奈良教育大学学術リポジトリNEAR
Degradation of Mitochondrial Genome of an
Unstable Strain in Saccharomyces cerevisiae as Revealed by Loss and Retention of the
Drug‑Resistance Factors (With 1 Text‑figure)
著者 SUDA Kohta, SUDA Kayoko, UCHIDA Akira journal or
publication title
奈良教育大学紀要. 自然科学
volume 27
number 2
page range 71‑83
year 1978‑11‑25
URL http://hdl.handle.net/10105/2488
Degradation of Mitochondrial Genome of an Unstable Strain in Saccharomyces cerevisiae as Revealed by Loss and Retention of the Drug-Resistance Factors
(With 1 Text-figure)
Kohta Suda
{Biological Laboratory, Nara University of Education, Nara 630) Kayoko Suda
(Depatment of Biology, Faculty of Science, Nara Women's University, Nara 630) and
Akira Uchida
{Biology Division, College of General Education, Kobe University, Kobe 657) (Received May 1, 1978)
Sammary
The instability, namely, high mutability to petite mutants, of an unstable strain in S.
cerevisiae was revealed to be under the control of a recessive nuclear gene. By using the mitochondrial genetic markers such as the risistance factors to oligomycin, erythromycin and chloramphenicol, the investigations were conducted on the genetic constitutions of the mitochondrial markers and the degrees of suppressiveness of the petites spontaneously ob- tained from the unstable and stable strains. The results suggest, on a mechanism of the instability, that a successive degradation of the mitochondrial genome of the unstable strain occurred during vegetative growth rather than that the overall mitochondrial genome was diluted out from the cell.
Introduction
The biogenesis of mitochondria of the yeast, Saccharomyces cerevisiae, is affected by both nuclear genes (pet) and the cytoplasmic factor (rho). Grande cells (pet+, rho+) can con- sequently utilize for growth both fermentable and nonfermentable carbon sources, but petite mutants, either (pet-) or (rho-), can only utilize fermentable carbon source.
Garande cells produce cytoplasmic petite mutants during vegetative growth at such rates that the cell population usually contains a few per cent of petites. There are some yeast strains, called unstable strains, which produce cytoplasmic petite mutants as high as 50%
or more. Ephrussi and Hottinguer (1951) first reported that the instability, namely, high mutability of grandes to petites, of an unstable strain was under the control of a recessive
71
72 Kohta Suda, Kayoko Suda, Akira Uchida
nuclear gene. In addition, it is also known that some nuclear pet mutations simultaneously result in rho- mutation, leading the progeny to double petite mutants {pet-, rho-) (cf. Beck etal.1971).
Since DNA was found in yeast mitochondrion (Schatz et al. 1964), it has been assumed that the mitochondrial DNA is the molecular basis of the rho factor or the mitochondrial genome. Many drug-resistance factors which cytoplasmically inherited have been roported (Thomas and Wilkie 1968 ; Linnane et al. 1972 ; Wolf et al. 1973 ; Michaelis 1976), and some of them are considered to be arranged in the mitochondrial genome (Uchida and Suda 1973 ; Deutsch et al. 1974 ; Molloy et al. 1975 ; Schweyen et al. 1976). Recently, gene- tic mapping of the mitochondrial genome or mitochondrial DNA has been, with genetic and biochemical methods, established by using many markers such as drug-resistance
factors, cytochrome-deficient (Slonimski and Tzagoroff 1976) and temperature sensitive re- spiratory-deficient mutants (Bolotin-Fukuhara et al. 1977). However, genetic investigations
on the mitochondrial genome for the unstable strain have not been reported.
Here described are the analyses of the mitochondrial genome of spontaneously arising petites from an unstable strain and those from a stable strain, by using the three cyto- plasmic drug-resistance factors as the the genetic markers of the mitochondrial genome.
Materials and Methods
Strains. The strains used are summarized in Table 1. [OLGr], [ERYr] and [CHLr] denote the cytoplasmic resistance factors to oligomycin, erythromycin and chloramphenicol, which reside in [07], [RHI] and [RI] loci defined by Slonimski's group (Avner et al. 1973), res- pectively. For the allelism analyses were used the strains, IL836-2A [OR1(OI)] and IL8-8D [C*321(RI), ER514(RIII)l
Media. YPD (1% yeast extract, 1% peptone and \% glucose) was used as a fermentable growth medium. YPG (\% yeast extract, 1% peptone and 2% glycerol) was a nonfermen-
table growth medium. SCD was a modified Wickerham's minimal medium (Wickerham
Table 1. Strains.
Strai n Genotype Source
C3516-1 A C 814-40 D OEC 1222 Sll-17R S51-41 R S 55-13 R IL 836-2 A IL8-8 D
a, ade 1 T. Takahashi
a, ura3 "
a, ade 1, his4, thr4, [OLGr, ERY", CHLr] Suda and Uchida, 1974
a, ura3 "
a, ura 3, [OLC, ERYr, CHL'~\ Segregant of OEC 1222
x Sll-17R
a, ade 1, [OLGr, ERY7, CHIS] Segregant of C 3516-1 A
x S51-41R
a, ura 1, [OR1(OI)] P. P. Slonimski
a, ura 1, [CB321(RI\ ER514(RIIIj\ "
1946) supplemented with 1% glucose and 0.2% casamino acids. SCG was as SCD with 2%
glycerol subtituted for glucose. For plating, 2% agar for glucose media and 1% for glycerol were added. All the experiments were done at 30°C unless otherwise stated.
Mating procedures and random spore analysis. Details were peviously described (Suda and Uchida 1974).
Determination of instability. Cells were precultured in YPG since the preculture in a non- fermentable medium was demonstrated to contain petites at the frequency which was equal to the petite-mutation rate (James and Spencer 1958). A small amount of cells was inoculated in YPG liquid medium and shaked for 3 to 5 days. The cells were transferred to YPD (1/100 inoculum size) and after being grown for 1 day, the cells were spread on YPD plates to form colonies. The respiratory capacity of the colonies were inspected by means of the color marker, adel (Sherman and Ephrussi 1962) or by tetrazolium overlay method (Ogur et al. 1957). The culture which contained more than 10% petites was, for convenience, de- fined as unstable.
Determination of loss or retention of the drug-resistance factors and of degree of suppressiveness of the petites. Cells of petite colony spontaneously generated were picked up by means of the color marker, adel, and crossed to a tester strain, Sll-17R or C814-40D. The mating mixture was inoculated in SCD (1/100 inoculum size) and allowed to stand for 1 day for zygotic growth.
One portion of the culture was used for the determination of loss or retention of the re- sistance factors of the petite. About 105 cells in the culture were spotted with a pipet on SCG plates with and without one of the drugs, oligomycin, erythromycin and chloramphenicol.
When the petite retained any drug-resistance factor, the zygote progeny could grow on SCG plates with and without the drug and when the petite had lost the resistance factor, they could grow only on SCG plate without the drug. The final concentrations of the drugs in the plates were 20 fig, 1 mg and 4 mg for oligomycin, erythromycin and chloramphenicol per ml of medium, respectively.
The other portion of the culture was used for the determination of degree of suppres- siveness of the petite. When the primary zygotes or the zygote progeny which had un- dergone only a few cell divisions from the cross between petites and grandes were plated, a considerable amount of variegated colonies (petites abcedees) were observed. As petites abcedees segregated to grandes and petites with generations of zygotes (cf. Fig. 1), the degree of suppressiveness of a petite was here determined by the use of the culture after the zygotic growth for 1 day in SCD liquid medium. The culture was spread on SCD plates after appropriate dilutions. The petite colonies were scored by the tetrazolium overlay method. The degree of suppressiveness was expressed by (X-Y)f(l-Y) after Sherman and Ephrussi (1962), where X was the fraction of petite colonies among total colonies and Y was the fraction of petites in the tester strain.
74 Kohta Sttda, Kayoko Suda, Akira Uohida
Results and Discussion
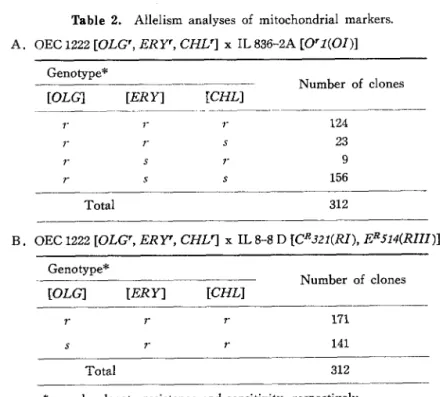
Allelism analyses of the mitochondrial markers. Allelism analyses were performed by crossing OEC1222 [OLC, ERY', CHL'] with IL836-2A [OR1{OI)\ and IL8-8D [CR321(RI), ER514 (RIII)]. The results are shown in Tables 2-A and 2-B. In Table 2-A, it was observed
Table 2. Allelism analyses of mitochondrial markers.
A. OEC 1222 [OLGr, ERY', CHL'} x IL 836-2A [O'1(OI)]
G e n o ty p e *
N u m b e r o f c lo n e s [O L G ] [E R Y ] [C H L ]
r r r 1 2 4 r r s 2 3 r s r 9 r s s 15 6
Total 312
B. OEC 1222 [OLGr, ERY', CHLr] x IL 8-8 D [CR321{RI), ERS14(RHI)]
Genotype*
[OL G] [ER T\ [CHL] Number of clones
171 141
Total 312
* r and it denote resistance and sensitivity, respectively.
that no oligomycin sensitive colonies appeared among 312 zygotic colonies whereas both erythromycin and chloramphenicol markers were segregated, for resistance and sensitivity, to the ratio of about 1 : 1 (The transmission frequencies of [ERY] and [CHL] alleles from
a parent were 47% and 43%, respectively). In Table 2-B, both erythromycin and chloram- phenicol sensitive colonies did not appear whereas oligomycin marker was segregated about 1 : 1 (The transmission frequency of [OLG] allele from a was 55%). These findings suggest that [OLGr], [ERYr] and [CHL'] resulted from the mutations at [01], [RIII] and [RI] loci,
respectively. Similar observations were made by other investigator (Birky personal com- munication).
Genetic analysis of instability. C3516-1A (a, adel) was an unstable strain. S51-41 R (a, ura 3, [OLGT, ERY', CHL']) was a segregant from zygotes made with OEC1222 (a, adel, his4, thr4, [OLG\ ERY', CHL']) and Sll-17R (a, uraS). A diploid strain, S55-R4, which wasr esistant to oligomycin, erythromycin and chloramphenicol was constructed by crossing C3516-1A to S51-41R. Since the population of S55-R4 cells contained petite mutants not exceeding 10
%, S55-R4 was determined to be stable concerning petite production, indicating that the
instability was under the control of a recessive gene(s). Further confirmation was done by the following random spore analysis of S55-R4. Haploid clones issued from S55-R4 asci
were selected by means of adel color marker. The cells of the adenine-requiring haploid colonies were examined form mating types and characters of the instability for petite produc-
tion. As shown in Table 3, it can be said that unstable and stable characters segregated 1 to 1 as well as mating types. It was, therefore, concluded that the instability, high muta-
bility to petites, was controled by a recessive nuclear gene.
Table 3. Random spore analysis of S55-R 4 diploid.
Mating type Stabi l ity Number of haploid clones
a s t a b l e 4 1 a u n s t a b l e 4 1 a s t a b l e 3 0 a u n s t a b l e 3 4
Total 146
The maintenance of the mitochondrial genome in yeast cells has been revealed to be controled by nuclear genes (Ephrussi and Hottinguer 1951 ; James and Spencer 1958) in- cluding pet loci (cf. Beck et al. 1971) as well as a mitochondrial gene itself (Handwerker et at. 1973). The contribution of nuclear genes for the yeast to the maintenance of the mito- chondrial genome is evident, but the mechanism has not been fully understood.
Loss or retention of the mitochondrial markers in spontaneously arising petites. An unstable and adenine-requiring strain, S55-13 R, was isolated from one of the S55-R4 segregants, carrying the three mitochondrial drug-resistance factors, [OLGr], [ERYr] and [CHLr], Cells of S55-13R were spread and spontaneously arising petite colonies were randomly selected.
The ratio of petite colonies to total colonies was about 30%. On a mechanism concerning instability of the unstable strain, one can assume that the overall mitochondrial genome of the unstable strain would be diluted out due to unequal rates between cell division and duplication of the mitochondrial genome and/or due to an unbalanced distribution of the genome to daughter cells (dilution model). If so, the petites from the unstable strain would retain no mitochondrial markers. To test this assumption, the petites from the unstable strain were analysed to see whether each mitochondrial marker had been lost or retained.
Table 4A shows the results together with those in the petites from a stable strain OEC1222, of spontaneous origin. One can see that there observed petites from both S55-13R and OEC1222 with the varieties of genetic constitutions of the mitochondrial markers and that more than 50% of the petites from both strains retained, at least, one of the three drug- resistance factors. These indicate that the majority of the spontaneously arising petites from both strains retained the mitochondrial genome, though partially defective. Therefore, it is difficult to elucidate the mechanism of higher rates of petite production in the unstable strain in terms of "dilution model", because the model predicts that the petites so produced may have no mitochondrial genome at all.
76 Kohta Siida, Kayoko Suda, Akira Uchida
Table 4A. Distributions of genetic constitutions of the spontaneously arising petites from S55-13R and OEC 1222, both of which harbor the cytoplasmic drug-resistance factors to oligomycin, erythromycin and chloramhenicol.
G e n e tic c o n s titu tio n *
S 55 ‑ 13 R O E C 1 22 2 [O L G ] [E R Y ] [C H L ]
r r r 5 7 86 r r o 5 1 r o r 0 1 r o o 2 3 5 1 o r r 3 4 2 3 o r o 6 1 o o r 0 2 o o o 10 1 87 T o ta l 2 2 6 2 5 2
F ra c tio n o f p e tite s (% 34 . 7 3 . 6
* r and o denote retention and loss, respectively, of the resistance factors in the petites.
Table 4B. Survival fractions of the three drug-resistnce factors and relative distances between the pairs of the three factors in the petite populations calculated from the data shown in Table 4A.
Strai n S 55-13R OEC 1222
Survial fraction Relatative distance between
[OLG] [ERY] [CHL] [OLGHERY] {ERY\-[CHL\ [CHL]-[OLG]
0.38 0. 55
0. 45 0. 44
0. 40 0.44
0.68 0. 69
0. 15 0. 06
0. 69 0. 70
Table 4 A shows that the spontaneous petites were frequently observed to loss or retain [ERY] together with [CHL] but [OLG] behaved itself differently from the other two markers.
If the breakage of the mitochondrial genome occurs at random sites, the chance of the two given markers to be lost simultaneously will reflect the distance of the two markers in the mitochondrial genome. To express this numerically, a method was proposed (Suda and Uchida 1974) which was analogous to that described by Nester and Lederberg (1961). When (X) and (Y) represent the survival fractions of petites retaining X and Y markers, respectively, in the petite population and (XY) represents the fraction of petites retaining both markers, the relative distance (D) between X and Y markers can be expressed by
D=(l -C)/(1 -Cu),
where
C=(XY)/[(X)+(Y)-(XF)] and
CB=(X)(F)/[(X ) +(F) -(X) (F)].
The culculated data are summarized in Table 4B. As seen in the table, the relative dis- tances between the two of the three markers calculated from the spontaneously arising petite population of S55-13R were almost comparable to those from the spontaneously aris-
ing petite population of OCE1222. From these observations it can be said that [ERY] and [CHL] were closely linked each other in the mitochondrial genome but [OLG] seemed not to be closely linked from the other two markers though carried by different strains. This linkage relationship is in agreement with that described by recombination analysis and by co-retention analysis with ethidium bromide (Uchida and Suda 1973; Suda and Uchida 1974).
Although almost the same linkage relationship of the mitochondrial markers was indicated in the preceding section, the unstable and stable strains showed different patterns as for the survival fractions of the markers (Table 4B). In the case of S55-13R, [OLGr] was the first, [CHLr] the second and [ERYr] the last to be lost from the mitochondrial genome.
On the other hand, [ERYr] and [CHL'] were almost simultaneously lost from the genome of OEC1222 and [OLGr] was followed. It can be envisaged that the mitochondrial genome of the unstable strain may be attacked at its specific site(s) by defect of the gene function responsible for maintenance of the mitochondrial genome. If so, the relative distances of the markers would differ from those obtained in the stable strain. But this was not the case.
An interpretation was presented that the order of the survival fractions of the mitochon- drial markers in an ethidium bromide-induced petite population would reflect the distances of the markers from a presumable attachment site of the mitochondrial DNA to the mem- brane structure (Suda and Uchida 1974). If the assumption that the farther a marker lies from the attachment site the more easily the marker will be lost is correct, it seems unlikely that the attachment site of the genome in the unstable strain would lie at the same point as that in the stable strain. One possible explanation is that the attachment site would not be fixed at a definite position of the mitochondrial genome but would differ from a strain to another, that is, the order of the survival fractions of the mitochondrial markers of a given strain will be different from that of another strain. In this connection, Linnane et al.
(1974) pointed out that multiple origins of the transmission of the mitochondrial genomes, Table 5. Genetic constitutions of the subclones from S55-13R petites (A, B
and C) and OEC1222 petites (D,E and F) with primary [OLGr, ERY', CHL'] type.
G e n e tic c o n stitu tio n * 蝣 S 5 5‑ 1 3 R I O E C 1 2 22 [O L G ] [E R Y ] [C H L ] A B D E F
r r r 3 4 0 6 14
r r o 0 0 0 0 0 0 r o r 0 0 0 0 1 0 T O O 0 1 1 1 2 1 o r r 2 2 0 l l 4 2 o r o 0 0 1 0 1 0 o o r 1 0 0 0 0 0 o o o 14 1 3 1 8 2 7 3
T o ta l 2 0 2 0 2 0 2 0 2 0 2 0
* r and o denote retention and loss, respectively, of the resistance factors.
78 Kohta Suda, Kayoko Suda, Akira Uohida
though intraclonal, to the zygote progeny upon mating must be considered in some crosses.
Instability of the mitochondrial genome of the petites during vegetative growth. Spontaneously arising petites which retained the mitochondrial markers, [OLGr], [ERYr] and [CHLr] were isolated from the unstable strain, S55-13R and from the stable strain, OEC1222. After each petite was grown for 1 day in YPD, cells were spread on YPD plates to form colonies.
Twenty colonies were selected at random and the cells of each colony were examined for loss or retention of the mitochondrial markers. The results in Table 5 show that the three mitochondrial markers of both S55-13R and OEC1222 were successively lost during vegeta- tive growth and that the markers of S55-13R were lost more frequently than those of OEC1222.
It was reported that the cytoplasmic drug-resistance factors of petites showed marked reductions in ability to transmit the factors to zygotic clones after vegetative growth (Gin- gold et al. 1969 ; Michels et al. 1974). The mitochondrial DNA was also degraded after ethidium bromide mutagenesis (Perlman and Mahler 1971). By subcloning, the primary petites were observed to be very heterogeneous with regard to the retention or loss of a mitochondrial erythromycin-resistance factor as well as the degree of suppressiveness (Nagley and Linnane 1972). Therefore, together with those findings, it appears that the mitochond- rial genome, once damaged by any event, is subsequently degraded to result in less infor- mative molecules. The degradation of the genome would probably be due to nucleolytic action and the degradation would be amplified in the case of the unstable strain.
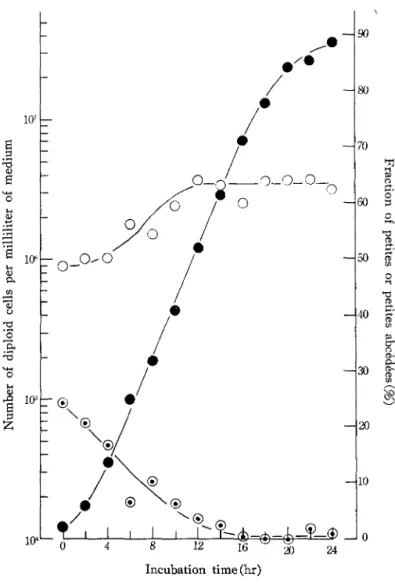
Determination of suppressiveness. On plating the primary zygotes arising from the cross between petite and grande, a considerable amount of variegated colonies (petite abcedees) were observed. The variegated colonies were decreased and, on the other hand, grande and petite colonies were increased with the growth of zygotes. An example is shown in Fig. 1, indicating that petites abcedees mitotically segregated to grandes and petites during the growth of zygotes. The segregation was completed by late exponential or stationary phase. The degree of suppressiveness was determined in further experiments by using the culture in which zygotes had been grown 1 day in the minimal liquid medium (SCD).
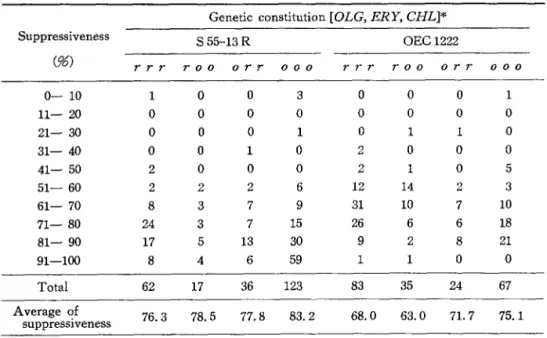
Suppressiveness of spontaneously arising petites from unstable and stable strains. The degrees of suppressiveness of the petites with various genetic constitutions of both unstable strain, S55-13R and stable strain, OEC1222 were studied. Spontaneously arising petites were isolated and crossed to tester strains. After growth of the zygotes for 1 day in SCD liquid medium, one portion of the culture was spread on SCD plates to form colonies after appro- priate dilutions, and the other portion was used for examining the ability to grow on SCG plates with and without the drugs. Table 6 shows that the majority of the petites from S55-13R and from OEC1222 were highly suppressive, only 4 among 238 petites for the former and 1 among 209 for the latter being neutral or low suppressive. As the petites without the mitochondrial DNA were expected to be neutral (Michaelis et al. 1971 ; Nagley and Linnake 1970 ; Nagley and Linnane 1972), or to be very low suppressive (Bech-Hansen and Rank 1973), almost all the spontaneously arising petites from both strains would have
io7 b-
10s
t-t
y
å a
I
105
b^_00
C ^"
104'
o
O_O_CL
o
70
60
®
I I I I L
ff-^^-ff-dU
=«4v=o u. it, 20
Incubation time (hr)
90
40
30
20
10
24
^
50 ff
§-
Fig. 1. An example of the time course of the growth of zygotes.
The mating mixture of the cross between a petite mutant with [OLGT, ERYr, CHL'~\ type originated from OEC1222 and a tester strain, Sll-17 R, was trans- ferred to SCD liquid medium. At given times of incubation, the samples were spread on SCD plates after appropriate dilutions. After a few days' incubation, the colonies of grandes, petites and petites abcedees formed on the plates were scored. % denotes the number of diploid cells per milliliter of medium; O and ® denote the fractions of petites and petites abcedees, respectively, in per cent.
the mitochondrial DNA, though defective. Therefore, the "dilution model", again, could not elucidate the mechanism of the production of petite mutants in the unstable as well as stable strains.
Table 6 shows that the average degree of suppressiveness of S55-13R petites (76.3-83.2
%) was higher than that of OEC1222 petites (63.0-75.1%). The mitochondrial genomes of S55-13R petites could be supposed to be more grossly damaged than those of OEC1222