九州大学学術情報リポジトリ
Kyushu University Institutional Repository
マウス味細胞の分化・増殖調節機構におけるインス リンとヒアルロン酸の働き
渡邉, 雄
http://hdl.handle.net/2324/4475044
出版情報:九州大学, 2020, 博士(歯学), 課程博士 バージョン:
権利関係:
マウス味細胞の分化・増殖調節機構における インスリンとヒアルロン酸の働き
渡邉 雄
九州大学大学院歯学府歯学専攻
指導教員 古谷野 潔 教授 九州大学大学院歯学研究院
口腔機能修復学講座インプラント・義歯補綴学分野
研究指導教員 重村 憲徳 教授 九州大学大学院歯学研究院
口腔常態制御学講座口腔機能解析学分野
目次
略語・・・1
発表論文・・・2
要旨・・・3
序論・・・5
材料と方法・・・7
第一章 マウス味覚器におけるインスリンシグナルの機能
緒言・・・15
結果・・・17
考察・・・26
第二章 マウス味覚器におけるヒアルロン酸の機能
緒言・・・31
結果・・・33
考察・・・47
総括・・・51
謝辞・・・53
参考文献・・・54
1
略語
4MU:4-methylumbelliferone ANOVA:analysis of variance BMP:bone morphogenetic proteins CM:conditioned medium
CV:circumvallate papillae Ca4:carbonic anhydrase 4
DMEM:Dulbecco's modified Eagle medium ELISA:enzyme-linked immuno sorbent assay ENaC:epithelial sodium channel
Entpd2:ectonucleoside triphosphate diphosphohydrolase 2 ES細胞:embryonic stem cells FGF:fibroblast growth factors FP:fungiform papillae
GAD67:glutamic acid decarboxylase 67 GFP:green fluorescent protein
GLUT4:glucose transporter member 4 GLUT8:glucose transporter member 8 Gnat3:guanine nucleotide-binding protein G(t)
subunit alpha-3(α-Gustducin)
HA:hyaluronic acid
HEPES:hydroxyethylpiperazine ethane sulfonic acid
Has:hyaluronan synthase
Hmmr:hyaluronan-mediated motility receptor
(RHAMM)
Hyal:hyaluronidase
IGFBP:insulin-like growth factor-binding protein
IFN:interferon
IGF:insulin-like growth factor
IGFR:insulin-like growth factor receptor
IR:insulin receptor
Lgr5:leucine-rich repeat-containing G-protein coupled receptor 5
mTOR:mechanistic/mammalian target of rapamycin
NT:non-taste epithelium NF-κB:nuclear factor-kaapa B OCT:optimal cutting temperature Otop1:otopetrin-1
PBS:phosphate-buffered saline PFA:paraformaldehyde Pkd:polycystic kidney disease PI3K:phosphatidylinositol 3-kinase PLCβ2:phospholipase C β2 QOL:quality of life
RT-PCR:reverse transcription polymerase chain reaction
SGLT:sodium/glucose cotransporter SNAP25:synaptosome-associated protein 25 SOX2:sex-determining region Y-box 2 SPF:specific pathogen free animals SSC:standard saline citrate SHH:sonic hedgehog TBS:tris buffer saline
TGF:transforming growth factor TNT:tris-NaCl-tween buffer TLR:Toll-like receptor Vim:vimentin
Wnt signal:wingless signal
2
発表論文
本論文の内容の一部は
Effects of insulin signaling on mouse taste cell proliferation
Shingo Takai, Yu Watanabe, Keisuke Sanematsu, Ryusuke Yoshida, Robert F. Margolskee, Peihua Jiang, Ikiru Atsuta, Kiyoshi Koyano, Yuzo Ninomiya, Noriatsu Shigemura
https://journals.plos.org/plosone/article?id = 10.1371/journal.pone.0225190
PLoS One, 2019 Nov. 12; 14(11): e0225190. doi: 10.1371/journal. pone. 0225190. eCollection 2019.
で発表済みである。
また、本文の内容の一部は下記の学会において発表した。
第62回歯科基礎医学会学術大会、鹿児島(Web開催)、2020年10月 モリタ優秀発表賞生理学部門 受賞
3
要旨
味蕾に含まれる個々の味細胞は絶え間なくターンオーバーを繰り返しているが、味蕾 としての恒常性は常に維持されている。このターンオーバーは、様々なケミカルメディ エーターの影響下にあることが予想されるが、その調節メカニズムには不明な点が多い。
本研究では、全身のエネルギー代謝において極めて重要な役割を担うホルモンであるイ ンスリンと、細胞外基質の主要な構成成分であり多様な生理活性を持つヒアルロン酸
(以下HA)に着目し、その末梢味覚器における働きを探索した。
はじめに、マウス末梢味覚器におけるインスリン関連分子の発現を探索した結果、イ ンスリン受容体は味蕾に広く発現していることわかった。味蕾前駆/幹細胞3次元培養 系である味蕾オルガノイドを用いた実験の結果、培地中にインスリンを添加すると、イ ンスリン濃度依存的にオルガノイドコロニーに含まれる味細胞の数、各種味細胞マーカ ーmRNAの発現量が有意に減少することが明らかとなった。さらに、インスリンシグナ リングの下流に存在し、細胞分裂や生存の調節に重要な働きを持つことが知られる mTOR(mechanistic target of rapamycin)は味細胞、またLGR5(leucine-rich repeat-containing G-protein coupled receptor 5)陽性の味蕾前駆/幹細胞に発現していた。味蕾オルガノイ ドで、mTOR経路を薬理学的に阻害すると、コロニー中の味細胞の増加、および各種味 細胞マーカーmRNAの発現量増加が見られた。以上の結果は、インスリン-mTOR経路 が味細胞の分化・増殖の調節に関わる可能性を示唆する。
次に、HA の末梢味覚器における役割を探索した。マウスの味蕾には HA 受容体であ るCd44とHmmr(RHAMM(receptor for hyaluronan-mediated motility))、およびHA合 成酵素のHas2、Has3(Hyaluronan synthase 2、3)とHA分解酵素のHyal1–4(Hyaluronidase
1–4)mRNA の発現が見られた。免疫組織化学染色では、CD44 が有郭乳頭領域に広範囲
に発現しており、HA自体も味蕾内、味蕾周囲に分布していることがわかった。さらに、
4
味蕾オルガノイド培地中にHA合成酵素阻害剤4-methylumbelliferon(4MU)を添加する と、4MU濃度依存的に味蕾オルガノイドのサイズ、および味蕾前駆/幹細胞マーカーで あるLgr5、間葉系細胞マーカーであるVim(Vimentin)、HA受容体であるCd44のmRNA 発現が有意に減少した。最後に、4MU を 40 日間継続してマウスに経口投与することに より、オルガノイドを用いた実験同様、各種味細胞マーカーのほか、Lgr5、VimのmRNA 発現が有意に減少した。さらに、この4MU投与マウスでは味蕾周囲の細胞増殖が劇的 に抑制されていることが明らかになった。以上の結果から、HAは味蕾、および周囲組 織で合成されており、味細胞に発現する HA の受容体を介して、味蕾前駆/幹細胞の分 化・増殖を調節することで味覚器のホメオスタシス維持に関与する可能性が示唆された。
5
序論
味蕾は食物中に含まれる呈味物質を感知するための感覚器官である。我々の舌表面の 味蕾は舌前方部の茸状乳頭(fungiform papillae: 以下FP)、舌後方部の葉状乳頭や有郭 乳頭(circumvallate papillae: 以下CV)に多く分布しており、1個あたり100個~150個 の味細胞が包含されている(Chandrashekar et al., 2006)。味蕾には解剖学的、生理学的 な特徴の違いから少なくとも4種類の味細胞(I型、II型、III型、IV型細胞)が含まれ ている。I型細胞は味細胞の中で最も数が多く、支持細胞として味蕾の形態維持にも関 わるが、その機能には不明な点も多い。II型細胞には甘味受容体(T1R2 + T1R3ヘテロ ダイマー(taste receptor type 1 family,members 2 + 3))、うま味受容体(T1R1 + T1R3 ヘテロダイマー(taste receptor type 1 family,members 1 + 3))、苦味受容体(T2R(taste receptor type 2 family))や、Gustducin、PLCβ2(phospholipase C β2)、といった分子が発 現しており、甘味、苦味、うま味の受容を担っている(Roper and Chaudhari, 2017)。III 型細胞には、OTOP1(otopetrin-1)やPKD2L3、PKD2L1(polycystic kidney disease 2-like 3や2-like 1)(Huang et al., 2006、Ishimaru et al., 2006、Teng et al., 2019)が発現する酸 味受容細胞が含まれており、味神経とシナプス接続を有することや GAD67(glutamate decarboxylase67)が発現しているのが特徴である(Roper and Chaudhari, 2017)。IV型細 胞は味蕾の基底部に位置する細胞群であり、味孔には接しておらず、味の受容には関係 しないとされる。SHH(sonic hedgehog)やSOX2(sex-determining region Y-box 2)等の 発現が見られ、味蕾の前駆/幹細胞がこのIV型細胞に含まれていると考えられている
(Miura et al., 2005、Okubo, Clark and Hogan, 2009、Shin et al., 2011、Castillo et al., 2014)。
味覚上皮は体内でも最もターンオーバーが早い器官の一つとして知られ、成熟味細胞 の寿命は1~2週間程度と考えられている(Beidler et al., 1965、Hamamichi et al., 2006、
Perea-Martinez et al., 2013)。近年、全ての味細胞は、LGR5(luecine-rich orphan G-protein-
6
coupled receptor 5)またはLGR4、LGR6を発現する味蕾前駆/幹細胞から分化・成熟す
ることが報告された(Yee et al., 2013)。味細胞分化・成熟の過程には、bone morphogenetic proteins(Zhou et al., 2006)、neurotrophin(Liebl et al., 1999)、fibroblast growth factors
(Petersen et al., 2011)、IGF1(insulin-like growth factor-1)(Biggs et al., 2016)、SHH
(sonic hedgehog)(Lu et al., 2018)、Wnt(wingless)タンパク質(Iwatsuki et al., 2007)
などの様々な因子の関与が指摘されているが、味細胞の分化・増殖の調節メカニズムに はまだ不明な点も多く、味蕾前駆/幹細胞の高い活性がどのように維持されているのか もわかっていない。味細胞の分化・増殖調節メカニズムを解明し、味覚の恒常性維持機 構を理解することは、動物の摂食行動を考える上で極めて重要である。本研究では、第 一章で全身の糖代謝を司るホルモンであるインスリン、および第二章では細胞外基質の 主要な成分であるヒアルロン酸(HA)に着目し、味蕾の維持機構への関与を明らかに することを目的とした。
7
材料と方法
・動物
マウスの飼育、およびすべての実験は、九州大学の倫理指針に基づいて実施した。す べての実験プロトコルと手順は、「National Institutes of Health Guide for the Care and Use of Laboratory Animals」に準拠したものであり、九州大学の動物実験委員会によって承認 された(承認番号A29-206、A20-292)。C57BL/6Jマウスは、日本チャールズリバーラ ボラトリーズジャパン(横浜、日本)から購入した。C57BL/6Jを遺伝背景にしたGAD67- GFP(glutamic acid decarboxylase 67-green fluorescent protein)マウスは、群馬大学 柳川 右千夫 教授により作製されたものを使用した(Tamamaki et al., 2003)。すべてのマウ スは、23℃で12/12時間の明暗サイクル、SPF(specific pathogen free)状態で維持され、
水および餌(CE-2、日本クレア、東京、日本)への自由摂取が可能な状態で維持した。
すべての実験において、8~12週齢の雄もしくは雌のどちらかを使用した。
・4MUの投与
4MU(Nakarai Tesque、京都、日本)を、0.4% キサンタンガム(MP Biomedicals、LLC、
USA)に懸濁し、C57BL6/Jマウス(n = 44、生後8~9週齢、25.0 ± 1.2 g)に体重あたり 1200 mg/kg、600 mg/kg、60 mg/kg、0 mg/kgの濃度で経口カニューレを用い胃内投与し た。投与期間中は毎日体重を計測した。その後、マウスをイソフルラン麻酔下で頸椎脱 臼にて安楽死させた後、舌からCVを摘出した。
・免疫組織化学染色
C57BL6/Jマウス(n = 4、生後8~12週齢、25.7 ± 4.2 g)、GAD67-GFPマウス(n = 3、生後8~12週齢、25.7 ± 1.9 g)の舌を4% ホルムアルデヒド(以下PFA)を含むリ ン酸緩衝液(以下 PBS)で 50分間浸漬固定した。ショ糖液で脱水した後(10%:1 時
8
間、20%:1 時間、30%:3 時間)、OCT(Optimal Cutting Temperature)コンパウンド
(Sakura Finetek、東京、日本)に包埋し凍結、10 μm厚の切片を作製し、これをシラン コートしたスライドガラスに貼付した。切片を、抗原賦活のためにHisto VT One(Nakalai Tesque)中にて80℃で20分間インキュベートした後、Blocking One溶液(Nakalai Tesque)
中で1時間インキュベートした。その後、切片を一次抗体で4℃にて一晩インキュベー トした。tris-NaCl-tween buffer(以下TNT)液で洗浄した後、スライドを二次抗体で2時 間インキュベートし、再度洗浄した。味蕾オルガノイドについては、ホールマウント免 疫組織化学染色を行った(Ren et al., 2014)。各ウェルからコロニーを1.5 mLエッペン ドルフチューブに集め、PBSで洗浄し、4% PFA を含む PBSで室温にて15分間固定し た。その後、TNT液で洗浄し、Blocking One溶液中で1時間インキュベートした後、一 次抗体で一晩、二次抗体で2時間インキュベートした。
味細胞における免疫蛍光はレーザー走査型顕微鏡(FV-1000、オリンパス、東京、日 本)を用いて観察し、画像はFluoviewソフトウェア(ver. 04. 01、オリンパス)を用い て取得した。GFP、T1R3、IR、およびMki67を発現する細胞数を決定するために、CV またはオルガノイドの水平切片において、各味蕾の陽性細胞数をカウントした。アーチ ファクトを除外するためにImage-ProPlus(ver.4.0、Mediacybernetics、MD、USA)を使 用した。シグナル密度がネガティブコントロール(一次抗体を省略)の味細胞の密度の 平均値 + 2 SDよりも大きい値を示す細胞を陽性とした。IRシグナルのみtyramide signal amplification kit Alexa 647 or 568(cat. no. T20926、B40956、Thermo Fisher Scientific、MA、
USA)を用いて増幅した。使用した抗体の一覧は表1に示す。
9
・味蕾オルガノイド
3-D味蕾オルガノイドは、過去の論文のプロトコルに準じて作製した(Ren et al., 2017)。
C57BL/6Jマウス(8~12週齢、25.1 ± 1.8 g、9匹を1回の実験に使用)のCVを摘出し、
トリプシンでシングルセル化した後セルストレーナ(cat. no. 352235、Corning、NY、USA)
でフィルタリング後、24ウェル超低付着ディッシュ(cat. no. CLS3473、Corning)を用
いてconditioned medium(以下CM)(500 μL/ウェル)中で培養した。インスリンの効
果を評価するために、インスリンフリー培地を調整、使用した:20% DMEM/F12 培地
(Life Technologies、OR、USA)、50% Wnt3a CM(Hans Clevers博士から提供されたWnt3a 産生細胞株(DMEM/F12培地中の125 μg/mL Zeocin によって選択培養)から作製)、
10
20% R-spondin CM(Jeffery Wittsett 博士より提供されたR-spondin細胞株(DMEM/F12 培地中の600 μg/mL zeocinによって選択培養)から作製)、および10% Noggin CM(pEAK-
Rapid細胞株(DMEM/F12培地中の400 μg/mL Zeocinによって選択培養)から作製)、
EGF(epitherial growth factor、50 ng/mL; cat. no. 315-09、Peprotech、NJ、USA)、N21-MAX insulin-free media supplement(2% vol/vol; cat. no. AR010、R&D Systems、MN、USA)、
B-27 supplement minus insulin(2% vol/vol; cat. no. A1895601、Life Technologies)、penicillin- streptomycin(1 × ; cat. no. 15140122、Thermo Fisher Scientific)、Matrigel(5% vol/vol; cat.
no. 356231、Corning)を添加した。単一細胞化した細胞については、細胞ダメージによ るアポトーシスを防ぐために、Y-27632(10 μM; cat. no. Y050330、Sigma-Aldrich、MO、
USA)を培地中に添加した。CM中のインスリン濃度をインスリンELISAキット(cat.
no. 10-1113-01、Mercobia、Sweden)を用いて定量した。本試験で使用した全てのCMの インスリン含有量は、このELISAキットの検出限界値(3 mU/L)以下であった。これ に各濃度のインスリン(0–50 nM; cat. no. 093-06471、和光、東京、日本)をCMに添加 した。mTOR の阻害には、ラパマイシン(20 ng/mL = 22.8 nM、cat. no. R-5000、LC
Laboratories、MA、USA)を培地に添加した。オルガノイドの明視野画像は、CKX-41(オ
リンパス)で取得し、Image Jソフトウェア(ver. 1.53g、NIH、MD、USA)で処理した。
・RT-PCR(Reverse transcription polymerase chain reaction)
マウスの味蕾組織を採取するために、動物(C57BL/6J マウス、n = 6、8~12週齢、
25.9 ± 2.5 g)をイソフルランで麻酔し、頸椎脱臼で安楽死させた。エラスターゼ注射(0.5–
1 mg/mL; Elastin Products、Owensville、MO、USA; 室温で10分間インキュベート)後、
舌の前部および後部の上皮を剥離した。周囲組織を除去した後、FP(マウス1匹あたり 50個)をガラスピペットでタイロード溶液(NaCl 140 mM、KCl 5 mM、CaCl2 1 mM、
MgCl2 1 mM、NaHCO3 5 mM、グルコース 10 mM、ピルビン酸ナトリウム 10 mM、HEPES
11
10 mM、NaOHでpH7.4に調整)中に採取した。CVを含む上皮は周囲の組織から解剖
し、von Ebner 腺を顕微鏡下で除去した後、ガラスピペットで味蕾を採取した。また、
味蕾を含まない舌上皮を1 mm2採取した。マウスの味覚組織またはオルガノイドのtotal RNAをFastGene RNA Premium キット(cat. no. FG-81050、日本ジェネティクス社、日 本)で抽出し、LSM NanoDrop ND-1000(Fisher Scientific)を用いてA260/A280で質、
濃度を評価した。cDNAは、各組織から等量の RNA を使用し SuperScript VILO master mix(cat. no. 11755050、Thermo Fisher Scientific)を使用して合成した。RT-PCRは以下 のように行った。15分95℃(1サイクル);30秒 94℃、30秒 58℃、60秒 68℃(35 サイクル);5分 75℃(1サイクル)。増幅産物は、ゲル電気泳動を用いて、紫外線照 明下で可視化した(GelRed nucleic acid gel stain(cat. no. 41001–41003-T、Biotium))。
ゲノムDNAからのシグナルを排除するために、すべてのプライマー対は、各プライマ ーが異なるエクソンからのものであるように設計した。精製されたRNAサンプルを、
逆転写酵素を加えないサンプルも同時に作製した。定量RT-PCRについては、Fast SYBR green master mix(cat. no. 4385612、Fisher Scientific)を使用し、各サンプルについて3つ のreplicate 反応の平均値を用いた。データは、StepOneソフトウェア (ver. 2.3、Applied
Biosystems、CA、USA)を用いて分析した。すべての結果は、各サンプル中のGapdh遺
伝子を参照として、ΔΔCt 法を用いて定量した。各遺伝子に使用したプライマーは表 2 に示す。
12
13
・In situ hybridization
In situ hybridizationは通法に基づき行った(Shigemura et al., 2004)。目的遺伝子のRT- PCR産物を精製し(In situ hybridizationのプライマー配列は表3に記載)、pGEM T-Easy vector(Promega、Madison、WI、USA)でサブクローニングした。次に、ジゴキシゲニ
ン(以下DIG)標識アンチセンスおよびセンスRNAプローブを、SP6/T7 RNA polymerase
kit(Roche、Mannheim、Germany)を用いて調製した。試料はC57BL/6Jマウス(n = 4、
8~12週齢、25.5 ± 3.8 g)の舌を、OCTコンパウンド(Sakura Finetek)に埋め込み、厚 さ6 μmの切片を作成し、これをシランコートスライドガラス上に貼付した。凍結切片
を4% PFA in PBSで室温にて10分間固定し、5 × 標準食塩-クエン酸緩衝液(以下SSC)
にて室温で15分間洗浄した後、プレハイブリダイゼーション溶液(5 × SSC/50% ホル ムアミド)で室温にて1時間インキュベートした。50% ホルムアミド、5 × SSC、5 × デ ンハルト液、250 μg/mL酵母tRNA、500 μg/mL変性サケ精巣DNA、1 mMジチオスレイ
トール、20–200 ng/mLアンチセンスRNAプローブを含むハイブリダイゼーションバッ
ファー中にて 60℃で 18 時間ハイブリダイゼーションを行った。その後、切片を 5 ×
SSC/50%ホルムアミドで洗浄(5分間 × 2回)、0.2 × SSC/50%ホルムアミドで洗浄した
(60分間 × 2回)。その後、切片を50 mM Tris/HCl(pH 7.5)と150 mM NaClからな るトリス緩衝生理食塩水(TBS)に室温で 5 分間浸漬した後、1% ブロッキング試薬
(Roche)を含むブロッキング液で60分間インキュベートし、アルカリホスファターゼ
(1: 400希釈、Roche)で標識したanti-DIG Fabフラグメントを含むブロッキング液中 で60分間、室温でインキュベートした。TNTバッファーで洗浄(5分間 × 3回)した 後、切片をalkaline phosphatase buffer(100 mM Tris/HCl(pH 9.5)、100 mM NaCl、50 mM MgCl2)に5分間浸漬した。発色基質として、nitoroblue-tetrazolium chlorideと5-bromo-
4-chloro-3-indolyphosphateを用いてシグナルを可視化した。各遺伝子のmRNAのシグナ
ル特異性は、対応するセンスプローブをネガティブコントロールとして用いて確認した。
14
・統計解析
標準誤差の計算を含む統計解析は、IBM SPSS Statistics(ver. 19、IBM、NY、USA)を使 用して実施した。各インスリン濃度の定量RT-PCR データを評価するために、one-way
ANOVA、およびpost-hoc Tukey HDS検定を使用した。オルガノイドに対するラパマイ
シンの効果を評価するために、t-testを使用した。各4MU濃度の定量RT-PCRデータと 群間体重変化の差を評価するために、one-way ANOVA、two-way ANOVA、post-hoc Tukey HDS検定、およびunpaired t-testを使用した。
15
第一章 マウス味覚器におけるインスリンシグナルの機能
緒言
インスリンは、血糖値の上昇に応じて膵島β細胞から放出され、脂肪細胞および骨格 筋でのグルコース取り込みを促進し、肝臓でのグルコース産生を阻害することによって 血糖値を低下させ、炭水化物および脂肪代謝の調節において極めて重要な役割を果たし ている。過去の研究では、げっ歯類の味細胞において、インスリンとinsulin receptor(IR)
subunit α(IRα)の一部は成熟味細胞のマーカーであるkeratin 18と共発現することが示
されている(Zhang et al., 1995、Suzuki et al., 2005、Doyle et al., 2018)。また最近、マウ ス有郭乳頭では、インスリン自体がphosphplipase C beta-2(PLCβ2:II型味細胞マーカ ー)やsynaptosome-associated protein 25(SNAP25:III型味細胞マーカー)で合成されて いるという報告もなされた(Doyle et al., 2018)。単離味細胞を用いた電気生理学的実験 では、インスリンが塩味感受性に影響を与える可能性が報告されている(Heck GL et al.,
1984、Baquero et al., 2011)。しかし、味蕾における詳細なIRの発現様式は不明であり、
またインスリンシグナルが末梢味覚器でどのような機能を持つかについてはほとんど 解明されていない。
インスリンは様々な組織で細胞の成長と分化に関与していることが報告されている
(Yoon et al., 2017)。例えば、インスリンは、細胞の成長やタンパク質合成、オートフ ァジーの調節において重要なセリン―スレオニンキナーゼである mTOR を活性化する
(Dazert et al., 2011、Avruch et al., 2009、Ma et al., 2009、Zoncu et al., 2011)。また、膵 臓由来の培養細胞であるαTC1細胞を用いた実験では、インスリン添加によりmTORシ グナルが活性化され、濃度依存的に細胞が増殖するという報告がある(Liu et al., 2011)。
16
味細胞の分化や増殖においても、インスリンシグナルと下流の mTOR が重要な働きを 担うことが想定されるが、その理解は全く進んでいない。
そこで本研究では、マウス味覚器におけるインスリンシグナルの役割について解析を 行った。まず、マウス舌のFPとCVの味蕾でIr のmRNAとタンパク質の発現を調べ た。次に、マウスCV味蕾前駆/幹細胞由来の味蕾オルガノイドを用いて、インスリン が味細胞の分化・増殖に対する影響について調べた。さらに、オルガノイドの実験系を 用い、インスリン受容体の下流分子である mTOR を薬理学的に阻害することにより、
味細胞の分化・増殖におけるmTORの関与について検討した。
17
結果
味細胞ではインスリン受容体が発現していた
RT-PCRにより、Ir mRNAはCVとFPの両方で発現していることがわかった。一方、
非味覚上皮(NT)でのIrのmRNA発現は極めて弱かった(図1A)。免疫組織化学染 色による解析では、T1R3陽性味細胞の約80%、GAD67-GFP陽性味細胞の約60%がIR を発現していた(IRとT1R3またはGAD67との共発現比率は、CVでは (T1R3 + IR)/IR
= 225/281 (80.1%)、(GAD67 + IR)/IR = 111/159 (69.8%)、FPでは (T1R3 + IR) /IR = 73/88 (83.0%)、(GAD67 + IR)/IR = 18/30 (60.0%) )(図1B、C)。IR発現は味蕾全体で見ら れ、T1R3 や GAD67-GFP を発現していない多くの味細胞でも IR シグナルが観察され た。また、LGR5 陽性の味蕾前駆/幹細胞でも IR シグナルが認められたことから味蕾 前駆/幹細胞にもインスリンが作用する可能性が示唆された(図 1D)。一次抗体を用 いない同じサンプルでは特異的な蛍光は観察されなかった(図1E)。
18
(Takai et al., 2019 より改変)
図1.マウス味覚組織におけるインスリン受容体mRNAとタンパク質の発現
(A)有郭乳頭(CV)、茸状乳頭(FP)、味蕾を含まない上皮(NT)でのRT-PCR解析。(B、
C)マウス味覚器における T1R3(緑)、GAD67(赤)、IR(シアン)の免疫組織化学染色像。
CV(B)およびFP(C)。矢印および矢頭は、それぞれIR + GAD67共発現細胞とT1R3 + IR共
発現細胞を示す。(D)CV底部におけるT1R3(緑)、IR(赤)、LGR5(シアン)の免疫組織 化学染色。矢印はIR + LGR5共発現細胞を示す。(E)一次抗体を省略した切片。scale bar: 50 μm。点線は味蕾を示す。
19
インスリンはオルガノイドコロニー中の味細胞を減少させた
味細胞の増殖に対するインスリンの効果を調べるために、単離した味蕾前駆/幹細胞 に様々な濃度のインスリンを添加し培養した。味蕾の前駆/幹細胞は主にCVのトレン チ底部に位置しているため、CV組織を採取して単一細胞に分離した後Matrigelを含む
Conditioned Medium(CM)で培養することで、3次元コロニーを作製した。20日間培養
したほとんどのオルガノイドコロニーにおいて、インスリンを含まない CM 中では
Gustducin陽性 II型味細胞や CA4陽性 III 型味細胞などが含まれていた(図2A)。ま
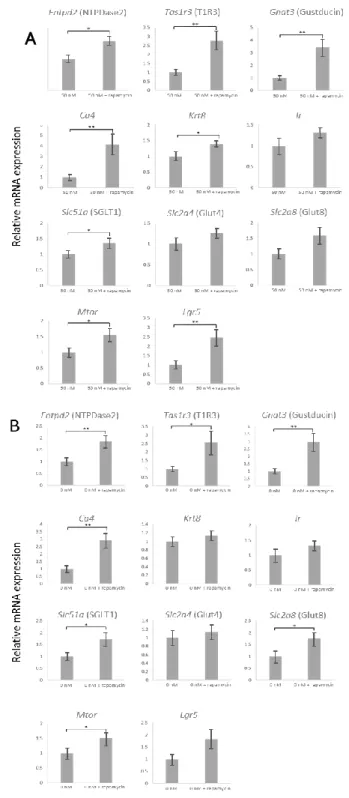
た、インスリン濃度の上昇に伴ってオルガノイドの平均コロニーサイズの増加傾向が見 られたが、サイズのばらつきが大きいため統計学的有意差はなかった(図2E)。
Gustducin 陽性味細胞を含むオルガノイドのコロニー数は、インスリン濃度の上昇に
伴い減少した(図2A–D、F)。また、定量RT-PCR解析では全ての味細胞マーカー遺伝
子 Entpd2、Tas1r3、Gnat3(Gustducin)、Ca4、Krt8、味蕾に発現する糖輸送体 Slc51a
(SGLT1: sodium-glucose cotransporter 1)、Slc2a8(GLUT8: glucose transporter 8)、Mtor、 および味蕾前駆/幹細胞マーカー(Lgr5)の mRNA 発現がインスリン濃度依存的に減 少した。(図3)。
20
(Takai et al., 2019 より改変)
図2.味蕾オルガノイド中の味細胞の分化・増殖に対するインスリンの作用
(A–D)様々なインスリン濃度のCM中で培養した味蕾オルガノイド。(A)インスリン0 nM、
(B)インスリン1 nM、(C)インスリン10 nM、(D)インスリン50 nM。抗Gustducin(マゼ ンタ)抗体と抗CA4(緑)抗体を用いた20日目の免疫組織化学染色像。scale bar: 50 μm。(E)
各インスリン濃度におけるオルガノイドコロニーの大きさと数を7日目、14日目、20日目に測 定した結果。データは平均値 ± SEMで示す。(F)全コロニー中のGustducinを含むコロニーの 割合。
21
(Takai et al., 2019 改)
(Takai et al., 2019 より改変)
図3.味蕾オルガノイドにおける味細胞マーカーmRNA発現に対するインスリンの効果
様々な濃度のインスリンを添加し 20 日間培養した味蕾オルガノイドを用いた定量 RT-PCR 解析の結果。データは平均値 ± SEMで示す。* p < 0.05、** p < 0.01、*** p < 0.001、one-way ANOVAおよびpost-hoc Tukey HDS検定。
22
mTOR阻害により味細胞マーカーmRNAの発現が増加した
mTOR経路は、インスリンを含む様々な成長因子によって活性化される(Zoncu et al.,
2011)。RT-PCRにより、mTORのmRNAはCV、FP、およびNTで発現していること
がわかった(図4A)。免疫組織化学染色による解析では、mTORのタンパク質が、IR 陽性細胞、T1R3陽性細胞、およびCA4陽性細胞を含むCVの味細胞において広範に発 現していることが確認された(図4D)。さらに、味細胞におけるmTORの役割を調べ るために、mTOR阻害剤であるラパマイシン20 ng/mL(22.8 nM)をCMに添加し、味 蕾オルガノイドを 20 日間培養した。ラパマイシンはmTOR の複合体のうちmTOR 複
合体1(mTORC1)を阻害することが報告されている(Zoncu et al., 2011)。結果、イン
スリンの有無にかかわらず、ラパマイシン添加は、Gustducin 陽性味細胞を含むコロニ ー数の割合を増加させた(図5 A、B、E)。ラパマイシン添加により、オルガノイドの 平均コロニーサイズは減少したが、個々のコロニーサイズのばらつきの大きさから統計 的に有意な差は見られなかった。また、定量RT-PCR解析の結果、50 nMインスリンCM では、ラパマイシン添加でEntpd2、T1r3、Gnat3、Ca4、Krt8、Slc51a、Mtor、およびLgr5 でmRNA発現量の有意な増加が認められた。0 nM インスリンCMでは、ラパマイシン
添加でEntpd2、T1r3、Gnat3、Ca4、Slc51a、Slc2a8、およびMtorのmRNA発現量が有
意に増加した(図6 A、B)。これらの結果から、mTOR経路が味細胞の分化・増殖に 関与している可能性が示唆された。
23
(Takai et al., 2019 より改変)
図4.マウス味覚組織におけるmTORの発現
(A)CV、FP、およびNTにおけるRT-PCR解析。(B–D)CVにおけるmTOR(緑)の免疫 組織化学染色像。(B)IR(マゼンタ)、(C)T1R3(赤)、(D)CA4(シアン)との二重染色 像。scale bar: 20 μm。点線は味蕾を示す。
24
(Takai et al., 2019 より改変)
図5.mTORC1である阻害剤ラパマイシンによるの味細胞の分化・増殖への影響
(A、B)インスリンを含まない培地(A)および50 nMのインスリンを含む培地(B)にラパ マイシンを添加して培養した20日目の味蕾オルガノイドの免疫組織化学染色像(Gustducin(マ ゼンタ)およびCA4(緑))。scale bar: 50 μm。(C、D)その際の平均コロニーサイズ。インス リン0 nM(C)および50 nMインスリン(D)。データは平均値 ± SEMで示す。(E)20日目 のコロニー数とGustducin陽性細胞を含むコロニーの割合。
25
(Takai et al., 2019 より改変)
図6.ラパマイシン添加による味細胞マーカーmRNA発現量の変化
(A、B)培地に22.8 nMラパマイシンを添加し、20日間培養したオルガノイドの各種味細胞
マーカーの発現量。インスリンを含まない培地(A)および50 nMインスリンを含む培地(B)。
データは平均値 ± SEMで示す。unpaired t-test。* p 0.05、** p < 0.01。
A
26
考察
本研究では、マウス味覚器におけるIrのmRNAとタンパク質の発現を明らかにした。
味蕾オルガノイドを用いた実験では、インスリンが味細胞の分化・増殖を調節すること を示した。また、mTORC1(mechanistic target of rapamycin complex 1)阻害剤であるラパ マイシン添加実験により、インスリンの作用は、mTORを介して生じている可能性が示 唆された。
味蕾オルガノイドは、幹細胞マーカーLGR5またはLGR6を発現する細胞から機能的 な味細胞を分化させる3次元幹細胞培養法である(Ren et al., 2014)。味蕾オルガノイ ド作製の基本的手順は、腸陰窩から腸管オルガノイドを培養するために開発された方法 を踏襲している(Sato et al., 2009、Schuijers et al., 2012)。これらの研究で使用した培地 には、3種の成長因子(Wnt3a、R-spondin、Noggin)や、B-27、N2、EGFなど多くのサ プリメントが含まれている。これらの成分のうち、R-spondinとNogginのCMを作製す るために使用する無血清培地Opti-MEM といくつかのサプリメント(B-27と N2)は、
生理的濃度をはるかに超えるインスリンを含んでいる(従来の培地は5000倍希釈でも
MercodiaインスリンELISAキットの測定限界(200 mU/L)よりも高い)ため、腸管オ
ルガノイド用の CM を用いて味蕾前駆/幹細胞の増殖に対するインスリンの効果を評 価することはできなかった。そこで、インスリンを含まない培地を用いて改変CMを調 製し、インスリン ELISA キットによる定量において検出限界以下の濃度であることを 確認した。In vivoでは、マウスの正常な血中インスリン濃度は8時間の絶食後で16.4 ±
4.5 μU/mL(0.11 ± 0.031 nM)程度であり、高脂肪食を16週間与えた後でも、その血清
インスリン濃度は170.9 ± 14.4 μU/mL(1.19 ± 0.01 nM)であった(Surwit et al., 1988)。
多くの細胞培養研究では、インスリンによる増殖効果を期待して高濃度のインスリンが 添加されている。それと比べると、本研究で用いたインスリンの濃度は、比較的高イン
27
スリン血症を発症した動物のそれに近い。味蕾オルガノイドシステムは、味覚器を多く の面で模倣しているが、味覚組織やその周辺環境を完全に再現することはできていない。
インスリンを含む様々な成長因子の相互作用を明らかにすることが、このシステムを発 展させるための次のステップとなるかもしれない。
味蕾で発現していることが報告されているグルコーストランスポーターである
SGLT1(Slc21a)やGLUT8(Slc2a8)のmRNAレベルは、インスリン投与により低下し
たが、GLUT4(Slc2a8)のmRNA発現には有意な変化は認められなかった(図3)。こ
れは、GLUT4 の発現が味細胞に限らず、味蕾を含まない上皮細胞や間葉系の細胞でも 発現しているためと考えられる(Yee et al., 2013)。我々が作出したオルガノイドはマ ウスのCVとその周囲の組織に由来するものである。そのため、味蕾前駆/幹細胞と味 蕾を含まない上皮の前駆細胞が含まれており、一部のコロニーは味細胞を含まない上皮 組織から成立した可能性があるため味細胞特異的にGLUT4発現の変化を観察すること は困難であったと考えられる。
mTORは細胞の栄養状態を感知することが知られている。様々な刺激に対し、細胞の 成長、増殖、発生、またオートファジーを制御する 2 つの mTOR 複合体(mTORC1、
mTORC2)の存在が報告されている(Ma et al., 2009)。インスリンはphosphatidylinositol
3-kinase(PI3K)やRasに依存するシグナル経路を制御することでmTORC1を活性化す
ることが知られている(Roux et al., 2004)。我々のデータでは、味細胞とその周囲の味 蕾を含まない上皮組織の両方でmTOR が発現しており、T1R3 発現細胞や CA4 発現細 胞を含む多くの味細胞でIRとmTORのシグナルが共発現していることが明らかとなっ
た(図4)。mTORC1阻害剤であるラパマイシンは、インスリンの作用を阻害し、オル
ガノイドにおける味細胞の増殖を促進した(図5A、5E、6A)。インスリンを含まない CMでも、mTORC1阻害は味細胞の生成を増加させた(図5B、5E、6B)が、これはお
28
そらく培地中のグルコースやアミノ酸によって mTOR シグナル伝達が既にある程度活 性化していたことによると考えられる。過去の研究では、T1R1 + T1R3うま味受容体が アミノ酸誘導性の mTOR シグナル伝達を調節している可能性が示唆されていることか ら、特定の味覚刺激もmTORを活性化する要因となるかもしれない(Wauson et al., 2012)。
また、mTOR の阻害は、放射線誘発性唾液腺損傷を改善することが報告されている
(Morgan-Bathke et al., 2014)。ラパマイシン類似物質はオートファジーを誘導し、悪化 した代償に増殖を抑制し、唾液腺機能を改善した(Morgan-Bathke et al., 2014)。オート ファジーは、基本的にすべての真核生物の組織において、本質的に活性化されている恒 常的なプロセスであり(Wirawan et al., 2012)、味覚器においても、絶え間ない味細胞 のターンオーバーの中で、新たに増殖した細胞のための新たな空間を生み出すために必 要なプロセスであると考えられる。過剰なインスリンは、正常なオートファジーと、新 たな味細胞分化からなる円滑な細胞のターンオーバーを妨げる可能性がある。すなわち、
インスリンは生理的な味細胞のターンオーバーを維持するための重要な調節因子の一 つであると考えられる。
我々の免疫染色では、IR シグナルはT1R3発現味細胞、GAD67発現味細胞で観察さ
れた(図1B、1C)。先行研究では、IRαが紡錘形の味細胞に発現し、CVではkeratin 18
と共発現していることが報告されている(Suzuki et al., 2005)。また、単離味細胞を用 いた電気生理学的実験では、インスリンによってepithelial sodium channel(ENaC)の開 口確率の制御とENaCタンパク質の膜輸送を介して、アミロライド感受性の塩味応答が 増強されたことから、ENaC を発現する塩味応答性味細胞のサブセットが IR を発現し ている可能性を示唆している(Heck GL et al., 1984、Baquero et al., 2011)。ENaCを発 現している細胞の一部はI型味細胞であるという報告もあることから、I型味細胞にも IRが発現する可能性は高い(Vandenbeuch et al., 2008)。以上のことから、味蕾におけ
29
るIR の発現は特定の味細胞のタイプに限定的なものではなく、多くの味細胞の集団に 非特異的に発現しており多様な生理機能を持つと考えられる。
IRのリガンドにはインスリンのほかにインスリン様成長因子(IGF)が知られている。
IGFは哺乳類の身体のほぼ全ての組織で発現しており、味蕾においてもこれまでにいく つかの研究でその発現に関する報告がある。ラット、マウスの味蕾では、IGFシグナル の一部が成熟味細胞のマーカーであるkeratin 18とオーバーラップしていた(Suzuki et al., 2005、Biggs et al., 2015、Zhang et al., 1995)。IGFはインスリン様成長因子受容体
(IGFR)およびインスリン様成長因子結合蛋白(IGFBP)に結合することが知られてい る。IGF1Rの発現はkeratin 8またはkeratin 18を有する味細胞で観察され、IGFBP-2お よびIGFBP-5も一部の味細胞で発現が見られた(Suzuki et al., 2005、Biggs et al., 2015、
Biggs et al., 2016)。他にも、IGFBP-6はCV味蕾を周囲の神経線維で発現することが示
されている(Suzuki et al., 2005)。舌上皮に特異的にIGF1Rを遺伝的に欠損させたマウ ス(Igf1r-KOマウス)を解析した結果、若齢Igf1r-KOマウス(30日齢)では野生型マ ウスと比べて味蕾数の減少が認められるが、80 日齢の成体マウスではこの様な変化は 観察されなかった。また、若齢および成体 Igf1r-KO マウスのいずれにおいても、味蕾 のサイズおよび味蕾内の味細胞ポピュレーション(I型、II型、III型細胞の割合)は野 生型と変わらず、舌上皮の構造および厚さにも Igf1r-KO による影響は見られなかった
(Biggs et al., 2015)。以上の知見から、IGF1は味細胞において豊富な発現が認められ るが、恐らく初期の味蕾数の決定には影響する可能性があっても、成体味蕾の構造的な ホメオスタシス維持においては、全く関係しないかもしくは非常に限定された役割しか 持たないと予想されている。
本研究では、マウス味細胞におけるIRとmTORの広範な発現を見出した。また、ex vivo味蕾オルガノイド培養系では、高濃度インスリンがI型、II型、III型、味蕾前駆/
30
幹細胞を含む味細胞の生成を抑制的に調節すること、逆に mTOR の薬理学的阻害が味 細胞の増殖を有意に促進することを発見した。本研究はインスリンが味細胞の分化・増 殖に重要な役割を果たしている可能性を示唆する初めての研究であり、また、mTORが 味蕾の恒常性維持に重要な分子である可能性を示唆した。今後は、肥満や糖尿病などの 病態発生に関わる生体の栄養代謝状態と味覚インスリン-mTOR シグナルとの関係を 明らかにしていく予定である。
31
第二章 マウス味覚器におけるヒアルロン酸の機能
緒言
D-グルクロン酸とN-アセチル-D-グルコサミンで構成される二糖の繰り返し単位を持
つグリコサミノグリカンポリマーであるHAは、皮膚、軟骨、関節液、血管、脳などの 様々な臓器の細胞外マトリックスの主要成分である(Fraser et al., 1997、Laurent et al.,
1986)。HAは、細胞膜でHA合成酵素(HAS)によって合成された後、細胞外に分泌
され、HA 分解酵素であるヒアルロニダーゼ(HYAL)によってより小さな分子量のフ ラグメントに分解される。HAは細胞間質のコラーゲンマトリックスに存在し、水和し たゲルを形成することにより、組織の水分保持や、形態保持を助けるほか、細胞周囲を 取り囲むことにより細胞の移動や接着に関与することが知られている(Day 1952)。
近年、HAは、細胞外マトリックスの構成による組織構造の保持に関わるだけでなく、
2つのHA受容体、CD44とRHAMM(receptor for hyaluronan-mediated motility)を介し て様々な細胞機能を調節する生理活性物質でもあることが判明してきた。最近の研究で は、HAが細胞の分化と増殖に重要な役割を果たしていることが示されている(Li et al.,
2007、Spicer et al., 2004)。腸管では、HAの腹腔内投与が腸管上皮の増殖を促進するこ
と、逆にCD44を阻害しHAシグナルを遮断することで腸や結腸の長さや上皮の増殖が 有意に減少することが示されている(Riehl et al., 2012)。さらに、生後早期にHAシグ ナルを阻害すると、腸管LGR5陽性前駆/幹細胞の増殖と腸陰窩での細胞分裂が有意に 減少する(Riehl et al., 2015)といった知見は、HAシグナルが正常な腸管の成長と幹細 胞の維持に必要不可欠であることを示唆している。味蕾は、腸管上皮と同じLGR5を発 現する上皮前駆/幹細胞に由来する内胚葉性上皮由来組織であるが、HAの分布や機能 はこれまでに全く調べられていない。
32
本研究では、末梢味覚器でもHAシグナルによる味蕾前駆/幹細胞の活性調節系が存 在し、味細胞の恒常的なターンオーバーに維持に寄与しているという可能性を想定して、
3 次元味蕾前駆/幹細胞培養(味蕾オルガノイド)によるin vitro 実験と、動物モデル
(マウス)によるin vivo実験を用いて、HAとHA関連分子の味覚器における発現と機 能の探索を試みた。
33
結果
マウス味覚器においてHA受容体、合成酵素、分解酵素のmRNAが発現していた
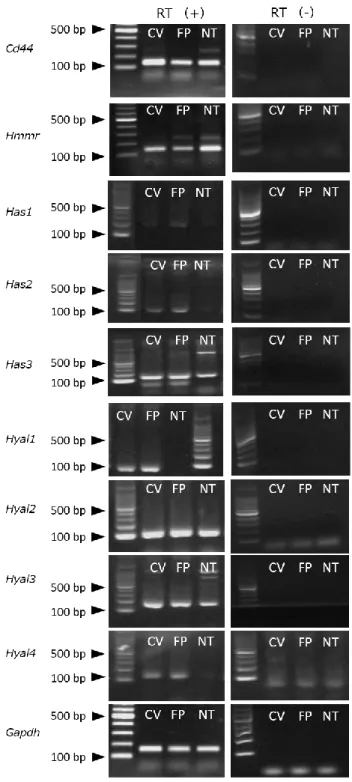
C57BL/6Jマウスの組織を用いたRT-PCR解析の結果、CV、FP、NTで、HA受容体で
あるCd44とHmmrのmRNAが発現していることがわかった(図7)。また、HA合成 酵素である Has2、Has3、および HA 分解酵素である Hyal1、Hyal2、Hyal3、Hyal4 の mRNA発現もCV、FPで認められた。しかしHas1に関しては、CV、FP、およびNTで は極めて発現が弱かった(図7)。さらに、Has2、Hyal1、Hyal4は味覚上皮特異的な発 現が確認された(図7)。
34
図7.RT-PCRによるマウス味覚器におけるHA関連分子mRNAの発現
マウスCV、FP、NTにおけるCd44、Hmmr、Has1、Has2、Has3、Hyal1、Hyal2、Hyal3、Hyal4、
およびGapdhのmRNA発現。RT+;逆転写酵素あり、RT-;逆転写酵素なし。
35
マウスCVではHA受容体、HA合成酵素、分解酵素は広範囲に発現していた
免疫組織化学染色とin situ hybridizationを用いて、マウスCVにおけるHAおよびそ の関連分子の局在を探索した。結果は、HAが味蕾、および味蕾基底部に存在するLgr5 発現味蕾前駆/幹細胞を含むCV上皮に広く分布していることが分かった(図8A、E)。
CD44はCVの味蕾内部または周囲に広範囲にわたって発現しており、その一部はPLCβ2
(II型味細胞マーカー)と共発現していた(図8B)。また、in situ hybridizationにより、
別のHA受容体であるHmmr(RHAMM)mRNA(図8C)もCVの味蕾内部または周囲 において広範囲な発現が認められた。Has2およびHas3 mRNA(図8D、E)も共にCV の味蕾内部とその周囲で発現していた。以上の結果から、HAは味蕾内の味細胞および 周囲の上皮において合成、分解され、近傍のHA受容体(CD44、RHAMM)に作用して いる可能性が示唆された。
36
図8.CVにおけるHAとHA受容体、HA合成酵素の局在
(A)CV におけるHA(緑)の免疫組織化学染色像。(B)CV におけるCD44(水色)と PLCβ2(緑)の免疫組織化学染色像。(C–F)CVにおける(C)Hmmr、(D)Has2、(E)Has3、
(F)Lgr5の in situ hybridization解析。scale bar: 25 μm。点線は味蕾の外形を示す。
37 4MUの添加は味蕾オルガノイドの増殖を抑制した
HAの味細胞への分化・増殖に対する効果を検討するために、マウス味蕾オルガノイ ドを用いた解析を行った。オルガノイドの培地中に HA 合成酵素阻害剤である 4- methylumbelliferone(以下4MU)を異なる3段階の濃度(0、100、500 μM)で添加し、
20日培養した(図9A)。20日経過時における味蕾オルガノイドの平均コロニーサイズ を計測したところ、高濃度の4MUを添加した群(500 μM)で、オルガノイドの平均コ ロニーサイズに有意な減少が見られた(0 μM:54693.8 ± 2559.0 μm2、100 μM:58055.6
± 2972.0 μm2、500 μM:11675.9 ± 591.2 μm2、**p < 0.01、one-way ANOVAおよびpost-hoc
Tukey HDS検定、図9B)。このことから、4MUによるHAの合成酵素阻害は味蕾オル
ガノイドの増殖を抑制することが示唆された。
38
図9.4MU添加による味蕾オルガノイドコロニーサイズへの影響
(A、B)味蕾オルガノイドの培地中に各濃度下の4MUを培地中に添加し培養し20日経過し た代表的なコロニー画像(A)および平均コロニーサイズ(B)。scale bar: 200 μm。 **p < 0.01、 データは平均値 ± SEMで示す。one-way ANOVAおよびpost-hoc Tukey HDS検定。
39
4MUの添加により味蕾オルガノイド中のHas2、Has3、Cd44、Entpd2、Vim、Lgr5 の mRNA発現が有意に低下した
次に、4MUを培地に添加し20日培養した味蕾オルガノイドから mRNAを抽出し、
定量 RT-PCR 解析にて味細胞マーカー、HA 関連分子の遺伝子発現解析を行った。HA
受容体のCd44、HA合成酵素であるHas2、Has3、HA分解酵素であるHyal1、Hyal3、
味細胞マーカーとして Entpd2(I 型味細胞マーカー)、Gnat3(Ⅱ型味細胞マーカー)、
Ca4(III型味細胞マーカー)、Krt8(成熟味細胞マーカー)、Lgr5(味蕾前駆/幹細胞
マーカー)、およびVim(間葉マーカー)のmRNA発現量を比較した。その結果、4MU 添加により味蕾オルガノイドのHas2、Cd44、Entpd2、Vim、およびLgr5 のmRNA発現 が有意に低下することが分かった(図10)(*p < 0.05、**p < 0.01、one-way ANOVAお
よびpost-hoc Tukey HDS検定)。一方、Has3のmRNA発現量は有意に増加していた。
以上の結果は、味蕾においてHAは成熟味細胞の前駆/幹細胞マーカーであるLGR5陽 性細胞に作用し、味細胞の分化・増殖に関与している可能性を示唆する。
40
図10.味蕾オルガノイドにおける味細胞マーカーmRNA発現に対する4MUの効果
4MUを添加し培養した味蕾オルガノイドの定量RT-PCR解析の結果。データは平均値 ± SEM で示す。それぞれの濃度群n = 5、* p < 0.05、** p < 0.01、*** p < 0.001、one-way ANOVAおよび post-hoc Tukey HDS検定。
41
マウスへの4MU投与は体重に影響せず、CVの成熟味細胞数にも有意な影響は見られ なかった
HA合成酵素阻害の影響をin vivoで検討するためにC57BL/6Jマウスを用いて異なる 4種類の濃度(体重あたり0、60、600、および1200 mg/kg)の4MUを40日間(1日1 回)経口投与した。非投与群と4MU投与群で40日間の体重変化率に有意な変化は見ら れなかった(図11A)(p > 0.05、two-way ANOVA、one-way ANOVA およびpost-hoc
Tukey HDS検定)。このことから4MUの投与はマウスの栄養摂取や全身的な健康状態
には影響しないものと考えられた。また、成熟味細胞への影響を検討するため CV の
Gustducin、CA4の免疫組織化学染色を行った(図11B)。その結果、各種濃度で4MU
を投与したマウスのCVにおいてGustducin陽性II型味細胞数と、CA4陽性III型味細 胞数には統計学的に有意な変化は認められなかった(p > 0.05、one-way ANOVA)。
42
図11.4MUを40日間投与したマウスの体重変化率とCVにおけるⅡ型、Ⅲ型味細胞数
(A)各種濃度で4MU投与開始から40日間のマウスの体重の変化率(投与前を100%、それ ぞれの濃度群n = 5、p > 0.05、two-way ANOVA、one-way ANOVA、およびpost-hoc Tukey HDS 検定)。(B)各種濃度で4MUを投与し40日経過後のマウスCVにおけるGustducin(緑)、
CA4(赤)の免疫組織化学染色像と味蕾1個あたりの各マーカー発現細胞数。各濃度n = 5。p >
0.05、one-way ANOVA。scale bar: 25 μm。点線は味蕾の外形を示す。
43
マウスへの4MU投与により味蕾の前駆/幹細胞、成熟味細胞の分化・増殖に影響し た
4MUを40日間(1日1回)経口投与した後、CVを採取しRNAを回収し、定量RT- PCR解析を用いて、HA受容体のCd44、HA合成酵素であるHas2、Has3、HA分解酵素
であるHyal1、Hyal2、Hyal3、各種味細胞マーカーのmRNA発現量の比較を行った(図
12)。その結果、4MU 非投与群と比較し、高濃度投与群では Entpd2、Gustducin、Ca4、
Lgr5、VimのmRNA発現が有意に減少していることが明らかになった(*p < 0.05、**p
< 0.01、one-way ANOVAおよびpost-hoc Tukey HDS検定)。味蕾オルガノイドの結果(図
10)と同様に、HA の合成酵素阻害により Lgr5 および成熟味細胞マーカー(Entpd2、
Gustducin、Ca4)の mRNA 発現が有意に抑制されたことから、生体においても HA が
LGR5陽性の味蕾前駆/幹細胞の活性や、成熟味細胞への分化・増殖にも影響すること が示唆された。
44
図12.マウスCVにおける各種マーカーmRNA発現に対する4MUの影響
4MUを各濃度で40日間投与したマウスCV組織より採取したRNAを用いた定量RT-PCR解 析。データは平均値 ± SEMで示す。それぞれの濃度群n = 6、* p < 0.05、** p < 0.01、*** p <
0.001、one-way ANOVAおよびpost-hoc Tukey HDS検定。
45
マウスへの4MU投与によりCV周囲の細胞増殖が劇的に抑制された
4MUを40日間(1日1回)経口投与したマウスのCVにおける細胞増殖の変化を比 較するため、Mki67抗体を用いた免疫組織化学染色を行った(図13)。4MU非投与マウ スでは CV の味蕾基底部を含む舌上皮基底層に限局して Mki67 陽性細胞が発現してい ることが分かった。一方、4MU投与マウスでは、Mki67陽性細胞が特に味蕾が存在する 部位で減少している様子が観察された(図13A)。このことから、4MU投与マウスCV の味蕾内部と味蕾に接している Mki67 陽性細胞数の定量化を行ったところ、有意な減 少が認められた(図13B、p < 0.05、unpaired t-test)。この結果は、HAの合成酵素を阻害 することにより、味蕾基底部に存在するLGR5陽性味蕾前駆/幹細胞を含むCV周囲の 細胞の細胞増殖が劇的に抑えられたことを示唆する。
46
図13.マウスCVにおけるMki67陽性細胞への4MUの影響
(A、B)0および600 mg/kg 体重の4MUを40日間投与したマウスのCVでのMki67(赤)
の代表的な免疫組織化学染色像、および味蕾内と味蕾に接する Mki67 陽性細胞数。それぞれの 濃度群n = 3。scale bar: 25 μm。データは平均 ± SEMで示す。unpaired t-test *** p < 0.001。
47
考察
本研究では、はじめにマウスCVにおいて味蕾内、味蕾周囲組織でHA受容体Cd44、
Hmmr mRNA の発現を認めた。さらに、HA 合成酵素である Has2 や Has3と分解酵素の
Hyal1–4 の mRNA の発現を見出した(図 1、2)。免疫組織化学染色および in situ
hybridizationでは、Cd44、Hmmrに加え、HAも CV領域の味細胞を含む周辺の上皮に広
範囲に発現していることがわかった。CVにおけるCD44 発現に関する結果は、過去の 報告と一致し、LGR5 発現味蕾前駆/幹細胞と近接した位置で見られた(Aihara et al.,
2015)。このことから、味蕾を含む舌上皮細胞でHAが合成・代謝され、味蕾と味蕾周
囲上皮に発現する HA 受容体を活性化することで味細胞および味蕾周囲の上皮の機能 に関与している可能性が示唆された。
HAは、細胞表面に存在するHA受容体であるCD44とRHAMMに結合する。CD44 は全身に広く存在する主要なHA受容体として知られ、4つのドメインからなる1回膜 貫通型糖蛋白質で、細胞周囲のマトリックス保持、細胞輸送や細胞凝集などのマトリッ クス-細胞間のシグナル伝達において多様な役割を果たす(Jiang et al., 2007、Laurent et al.,1996、Sherman et al.,1994、Aruffo et al.,1990)。また、CD44は様々な組織の幹細胞に 発現して分化・増殖にも深く関与している。例えば、海馬の神経幹細胞の分化に重要な 働きをしていることが報告されており、Cd44 遺伝子欠損により記憶障害が起こると報 告されている(Su et al., 2017)。他にもES細胞(embryonic stem cells)からの平滑筋分 化に必要な分子であること(Simpson et al., 2016)、さらに、癌幹細胞のマーカー分子の 1つであることも報告されている(Dalerba et al., 2007、Thapa et al., 2016)。これらのこ とから、CD44は味細胞の前駆/幹細胞においても成熟味細胞への分化・増殖の鍵分子 として機能している可能性が示唆されている。一方、RHAMMは移動中の細胞に多く発 現する HA と高い結合能を持つ糖タンパク質であり細胞内と細胞表面で多様な形態が
48
見られる分子である(Zhang et al., 1998、Crainie et al., 1999、Nedvetzki et al., 2004)。こ の2つの受容体CD44とRHAMMは、細胞増殖、分化、細胞移動を含むHAシグナル 伝達において協調して働くことが示唆されている。例えば、関節炎モデルマウスにおい てCd44のノックアウトをした場合、RHAMMがCD44の喪失を補償し、細胞移動のサ ポート、炎症に関与する遺伝子のアップレギュレートが起こる(Nedvetzki et al., 2004)。
我々の実験でもこの2 つの受容体の広範な発現が味蕾で確認されたことから(図 8)、
それぞれの受容体がそれぞれ特異的な役割を有して協調的に働いている可能性がある。
HA合成酵素であるHAS1、2は高分子HA(HMW-HA:2 × 105–2 × 106 Da)を産生し、
低分子HA(LMW-HA:1 × 105–2 × 106 Da)の産生にはHAS3およびHYALが関与する とされる(Jiang et al., 2007)。興味深いことに、HAはその長さにより機能が多様であ ることが報告されている(Chistyakov et al., 2005)。具体的には、低分子量HAは細胞接 着を促進するが、高分子HAは逆に細胞分離効果を示す(Tien et al., 2005)。また、Has1 またはHas3遺伝子欠損マウスは生存可能であるが、Has2遺伝子欠損マウスは致死であ る(Spicer et al., 1999)。加えて、HASは特定の組織では、部位特異的または時間特異 的な発現パターンを示すことが示されている。例えば、マウスの歯の発生過程でHasの 遺伝子発現を調べた研究では、Has1は発現していないが、Has2 は胎生 12~18 日で歯 胚周囲の間葉に発現しており、出生後にその発現は消失した。さらにHas3は胎生16日 までは、エナメル器で特異的な発現が見られることがわかっている(Morita et al., 2016)。
本研究でも、歯胚同様、味覚上皮でHas1のmRNAの発現は弱かったが(図7)、Has2 とHas3に関しては両遺伝子ともに味蕾において発現が見られた(図8D、E)。マウス の味細胞は胎生期終盤(E15前後)に発生し、成獣のような味蕾の構造が完成するのは 出生後1週間程度経ってからである(Barlow, 2015)。今回着目したターンオーバーに 寄与する幹細胞への影響という観点ではなく、味蕾の発生におけるHAシグナルの関与 は、今後の検討課題である。