43
日本甲殻類学会 ReportCarcinological Society of Japan
報告
Cancer 26: 43–46 (2017)
中部北太平洋の光孝海山で漁獲されたセンジュエビ類の
DNA種判別
Molecular species identification of polychelid species collected at Koko Seamount in the central
North Pacific
柳本 卓
1・小西光一
2Takashi Yanagimoto and Kooichi Konishi
はじめに 中部北太平洋に散在している天皇海山は,クサカ リツボダイやキンメダイを漁獲対象としたトロール 漁船の漁場になっている.そこでの漁獲物の中か ら,センジュエビ科の1種と考えられる個体が得ら れた.日本周辺において,駿河湾ではセンジュエビ (Polycheles typhlops)が,豊後海山ではセンジュエ ビ科の1種(Pentaceros laevis)やセンジュエビ科の 1種(Homeryon armarium)が漁獲されている(柳 本ら,2015).そこで,沖合域に分布する本個体が どの種になるのか調べた. 本個体は漁獲後にエタノールで固定されていた が,研究室で受け取った時点でエタノールが蒸発し てほとんどなかった.そのため,本個体の体色は変 色し,また第1胸脚は欠落していた.センジュエビ 科の分類については近年改変が著しく混乱が生じて いる(Torres, et al., 2014).これに対しては,外部 形態による種判別とDNA分析による化学的種判別 の両面から行うことで,より有効な情報と安定した 同定結果が期待される.本報告では,天皇海山で漁 獲されたセンジュエビ科の標本の種同定について, 外部形態とDNA分析により行った. 材料と方法 北 方 ト ロ ー ル 船 第5大 林 丸(稚 内 漁 業 所 属) によって2009年7月25日に北緯35度6.2分東経171 度30.8分の水深457 mで漁獲されたセンジュエビ
科の1種を用いた(Fig. 1).Galil (2000)やAhyong & Chan (2004)に基づき,外部形態から種判別した. 成体の腹節の筋肉片約10 mgを用いてQuickGene (Fuji Co.)によりDNAを抽出した.この抽出DNAを 用 い て,mtDNAのCOI領域について,PCR増幅を 行った. COI領域の増幅にはL5956(5′-cacaaagacattg-gcaccct-3′)とH6558(5′-cctcctgcagggtcaaagaa-3′)のユニ バーサルプライマーを用いた(Folmer, 1994).サー マルサイクラーにはABI9700(Applied Biosystems) を 用 い た.94℃2分加熱後,94℃30秒,55℃30秒, 72℃2分を30サイクル行い,最後に72℃で7分加熱し て,PCR反応を行った.DNA溶液1 μl, 2.5 mMdNTP 溶 液2.5 μl, 10×Buffer(TaKaRa)2.5 μl, 50 μMの 各 プライマー0.5 μl, TaKaRa Ex Taq Polymerase 0.125U,
総量が25 μlになるように超純水を加えPCR反応溶
1 国立研究開発法人 水産研究・教育機構 中央水産
研究所 水産生命情報研究センター
〒236–8648 神奈川県横浜市金沢区福浦2–12–4 Research Center for Aquatic genomics, National Research
Institute of Fisheries Science, 2–12–4 Fukuura, Kanaza-wa, Yokohama, Kanagawa 236–8648, Japan
E-mail: [email protected]
2 国立研究開発法人 水産研究・教育機構 中央水産
研究所 海洋・生態系研究センター
〒236–8648 神奈川県横浜市金沢区福浦2–12–4 Research Center for Fisheries Oceanography and Marine
Ecosystem, National Research Institute of Fisheries Sci-ence, 2–12–4 Fukuura, Kanazawa, Yokohama, Kanagawa 236–8648, Japan
44
Cancer 26 (2017) 柳本 卓・小西光一液 と し た.PCR産 物 を1.5% ア ガ ロ ー ス ゲ ル (NuSieve3:1, TaKaRa)で電気泳動し,エチジウムブ ロマイド染色によって増幅産物の確認をした.増幅 産 物 をQuick PCR Purification Kit(Qiagen)を用い て精製した.精製した増幅産物をテンプレートとし て,BigDye Terminator Kit Ver3.1(Applied Biosys-tems)を用いてシーケンス反応を行った.この反応
には,PCRで用いたのと同じプライマーを用いた.
シーケンス反応物をExo-SapIT(GEヘルスケア)で 精 製 し た 後,ABI PRISM 3730XL Genetic Analyzer (Applied Biosystems)にて電気泳動して塩基配列を 決定した. 決定した塩基配列について,インターネット上の 遺伝子相同性検索ソフトBlast(Altschul et al., 1990) によって,塩基配列がどのような種と近いかを検討 した.また,近縁種のデータがDNAデータバンクに 登録されている場合,それらの塩基配列と本研究で 得 ら れ た 塩 基 配 列 をClustal W(Thompson et al., 1994)によってアライメントを行った後,MEGA5.05 ソ フ ト ウ エ ア に よ っ て,Kimura two-parameter法 (Kimura, 1980)により遺伝距離を求め,近隣結合 法(Saitou & Nei, 1987)により系統樹を作成した.
系統樹の信頼性を1,000回のブーストラップ検定
(Felsenstein, 1985)によって評価した. 結 果
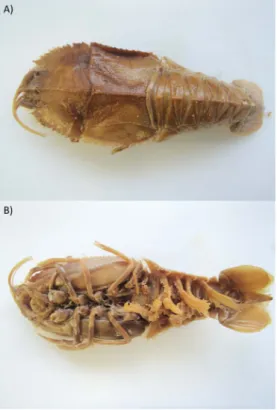
本個体は抱卵雌で,外部形態ではGalil (2000)や Ahyong & Chan (2004)に基づき,第3鍔脚の副肢が 座節よりとほぼ等長であり,頭胸甲は平滑で,かつ 眼窩の縁は深く V字状に切込むことなどからPen-tacheles laevisと同定された. 本サンプルは十分なエタノールによる固定がされ ていなかったため,変色していたが(Fig. 1),DNA を 抽 出 しPCR法により増幅産物が得られた.こ れをシーケンス分析した結果,717塩基配列が得 られた.Blast分析を行った結果をTable 1に示す. センジュエビ科のPentacheles laevis(LC126713)と
Fig. 1. Photographs of polychelid species collected at
Koko Seamount in the central North Pacific. A) Dorsal view, B) Ventral view.
Table 1. The results of Blast analysis for nucleotide sequences of Polychelidae collected at Koko Seamount.
Order DNA Acc. No. Species name Identities Identities (%)
1 LC126713 Pentacheles laevis 717/717 100 2 LC022129 Polychelida sp. 624/624 100 3 LC022127 Pentacheles laevis 622/624 99 4 LC022126 Pentacheles laevis 622/624 99 5 LC022128 Homeryon armarium 533/616 87 6 HQ241553 Polycheles enthrix 540/665 81 7 KF828019 Polycheles enthrix 529/654 81
8 KU496440 Galatheidae sp. JOD_0437 522/658 79
9 KU496438 Galatheidae sp. JOD_0212 521/658 79
45
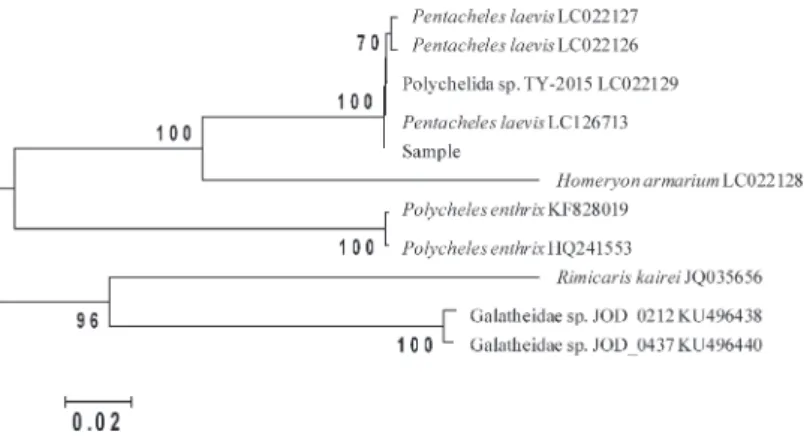
Cancer 26 (2017) 光孝海山で漁獲されたセンジュエビ類のDNA種判別 P. laevisの幼生(LC022129)が一致率100%で, P. laevis(LC022127, LC022126)が一致率99%で類似 していた.続いてセンジュエビ科のHomeryon armar-ium(LC022128) が 一 致 率87%, Polycheles enthrix (HQ241553, KF828019)が一致率81%で類似してい た.他はコシオリエビ科が一致率79%で類似して いた. サンプルと類似度が高かった上位10種の塩基配 列を用いて,NJ系統樹を作成した(Fig. 2).サン プルはP. laevisの塩基配列と1つのクラスターを形 成し,他の種から大きく離れていた. 考 察 光孝海山で漁獲されたセンジュエビ科の1種は, 外部形態からは第3顎脚の長さや頭胸甲の形によ り,Pentaceros laevisと同定された.mtDNAのCOI 領域の塩基配列のBlast分析によってP. laevisと一致 率100%で類似し(Table 1),またNJ系統樹を作成 した所,センジュエビ科の1種はP. laevisのクラス ターに属し,他種と大きく異なったいたことから (Fig. 2),P. laevisであることが分かった. P. laevisの分布(http://maps.iucnredlist.org)をみる と,全世界の熱帯や亜熱帯域に広く分布し,水深 200~3000 mに生息している(Galil, 2000).今回は 北太平洋の光孝海山での初めての採集結果となる. ただし,漁獲対象となっていないため,これまで漁 獲されても廃棄されていた可能性がある.また,他 のセンジュエビ類も採集される可能性もある.今 後,より具体的な種組成や生態などについてもモニ タリングする必要がある. センジュエビ科の分類については,近年の深海調 査の進展に伴い多くの標本が得られつつあることも あり,改変が著しいが(Torres et al., 2014),日本に おいてもこれに加え,異なる学名に対して同じ和名 が使われたり,またこの逆の事例もあり,更に混乱 している.センジュエビ科は一般的に漁獲量が少な く,また食料として利用されていない.そのため, 分類についての一般からの関心も低いために,取り 組みも進んでいないと考えられる.また,本科では 幼 生 に つ い て も ほ と ん ど 情 報 が な い(柳 本 ら, 2015).今後,これらの生態や生活史を含めた基本 的情報を充実させていく必要がある. 謝 辞 本研究で用いたセンジュエビ科の標本は稚内漁業 所属の第5大林丸によって漁獲された.この場を借 りて,漁労長はじめ乗組員の各位に厚くお礼申し上 げる. 文 献Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipan, D. J., 1990. Basic local alignment search tool. Journal of Molecular Biology, 215: 403–410.
Fig. 2. Neighbor-joining tree of the sequences of COI region constructed using Kimura two parameter distances
for polychelid species. Numbers beside internal branches indicates bootstrap probabilities based on 1,000 pseudoreplicates. Bootstrap probabilities higher than 70% are shown.
46
Cancer 26 (2017) 柳本 卓・小西光一Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783–791. Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrigenhoek,
R., 1994. DNA primers for amplification of mitochon-drial cytochrome c oxidase subunit I from diverse meta-zoan invertebrates. Molecular Marine Biology and Bio-technology, 3: 294–299.
Galil, B. S., 2000. Crustacea Decapoda: Review of the gen-era and species of the family Polychelidae Wood-Ma-son, 1874. Mémoires du Muséum national d’Histoire naturelle, 184: 285–387.
Kimura, M., 1980. A simple method for estimating evolu-tionary rate of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, 16: 111–120.
Saitou, N., & Nei, M., 1987. The neighbor-jointing method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4: 406–425. Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M.,
& Kumar, S., 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolution-ary distance, and maximum parsimony methods. Mo-lecular Biology and Evolution, 28: 2731–2739. Thompson, J. D., Higgins, D. G., & Gibson, T. J., 1994.
CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weight-ing, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22: 4673–4680. Torres, A. P., Palero, F., Dos Santos, A., Blanco, P. A.-E., &
Guerao, A.B-G., 2014. Larval stages of the deep-sea lobster Polycheles typhlops (Decapoda, Polychelida) identified by DNA analysis: Morphology, systematic, distribution and ecology. Helgoland Marine Research, 68: 379–397.
柳本 卓・小西光一・髙見宗広・猿渡敏郎,2015.本
州太平洋沿岸で採集されたセンジュエビ科の後期 幼生のDNA種判別.Cancer, 24: 7–13.