Decreased ■ n,
s,
Figure 1
.
Behavioral phenotypes in 163 strains of genetically engineered mice.
Each column represents a mutant mousestrainand each row represents a behaVior categery
.
The white and dark grey⊂olors indicate a higher or lower level ofbehav−
ior compared to wild
−
type control mice,
respectively.
Adapted
丘omTakao
et a1.
(2007).
行 動 を網 羅 的に解析し た結果
,
驚くぺ きこ とに,
遺伝 子を改 変したマ ウス系 統のほと ん どで その行 動 表 現 型に異 常が あ ることが わか り, 脳で発 現し てい る遺 伝子の大 半
が
,
行動の レベ ル に おいて も何ら かの機 能 的 役 割を果たして い る こ と が示 唆さ れ た (Figure l) (Takao
,
Yamasaki,
&
Miyakawa
, 2007)。
その中に は,
統 合 失 調 症 や双 極 性 障 害などの精 神 疾 患 患 者に見ら れ る行 動に類 似し た表 現 型 を 示 すマ ウス系 統が多 数 見 出さ れてお り,
その代表的な 例が α一
CaMKII HKO マ ウス で ある。
Ca2+ 1カル モ ジュ リン依 存 性タンパ ク質 リン酸 化 酵 素 II(CaMKII )は,
統合 失調 症の病 態に関与すると考え ら れ てい るグル タ ミン酸 受 容 体の一
種で あるN一
メ チルー
D一
アスパ ラ ギン酸 (NMDA )受 容 体の活 性 化と と もに活 性 上 昇す ることが知 られ てい る主 要な分子 である。
この分 子に は 4つ のサブユ ニ ッ ト (cr、
β,
7,
δ)が あり,
α!β一
CaMKII
は大 脳 皮 質 や 海 馬 を は じめとする中 枢 神 経 系に 多く発 現 し,
長期 増 強や長 期 抑 圧などの神 経 可 塑 性 をは じめ,
学 習・
記憶 機 能な ど に き わ め て重 要な役割を担っ て い ると考 え られて い る (Winder & Sweatt,
2001)。 こ の う ちα一
CaMKII 遺 伝子 をヘ テロで欠 損し た (heterozygous
knockout;HKO )マ ウス は,
同居 し てい る仲 間を殺 し て し まうほ ど激 しい攻 撃 行 動を示 すほか,
新 奇環境であ る オー
ブン フ ィー
ル ド装 置 内で の 自 発 活 動量 が非 常に多 く,
8方 向 放 射 状 迷 路やT
字 型 迷 路 課 題 を 用いた作 業 記 憶 課題の成 績 が 顕著に低下 して い る とい う特 徴を 示 す(Shoji
,
Hagihara,
Takao,
Hattori,
& Miyakawa,
2e12;Yamasakiet al
、
,
2008)。
さ らに,
α一
CaMKII HKO マ ウス は,
統 制 群のマ ウス と比べ ると ホ
ー
ム ケー
ジ内で の活 動 量に 10〜
20 日ほ どの緩徐な周期 性の あ る増減が見ら れ, そ れ は「気 分の波」 を反映する か の ような行動パ タ
ー
ン で あった (Figure 2a)
。
こ の ように,
α一
CaMKII
HKO
マ ウス の行 動の特性は
,
統合 失調症や双極性 障害の患 者に見 られ る症 状と似て いた。
α一
CaMKII HKO マ ウス の行動表現 型は,
どの ような脳 の異 常に基づいて い るの で あろ うか。
気 分の調 節や作 業 記 憶に は海 馬が深く関 与してい るこ と が知 られてい るこ と か ら,
α一
CaMKII HI(0 マ ウス の海 馬に おいて遺 伝子発 現の変 化を網 羅 的に捉えるた め,
DNA マ イクロ ァ レイ に よる解 析 を 行っ た。 α一
CaMKII
HKO マ ウス の海 馬で は,
統 制 群のマ ウス と比べ て,
2,
000個 以 上 もの遺 伝 子 で発 現変化に有意な変化があるこ とが確認さ れ た。 その う ち発 現 量が顕 著に増 加 してい た遺 伝 子の 中に ドー
パ ミ ンDIA 受 容 体 遺 伝子が あっ た た め,
この ドー
パ ミン D1 様受容体に対するリガン ド結 合能を調べ た ところ,
海馬 の中で も歯 状 回と い う部 位で選 択 的で顕著な結 合 能の上 昇が観 察さ れ た。 さ らに,
海 馬で発 現 変 化が見られた上 位30個の遺 伝 子の脳 内の発 現 部 位を調べ たと ころ,
面 白い こ と に6個の遺 伝子 が 海 馬歯 状回で選択 的に発現 す る遺伝 子で あっ た (Figure 2b)(Lein et al.
,
2007 参照)。 これらの結 果は,
α一
CaMKII HKO マ ウス の海 馬,
特に歯 状回に おい て何らかの異常が存在す る可能性を予想さ せ るもの であっ た。 歯状回 は,
成 体 神 経 新 生 (大 人に なっ た後で も神 経 細 胞が新し く生ま れ る現 象 )が生じ る領 域と し て知 られ.
脳の中で も過 去十 数年にわ たっ て 精 力的に研究が行わ れて き た場所で あ る。 歯状 回には 神 経幹細 胞 が 存 在 し,
神 経 幹 細 胞は神 経 前 駆 細 胞を経て未 成 熟な神 経 細 胞か ら 成 熟 した神 経 細 胞へ と分 化する。
新 生し た神 経 細 胞は数 週 間か ら数ヶ月の期 間を経て成 熟し,
こ の間に海 馬 内の 既 存の神 経 回路に組み込ま れ,
生理的な機 能を持っ よ う になる と考え ら れて い る。
BrdU染色コ
を行っ たと ころ,
3BrdU は細胞が増 殖す る際に起 こるDNA の合 成 期に チ ミジン の代わ りに細胞に取 り込 ま れ る物質で,
BrdU を取 り込ん で標 識 され た細 胞は増 殖 中の細 胞 であっ た こ とを示し細 胞が新 たに生 まれ たこ とを 示 唆す る。
N工 工一
Eleotronio Library104 基 礎 心 理 学 研 究 第32巻 第1号 a
=
≡・
着 三K”
;三
ヨ一
主 b 丶Vild−
type属
O灰
牌
箪
h
亀
国
ひ
’
亠 … 4ー
(
,
⊇.
< モ 三 翠 鐚 r.
oL − − U40DI 40 45 50 ∬ 60 65 ;O lj
I
醤 。坦 鼎 丁imじ〔day} 辜11 CTOFtophan2
、
3−itJD・
gc,
ntis’
t,
‘
Vep’1厂ont.
ct’fT厨
1 巨=
窮己
」
凝
亠
り
「
二
並u
留
■
一
δ一
芻り
乙 4』
り
.
’
「
冒
τ匡
α一
CaMKH HKO 40り □Ligh仁phasc…
。血血
血
血血
山副
’D”’kphase B400 5 10 1〜 21〕 25 ヨ0 ヨ5
1
。血
血
川
川 川
1
血
鍔血 齟
、
冖 15 $o 尺.
i 9冂 τiIne 【daン).
: .
−自
「
下
一
鬲
工
∋
國
臨
団
卍
ひ
’
一
】
”
[
翠
Z口
O一
晒
樽
り
b
ら
頁
噬
2
.
’
轟
署
[
2
〆nte〃げ‘面η1’ぞc{ヴノ’砿 4L・
pelP”egnt〃 κ:ドUi)r[
’
gu’Cl’t’
dnon−
itbiijuifou.
sl.
レ ∫〔’ノutL,
【’
ttrrittr.
〆α〃r”し’
39bnc’
tat ionEllコrc
,
.
∬ [t
ごκ‘
J.
,
Wkina.
sc’
f厂
an・
yp‘
,
rtet ソ.
niemh匚
’
r 6 rtd
囗W
.
’
ild−
tyPL・
■ a
・
CaMKIIHKOFigure 2
.
Home−
cage activity and iDGphenetype
in a−
CaMKII HKO mice.
(a)Temporalpattem
of locomotor activityof wild
−
type and α一
CaMKII HKO mice ill their home cage.
(b)Relative expression levels of the downregulated genesselectively expressed in the DG inα
一
CaMKII HKO mice (datafrom
Allen Brain Atlas,
http:〃mouse.
brail1−
maP.
org),
(c)Incr
−
CaMKII HKO mice,
expressions of calretillin amd PSA−
NCAM (immature−
neuron markers )markedly increased,
andexpression ofcalbindin (a mature neuron mark εr)decreased
.
(d)Exprεssion ofArc−
dVenus in the DG ef cr−
CaMKII HKOmice after the working nlemory task was complete1
γ abolished
.
Adapted 丘01n Yamasaki et al.
(2008)and Matsuo et al.
(2009)
.
α一
CaMKII
HKO マ ウス の歯 状回で は,
野 生型マ ウス に比 べ て,
こ の成 体 神 経新 生が 増 加 して い るこ とが わか っ た。
さらに,
α一
CaMKII HKO マ ウス の 歯状 回の神 経 細 胞の分 化の程 度 を調べ たとこ ろ,
成 熟 神 経 細 胞の分’
r
一
レ ベ ル で のマー
カー
(指 標 )で あ るカル ビン ジン の.
発現は低 トし (Fi8ure 2c), 未 成 熟.
神 経 細 胞の分子マー
カー
で あ る カル レチニ ン お よ びPSA−
NCAM の発 現が増 加して い た。
つ まり,
こ のマ ウス の歯 状 回の神 経 細 胞は,
分 子レベル で 未成 熟 な細 胞の特 徴 を 持っ てい た。 次にα一
CaMKII HKO マ ウスの歯 状回の神 経 細 胞をゴル ジ染 色 法に より形 態レ ベル で解析し た結果.
樹状突起の分枝が少なく,
全 長が 短い こ と が確認さ れ,
形 態 学 的に末 成 熟で ある こ と が わ か っ た。
続いて電 気 生 理 学 的レ ベ ルで解 析し た とこ ろ.
このマ ウ.
スの粛 状回 の神 経 細 胞は,
入 力 抵 抗が高く.
脱 分 極 時の スパ イ ク 発火 頻 度が少ない な ど,
通常の未 成 熟 な神経細胞が示す特徴を有してい るこ と が わか っ た. ま た,
こ のマ ウスで は,
歯 状 回か らCA3 領 域へ の神 経連 絡である苔 状 線 維 CA3 シ ナ プスで は,
シ ナプス促 通と 呼 ば れ る 可 塑 性 が 顕 著に低 ドして お り,
こ の点で も 幼 若 期の マウスの もの と同 様な特 徴が見 出さ れ た。 こ の よ う に,
α一
CaMKII HKO マ ウス の歯 状回で は神 経 細 胞が過 剰 に産 生さ れ ると ともに,
神 経 細 胞のほとんどが 分子,
形 態, 電 気 生理学 的レベ ル で未 成熟な特 徴を 示 していた。 α一
CaMKII HKO マ ウスの 歯状回の神 経 細 胞で は,
どの よ う な活 動が 生 じてい る の であろうか。 これを調べ るた め,
活 動が 生 じ た 神 経 細 胞が 蛍 光 を 発 するよ う なα一
CaMKII HKO マ ウス を作 製し た.
この マ ウスは,
神経 活 動依 存 的に発 現す る最 初 期 遺 伝 子Arcの プロモー
ター
制 御 下で分 解 促 進 型 蛍 光 蛋 白 質dVenusを 発 現 す る ト ラン ス ジェ ニ ッ クマ ウス(Eguchi& Yamaguchi
,
2009)とα
一
CaMKII HKO マ ウス とを交 配す るこ と に よっ て 得 ら れ た。
こ の交配に よ り生ま れ たマ ウス に作 業 記憶 課題 を経 験させ たと ころ,
統 制 群のマ ウス で は,
作 業 記 憶 課 題 経 験 後に歯 状 [bl]の神 経 細 胞の 多 くで dVcnusの発 現が観 察NII-Electronic Library Service 昌子
・
萩 原・
高 雄・
WALTON・
松 本・
宮川:精 神 疾 患の 中間表 現 型と して の未 成 熟 歯 状回 105 さ れ,
神 経 活動が増加して い る こ とが示さ れ た。一
方,
α一
CaMKII HKO マ ウス の 歯状回の神 経 細 胞で は, 経 験 依 存 的なdVenus
の発現が ほぼ完全に消失し (Figure 2d ),
神 経 細 胞が正常に機 能してい ない可 能 性が高い こ と が示 唆 された (Matsuo et aL,
2009)。
こ のようにα一
CaMKII を 欠 損 し たマ ウス は,
統 合 失 調 症や双極性 障害な どの精神疾 患様の行動異常の パ ター
ン を 示 し,
かつ歯 状 回で は神 経 細 胞のほとん ど が未 成 熟 な 状 態の ま まで留まっ て い るこ と が示唆さ れ た。
我々 は,
こ の ように歯状回の神 経 細胞が未 成 熟な状 態の ま ま留 まっ て い る状態を 「未成熟歯 状回 」 と命名し,
統 合失調 症や 双極 性 障 害の中 間 表 現 型の 1つ で は ないか と考 え た。3.
未 成 熟 歯 状 回 を もつ複 数の遺 伝 子 改 変マ ウス の同 定 未成熟 歯 状回とい う現象は,
ec−
CaMKII HKO マ ウス に だ け認められ る特 殊 な現 象 なの で あろうか,
それとも他 の マ ウス 系 統で も観 察され 得 る現 象 なのであろうか。
我々 は,
これ まで に得ら れ た多数の遺 伝子改 変マ ウスの 網 羅 的 行 動 解 析 デー
タ4 (Takao et al.
,
2007)か ら,
α一
CaMKII HKO マ ウス と同 様の行 動 表 現 型を示し たマ ウス 系 統を探 索 した結 果, Schnurri−
2 と呼 ばれ る転 写 因 子を 欠 損し たマ ウス (Shn−
2 KO マ ウス)が α・
CaMKII HKO マ ウス と同 様の行 動パ ター
ン を示 すこ とを確 認 し た。 Shn−
2 KO マ ウス は, 自 発 活 動 量の顕 著 な 増 加 や 作 業 記 憶の障害を示す ほ か (Figure 3),
社 会 的 行 動やプレパ ル ス 抑 制 の 低 下 な どの様 々 な行 動 異 常を示 して い た (Takao et al、
,
2013)。 そこ で,
Shn−
2 KO マ ウス の海 馬で の 遺 伝子発 現レ ベ ル をDNA マ イ クロアレ イに よっ て解 析 し,
α一
CaMKII HKO マ ウス と の比較を行っ た結 果, Shn−
2KO マ ウス の海馬で の遺 伝子群は
,
α一
CaMKII HKO マウス と顕 著に類 似し た発 現パ タ
ー
ンを示 し て い た。 驚 く べ き こ と に,
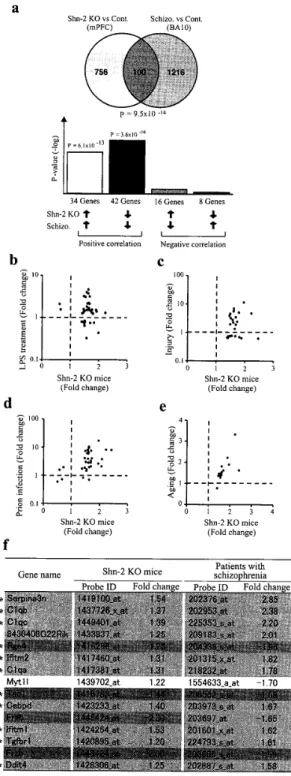
両 系 統で共 通し て発 現 量が有 意に変 化し て い る遺 伝 子は 100個 以上 も あ り,
しか も,
そ れ らの発 現 変 化 量の程 度 までもが両 系 統 間で酷 似 してい るこ とがわ か っ た (Figure 4)。
さらに,
Shn−
2 KQ マ ウス の歯 状回で は,
α一
CaMKII HKO マ ウス と同様に, 成 熟 神 経 細 胞の マー
カー
で ある カル ビンジン の発現量が顕著に低 下し て いた ほか,
Shn−
2 KO マ ウスの歯 状回の神 経細 胞は,
発 火 閾 値や発 火 潜 時が小さく,
脱 分 極 時のス パ イク発 火 頻 4 系 統 情 報 や 行 動 デー
タ はすべ て 「Mouse Phenotアpe Database」 に登 録 さ れ,
学 術 論 文と し て公 表 し た デー
タ につ い て は ウェ ブサ イ ト(http
:〃www.
mouse−
phenQtype.
orgf)にて公 開 して いる。 5 7 6 5 4り
う
0【
β=
00σ
軅
曽 暇 』 aり
石 』 U 巨ロ
置 髭 選一
⇔EighI
−
arm ra〔1ial maze testb 25 @0@ WiLd
一
卯e(
n=匸 j〔n= )
ユ
o@
詈鵠
i5@
籌
B 羅 3 @ 9 12BlockS 2trialsT −
ze test 〔force 己al重ernst
n
task
)
c
OWLId’1y
(
)n16
dl 戯 @ 4 6 8 s¢55jons 69
12Btoc of2trials
T−maze
test
〔
Lefl −righ重
discrination
重
a)Reveale
〔 u16 ) D〔nij246 呂10
Zl4 Se3s 匹onsFigure 3. Wbrking memory de 且cits
i
@Shn
−2 KO mice. (a)Different arm choice in first 8tries
and(b)re− visiting error in the spatial workimemory task
of
the8
−arm radial maze , (¢ )Corre
responses (%)in
the
T−maze forced−
alternation
ta.(d)Correct re− sponses(%)in the T−maze left −rig
discrimination task . Asterisks indicate stati$tica
significances
(p< 0.05),Adapted frQm]]
ao
et
al
, (2013
), 度 が少な いな ど 未 成 熟神経 細 胞に 認め
られ
る特徴 があ ることも 見出 され た。αCaMKII
HKO
マ ウ スの 場合
と 同 様に,精
神疾患様 行 動異 常パ ターンを示 すShn −2
KO
マウスの 馬歯状回では, ほとんどの 神 経細 胞が 成熟 な状態
に あ る こ とが明 ら か になったe そ 他 にも,精神疾患 様 の 行 動異 常を 示し,か つ未成 熟 状 回 を持
つ遺伝子改変
マ ウ ス系統が 次々に見つかっ て いる。
ナプトソー
ム 関 連タンパク 質25kDa
(SNAP −25 )
, シナプス 膜に 存在す るt
−SNARE
タ ン パク質 で シナ プス前 部からの神経伝達物質の細胞
外
放出に関与 し , 統合失調症(LeWis ・et・a
D
,2003
;Young
et
al.,1998 )や
注意欠陥多動性障害( HD)
(
Barr et aL ,2000;Ch。i etal ..2007 ),てん
かん(Kataeka e@al.,2006 ;Zhang
,
Vilaythong ,Yosho
C
&Noebels
,2004
) な どの 精神・
神経疾 患との 関 が示 唆 され てい る 分子 で ある。こ のSNAP
− 2ェ
リン酸 化 されない よ うに1
アミ ノ酸置換が行 われ た ノッ ク イン
( SP
−25
Knock
−in
(KI ) )マウ
スに お い て ,活 動 量 の 亢 進や作 業記 憶 課題 の成績低 下 が 認 られ,かつ歯状回 の神経
細
胞が未成熟な
態にあ るこ とが
確認 された (Ohiraet
al .,
2013
) カルシニューリン(calcineurin
,C
jは ,脳
の 細胞 内 シ グナル伝達 に 関与 す るタンパク 質 脱リン 酸lO6 基 礎心 理学研究 第32巻 第1号 5
4
3
へ
乙
1
0
(
ω
切戸
雨 ‘ り層
[
G 』〕
OU一
∈ O 冨 工 = 呂 Σ呵
∪ 16 a ;1・
O.
S]O{P・
:O.
−O曲1
.
3
旨1
.
じ も,
臨 嶺.
i・・
’
臣・
・
yl
” 8・
0 1 2 3 4 5 6 Shn・
2 KO mi匸
ビ
〔Foldじ
h齟
呂¢
, b へ,
.
,
1。
N 卜[
1】
I LO⊂.
547362 Spera13 NtG]/
19]OSO] 巳亅Dじ
5dFhm7 〔
.
cndl Wnk4 AI ]OOVOK工MRik c 丁dn2 D;
P NpnL 〔1alb1
匚apn.
1146 〔伽04]−
atM腸
nvks]ヨ PcdhlI [rpan19 R>.
rI D幽
3 』 5 0 1白
L
4,
bKO n O.
5 1 11 n.
5 [丶ttr111aLLxじd CNpre∬tttlL Norm
己
II∠
edじ
丶
prじ
sskpnHgure 4
.
Maturation abnormalities in DG neurons ef Shn−
2 KO mice.
(a)The hippocampal transcriptome pattern of Shn−
2KO mice is similar to that of cr
−
CaMKII HKO miGe,
which also demonstrated maturation abnormalities in the DG,
Genesshowing
differential
expressionbetween
genot}rpes atp
く0.
005 inboth
experimcnts were plotted.
(b
)Normalized gene ex−
pression of
djfferentjally
expressed genes in Shn−
2 KO and α一
CaMKII HKO mice,
The top lO genes are ind正cated in thegraphs
.
(c)Expression of the mature ]コeuronal marker calbindin wasdecreased
,
and the expression of the imlnature neuro−
na1 marker calretinin “was markedly increased in the DG of Shn
−
2 KO mice,
Adaptedfrom
Takao et al,
(2013).
つ で あ る。 こ の CN を前脳特異的に欠損さ せ たマ ウス (CN KO マ ウス)は
,
作 業記憶の 低 ドを は じ めとす る 様々 な統 合 失 調 症 様の行 動 異 常を示す (Miyakawa et al.
、
2003;Zeng et aL,
2001)。
このCN KO マ ウスで も,
歯状ltil
神 経 細 胞が未成熟な状 態で あ るこ とが 示唆さ れて い る (Hagihara et aL,
2011a)。 これ まで の研究か ら,
歯状回にお け る数種の遺伝 子の 発 現 量を リアル タ イ ムPCR 法に よ り測定しその発 現ハ ター
ンを 調べ る こ と に よっ て,
歯状回が未 成 熟 歯 状回 で あ る か どう か を評 価す る簡便な方法が確 立さ れてい る(Figure 5) (Hagihara
.
Toyama,
Yamasaki,
& Miyakawa,
2009)
。
こ の方 法で は,
歯 状「・1
に お け る ドー
パ ミン受 容 体DIA (Drdla )遺 伝子の発 現上界と1・
リブ トファ ン2,
3一
ジ オ キシゲ ナー
ゼ (Tdo2 )お よ び デス モプ ラ キン (Dsp) 遺 伝 亅孔の 発 現低 トを未 成 熟 歯状回 のマー
カー
と して 用い る (Yamasaki et al.
,
2008;Ha帥 ara et aL,
2009)。
現 在 まで に これ らの遺 伝 子マー
カー
を利 用 した解 析によ り,
自発 活 動 量の亢 進 や 作 業 記 憶の障 害な どを示した マウ ス系 統 の う ちの お よ そ半 数の系統で歯状回が未成熟で あ るこ と が示唆さ れ て お り,
未成熟 歯状回が精神疾患 様行動異常 を 示 すマ ウス の中で広 く認め ら れる表 現型で ある こ と が 3 21 宅 }
u
。
三 峰 石 匡矼
、
CaMKI [HKOmice
o
ノ
ゴ
Of“♂
85
4
2 q っ
尸
尋」
穿 劣.
蕚 三 3一
0 卜11uoxctiT
匸C−
t1.
ca 【cd mic じ HK(,ズ
ボ
♂♂
’
2 ヨ ユら L α
日
窪 曁 ロ 話り
.
三吊
[
齷 Shn・
2KOmiu じ 0
ノ
ボ
碑 虐」
4 3 2 1[
羣 誇」
【
門
蓊」
三 薯 匡 0P[k,ca Φine
−
1「じaLed mlee* *
ど
詞
・s避
Figure 5
.
Identification of the immature dentate gyrus(iDG)phenot}pe using real
−
time PCR,
iDG is charac−
terized by upregulation of dopamine receptor Dla
(Drdla)and downregulation of both desmoplakin
(Dsp )and tryptc)phan 2
,
3−
di(>xygenase (TdQ2)in thehippocampu5
,
Asterisks indicate statistical significanc.
es (* *
ρく0
.
Ol).
Adapted from Yamag.
aki et a],
(2008),
Takao et al
,
(2013),
Kobayashi e【a1、
(2010),
and ShinNII-Electronic Library Service 昌子
・
萩 原・
高雄・
WALTON・
松 本・
宮 川:精 神 疾 患の中 間 表 現 型と して の未成熟歯 状回 107 わ か りつ つ ある。
4. 薬物投与
に よ る歯状
回神 経 細 胞の 「脱 成 熟」 未 成 熟 歯 状 回とい う現象は,
遺伝子改変マ ウス の解析 か ら初め て見 出さ れ たもの であるが,
類 似 した現象が野 生 型マ ウスに薬 物を投与す る こ と に よっ て誘 導 可 能であ るこ ともわ かっ て きた。
K。bayashi et al.
(2010)は,
抗 う つ薬と して最も広く使 用さ れ てい る選 択 的セ ロ トニ ン再 取 込み阻害薬 (SSRI)の 1っで あ るフル オ キ セ チン を成 体マ ウス に慢 性 投 与 する と,
歯 状 回の神 経 細 胞が擬似 的 に未 成熟な状 態に戻っ て し ま うこ とを発 見 した。 フルオ キセ チンを慢性 投 与し たマ ウスの歯 状回で は,
成 熟 神 経 細 胞のマー
カー
である カ ル ビンジン の 発 現が減少し, 未 成 熟 神 経 細 胞の マー
カー
で ある カル レチニ ンの発現が増 加してい た (Kobayashi etal.
,
2010; Ohira & Miyakawa,
201 1)。ま た
,
未 成熟 歯 状回の 遺 伝子マー
カー
で あるDrdlamRNA 発 現は増 加 し
,
Tdo2やDspの mRNA 発 現は低 下 してい た。 電気生理学 的に も歯状回の神 経 細 胞は,
フルオ キセ チンの慢性投与によっ て未成熟な神 経 細 胞の特 徴
で あ る高い興 奮 性を示し, 苔 状 線 維の シナ プス促通 が幼
若な神経細 胞に見ら れ るレベ ル まで低 下 する ことが確 認
され た。 こ の よ う な分子的
・
電 気 生 理 学 的 特 徴は,
α一
CaMKII
HKO
マ ウス,
Shn−
2 KO マ ウスお よ びSNAP−
25 KI マ ウス の場 合と酷 似 してい た (Table 1) 。 こ の よ う に,
成熟 神 経 細胞が擬 似 的な未 成 熟 状 態へ と逆 戻 りする現象 を 「脱 成熟 (dematuration)」 と呼ん でい る (Kobayashi
et al.
,
2010)。
なお,
他のSSRIで ある パ ロ キセ チンの投 与に よっ て も歯状回神経 細 胞が脱 成 熟 する こ とが確 認さ れて い る。
パ ロキセ チンを慢 「生投与す る と,
フル オ キセ チン の場合と同 様に, 海 馬で の カ ル ビ ンジンお よ びDspの 遺 伝 子発現は低 下し (Smaber
et aL,
2008),
苔 状線 維一
CA3 間の シ ナプス促通は抑 制さ れ る (Kobayashi et al.
,
2010)。 これ らの知 見は,
セ ロ トニ ン神経 系が歯 状回神 経 細 胞の 成 熟 変 化に関 与 して い る こ とを示唆す る が, そ の詳 細な メ カニ ズ ム につ いて は未だ不 明である。 な お,
Kobayashi らの研究で 「脱 成熟」現 象が誘 導 されたと きの投薬量は,
野 生 型マ ウス (Dulawa
,
Holick,
Gundersen,
& Hen,
2004)や うつ 病モ デル マ ウス (Alenso et al
.
,
2003; DaVid et al.
,
20eg)に おい て抗うつ効 果が確 認さ れた際の投 薬 量より も多か っ た。 比 較 的 高 用量の フル オ キセ チンの投 与に よっ て活 動 量が 大 き く変 動 する よ う に な る (Kebayashi,
Ikeda,
& Suzuki,
2011)こと か ら,
フ ルオ キセチンの慢 性 投 与に よっ て誘導さ れ た 「脱 成 熟」現 象とその行 動の変 化は,
臨 床で観察さ れて い る抗うつ 薬 誘 発 性の躁 状 態 を 反映し た現 象で ある可 能 性が考え られ る。 歯状回で の 「脱 成熟」現象は, けいれん発 作に よっ て 誘 導さ れ るこ とも見出さ れ てい る。 て ん か ん は,
脳の神 経 細 胞が反 復 的かつ過 剰に活 動し た結 果,
けいれ ん や意 識障 害などの症 状が現れ る疾 患で あるが,
て ん かん患 者 の中には幻覚や気 分の不 安 定 化,
躁 状 態などの精 神 症 状 を 呈 する場 合があるこ とが知 られて い る。 近年,
遺 伝学 的 研 究に より,
染 色 体15q13.
3の微 小 欠 失がて んか ん や 精 神 遅 滞,
統合失調症の発症 リスク を高め ることが明ら か にされ (Sharp
et al.
,
2008),
て ん か ん と統 合 失調 症に は共 通の遺 伝 的 基 盤が あると推 察されて いる。 非 選択的 ム ス カ リン受容体作動 薬で あ るピロカル ピン は,
投 与す ると てん か ん様の けいれ ん発 作を誘 発す るこ と か らて ん か ん モデル動 物を作 製するた めに用い られてい る薬 物で あ る。
この ピロ カル ピン を投 与 した ラッ トの海馬で は カ ル ビ ンジン の遺伝子発 現およびタンパ ク発 現が低 下 す るとい う報 告がある (Carter
,
Harrisen,
Falenski,
Blair,
&De−
Lorenzo
,
2008)。
我 々は これ らの知 見 を もと に,
ピロ カ ル ピン誘発 性 けいれ ん発 作を 示 すマ ウス の 歯状回の神 経 細 胞が未 成 熟 な状 態にあると の仮説を立て詳 細に検 討し た。
こ の結 果,
ピロ カル ピン誘 発性の て ん か ん発 作を示 したマ ウス の 歯状回神経細 胞で は, 遺 伝 子・
タ ン パ ク発 現レベ ル か ら 電気生理学 的レ ベ ル に至るまで α一
CaMKII HKO マ ウ ス と類 似し た 未 成 熟 歯 状 回様の特 微を持つ こ とが明らかに なっ た (Shin et al.
,
2013)。 ま た,
ピロ カル ピン投 与マ ウス は、
自発 活 動 量の亢 進 や作 業記憶の障害 を 示 すこ とが確認さ れ,
行動レ ベ ル の異 常につ い て も未 成 熟 歯 状 回 を 持つ マ ウス と似て い るこ と が わか っ た (Shin et aL,
2Dl3)。
けいれ ん発作を示し た ピロカル ピン投 与マ ウス の歯 状 回神 経 細 胞が未成 熟な状態に ある と い う知 見は,
未 成 熟 歯 状回が けい れ ん発 作に伴 う神 経 細 胞の過剰な興 奮に よっ て誘 導さ れ た可 能 性が あるこ とを示して い る。 ピロ カル ピン の投 与 に よっ て けいれん発 作を示 し たマ ウス に おい て,
脱 成熟が生じてい た条 件を調べ た と ころ,
けい れん発 作が 1回 生じ た だ け で は十 分で は なく,
そ の後に 自発 的 再 発 性発 作が起こ る必 要があるこ とが わ か っ た (Shin et al.
,
2013)。 ま た,
SNAP−
25 KIマ ウス は, 生 後21〜
24日以 降に 自 発 性けい れ ん発 作を起 こすこと が報 告さ れてい た が (Kataoka et al.
,
2011),
こ の発 作は生後16日 か ら抗けい れん薬で あ るバ ル プロ酸 を 投 与 する こ と に よっ て抑 制す る こ とが 可能で あ る (Otsuka et aL,
2011)。
SNAP−
25 KIマ ウス に生 後16日 目か らバ ル プロ酸を投 与 し,
自発 性けいれん発 作を抑 制 した と ころ,
成長 後の歯 状 回の神経 細胞で はカル ビンジ ン と,
カル レチニ ン の発 現が 正常化 すると とも に,
作 業 記 憶の障 害が改 善さ れる N工 工一
Eleotronio Library108 基礎心理学研究 第 32 巻 第1号
Table 1
,
Behavioral
,
electrophysiological and rnolecular phenotypes in mice with iDGPhenetype a
−C
”MKIIShn
−
2 KO SNAP.
25.KI CNKO FI・・x・ti・ ・ Pi1・…pi
・・HKO t「eatment t「eatment
BehaVior LOCOmetOr aCtiVity
W ・rking mem ・ry Social interaction PPI ↑ ↓
Mutants
kill
cagemates ↑ 1 Ψ ー Ψ ↓ 夲 IIΨ
↓ ↓ ↑ ↓ ↓ ↑↓*L * 个 1 ↓ 1 ΨEle¢trophysiology ルfo∬T/ibre
−
CA3synapseBasal transmission
Frequen匸y facilitation Granule匚ell soma
Passive Input resistance
Active Excitability Spike number 小 1 ↓ ↑ ↑ ↓ ↑ ↓ ↑ 个 II Ψ ↓ ↑ ↑ ↓ ↑ ↑ ↑ ↑ Molecule
Markers
qブimmature granule‘跳Dopamine Dl receptor (Drdla)
Calretinin
Markers ・」伽 ure granule cells
Tryptophan 2
,
3−
dioxygenase(Tdo2)
Desmoplaldn (Dsp)
Calbindin GluRlImmediate
ear!
y
gene induct加 c−
Fos or Arc−
dVenusInflammationイelated mole ‘ules
Glial丘brillary acidic protein
(
GFAP
) Complement components MHC genes Adult neurogenesis BrdU incorporation ↑ ↓ ↓ 1 堂 1 Ψ ↓ ↑ ↑ ↓ 1 Ψ ↓ 1 Ψ ↓ ↑ ↑ 个1 ↑ ↑ ↓ ↓ 1 Ψ ↓ ↑ ↑ ↑ ↓ 2 * ↑ 个 1 ↓ ↓ 1 Ψ ↓ ↑*z ↑*2 ↑*1 ↑ ↑ ↑ ↓ ↓ ↓ ↑*4 ↑*5 ↑*5 ↑*4・
6 * K・b
・y・・hi・t・1,
(2011)・ *2,
Si
旺・ber
et・1.
(2008)・ *3,
・D・t・u・et・L (2005)、 * H・gih
… et・L(2005)、 *5,
・Ok、m 。t。 ,t、L (2010)、 *6・
Ch・ ・t・1・
(2004).
↑,
lncrea・ed ・・up ・eg・1
・t・d
・↓,
d… ea・ed ・ ・d・w・ ・eg・1・t・d
、一
,
… ig・i丘・ant・differen
。e,
こ とが わか っ た (Ohira et al
.
,
2013)。 これ らの知 見は,
歯 状回神 経 細 胞の脱 成 熟を誘 導 する要 因の 1つ と し て,
神経 細胞の断続 的 な過 剰 興 奮が あ り.
そ の結 果と して精 神 疾 患 様の行 動 異 常が生じ ること を 示唆して い る。
未 成 熟 歯 状回 を持つ マ ウス の 歯状 回の神 経 細 胞に は興 奮 しやすい 性 質が あ る。 例え ば,
α一
CaMKII HKO,
Shn−
2 KO,
フル オ キセチン慢 性 投 与マ ウスの歯 状 回の神経細胞 で は,
活 動 電 位 を 誘 発 する た め に必要な閾値電 流が低 く,
神 経が興 奮 しや すい。 ま た,
Shn−
2 KO マ ウス の海 馬で は,
GABA 抑制 性 神 経 細 胞に発 現 するパ ル バ ル ブミ ン (parvalbumin;PV )やGAD67 の発現が減 少して い る が (Takae et al.
,
2013),
こ の知 見は抑 制 性 神 経 系の機 能 低 下に よっ て神経細 胞が興 奮 しや す くなっ てい るこ とを 示唆し てい る。
ま た最近,
精 神 疾 患の脆 弱 性 遺 伝 子の有 力な候 補の一
っで あると考 え られ てい るDiscl遺伝子を 歯 状 回の新 生 神 経 細 胞のみで ノ ックダ ウンして そ の機 能 を抑 制す る と,
神経の興奮 性が.
トが り情 動・
認 知 機 能が 障 害さ れ ること が報告さ れ た (Zhou et al.
,
2013)。
SNAP−
25K 匸マ ウス に おい てバ ル プロ酸の投 与で各種異常が 正 常 化 される こ と (Ohira et aL,
2013)も合わせ て考 慮 する と,
これ らの知 見は,
歯状回神 経 細 胞の過 剰 興 奮に よっ て神経 細胞が未 成 熟 な状 態にな り,
精神疾患 様の行 動 異 常が生 じてい る可 能 性が あ るこ とを示して い る。
5.
未 成 熟 歯 状 回と精神疾患
精 神 疾 患 様 行 動 異 常を示すマ ウス の歯 状回の神 経 細 胞NII-Electronic Library Service 昌子
・
萩原・
高雄・
WALTON・
松本・
宮川;精神 疾 患の中 間 表 現 型としての未 成 熟 歯 状 回 lQ9 が未 成熟な状 態に あ る とい う知 見か ら, 精 神 疾 患 患 者の 歯 状回で も同 様の現象が起きて い る こ とが予 想さ れ た。
Yamasaki et al.
(2008)は , 精 神 疾 患 (統 合 失 調 症,
統 合 失 調 感 情 障害,
双極性 障害)患者 21例を含む166例の死 後 脳か ら得 ら れた海 馬の遺 伝 子 発 現 デー
タ を用い て,
α一
CaMKII HKO マ ウス の海 馬で変 動が見 られ た10個の遺 伝 子 (ADCY8
,
CCNDI,
LOCI51835・
LOC2S4018,
NTNGI,
PDYN ,
PIP3−
E,
PNCK,
SPATA13.
TDO2 ) を対 象に ク ラスタ