学位論文
Doctoral Thesis
超過剰排卵由来マウス凍結卵子を用いた体外受精における
N-アセチルシステインの有用性 (N-acetyl cysteine restores the fertility of vitrified–warmed mouse oocytes derived through
ultrasuperovulation)
椋木 歩
Ayumi Mukunoki
熊本大学大学院医学教育部博士課程医学専攻
HIGO
プログラム専門コース指導教員
中潟 直己 教授
熊本大学大学院医学教育部博士課程医学専攻資源開発学
2020
年3
月学位論文
論文題名:超過剰排卵由来マウス凍結卵子を用いた体外受精における
N-アセチルシステインの有用性
(N-acetyl cysteine restores the fertility of vitrified–warmed mouse oocytes derived through ultrasuperovulation)
著 者 名: 椋 木 歩
Ayumi Mukunoki
指導教員名:熊本大学生命資源研究・支援センター 資源開発分野 中潟 直己教授
審査委員名:産科婦人科学分野 教授 片渕 秀隆 発生医学研究所 損傷修復分野 講師 立石 智 生命科学研究部 生体微細構築学分野 教授 若山 友彦 老化・健康長寿学講座 感覚・運動医学分野 准教授 三浦 恭子
2020
年3
月目次
要旨 --- 1
本論文の骨格となる参考論文 --- 2
略語一覧 --- 3
1. 研究の背景と目的 --- 4
2. 実験方法 --- 6
3. 実験結果 --- 10
3-1. 超過剰排卵誘起法由来卵子を用いた凍結保存における形態的に正常な卵子の割合の評価 --- 10
3-2. N-アセチルシステインが超過剰排卵誘起法由来凍結卵子を用いた体外受精の 受精率に及ぼす影響 --- 12
3-3. 卵子の凍結処理が透明帯中チオール基量に及ぼす影響 --- 13
3-4. N-アセチルシステインが透明帯中チオール基量および透明帯の膨化に及ぼす影響 --- 14
3-5. 超過剰排卵誘起法由来凍結卵子を用いた体外受精により作製された二細胞期胚の発生能 --- 16
4. 考察 --- 18
5. 結語 --- 21
6. 参考文献 --- 22
要旨
【目的】
生命科学研究において、遺伝子改変マウスは重要な研究資源として利用されている。近年、ゲノム編 集技術の進展に伴う遺伝子改変マウスの効率的な作製、保存を行うため、生殖工学技術が利用されてき ている。卵子の凍結保存技術は、いつでも体外受精が可能なことから、効率的な遺伝子改変マウスの作 製に有用である。しかしながら、既存の過剰排卵誘起法で作製できる凍結卵子数が少ないことや凍結/加 温卵子を用いた体外受精における受精率の低下が課題であり、技術改良が必要とされている。これまで に私たちは、新たな過剰排卵誘起法として、インヒビン抗血清を用いた超過剰排卵誘起法を開発し、1匹 の雌マウスから100 個の卵子を排卵させることに成功した。また、体外受精技術では、システイン類似
体であるN-アセチルシステイン(NAC)が受精率を向上させることを明らかにしている。これらの知見
から、卵子凍結保存の課題である卵子数や受精率の低下に対して、超過剰排卵誘起法とNACを用いた体 外受精法が有用であると考えた。そこで本研究では、卵子凍結保存技術を活用した遺伝子改変マウスの 効率的な作製システムを開発するため、超過剰排卵誘起法が凍結卵子の生存能に及ぼす影響(実験1)お よびNACが凍結卵子の受精能に及ぼす影響(実験2)を評価した。また、凍結卵子を用いた体外受精に おけるNACの受精能向上作用について、凍結操作による卵子透明帯中チオール基量(実験3)、NAC処 理による凍結/加温卵子透明帯中チオール基量および透明帯断面積(実験4)について評価した。さらに、
本技術で得られた二細胞期胚の発生能(実験5)を評価した。
【結果および考察】
実験1:超過剰排卵誘起法由来凍結卵子の加温後の生存能
既存の過剰排卵誘起法と比較し、超過剰排卵誘起法を用いることで、凍結卵子の作製効率が2倍となり、
加温後の形態的に正常な卵子の割合は既存の過剰排卵誘起法と同様に高値であった。
実験2:NACによる超過剰排卵誘起法由来凍結卵子の体外受精能に対する改善効果
NAC処理は、超過剰排卵誘起法由来凍結卵子の体外受精率を向上させた。
実験3:凍結操作が透明帯中チオール基量に及ぼす影響
卵子への凍結保護剤の処理は、透明帯中チオール基量を有意に減少させた。
実験4:NACが透明帯中チオール基量および透明帯の膨化に及ぼす影響
NAC処理は、透明帯中チオール基量を増加させ、さらに卵子の透明帯を膨化させた。
実験5:超過剰排卵誘起法由来凍結卵子によって得られた二細胞期胚の発生能
超過剰排卵誘起法由来凍結卵子を用いて得られた二細胞期胚は、体外培養において正常に胚盤胞期胚ま で発生し、さらに胚移植により正常に産子へと発生した。
【結論】
本研究において、超過剰排卵誘起法を用いることで、1匹の雌マウスから作製できる凍結卵子数を増加 させ、加温後の卵子に NAC を処理することで、凍結/加温卵子の受精能を向上させることに成功した。
さらに、超過剰排卵誘起法由来凍結卵子は正常に産子まで発生した。これらの知見より、本技術は遺伝 子改変マウスの効率的な作製システムの構築に繋がる有用な技術であり、今後の医薬研究の更なる発展 に寄与するものである。
本論文の骨格となる参考論文
本論文は、学術誌に掲載された次の論文を基礎とするものである。
1. Ayumi Mukunoki, Toru Takeo, and Naomi Nakagata. N-acetyl cysteine restores the fertility of vitrified–warmed mouse oocytes derived through ultrasuperovulation. PLoS One 2019 14(10):
e0224087.
その他の参考論文リスト
1. Toru Takeo, Ayumi Mukunoki, Naomi Nakagata. Ovulation of juvenile, mature, and aged female C57BL/6 mice following coadministration of inhibin antiserum and equine chorionic gonadotropin.
Theriogenology. 135 (2019) 1-6
本論文で使用した略号一覧表
本論文において、次の略語を使用した。
eCG : equine chorionic gonadotropin hCG : human chorionic gonadotropin FSH : follicle stimulating hormone IAS : inhibin antiserum
NAC : N-acetyl cysteine SPF : specific pathogen-free KLH : keyhole-limpet hemocyanin DMSO : dimethyl sulfoxide
DAP213 : 2 M DMSO, 1 M acetamide and 3 M Propylene glycol mHTF : modified human tubal fluid
TYH : Toyoda Yokoyama Hosi medium MBCD : methyl-β-cyclodextrin
cTYH : Toyoda Yokoyama Hosi medium with methyl-β-cyclodextrin KSOM/AA : potassium simplex optimized medium with amino acid
IASe : mixed solution of inhibin antiserum and equine chorionic gonadotropin COCs : cumulus oocytes complexes
FBS : fetal bovine serum
PBS-PVA : 1.0 mL phosphate buffered saline containing 1.0 mg/mL polyvinyl alcohol AFM : Alexa Fluor 488 C5 Maleimide
ICSI : intracytoplasmic sperm injection IVF : in vitro fertilization
GSH : reduced glutathione LH : luteinizing hormone
IMSR : International Mouse Strain Resource SD : standard deviation
1.
研究の背景と目的遺伝子改変マウスは、医学研究や新薬の開発に利用され、生命科学研究における重要な研究資源 である [1-3]。近年、ゲノム編集技術の発展により、ヒト疾患を模倣した精度の高い疾患モデルマウ スを短期間で作製できるようになり、遺伝子改変マウスの有用性がますます高まってきている [4]。
現在までに、世界中の研究機関において54,000系統を超える遺伝子改変マウスが作製されており、
これらの遺伝子改変マウスを効率的に活用するために、マウスの作製や保存に有用な生殖工学技術 の開発が求められている。
現在、遺伝子改変マウスの効率的な作製や保存には、精子や二細胞期胚の凍結保存技術が広く用 いられている [5]。精子や二細胞期胚を凍結保存することで、計画的な個体作製や系統維持に必要な スペースやコストの削減が可能になる。一方で、卵子の凍結保存技術は、体外受精における時間的 制約や飼育コストやスペースの削減に有用だと考えられているが、既存の過剰排卵誘起法で作製で きる凍結卵子数が少ないことや凍結/加温卵子を用いた体外受精における受精率の低下等、解決すべ き技術的課題が残されているため実用化されていないのが現状である [6]。
これまでに私たちは、新たな過剰排卵誘起法として、インヒビン抗血清(IAS)とウマ絨毛性性腺 刺激ホルモン(eCG)を同時に投与することにより、1 匹の雌マウスから 100 個の卵子を排卵させ る超過剰排卵誘起法を開発した(IAS + eCG: IASe)[7, 8]。また、体外受精技術では、還元型グル タチオン(GSH)、L-システインおよびD-システイン、システイン類似体であるN-アセチルシステ イン(NAC)が、透明帯のジスルフィド結合を切断し、透明帯を膨化させることにより、精子の侵 入を補助し受精率を向上させることを明らかにしている [8-10]。特に、NACは、他のシステイン類 似体に比べて溶媒中における安定性が高いことから、体外受精培地への応用に適している [8]。これ らの知見から、卵子凍結保存における課題である卵子数や体外受精における受精率の低下に対して、
超過剰排卵誘起法とNACを用いた体外受精法が有効ではないかと考えた。
そこで本研究では、卵子凍結保存技術を活用した遺伝子改変マウスの効率的な作製システムを開 発するために、超過剰排卵誘起法を用いて作製した凍結卵子の形態的に正常な卵子の割合および作 製効率やNACが凍結卵子を用いた体外受精における受精率に及ぼす影響を検討した。さらに、NAC 処理を行った凍結卵子の体外培養での発生能および移植による産子への発生能を評価した。以下に、
本研究から得られた知見を詳述する。
2.
実験方法2-1.動物
精子および卵子は、性成熟した 12 週齢以上の C57BL/6J 系統雄マウス(日本クレア)および 4
週齢のC57BL/6J系統雌マウス(日本クレア)からそれぞれ採取した。二細胞期胚の移植には、12
週齢以上のICR系統のマウス(日本クレア)を受容雌(レシピエント)として使用した。これらの マウスは、特定病原体除去(SPF)の飼育室内において、7-19時までは明期、19-7 時までは暗期、
室温は22 ± 1°C、固形飼料及び水は不断給与できる環境で飼育した。なお、全ての動物実験は、熊
本大学動物実験指針に準じて行った。
2-2.試薬
IASはKishiらの方法に準じて作製した [11]。KLH修飾下マウスインヒビン合成ペプチドを抗原
として、ヤギに免疫させた。免疫したヤギから血清を採取し、インヒビンに対するアフィニティを 評価することで、インヒビン抗体の力価を評価した。本実験においては、そのアフィニティ検査に おいて、力価の上昇が確認された抗血清を使用した。
卵子の凍結保存には、1 M ジメチルスルフォオキサイド(DMSO)含有リン酸緩衝液と DAP213
(2 M DMSO, 1 M アセトアミド, 3 M プロピレングリコール)を用いた [12]。凍結した卵子の加温 には、0.25 M スクロース含有PB1溶液を用いた。体外受精培地には、modified human tubal fluid(mHTF)
あるいは各種濃度(0.25, 0.5 mM)のNACを含有したmHTFを使用した [13, 14]。精子前培養培地 は、modified Krebs-Ringer bicarbonate solutionをもとに作製されたToyoda Yokoyama Hosi培地(TYH)
に、0.75 mMメチル--シクロデキストリン(MBCD)を添加したcTYHを使用した [15, 16]。また、
胚培養には、potassium simplex medium (KSOM/AA)を使用した [17]。全ての試薬は、4°Cで冷蔵 保存し、体外受精培地(mHTF)、精子前培養培地(cTYH)および体外培養培地(KSOM/AA)は使 用時に30分間インキュベーター内で加温した(37°C、5% CO2)。
2-3.超過剰排卵誘起法および採卵
超過剰排卵誘起は、Takeo & Nakagataの方法に従って行った [7]。雌マウスの腹腔内にIASとeCG の混合液(3.75 IU eCG + 0.1 mL IAS: IASe)を0.2 mL投与し、48時間後にhCGを腹腔内投与した。
hCG投与から15~17時間後に、雌マウスを頸椎脱臼によって安楽死させ、体外受精培地を被覆して
いる流動パラフィン内に採取した卵管膨大部を含む卵管を導入した。続いて、解剖針を用いて卵管 膨大部を切開し、卵丘-卵子複合体(COCs)を200 µLのmHTFあるいは各種濃度のNACを含有し たmHTFドロップ中に導入後、採卵した卵子を凍結保存した。
2-4.卵子の凍結保存
卵子の凍結保存は、Nakagataらの方法に従って行った [18]。まず、IASeおよびhCGを投与した4 週齢の雌マウスから採取したCOCsを含有しているmHTFのドロップに、最終濃度が0.1%になるよ うにヒアルロニダーゼ溶液を加え、1分間加温した後に、流動パラフィンで被覆した80 µLのmHTF を用いて3回洗浄を行い、卵丘細胞を剥離した。裸化された卵子は、20%のウシ胎児血清(FBS)を
含有した100 µL mHTFドロップに導入し、インキュベーター内で10分間培養を行った(37°C、5%
CO2)。次に形態的に正常な卵子のみを回収し、100 µLの1 M DMSO中に導入後、5 µLの1 M DMSO 溶液とともにクライオチューブ移し(40~100個/チューブ)、5分間静置した(0°C)。続いて、予冷 しておいた45 µLのDAP213をクライオチューブ内に添加し、5分間静置した後に、あらかじめ液体 窒素中で予冷しておいたケーンにクライオチューブを取り付け、液体窒素中に浸漬した。凍結卵子 の加温は、あらかじめ37°Cに温めておいた0.25 M スクロース溶液(0.9 mL)をチューブ内に添加 して行い、10回ほど静かにピペッティングを行った後に、100 µLのmHTFドロップ中に卵子を回収 した。続いて10分間インキュベーター内に静置した後に、回収した卵子を100 µLのmHTFドロッ プ中で2回洗浄し、回収卵子数および生存卵子数を確認し、形態的に正常な卵子の割合を評価した。
2-5.体外受精
形態的に正常な凍結/加温卵子を、200 µLのmHTFあるいはNAC含有mHTF(0.25, 0.5 mM)に導 入し、インキュベーター内で1 時間培養を行った(37°C、5% CO2)。一方、精子は、安楽死させた
した。採取した精子はインキュベーター内で1時間培養(37°C、5% CO2)した後に、凍結/加温卵子 を含む体外受精培地に3 µL添加し、インキュベーター内で共培養を行った(37°C、5% CO2)。媒精 から3時間後に、流動パラフィン内で被覆した80 µLのmHTFドロップで卵子を3回洗浄し、一晩 インキュベーター内で培養した(37°C、5% CO2)。受精率は、体外受精の翌日(媒精後24時間後)
に、二細胞期胚をカウントすることにより算出した(受精率:二細胞期胚数/(二細胞期胚数+未受 精卵子数) x 100)。なお、体外受精培地中の精子濃度は、800~1000 sperm/µLであった。
2-6.透明帯中チオール基量および透明帯断面積の評価
透明帯中チオール基量の評価は、Takeoらの方法に従って行った [8]。ポリビニルアルコール含有 リン酸緩衝液(PBS-PVA)に、10 mg/mL Alexa Fluor 488 C5 Maleimide(AFM, Life technologies)を溶 解し、ストック溶液を調製した。超過剰排卵誘起法によって得られたCOCsを0.1%のヒアルロニダ ーゼを用いて卵丘細胞を剥離した後、未処理(control)、1M DMSO処理群、1M DMSOおよびDAP213 処理群、凍結/加温群に分配した。各処理を行った卵子は、流動パラフィンで被覆した100 µL PBS-PVA ドロップ中で2回洗浄し、0.1 mg/mL AFMを含有した100 µL PBS-PVA中に導入し、30分間反応(室 温、遮光)させた。染色処理後、卵子を100 µL PBS-PVAで3回洗浄し、蛍光顕微鏡(Biorevo BZ-9000、
Keyence Co.)にて観察し、透明帯中のチオール基と反応した AFMの蛍光強度を画像解析ソフトに
て評価した(BZ-H2A version 1.42、Keyence Co.)。なお、透明帯中チオール基量は、AFMの相対的 蛍光強度=凍結保護剤(1M DMSOあるいはDAP213)処理卵子あるいは凍結/加温卵子の蛍光強度/
未処理卵子の蛍光強度により評価した。また、透明帯の断面積は、相対的透明帯面積=凍結保護剤
(1M DMSOあるいはDAP213)処理卵子あるいは凍結/加温卵子の透明帯断面積/未処理卵子の透明 帯断面積により評価した。
2-7.胚の体外培養および卵管内胚移植
二細胞期胚は、流動パラフィンで被覆した100 µLのKSOM/AAドロップで2回洗浄した後、イン キュベーター内で72~96時間体外培養を行った(37°C、5% CO2)。胚の観察は24時間ごとに行い、
胞期胚数(体外培養開始から72あるいは96時間後)を確認し、発生率(各発生ステージの胚数/二 細胞期胚数 x 100)を算出した。
胚移植は、Nakagata の方法に従って行った [19]。移植を行う前日に、精管結紮雄マウスと雌マウ スを同居させ、朝に交配が確認(プラグ形成を目視で観察)できた雌マウスをレシピエントとして 使用した。麻酔は、ドミトール、ミダゾラムサンドおよびベトルファールを混合した三種混合麻酔 を腹腔内投与によって行い(0.01 mL/g body weight)、麻酔が効いたことを確認した後に、レシピエ ントマウスの卵管を外科的に露出させ、両側の卵管内に各10個の二細胞期胚を、ガラスキャピラリ ーを用いて移植した。移植終了後は、拮抗剤としてアンチセダンを腹腔内投与し(0.01 mL/g body
weight)、覚醒するまで37°Cで保温した。産子は移植後 19日に確認、産子への発生率(産子数/移
植胚数 x 100)を算出した。
2-8.統計解析
本実験における統計解析は、Prism version 7.02(GraphPad Software)を用いて行った。得られた値 は、平均値±標準偏差(standard deviation, SD)で示し、パーセントデータは逆正弦変換した後に分 散分析を行い、t検定あるいはDunnettの方法により各実験区間の有意差検定を行った。
3.
実験結果3-1. 超過剰排卵誘起法由来卵子を用いた凍結保存における形態的に正常な卵子の割合の評価
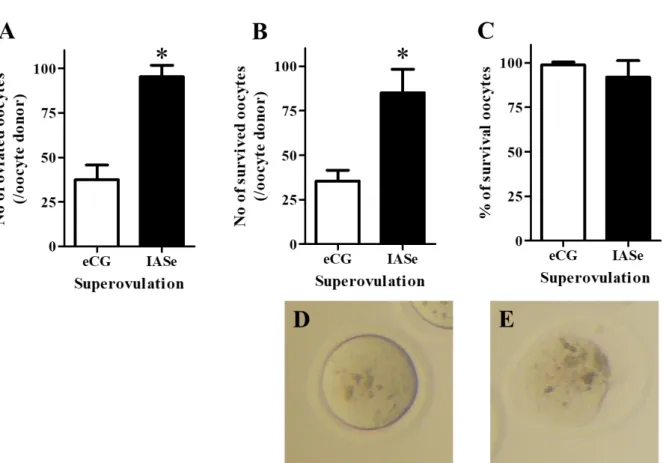
超過剰排卵誘起法(IASe)由来卵子における排卵卵子数、凍結卵子数、加温後の形態的に正常な 卵子の割合を、既存の過剰排卵誘起法(eCG)と比較した。その結果、IASe投与によって1匹のマ ウスから得られる卵子の数は約 3 倍になった(Fig. 1A)。また、それに伴いIASe 投与によって 1 匹のマウスから作製できる凍結卵子数も増加した(Fig. 1B)。また、IASe由来凍結卵子の加温後の 形態的に正常な卵子の割合も、eCG由来凍結卵子と同等であった(Fig. 1C)。本検討により、IASe 投与由来卵子は凍結保存技術へ応用が可能であると確認できたため、今後の検討では、IASe投与由 来凍結卵子を用いた。
Fig 1. Effect of superovulation induced by eCG or IASe treatment on the viability and yield of vitrified–
warmed oocytes.
A) The number of ovulated oocytes per oocyte donor was counted after removing the cumulus cells. B) The number of surviving oocytes per oocyte donor was counted after vitrifying and warming. C) The rate of
survival of oocytes (%) was calculated as the number of morphologically normal oocytes divided by the sum of morphologically normal and dead oocytes x 100. Morphologically normal oocyte (D) and dead oocyte (E) after vitrifying and warming are shown. The results are expressed as mean ± SD (n = 5). *P < 0.05 compared with eCG.
3-2. N-アセチルシステインが超過剰排卵誘起法由来凍結卵子を用いた体外受精の受精率に及ぼす影響
超過剰排卵誘起法由来凍結卵子の体外受精におけるNACの有用性を調べるために、0.25 mMあ
るいは0.5 mMのNACを添加した体外受精培地中に凍結/加温卵子を導入し、新鮮精子と体外受精
を行った。その結果、0.5 mM NACは、体外受精における受精率を上昇させた(Fig. 2)。
Fig 2. Effects of NAC on the fertilization rate of vitrified–warmed oocytes.
The fertilization rate was calculated as follows: total number of two-cell embryos divided / (the total number two-cell embryos + unfertilized oocytes) × 100. Results are expressed as mean ± SD (n = 6). *P < 0.05 compared with control (0 mM NAC).
3-3. 卵子の凍結処理が透明帯中チオール基量に及ぼす影響
凍結/加温卵子の受精能低下の要因として、透明帯中チオール基の酸化に伴う透明帯硬化が関与す ると考えられている。そこで本実験では、凍結卵子作製の際に行う各操作(1 M DMSO処理、DAP213 処理、凍結/加温操作)が、透明帯中チオール基量に及ぼす影響を評価した。その結果、1M DMSO 処理を行うことで、透明帯中チオール基量が低下しており、DMSO処理が透明帯硬化に関与するこ とが明らかになった(Fig. 3)。
Fig 3. Thiol levels following treatment with cryoprotectants.
Relative fluorescence intensity was calculated as follows: fluorescence intensity of vitrified–warmed oocytes precultured with NAC / fluorescence intensity of fresh oocytes. Results are expressed as mean ± SD (fresh: n
= 85, DMSO: n = 85, DAP213: n = 52, thawed: n = 86). *P < 0.05 compared with fresh oocytes [DMSO (−), DAP213 (−), vitrified–warmed (−)].
3-4. N-アセチルシステインが透明帯中チオール基量および透明帯の膨化に及ぼす影響
DMSO処理により生じた透明帯中チオール基の酸化に対するNAC の影響を調べるために、各濃 度(0, 0.25, 0.5 mM)のNACを含有した培地中で凍結/加温卵子を培養し、透明帯中チオール基量 を評価した。その結果、凍結/加温卵子の透明帯中チオール基量は、NACにより増加した(Fig 4)。
本知見から、NACが凍結/加温卵子の透明帯中ジスルフィド結合を切断し、チオール基を増加させて いることが明らかになった。
Fig 4. Effects of NAC on thiol levels in vitrified–warmed oocytes.
A, C, E): bright field, B, D, F): fluorescence, scale bar shows 50 µm. G) Thiol levels in the ZP were calculated as follows: Fluorescence intensity of each group / Fluorescence intensity of 0 mM NAC) × 100. Results are expressed as mean ± SD (n = 4). *P < 0.05 compared with control (0 mM NAC).
各種システイン類似体による受精能改善効果において、透明帯の膨化が関与していると考えられ ている。そこで本実験では、NAC が凍結/加温卵子の透明帯の膨化に及ぼす影響を調べるために、
NAC 処理が透明帯断面積に及ぼす影響を評価した。その結果、凍結/加温卵子の透明帯断面積は、
NAC処理により増加した (Fig 5)。
Fig 5. Effects of NAC on zona pellucida expansion in vitrified–warmed oocytes.
A) Cross-sectional areas of the zona pellucida (n = 31–34 oocytes) were measured. B) The extent of
expansion of the zona pellucida was calculated as follows: (cross-sectional area of each group / cross-sectional area in 0 mM NAC) × 100. Results are expressed as mean ± SD (n = 4). *P < 0.05 compared with control (0 mM NAC).
3-5. 超過剰排卵誘起法由来凍結卵子を用いた体外受精により作製された二細胞期胚の発生能
超過剰排卵誘起法由来凍結卵子を用いた体外受精によって得られた二細胞期胚の発生能を確認す るために、加温後の体外受精で得られた二細胞期胚の体外培養およびレシピエント雌マウスの卵管 内への移植を行った。その結果、超過剰排卵誘起法由来凍結卵子を用いた体外受精によって得られ た二細胞期胚は、体外培養において胚盤胞期胚まで正常に発生(Table 1)するとともに、移植によ って正常に産子まで発生した(Table 2)。なお、いずれの検討においても、NAC未処理の凍結/加温 卵子とNAC処理を行った凍結/加温卵子との間に、発生能に差はみられなかった。
Table 1. Effects of NAC on the development of vitrified–warmed oocytes
NAC (mM)
Number of inseminated
oocytes
Number of two-cell embryos (%)
Number of four-cell embryos (%)
Number of morulae (%)
Number of blastocysts
(%)
0 263 165
(65.5±5.5)
156 (94.5 ± 3.9)
150 (90.9 ± 9.1)
138 (83.6 ± 8.8)
0.25 261 207
(80.5±11.0)
199 (96.1 ± 5.0)
193 (93.2 ± 5.6)
169 (81.6 ± 7.5)
0.50 264 219
(85.9±9.1*)
214 (97.7 ± 2.8)
209 (95.4 ± 3.1)
181 (82.6 ± 13.5)
Vitrified–warmed oocytes were inseminated and fertilization rate was calculate as follows: Number of two-cell embryos / Number of inseminated oocytes) × 100. After IVF using vitrified–warmed oocytes, two-cell embryos were cultured in KSOM/AA. Developmental rate was calculated as follows: (Number of four-cell embryos, morulae, or blastocysts / Number of two-cell embryos) × 100. Results are expressed as
Table 2. The birth rate of vitrified–warmed oocytes derived from IVF with NAC
NAC
(mM) Oocyte
Number of transferred two-cell embryos
Number of recipients
Number of live pups (%)
0
Fresh
80 4 31
(38.8 ± 16.5)
0 Vitrified–
warmed
80 4 32
(40.0 ± 15.8)
0.5 Vitrified–
warmed
80 4 37
(46.3 ± 11.1)
Two-cell embryos were transferred in each oviduct of the recipient mice (20 embryos/recipient), and the number of pups was recorded after 19 days. Two-cell embryos derived from fresh oocytes were used as controls. The birth rate was calculated as follows: (Number of the pups / Number of transferred two-cell embryos) × 100. Results are expressed as mean ± SD (n = 4).
4.考察
本研究により、超過剰排卵誘起法を用いることで 1 匹の雌マウスから作製できる凍結卵子数が増 加し、形態的に正常な卵子の割合は凍結/加温卵子も高い値を有していることが示された。また、凍 結/加温卵子を用いた体外受精において、NACを凍結/加温卵子に処理することで受精率が向上した。
さらに、卵子の透明帯中チオール基量は、凍結操作によって減少するものの、NACを処理すること によって改善した。超過剰排卵誘起法由来凍結卵子を用いた体外受精で得られた二細胞期胚は、胚 移植により産子へと発生することが示された。以下に、本研究から得られた知見を考察する。
超過剰排卵誘起法(IASe)は、雌マウスから排卵される卵子数の増加に有用である[7, 20]。また、
超過剰排卵誘起由来卵子は、体外受精の受精率、凍結/加温した二細胞期胚の融解後における形態的 に正常な胚の割合および胚移植による産子への発生率について、既存の過剰排卵誘起法と同等の成 績を示すことを明らかにしている。本研究では、超過剰排卵誘起法由来卵子の凍結/加温後における 形態的に正常な卵子の割合を評価し、既存の過剰排卵誘起法と同様に高値を示すことを明らかにし た(Fig. 1)。超過剰排卵由来卵子の凍結保存が可能になったことで、遺伝子改変マウスの雌由来生 殖細胞の保存や凍結卵子を用いた体外受精による効率的な個体作製が可能になった。また、超過剰 排卵誘起法において 100 個以上の卵子を得るためには、4 週齢の雌マウスを使用する必要があり、
採卵と体外受精の時期が制限されていたが、卵子を凍結保存することで体外受精の時期を自由に選 択することも可能になった。
体外受精における検討では、NACが凍結/加温卵子の受精能の向上に有効であることを明らかにし た(Fig. 2)。凍結/加温卵子における受精能低下の原因として、透明帯硬化が関与することが知られ ている[21-23]。通常、透明帯硬化は、精子が卵子の細胞膜と接着することが刺激となり、卵細胞質 表面に存在する表層顆粒が崩壊し、透明帯の構造が変化することによって起こる [24]。また、透明 帯硬化には透明帯構成タンパク質のチオール基の酸化が関与しており、分子内あるいは分子間でジ スルフィド結合を形成し、透明帯の構造変化が起こる [25]。透明帯硬化は、受精時のみに起こるの
卵子の凍結保護剤であるDMSOは、チオール基の酸化作用を有すると共に、卵子の表層顆粒の崩壊 を誘発し、透明帯硬化および卵子の受精能の低下を引き起こす [27, 28]。本研究においても、1M DMSOを処理することによって、卵子の透明中チオール基量が低下しており、透明帯硬化が生じて いることが示唆された(Fig. 3)。一方で、NACは、透明帯中チオール基量を増加させたことから、
ジスルフィド結合の切断により透明帯硬化の抑制に関与した可能性がある(Fig. 4)。さらに、NAC は、凍結卵子の透明帯を膨化させたことから、透明帯硬化を引き起こした卵子に対しても有効であ ることが示された(Fig. 5)。これらの知見から、マウス凍結卵子における受精率低下の原因は、DMSO 処理による透明帯硬化であり、NACによる受精率の改善は透明帯硬化からの回復に起因すると考え ている。
これまでの報告から、NACは、凍結/加温卵子のミトコンドリア活性および胚の発生能を向上させ ることが知られている [29]。また、前核期受精卵の凍結保存では、1.5 mM NACを含有した凍結保 存液を用いることによって、活性酸素の産生を抑制し、加温後の胚の発生能を向上させることも明 らかとなっている [30]。さらに、抗酸化剤であるレスベラトロールが、凍結/加温卵子のミトコンド リア活性および胚の発生能を改善することが知られている [31]。本研究では、凍結/加温後の卵子に のみNAC処理を行っており、今後、凍結保護剤にNACを添加することにより、卵子の機能改善に 有効であるか否か等についても、検討する必要がある。
超過剰排卵誘起法由来凍結卵子およびNACを用いた体外受精において、二細胞期胚が作製され、
体外培養により胚盤胞期胚に発生し、胚移植により産子へと発生することを確認した(Table 1, 2)。 これらの結果から、凍結保存操作およびNACを用いた体外受精が、卵子の発生能に及ぼす影響は認 められず、遺伝子改変マウスの作製に利用可能であると考えている。一方で、近年の解析技術の進 歩により、過剰排卵誘起法、凍結保存および体外培養等の操作が、エピジェネティクス変化を引き 起こし、初期胚の遺伝子発現に影響するということも明らかになっていきている [32]。従って、本 知見で確立した技術を畜産やヒトの不妊治療などに応用する場合は、エピジェネティクス変化の影
以上、本研究において私は、超過剰排卵誘起法由来卵子を用いた卵子凍結保存法およびNACを用 いた体外受精法を開発し、遺伝子改変マウスの効率的な作製や保存に有用な技術を確立した。本技 術を用いてマウス凍結卵子を保管することは、遺伝子改変マウスの効率的な保存、凍結卵子を用い た迅速の個体の作製を可能にした。また、一個体から得られる卵子数の増加により、二細胞期胚の 凍結保存や個体作製に必要になる卵子ドナーの個体数を削減することが可能になり、動物実験を行 う際に遵守すべき3Rsの1つである「使用動物数の削減」を実現した。以上、本知見は、超過剰排 卵誘起法由来凍結卵子とNACを用いた体外受精技術を確立し、遺伝子改変マウスを用いた医学研究 や創薬研究の効率化と迅速化に有用だと考える。
結語
本研究では、超過剰排卵誘起法由来卵子を用いた凍結卵子の作製が可能であること、体外受精培 地にNACを添加することで受精能を向上させることができることを明らかにし、凍結卵子を用いた マウスの効率的な保存および作製技術を開発した。本知見は、卵子の凍結保存技術における課題で あった、卵子数の制限および加温後の体外受精における卵子の受精能低下を克服し、医薬研究にお ける重要な研究資源である遺伝子改変マウスの作製技術として応用することによって、今後の医薬 研究の迅速化に貢献する技術として広く利用されることが期待できる。
参考文献
1. Donahue LR, Hrabe de Angelis M, Hagn M, Franklin C, Lloyd KC, Magnuson T, et al.
Centralized mouse repositories. Mamm Genome. 2012;23(9-10):559-71. Epub 2012/09/05. doi:
10.1007/s00335-012-9420-4. PubMed PMID: 22945696; PubMed Central PMCID: PMCPMC3709583.
2. Nakagata N, Yamamura K. Current activities of CARD as an international core center for mouse resources. Exp Anim. 2009;58(4):343-50. Epub 2009/08/06. PubMed PMID: 19654431.
3. Sacca R, Engle SJ, Qin W, Stock JL, McNeish JD. Genetically engineered mouse models in drug discovery research. Methods Mol Biol. 2010;602:37-54. Epub 2009/12/17. doi:
10.1007/978-1-60761-058-8_3. PubMed PMID: 20012391.
4. Singh P, Schimenti JC, Bolcun-Filas E. A mouse geneticist's practical guide to CRISPR applications. Genetics. 2015;199(1):1-15. Epub 2014/10/02. doi: 10.1534/genetics.114.169771. PubMed PMID: 25271304; PubMed Central PMCID: PMCPMC4286675.
5. Landel CP. Archiving mouse strains by cryopreservation. Lab Anim (NY). 2005;34(4):50-7. Epub 2005/04/05. doi: 10.1038/laban0405-50. PubMed PMID: 15806091.
6. Moussa M, Shu J, Zhang X, Zeng F. Cryopreservation of mammalian oocytes and embryos:
current problems and future perspectives. Sci China Life Sci. 2014;57(9):903-14. Epub 2014/08/12. doi:
10.1007/s11427-014-4689-z. PubMed PMID: 25104318.
7. Takeo T, Nakagata N. Superovulation using the combined administration of inhibin antiserum and equine chorionic gonadotropin increases the number of ovulated oocytes in C57BL/6 female mice.
PloS one. 2015;10(5):e0128330. doi: 10.1371/journal.pone.0128330. PubMed PMID: 26024317; PubMed Central PMCID: PMC4449130.
8. Takeo T, Horikoshi Y, Nakao S, Sakoh K, Ishizuka Y, Tsutsumi A, et al. Cysteine analogs with a free thiol group promote fertilization by reducing disulfide bonds in the zona pellucida of mice. Biol Reprod. 2015;92(4):90. Epub 2015/02/27. doi: 10.1095/biolreprod.114.125443. PubMed PMID: 25715791.
9. Takeo T, Nakagata N. Reduced glutathione enhances fertility of frozen/thawed C57BL/6 mouse sperm after exposure to methyl-beta-cyclodextrin. Biol Reprod. 2011;85(5):1066-72. Epub 2011/07/23. doi:
10.1095/biolreprod.111.092536. PubMed PMID: 21778138.
10. Takeo T, Tsutsumi A, Omaru T, Fukumoto K, Haruguchi Y, Kondo T, et al. Establishment of a transport system for mouse epididymal sperm at refrigerated temperatures. Cryobiology. 2012;65(3):163-8.
Epub 2012/06/23. doi: 10.1016/j.cryobiol.2012.06.002. PubMed PMID: 22722060.
11. Kishi H, Okada T, Otsuka M, Watanabe G, Taya K, Sasamoto S. Induction of superovulation by immunoneutralization of endogenous inhibin through the increase in the secretion of follicle-stimulating hormone in the cyclic golden hamster. J Endocrinol. 1996;151(1):65-75. Epub 1996/10/01. PubMed PMID:
8943770.
12. Nakao K, Nakagata N, Katsuki M. Simple and efficient vitrification procedure for cryopreservation of mouse embryos. Exp Anim. 1997;46(3):231-4. Epub 1997/07/01. PubMed PMID:
9250485.
13. Kito S, Ohta Y. In vitro fertilization in inbred BALB/c mice II: effects of lactate, osmolarity and calcium on in vitro capacitation. Zygote. 2008;16(3):259-70. doi: 10.1017/S0967199408004619. PubMed PMID: 18578945.
14. Quinn P, Kerin JF, Warnes GM. Improved pregnancy rate in human in vitro fertilization with the use of a medium based on the composition of human tubal fluid. Fertil Steril. 1985;44(4):493-8. Epub 1985/10/01. PubMed PMID: 3902512.
15. Takeo T, Nakagata N. Combination medium of cryoprotective agents containing L-glutamine and methyl-{beta}-cyclodextrin in a preincubation medium yields a high fertilization rate for cryopreserved C57BL/6J mouse sperm. Lab Anim. 2010;44(2):132-7. Epub 2010/04/02. doi: 10.1258/la.2009.009074.
PubMed PMID: 20357317.
16. Toyoda Y. Studies on fertilization of mouse eggs in vitro. I. In vitro fertilization of eggs by fresh epididymal sperm. Jpn J Anim Reprod. 1971;16:147-51.
17. Ho Y, Wigglesworth K, Eppig JJ, Schultz RM. Preimplantation development of mouse embryos in KSOM: augmentation by amino acids and analysis of gene expression. Mol Reprod Dev. 1995;41(2):232-8.
Epub 1995/06/01. doi: 10.1002/mrd.1080410214. PubMed PMID: 7654376.
18. Nakagata N, Takeo T, Fukumoto K, Kondo T, Haruguchi Y, Takeshita Y, et al. Applications of cryopreserved unfertilized mouse oocytes for in vitro fertilization. Cryobiology. 2013;67(2):188-92. Epub 2013/07/13. doi: 10.1016/j.cryobiol.2013.06.011. PubMed PMID: 23846105.
19. Nakagata N. [Embryo transfer through the wall of the fallopian tube in mice]. Jikken dobutsu Experimental animals. 1992;41(3):387-8. PubMed PMID: 1505632.
20. Takeo T, Nakagata N. Immunotherapy using inhibin antiserum enhanced the efficacy of equine chorionic gonadotropin on superovulation in major inbred and outbred mice strains. Theriogenology.
2016;86(5):1341-6. doi: 10.1016/j.theriogenology.2016.04.076. PubMed PMID: 27242176.
21. Matson PL, Graefling J, Junk SM, Yovich JL, Edirisinghe WR. Cryopreservation of oocytes and embryos: use of a mouse model to investigate effects upon zona hardness and formulate treatment strategies in an in-vitro fertilization programme. Hum Reprod. 1997;12(7):1550-3. Epub 1997/07/01.
PubMed PMID: 9262294.
22. Carroll J, Depypere H, Matthews CD. Freeze-thaw-induced changes of the zona pellucida explains decreased rates of fertilization in frozen-thawed mouse oocytes. J Reprod Fertil.
1990;90(2):547-53. Epub 1990/11/01. PubMed PMID: 2250252.
23. Ishizuka Y, Takeo T, Nakao S, Yoshimoto H, Hirose Y, Sakai Y, et al. Prolonged exposure to hyaluronidase decreases the fertilization and development rates of fresh and cryopreserved mouse oocytes.
J Reprod Dev. 2014;60(6):454-9. Epub 2014/09/17. doi: 10.1262/jrd.2014-045. PubMed PMID: 25225080;
PubMed Central PMCID: PMCPMC4284320.
24. Hoodbhoy T, Talbot P. Mammalian cortical granules: contents, fate, and function. Mol Reprod Dev. 1994;39(4):439-48. Epub 1994/12/01. doi: 10.1002/mrd.1080390413. PubMed PMID: 7893493.
25. Iwamoto K, Ikeda K, Yonezawa N, Noguchi S, Kudo K, Hamano S, et al. Disulfide formation in
cross-linkages in hardening of the zona pellucida. J Reprod Fertil. 1999;117(2):395-402. Epub 2000/02/26.
doi: 10.1530/jrf.0.1170395. PubMed PMID: 10690208.
26. Larman MG, Sheehan CB, Gardner DK. Calcium-free vitrification reduces cryoprotectant-induced zona pellucida hardening and increases fertilization rates in mouse oocytes.
Reproduction. 2006;131(1):53-61. Epub 2006/01/03. doi: 10.1530/rep.1.00878. PubMed PMID: 16388009.
27. Vincent C, Pickering SJ, Johnson MH. The hardening effect of dimethylsulphoxide on the mouse zona pellucida requires the presence of an oocyte and is associated with a reduction in the number of cortical granules present. J Reprod Fertil. 1990;89(1):253-9. Epub 1990/05/01. doi: 10.1530/jrf.0.0890253.
PubMed PMID: 2374118.
28. Pickering SJ, Braude PR, Johnson MH. Cryoprotection of human oocytes: inappropriate exposure to DMSO reduces fertilization rates. Hum Reprod. 1991;6(1):142-3. Epub 1991/01/01. doi:
10.1093/oxfordjournals.humrep.a137248. PubMed PMID: 1874947.
29. Matilla E, Martin-Cano FE, Gonzalez-Fernandez L, Sanchez-Margallo FM, Alvarez IS, Macias-Garcia B. N-acetylcysteine addition after vitrification improves oocyte mitochondrial polarization status and the quality of embryos derived from vitrified murine oocytes. BMC Vet Res. 2019;15(1):31.
Epub 2019/01/19. doi: 10.1186/s12917-018-1743-2. PubMed PMID: 30654800; PubMed Central PMCID:
PMCPMC6337864.
30. Yue SL, Zhang YT, Wang SW, Sun M, Xing YC, Wen J, et al. EFFECT OF NAC ON MOUSE GV OOCYTE SURVIVAL AND SUBSEQUENT EMBRYONIC DEVELOPMENT FOLLOWING VITRFICATION. Cryo Letters. 2016;37(4):295-302. Epub 2016/12/08. PubMed PMID: 27925012.
31. Wang Y, Zhang M, Chen ZJ, Du Y. Resveratrol promotes the embryonic development of vitrified mouse oocytes after in vitro fertilization. In Vitro Cell Dev Biol Anim. 2018;54(6):430-8. Epub 2018/05/31.
doi: 10.1007/s11626-018-0262-6. PubMed PMID: 29845451.
32. Bakhtari A, Rahmani HR, Bonakdar E, Jafarpour F, Asgari V, Hosseini SM, et al. The interfering effects of superovulation and vitrification upon some important epigenetic biomarkers in mouse blastocyst.
Cryobiology. 2014;69(3):419-27. Epub 2014/10/14. doi: 10.1016/j.cryobiol.2014.09.379. PubMed PMID:
25307438.