DOI: http://dx.doi.org/10.14947/psychono.34.3
ラットのピーク法を用いた計時行動のセッション間安定性の検討
氏 田 麻 美
a・服 部 稔
b・坂 田 省 吾
a,*
a 広島大学大学院 総合科学研究科
b 広島大学大学院 医歯薬保健学研究院
Behavioral stability of timing in a peak-interval procedure in rats
Asami Ujita
a, Minoru Hattori
b, and Shogo Sakata
a,*
a Graduate School of Integrated Arts and Sciences, Hiroshima University, b Institute of Biomedical and Health Sciences, Hiroshima University
The peak-interval procedure is an effective tool for measuring interval-timing behavior in rats. However, its stability has not been previously examined. Therefore, the purpose of this study was to examine behavioral stability across sessions when rats were trained using the peak-interval procedure. To this end, we performed 100 sessions of the peak-interval 30-s schedule with rats. Responses during 90-s probe trials were classified into 3-s bins individual-ly and were averaged across five session-blocks in order to examine behavioral stability between sessions. Data anal-ysis revealed the stability with a relative response rate for one block in five sessions. Furthermore, we applied the provided relative response rate to a normal distribution curve and calculated the kurtosis of the curve at each re-sponse rate and Gaussian fitting, i.e., the peak time of the curve. Examination of the stability showed a Cronbach’s coefficient of α≧0.86 (R2=0.98, peak time=0.93, kurtosis=0.86). This indicates that the peak-interval procedure
is an excellent reliable tool for measuring the interval-timing behavior in rats.
Keywords: interval-timing, peak-procedure, stability, rats
序 論 インターバルタイミングとよばれる秒・分単位の時間 によって制御される計時行動は,行動の生起,ある行動 の持続時間の推定,あるいは生きていくために重要な採 行動など,日常の行動と関連する部分が多い(Buhusi & Meck, 2005)。したがって,生体が計時行動をどのよ うな分子動態メカニズムで処理しているかを明らかにす ることは,生体の行動を理解することにつながると考え られる。 動物のインターバルタイミングを検討するために,多 く の計 時 行 動 を 測 定 す る 課 題 が 考 案 さ れ て い る。 Church (1978)は動物の計時行動を測定する方法を,評 価(estimate)法と産出(production)法の2つのタイプ に分類した。評価法は標準刺激を提示した後,プローブ 刺激を提示し,プローブ刺激の提示時間が標準刺激と同 じであったかどうかを判断するような方法である。この 方法は過去に経験した時間と,今目の前にある時間を比 較して弁別する課題であり,計時メカニズムを検討でき る可能性がある。それに対して産出法は,実験者によっ て示された時間を被験体が何らかの方法で再生する方法 で,流れている時間に対して心理的な期待を行動に反映 させることができる。産出法の課題例としては,試行開 始を合図する刺激の提示から一定間隔後の最初の反応が 強化され,試行と試行は試行間間隔(inter-trial interval: ITI)でそれぞれ分離している離散試行型の固定間隔 (discrete fixed interval: DT-FI)強化スケジュールがあげ
られる。 近年ではDT-FI強化スケジュール法の変形であるピー ク法(peak-interval procedure)がインターバルタイミング を検討するうえでよく用いられている(Roberts, 1981)。 ピーク法は,強化試行である FI試行と,非強化試行で あるプローブ試行がランダムに提示される課題である。 FI試行では,刺激が提示されてから一定時間経過後の初 Copyright 2015. The Japanese Psychonomic Society. All rights reserved. * Corresponding author. 1–7–1 Kagamiyama,
Higashi-Hiroshima, Hiroshima 739–8521, Japan. E-mail: ssakata@ hiroshima-u.ac.jp
発反応が強化され,試行と試行はITIによって区切られ る。しかし FI試行のみでは,強化子が与えられた時点 でFI 試行が終了してしまうため,実際に計時行動を 行っているかどうかは不明である。そこでプローブ試行 を課題の中に組み込むことで,試行中の反応分布を明ら かにすることが可能となる。プローブ試行は刺激の提示 により試行が開始されるが,反応しても強化されない試 行であり,強化時間が過ぎるまではその試行が FI試行 かプローブ試行かは被験体にとって不明であるため,FI 強化スケジュールの設定値付近の反応が多くなると考え られる。 一般に,時間を横軸,プローブ試行の反応頻度を縦軸 として度数分布を求めると,FI強化スケジュールの設定 値付近に反応率の頂点をもつ正規分布様の反応分布が得 られる。多くの研究では,正規分布から回帰曲線をもと め,回帰曲線の頂点を示す時間をピーク時間と定義し, 計時行動の指標としている。またその反応度数分布の散 布度を指標として,尖度という観点から計時行動をみる ことができる。つまり,分布が狭く尖度が小さいほど, 正確に時間を捉えているということができる。ピーク時 間は,ヒトもヒト以外の動物も FI強化スケジュールの 設定値あたりにみられるが,反応分布の散布度で表され る尖度は,ヒトとヒト以外の動物で違いがみられること が報告されている(Rakitin et al., 1998)。 ピーク法の妥当性は多くの研究で示されている。時間 の評価値の標準偏差が計時される時間間隔に比例するス カラー特性は動物種を超えて成立することが報告されて いる。これは時間知覚における最も代表的な特徴である。 他のインターバルタイミングの課題と同様に,ピーク法 で得られたラットの相対反応率も,スカラー特性として 表現できることが報告されている(Gibbon, 1977)。また ドーパミン系の機能不全と関連するパーキンソン病患者 は計時行動の困難が報告されている(Malapani, Deweer, & Gibbon, 2002)が,ピーク法で訓練された動物にドーパ ミン作用薬(methamphetamineもしくはd-amphetamine) を投与するとピーク時間が前進し,ドーパミン拮抗薬 (haloperidol)を投与するとピーク時間が後退すること が報告されている(Buhusi & Meck, 2002; Kraemer, Ran-dall, Dose, & Brown, 1997; Maricq, Roberts, & Church, 1981; Meck, 1996)。またラットのピーク法遂行中にドーパミ ン作動性経路である線条体から脳波の一種であるθ波を 測 定 し た 先 行 研 究 で は, 試 行 開 始 か ら FI 強 化 ス ケ ジュールの設定値に近づくにつれてレバー押しの反応回 数が増加し,それにともない線条体から測定したθ波の パワー値も増加したことが報告されている(Hattori & Sakata, 2014)。これらの結果は,ヒトと動物の計時に関 する神経基盤が共通していることを示している。 目 的 以上のことから多くの先行研究の結果は,ピーク法に よって得られるピーク時間が動物のインターバルタイミ ングの有効な指標になることを示唆している。しかし ピーク法の学習基準や解析基準は先行研究によって一貫 しておらず(Buhusi & Meck, 2000; Roberts, 1981; Swanton, Gooch, & Matell, 2009),ピーク法の指標であるピーク時 間や尖度,ピーク曲線の適合率がセッションを繰り返す 中でどの程度変動するかを検討した研究は少ない。そこ
で本研究ではラットのピーク法を100セッション連続で
実施し,その安定性を検討する。
また,Matell & Portugal (2007)はピーク法における ラットの相対反応率に対して正規分布の回帰曲線の適合
率 R2≧0.90を5日間連続で超えることを学習基準とし
た。MacDonald & Meck (2005)は正規分布の回帰曲線の 適 合 率 R2≧0.80 を,西村・高橋・坂田(2003)は R2≧ 0.80,回帰曲線の尖度が>1.3のセッションを解析対象と した。しかし西村他(2003)は任意の基準でデータを取 捨選択しており,それらの基準が適切か検討は行ってい ない。そこで本実験ではさらに学習基準として正規分布 の適合率R2はどの程度が適切か,探索的な検討を行っ た。 方 法 被験体 被験体は,FI30秒のピーク法を35セッション経験し たオスのWistar系アルビノラット6匹を用いた。本実験 開始時の日齢は470日であった。実験終了後時の日齢は 576日であった。すべての被験体は最初のピーク法訓練 から60日間を経過していた。実験期間中ラットはアク リル製の透明なケージ(奥行き 400×幅250×高さ200 mm)で個別に飼育し,ラットの体重は自由摂食時のお よそ85%に維持した(実験時体重約395 g)。飼育室は明 期12時間(08 : 00–20 : 00),暗期12時間(20 : 00–08 : 00) の明暗サイクル下に置かれ,実験はすべて明期に行っ た。水は自由摂取とし,給 は実験終了後に行った。な お本実験は広島大学の動物実験委員会の承認を受けて 行った。 装置 6台のアクリル製のオペラントボックス(260×300× 250 mm)を用いた。側面と天井の壁は透明塩化ビニー ル製,床はステンレス・グリッドであった。前面と後面
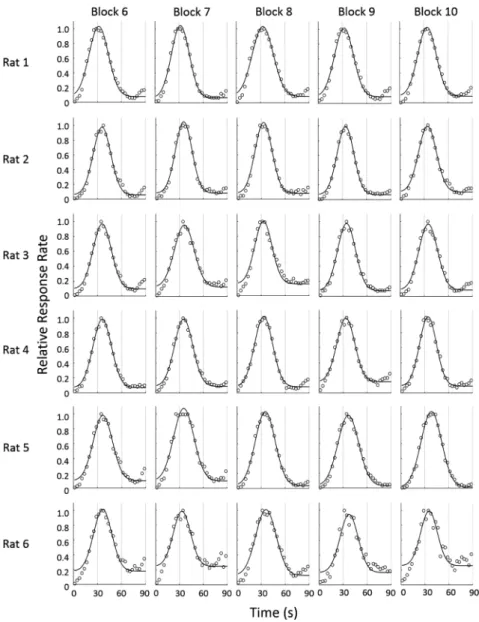
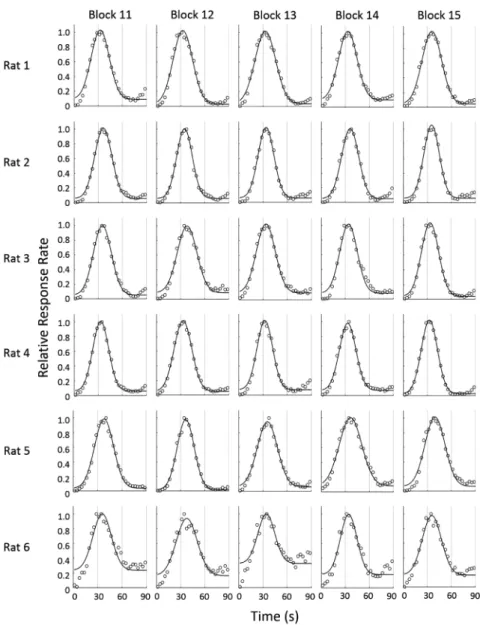
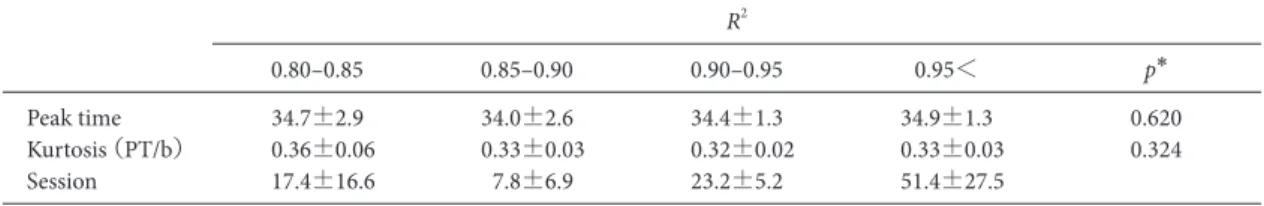
の壁はアルミニウム製で,前面の壁には格納式のレバー を設置した。強化される反応が行われた時には,前面の 壁の中央に取り付けられた 皿に45 mgのペレットが給 されると同時に,実験箱外のブザー(75 dB, EB2116, 松下電工製)が鳴るように設計した。実験箱の天井に設 置したライト(6.3 V, 0.15 A)をハウスライトとして利 用した。音刺激(90 dB, 2000 Hz, 125 ms on/offのパルス 音)は背面の壁に設置したスピーカーから提示した。オ ペラントボックスは,外部の音や光の影響を低減させる ために木製の防音箱(450×620×460 mm)内に設置し た。実験の制御・記録と音の提示は,2台のパーソナル コンピュータ(それぞれ Aptiva 2168-S65, Endeavor MT-6000)によって行った。 手続き ピーク法を 1日1セッション行った。1セッションは 130試行で,最初の10試行は試行開始30 s経過後の初発 反応を強化する FI試行を提示し,残りは3 : 1 (90 : 30) の割合でFI試行とプローブ試行をランダムに提示した。 FI試行では試行開始から30 s経過後の最初の反応が強化 されると同時に試行が終了した。プローブ試行では試行 開始から30 s経過後の反応に対しても強化子は与えられ ず,反応とは無関係に 90 s後に終了した。ITIは35±3 s であった。プローブ試行が4試行以上連続で提示されな いように擬似ランダムに試行の順序を決定した。この手 続きを100セッション実施した。 行動解析 まず安定性の検討としてプローブ試行のみを取り出 し,試行開始から90 s間のデータを用い,試行開始から 3 sごとの反応度数を求めた。次に最大反応度数が1にな るように相対化し,相対反応度数を求め,5セッション 1ブロックとして平均を算出した。ラットの相対反応度 数分布に対して,市販のソフトウェアを用い(MATLAB 2014, MathWorks),先行研究(西村他,2003; 西村・坂 田,2004)による下式を用いて正規分布からの回帰曲線 を求め,ピーク時間,尖度,回帰分析の適合率を算出し た。 y=y0+a×exp[(−0.5((x−x0)/b)2)] ここでxは試行開始からの経過時間,yは相対反応率, x0は回帰曲線の頂点,bは回帰曲線の標準偏差,y0は回 帰曲線の切片を表している。この回帰曲線によって算出 されたx0を頂点時間,標準偏差bを頂点時間x0で割った もの(b/x0)を尖度,回帰曲線の決定係数(R2)を適合 率と定義した。次に学習基準の検討のため,1セッショ ンずつの解析を実施した。解析はブロック時の方法と同 様に,プローブ試行の反応度数分布に関して,回帰曲線 を求め,ピーク時間,尖度,回帰分析の適合率を算出し た。 統計解析 セッションブロック間の内的一貫性を検討するために Cronbach’s αを用いて検討を行った。またセッションブ ロックを前半・後半の2つに分類し,適合率R2・ピーク 時間・尖度(b/x0)を t検定によって比較検討した。効 果量としてCohen’s dを算出した。次に学習基準の解析 として,被験体ごとに100セッションを回帰分析の適合 率R2より0.80–0.85, 0.85–0.90, 0.90–0.95, 0.95<の4区間に 分類した。4区間でピーク時間・尖度(b/x0)にどのよ うな違いがあるか,対応のある分散分析を用いて解析し た。多重比較にはBonferroniの補正を行ったt検定を用 いた。すべての指標は平均値±標準偏差で示した。統計 分析はSPSS (IBM SPSS statistics 22.0, IBM Corporation, NY, USA)を用いて行った。すべての統計的検定における有 意水準は5%とした。 結 果 Figure 1 に 1–5 ブ ロ ッ ク,Figure 2 に 6–10 ブ ロ ッ ク, Figure 3 に 11–15 ブロック,Figure 4 に 16–20 ブロックの ピークの形成過程を示した。すべてのラットにおいて第 4ブロックで正規分布様の相対反応率が得られた。適合 率 R2,ピーク時間,尖度(b/x 0)を Figure 5 に示した。 すべての指標において初期の3ブロックと比較して第4 ブロック以降で分散が小さくなっていた。したがって, 最初の3ブロックは学習形成途中だと考えられるため, 以降の分析では4–20ブロックを解析対象とした。3つの 指標それぞれについて4–20ブロックの平均を算出した 結果,適合率R2は0.97±0.02,ピーク時間は34.5±1.8 s, 尖度(b/x0)は 0.32±0.03であった。本研究においても FI強化スケジュールの設定値付近に反応の頂点をもち, それを中心に反応が増加・減少するという,ピーク法に おける典型的な正規分布様の反応パターンを示した。こ のことからラットは強化設定時間を計時していたことが 示唆された。Cronbach’s αは適合率R2で0.98,ピーク時 間で 0.93,尖度(b/x0)で 0.86と高い値を示した。また 4–10ブロックを前半,11–20を後半に分類し再度分析し た結 果, 適 合 率 R2 (前 半: 0.97±0.03, 後 半: 0.97± 0.03),ピーク時間(前半: 34.5±1.4 s,後半: 34.5±1.3 s), 尖度(b/x0)(前半: 0.33±0.03,後半: 0.32±0.02)いず れも有意な差は認められなかった(適合率 R2: t(5)= 2.37, p=0.064, Cohen’s d=0.12, ピーク時間t(5)=0.06, p= 0.952, Cohen’s d=0.01, 尖度: t(5)=1.79, p=0.133, Cohen’s
d=0.39)。 次にセッションごとの解析を行い適合率とその他の指 標にどのような関連があるか検討を行った。セッション を,適合率R2より0.80–0.85, 0.85–0.90, 0.90–0.95, 0.95<の 4区間に分類し,それぞれの区間内におけるピーク時間 と尖度の平均値と標準偏差を算出した(Table 1)。1匹は 0.85–0.90の基準値に当てはまるセッションが存在しな かったため,以降の統計解析から除外した。したがって 以降の解析では5匹を解析対象とした。ピーク時間はそ れ ぞ れ,34.7±2.9 s, 34.0±2.6 s, 34.4±1.3 s, 34.9±1.3 s で あった。また尖度(b/x0)はそれぞれ 0.36±0.06, 0.33± 0.03, 0.32±0.02, 0.33±0.03 であった。統計解析の結果 4 つの適合率R2によるピーク時間に有意な差は認められ なかった(F(3, 12)=0.48, p=0.620)。尖度(b/x0)にお いても有意な差は認められなかった(F(3, 12)=1.28, p= 0.324)。 考 察 インターバルタイミングを測定する課題では,長潜時 分化強化スケジュール(differential reinforcement of long Figure 1. Response rate of a probe-trial in ten successive blocks (1–5). The mean proportion of the maximum response rate
latencies: DRLL), 間 隔 二 等 分 課 題(temporal bisection task),ピーク法(peak-interval procedure)などが用いら れている。この中でもピーク法は時間経過における心理 的予期を行動(相対反応率)に反映させることが可能な ため,方法論的に我々の日常の計時行動を最も反映して いる方法である(坂田,2003)。そのため多くの研究で ピーク法が用いられているが,今まで安定性についての 検討は不十分であった。そこで本研究ではラットのピー ク法における安定性を検討するために,ピーク法を100 セッション実施しその適合率・ピーク時間・尖度の変動 性を検討した。 先行研究と同様に本研究においても正規分布様の反応 パターンのタイミング行動が第4ブロック以降すべての ラットで観察できた。本研究では以前にピーク法を経験 したことのある被験体を用いたため,ピーク曲線の形成 が早かった可能性が考えられる。またピーク形成後の 17 ブロックにおける Cronbach’s αは,適合率 R2が0.98, ピーク時間が 0.93,尖度(b/x0)が 0.86 といずれも高 い値を示した。また前半(4–10ブロック)と後半(11– 20 ブ ロ ッ ク) に も 有 意 な 差 は 認 め ら れ な か っ た。 Figure 2. Response rate of a probe-trial in ten successive blocks (6–10). The mean proportion of the maximum response rate
Cronbach’s αが高値を示したこと,前半と後半によって 適合率・ピーク時間・尖度に有意な差が認められないこ とから,ピーク法はセッションブロック間の変動性が小 さく,安定性の高い課題であることが示唆された。 次にセッションごとの解析を行い,適合率 R2より 0.80–0.85, 0.85–0.90, 0.90–0.95, 0.95<の 4 区間に分類し, ピーク時間を比較した。その結果,いずれの群も34 s付 近にピーク時間が見られ,群間に有意な差は認められな かった。これは尖度についても同様であり,4群間で有 意な差は認められなかった。適合率R2>0.80はピーク時 間において有意な差が認められなかったため,適合率に ついてはR2>0.80に到達するまでトレーニングを行うの が1
つの基準であると考えられる。したがって,Mac-Donald & Meck (2005)や西村ら(2003)の正規分布の回
帰曲線の適合率R2>0.80を基準とすることは妥当である と思われる。また,ブロック間の解析において,前半 (4–10ブロック)と後半(11–20ブロック)で有意な差 は認められなかったものの,後半の適合率は前半よりも わずかに高く,さらに適合率R2>0.90になると,ばらつ きも小さくなることが示唆された(Table 1)。よって, Figure 3. Response rate of a probe-trial in ten successive blocks (11–15). The mean proportion of the maximum response
ばらつきが小さい状態において,より厳密にピーク時間 の違いなどを検出したい場合はMatell & Portugal (2007)
のように適合率R2>0.90を基準とするのがよいと思われ る。 本実験ではピーク法の経験のあるラットを被験体とし て用い,100セッションという非常に長期間のトレーニ ングを実施した。したがって実験開始時は470日齢,実 験終了後は576日齢と高齢であった。ヒトやラットを対 象とした実験では,加齢によって計時行動が変化するこ とが明らかになっている。ラットを対象とした実験にお いて,Meck (2006b)は180日齢(6か月齢)と780日齢 (26か月齢)のラットを用い,強化設定時間を20秒とし てピーク法を行ったところ,高齢のラットにおいてピー ク時間が約4 s遅くなることを報告した。本研究におい て,ピーク法の強化設定時間は30 sであったが,結果と してピーク時間は34 s付近であった。この原因について は加齢の影響を受けている可能性がある。今後は幼齢か ら高齢まで縦断的な実験を行い,インターバルタイミン グと加齢の関連について検討する必要がある。 多くの先行研究と同様に,おおよそ強化設定時間に反 Figure 4. Response rate of a probe-trial in ten successive blocks (16–20). The mean proportion of the maximum response
応の頂点をもち,それを中心に反応が増加・減少すると いう,正規分布様の反応パターンのタイミング行動を示 した。このことは,先行研究と同様にラットにおいて も,ヒトと同様にピーク法によって形成されるタイミン グ行動に内在する脳内神経基盤が共通していることを示 唆するものである。動物のタイミング行動を説明するモ デルは多く存在するが,現在のところはまだ,Gibbon,
Church, & Meck (1984)の情報処理モデルが有効である と考えられている。
情報処理モデル(Figure 6 (Gibbon et al., 1984を改変)) では,タイミング行動は大きく分けて時計部,記憶部, 決定部の3つの段階からなるシステムであると仮定され ている。時計部ではパルスを発生させるペースメーカー を想定しており,ペースメーカーから発信された信号が ゲートを通過し,蓄積される。次に記憶部では,時計部 で蓄積されたパルスの数を作業記憶に送っている。作業 記憶は常に変化しており,強化初期の時点では参照記憶 はまだ形成されていないが,行動が強化されるにつれて 参照記憶が形成されていくようになる。記憶部では,こ の変化し続ける作業記憶と,強化される参照記憶の比較 を行い,最後の決定部において行動するかどうかを決定 すると想定している。 情報処理モデルは時間弁別学習における行動特性から 考えられたモデルであり,先行研究によって前頭葉・海 馬・大脳基底核を構成する黒質・線条体などが重要な役 割を果たしていると考えられている。(Buhusi & Meck, 2005; MacDonald, Fortin, Sakata, & Meck, 2014; Meck, 1996)。今までの先行研究では特に黒質・線条体の検討 が多く行われており,Meck (2006a)は,黒質からのドー パミン投射を阻害すると,ピーク法において課題成績が 悪くなり,ピーク曲線が形成されなくなることを報告し ている。また,線条体背側部のドーパミン作動性システ ムが課題を行う上で重要であることも報告されている Figure 5. The group mean value of each twenty-block group (and ±S.D., N=6). Left: The coefficient of determination of
Gaussian fitting (R2), Middle: Peak time, Right: Kurtosis.
Table 1.
The mean peak time and Kurtosis (and ±S.D.) of each coefficient of determination of Gaussian fitting (R2) (N=5).
R2 0.80–0.85 0.85–0.90 0.90–0.95 0.95< p* Peak time 34.7±2.9 34.0±2.6 34.4±1.3 34.9±1.3 0.620 Kurtosis (PT/b) 0.36±0.06 0.33±0.03 0.32±0.02 0.33±0.03 0.324 Session 17.4±16.6 7.8±6.9 23.2±5.2 51.4±27.5 *対応のある分散分析
Figure 6. Information processing model for the time perception (Gibbon, Church, & Meck, 1984, partial modification). The upper part shows a clock process, the middle part shows a memory process, and the low-er part shows decision process.
(Cheng, MacDonald, & Meck, 2006; Matell, Bateson, & Meck, 2006; Meck, 1996)。上記のような先行研究によって,黒 質–線条体系のドーパミン系の働きがタイミング行動に 影響を与える可能性が高いと考えられる。しかし情報処 理モデルにおいて初期段階である時計部との関連につい てはいまだ不明瞭である。本研究によってピーク法は計 時行動を測定する指標として安定性が高いことが示唆さ れたため,今後は神経科学的手法などを取り入れて検討 を行うことで,計時行動の分子動態メカニズムを説明す る神経基盤について言及することが可能になると考えら れる。 最後に,本実験では1セッションが130試行で構成さ れているピーク法を100セッション行い,セッションの 進行に伴った各指標の変化を検討した。しかしセッショ ン内における各指標の変化については未検討であり,今 回の結果をふまえた上で今後の課題として検討を行う必 要があると考えられる。 結 論 本実験ではインターバルタイミングの課題であるピー ク法を100セッション連続で実施し,ピーク法の安定性 を検討した。その結果,ピーク法はセッションブロック 間の変動性が小さく,安定性の高い課題であることが明 らかになった。また適合率R2>0.80が1つの学習基準と なることが示唆された。今後はピーク法を用いて計時行 動の分子動態メカニズムを検討していくことで,生体の 計時行動について,より言及することが可能になると考 えられる。 謝 辞 本論文を作成するにあたり,貴重な助言をいただいた 玉川大学脳科学研究所酒井裕先生に深謝いたします。本 研究は広島大学大学院総合科学研究科総合科学推進プロ ジェクト,広島大学特別経費概算要求(脳科学研究を基 盤とした発達障害の行動科学研究プロジェクト)及び日 本学術振興会科学研究費補助金(16530471, 16203038, 21300125, 25280051, 26119519, 26780419)の補助を受けた。 記して感謝の意を表する。 引用文献
Buhusi, C. V., & Meck, W. H. (2000). Timing for the absence of a stimulus: the gap paradigm reversed. Journal of Experi-mental Psychology: Animal Behavior Processes, 26, 305–322. Buhusi, C. V., & Meck, W. H. (2002). Differential effects of
methamphetamine and haloperidol on the control of an in-ternal clock. Behavioral Neuroscience, 116, 291–297.
Buhusi, C. V., & Meck, W. H. (2005). What makes us tick? Functional and neural mechanisms of interval timing. Na-ture Reviews. Neuroscience, 6, 755–765.
Cheng, R. K., MacDonald, C. J., & Meck, W. H. (2006). Differ-ential effects of cocaine and ketamine on time estimation: implications for neurobiological models of interval timing. Pharmacology, Biochemistry, and Behavior, 85, 114–122. Church, R. M. (1978). The internal clock. In S. H. Hulse,
H. Fowler & W. K. Honig (Eds.),Cognitive Processes in Animal Behavior. Hillsdale, NJ: Erlbaum, pp. 227–310. Gibbon, J. (1977). Scalar expectancy theory and Weber’s law
in animal timing. Psychological Review, 84, 279–325. Gibbon, J., Church, R. M., & Meck, W. H. (1984). Scalar
tim-ing in memory. Annals of the New York Academy of Sciences,
423, 52–77.
Hattori, M., & Sakata, S. (2014). Brain electrophysiological ac-tivity correlates with temporal processing in rats. Behav-ioural processes, 101, 97–102.
Kraemer, P. J., Randall, C. K., Dose, J. M., & Brown, R. W. (1997). Impact of d-amphetamine on temporal estimation in pigeons tested with a production procedure. Pharmacolo-gy, Biochemistry, and Behavior, 58, 323–327.
MacDonald, C. J., Fortin, N. J., Sakata, S., & Meck, W. H. (2014). Retrospective and prospective views on the role of the hippocampus in interval timing and memory for elapsed time. Timing and Time Perception, 2, 51–61. MacDonald, C. J., & Meck, W. H. (2005). Differential effects of
clozapine and haloperidol on interval timing in the supra-seconds range. Psychopharmacology, 182, 232–244.
Malapani, C., Deweer, B., & Gibbon, J. (2002). Separating storage from retrieval dysfunction of temporal memory in Parkinson’s disease. Journal of Cognitive Neuroscience, 14, 311–322.
Maricq, A. V., Roberts, S., & Church, R. M. (1981). Metham-phetamine and time estimation. Journal of Experimental Psychology: Animal Behavior Processes, 7, 18–30.
Matell, M. S., Bateson, M., & Meck, W. H. (2006). Single-trials analyses demonstrate that increases in clock speed contrib-ute to the methamphetamine-induced horizontal shifts in peak-interval timing functions. Psychopharmacology, 188, 201–212.
Matell, M. S., & Portugal, G. S. (2007). Impulsive responding on the peak-interval procedure. Behavioural Processes, 74, 198–208.
Meck, W. H. (1996). Neuropharmacology of timing and time perception. Cognitive Brain Research, 3, 227–242.
Meck, W. H. (2006a). Neuroanatomical localization of an in-ternal clock: a functional link between mesolimbic, nigros-triatal, and mesocortical dopaminergic systems. Brain Re-search, 1109, 93–107.
Meck, W. H. (2006b). Temporal memory in mature and aged rats is sensitive to choline acetyltransferase inhibition. Brain Research, 1108, 168–175.
西村優基・坂田省吾(2004).ラットのピーク法におけ るタイミング行動の検討―セルフスタートレバーと ギャップ手続きの効果― 広島大学総合科学部紀要
IV理系編,30, 1–18.
(Nishimura, Y., & Sakata, S. (2004). Timing behavior on peak procedure in rats: effect of self-start lever and gap procedure. Memoirs of the Faculty of Integrated Arts and Sciences, Hiroshima University, IV, Science Reports, 30, 1–18.)
西村優基・高橋絵理佳・坂田省吾 (2003).ピーク法に よるタイミング行動の形成と時間的分布 広島大学総 合科学部紀要IV理系編,29, 63–75.
(Nishimura, Y., Takahashi, E., & Sakata, S. (2003). Shaping of interval timing behavior by the peak procedure and re-sponse temporal distribution. Memoirs of the Faculty of Inte-grated Arts and Sciences, Hiroshima University, IV, Science Reports, 29, 63–75.)
Rakitin, B. C., Gibbon, J., Penney, T. B., Malapani, C., Hinton, S. C., & Meck, W. H. (1998). Scalar expectancy theory and
peak-interval timing in humans. Journal of Experimental Psychology: Animal Behavior Processes, 24, 15–33.
Roberts, S. (1981). Isolation of an internal clock. Journal of Ex-perimental Psychology: Animal Behavior Processes, 7, 242– 268.
坂田省吾(2003).時間弁別の行動特性と脳の情報処理 モデル 生理心理学と精神生理学,21, 39–48. (Sakata, S. (2003). Behavioral property of temporal
discrim-ination and the information-processing model of the brain. Japanese Journal of Physiological Psychology and Psychophys-iology, 2, 39–48.)
Swanton, D. N., Gooch, C. M., & Matell, M. S. (2009). Averag-ing of temporal memories by rats. Journal of Experimental Psychology: Animal Behavior Processes, 35, 434–439.