近畿大学学術情報リポジトリ
21
0
0
全文
(2) Doctoral. Changes. in activity. neurons. after. Dissertation. of substantia middle. cerebral. in the. School, Course. artery. rat. Toshiharu. Graduate. nigra pars. Asai. Kinki. University. of Medicine. Department. of Physiology. ( Director : Prof. Shiko Chichibu ). September. 1991. reticulata. occlusion.
(3) Changes in activity of substantia neurons after middle cerebral in the rat. Toshiharu. nigra pars reticulata artery occlusion. Asai. Department of Physiology, Kinki University School of Medicine, Osaka, Japan ( Director : Prof Shiko Chichibu. ABSTRACT. The spontaneous single unit activity of substantia nigra pars reticulata (SNR) neurons was studied in control and in ipsilateral middle cerebral artery (MCA) -occluded rats. The activity of MCA-occluded preparations was recorded in 3 groups; 1 hour, 24 hours, and 7 days after MCAocclusion. Activity was recorded extracellularly with glass microelectrodes and was analyzed using time interval histograms. In the control, half of the SNR units (47%) showed a regular firing pattern. There was no change 1 hour after MCA occlusion. After 24 hours and 7 days, however, there was increased SNR neuronal unit activity with irregular firing patterns (67% and 57%, respectively), and a slightly decreased mean firing rate. In the MCA-occluded groups, no hyperexcited neurons at all were found in the SNR. Our results indicated that SNR neuronal degeneration accompanied by striatal ischemia does not depend simply on hyperexcitation due to a disinhibition mechanism.. Key words:. substantia. nigra pars reticulata. hyperexcitation. , unit activity,. MCA occlusion,. disinhibition,.
(4) INTRODUCTION In the basal ganglia, the striatum It has been well-established, that. GABA-ergic. nigra are closely connected.. by both anatomical and physiological. striato-nigral. influence on substantia. and the substantia. neurons,. in particular,. investigations,'. exert a potent. inhibitory. nigra pars reticulata (SNR) neurons.. It has been recently shown that, in the rat, striatal ischemia induced by middle cerebral. artery. (MCA). neuronal. death within. occlusion. led to delayed. within. have speculated. of glucose utilization. the SNR after ipsilateral. themselves,. the deafferentation. technique,. due to a disinhibitory. 205. and. began.. 305g.. The. which occurs as a consequence. inhibitory. to discover. these neurons. whether. pathway.. However,. of. there. they actually. exhibited. hyperactivity. AND. METHODS. and anesthesia were. performed. Animals. anesthetic. is. directly, using a single unit. mechanism.. MATERIALS. Experiments. many investigators. data that supports this idea.. In the present study, we examined. Preparation. From. and of blood flow were. striatal ischemia,5. hyperexcitation. of the striato-nigral. no electrophysiological. 1.. area.'. or. that the changes in SNR neurons are caused by hyperexcitation. within the neurons. recording. degeneration. the SNR in areas remote from the ischemic. evidence showing that augmentation observed. pathological. were. on. 57 adult. usually. used during. kept. the operation. —2—. male. Wistar. for several and. rats days. recording. weighing before periods. between. experiments was chloral.
(5) hydrate (400 mg/kg i.p.). Additional injections of chloral hydrate were administered intramuscularly when necessary. Body temperature was maintained at 37°C, with a warming pad, throughout the experiment.. 2. Middle cerebral artery occlusion We exposed the left MCA by the transretroorbital. approach and occluded it. by electrocoagulation, using bipolar forceps. The occlusion site was made proximal to the olfactory bulb to produce focal striatal ischemia. Surgery was performed visually under an operating microscope. Animals in the control group were intact; sham MCA occlusion was not carried out.. 3. Extracellular unit recording At periods of 1 hour, 24 hours, and 7 days after the MCA occlusion, each animal was re-anesthetized and fixed on a stereotaxic apparatus. A small burr hole was drilled in the skull at a site 2.0 mm lateral to the sagittal suture and 5.0-6.0 mm posterior to the bregma on the left side. The dura was removed, and a singlebarrel electrode was passed through the hole and lowered into the brain with a hydraulic microdrive to the level of the SNR. The neurons recorded were located at the stereotaxic coordinates of 4.0-4.9 mm A, 1.9-2.5 mm L and 6410-8900 um H according to the atlas of Paxinos and Watson.6 Using a Narishige PE-2 electrode puller, we pulled single-barrel recording electrodes from 2.0 mm-glass capillary tubing prefilled with several fine glassfibers. After completing this process, we filled micropipettes with 1% Pontamine Sky Blue (Tokyo Kasei, Tokyo) in 2M NaC1 solution. Tip diameters of the record-. - 3—.
(6) ing electrodes. were approximately. I pm, and the ohmic resistance typically ranged. from 2.0 to 5.0 Ms/. Electric signals passed through a high input-impedance W-P Instruments. Inc., New Haven, Conn.) were filtered with a bandpass. 3000 Hz, and displayed SNR. amplifier (Model M701,. neurons. on a storage oscilloscope. were. (1) They had smooth,. easily. identified. of 50-. (VC-11, Nihon Koden, Tokyo).. according. to. sharp, biphasic action potentials. the. following. of 0.5-0.7. criteria:. msec duration,. and higher firing rates than SNC neurons, normally ranging from 10 to 40 spikes/ sec. (2) dopamine. They. were. neurons,. located. ventral. which mostly. to substantia. showed. longer. nigra duration. pars compacta. (SNC). and lower firing rates. than SNR neurons.' Spontaneous TEAC, Tokyo). Ltd., Tokyo). unit activity was recorded. on a magnetic tape recorder (RD-110T,. The output was fed to a signal processor (7T07, San-ei Instruments, to obtain. a time interval histogram. At the end of each experiment, ejection of Pontamine. the electrode. Sky Blue (15 pA cathodal,. (TIH) for subsequent. tip was marked. analysis.. by iontophoretic. 20 min).. 4. TTC staining and cell identification After. the experiments. above were completed,. the animals were decapitated. and the brains were quickly removed. The anterior with 2,3,5-triphenyltetrazolium. chloride. (TTC). half of the brain was stained. for evaluation. brain region. TTC staining was carried out according. of the infarcted. to the method. of Bederson. et aL 8 The. posterior. half of the brain was preserved. —4—. in a 10% buffered. formalin.
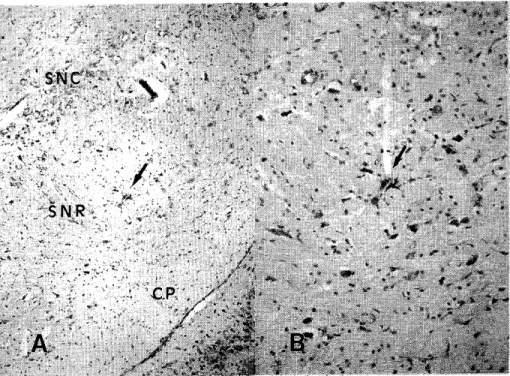
(7) solution,. and then immersed in a 30% sucrose solution overnight.. 20 µm thick, were cut on a cryostat, localization. mounted,. Frozen sections,. and stained with neutral. red for. of electrode tips and tracks.. RESULTS. 1. Localization. of recorded neurons. A total of 75 spontaneous out the experiments.. unit activities was recorded. Of these,. 15 were recorded. from 57 animals through-. from 15 normal rats (control. Fig. 1 Location of a recorded neuron in the SNR A: The arrow indicates the ejection site of Pontamine Sky Blue (X100). Counterstain: neutral red. B: Higher magnification (X200). Counterstain: neutral red. Abbreviations: SNC, substantia nigra pars compacta; SNR, substantia nigra pars reticulata; CP, cerebral peduncle..
(8) o. CONTROL. • 1 hr. Fig. 2 Localization of recording drawings of the SNR in control rations Abbreviations:. A 24. hr. *. day. sites, shown on serial antero-posterior and in ipsilateral MCA-occluded prepa-. refer to the legend in Fig.l..
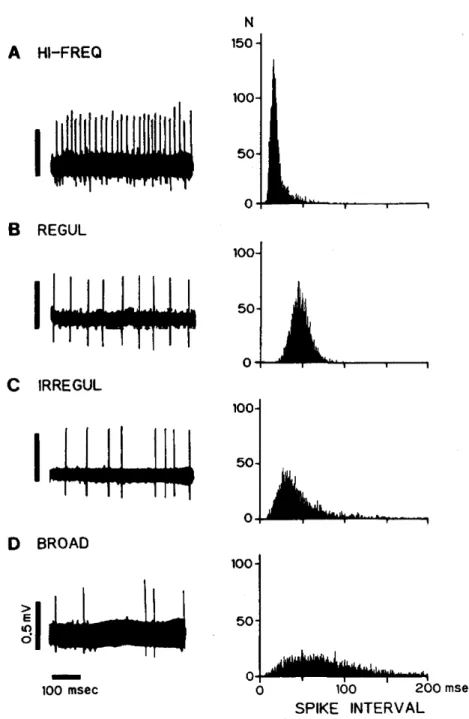
(9) group);. 18 were recorded. 21 were recorded. from. from 14 rats,. 1 hour after MCA occlusion. 14 rats, 24 hours after MCA occlusion. ( 1h group);. (24h group); and. 21 were recorded from 14 rats, 7 days after MCA occlusion (7d group). Figure 1 shows an example of an electrode verified the location of a recorded. neuron. track and typical dye-marking. within the SNR. The recorded. that. neuron. is stained as a blue spot close to the soma and is located in the center of the photograph. The electrode was advanced along a dendrite. Localizations. of neurons recorded. Figure 2. All neurons. recorded. dorsal to the cerebral peduncle.. in the four experimental. were located. groups are shown in. in the SNR, ventral to the SNC and. No topographical. differences. were found among. the groups, and similar samples were obtained from all groups.. 2. Four firing patterns, as classified by TIH Spontaneous unit activities recorded in the SNR were subsequently divided into four patterns by TIH analysis. In Figure 3, inset A had a high-frequency pattern, in which every spike interval was short (8.5 ± 7.6 msec) and in which the firing rate was very high (up to 117.6 spikes/sec). The TIH was narrow in shape and the peak interval was remarkably short (15 msec). Inset B had a regular pattern, in which most spike intervals were identical (49.6 ± 10.5 msec). The TIH had a symmetrical shape, reflecting a binomial distribution. Inset C had an irregular pattern, with longer intervals which were often varied. The TIH had an asymmetrical shape, but the peak interval was recognizable (40 msec). Inset D had a broad pattern, the spike intervals were unequal, and intervals which were even longer than those in inset C appeared. The TIH.
(10) Fig. 3 Examples of the four spontaneous firing patterns recorded in SNR neurons Left: Representative unit activities. Right: Time interval histograms (TILL bin width: 0.2 msec, sample size: 5000 intervals). Abbreviations: HI-FREQ, high-frequency; REGUL, regular; IRREGUL, irregular.. — 8 —.
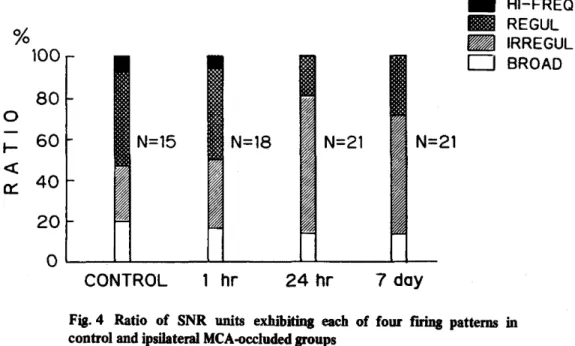
(11) had a flat shape and no distinguishable peak. No significant differences among the patterns of the recording sites could be found by histological examination.. 3. Changes in unit activity after MCA occlusion Firing patterns in the four experimental groups are shown in Figure 4. (Ratio = representative units / total units (%) ) In the control group , 47% of the total showed a regular pattern, which was consistent with earlier reports.'. In the 1h. group, the ratio was similar to that in the control (44%). Compared with the control group and the 1h group, there was a relatively increased ratio of irregular pattern in the 24h group and the 7d group (67% and 57%, respectively). In contrast, the ratio of their regular patterns was reduced (19% and 29%, respectively). No highfrequency pattern was found at all.. Fig. 4 Ratio of SNR units exhibiting each control and ipsilateral MCA-occluded groups Abbreviations: refer to the legend in Fig.3.. —9—. of four. firing. patterns. in.
(12) ..f1.1. •1.1.J. :".1. UV. f 1.0. OV. =LI. 1VU. I IIJ. IGLI. spikes/sec. MEAN. FIRING RATE. Fig. 5 Distribution of mean firing ipsilateral MCA-occluded rats. rates of SNR neurons. — 10 —. in control. and.
(13) The mean firing rate for 5000 spikes was also analyzed. The histogram for each group is shown in Figure 5. In the control group, the rates ranged from 10 to 65 spikes/sec, while in the lh group, the rates ranged from 5 to 120 spikes/ sec. In comparison with the control group and the 1h group, the rates of the 24h group were relatively low, ranging from 5 to 30 spikes/sec. Similarly, rates in the 7d group ranged from 10 to 35 spikes/sec. In the latter two groups, the firing rate did not exceed 35 spikes/sec.. 4. Evaluation of ischemic area The ischemic area was easily recognized by TTC staining. As shown in Figure 6, the entire left striatum became ischemic after ipsilateral MCA occlusion. The cerebral cortex became partially ischemic. No apparent ischemic change was found in the ipsilateral globus pallidus. Of the total of 42 MCA-occluded rats, only three exhibited striatal ischemia only. In these cases, the cerebral cortex was intact and did not become ischemic..
(14) Fig. 6 TTC-stained coronal brain slices 24 hours after left MCA occlusion The white regions show areas that failed to react with tetrazolium. A: Section at the level of the anterior commissure. B: Section through the globus pailidus.. Scale bars = 5 mm. — 12 —.
(15) DISCUSSION In this study, unit recording. we examined. technique. neuronal. activity.. neuronal. changes. be necessary. Using in the. for. all experimental. 1.. our. activity. directly. had previously. proven. to be useful. this. technique,. however,. same. preparation. before. discussion. to note. that. we. , using an extracellular. did. for detecting. not. and after. observe. dynamic. MCA occlusion. we obtained. similar. such. . It will. samples. from. groups.. TIH analysis We divided. classification. SNR. in Figure. modes. to SNR. inputs. either. from. may. B pattern. presented. longer. drive. spike. stronger Since. received. a quite. from. stronger. into. kinds. as flat. four patterns,. indicating. preparations. wide. TIH. and. other. influence.". tonic. discharge,. that. showing. neurons. in which. influence. showing. the. that. striatum. SNR. inhibitory. Neurons. showing. the. Neurons. showing. the. and. appeared. to receive. the C and D patterns these. patterns. included. had received. the B pattern. became. neurons. ischemic. in the striatal. than those in the control. — 13 —. in the input. for dense. afferents.. Both. than neurons. TIH analysis . This. differences. distribution. in the CNS.. we postulated. inhibitory. of TIH reflected. facilitatory. regular. using. reports.9. or from. some. somewhere. influence. the C pattern,. such. received. intervals,. the. 3, the four. inter-neurons)°. have. inhibitory. activity. to that in previous. neurons,. A pattern. a regular. neuronal. corresponded. As shown. hibit. that. SNR neuronal. group.. tended ischemic. to exgroups.
(16) 2.. An earlier proposal for explaining SNR changes It is known that a pathological. the occurrence after striatal. of striatal. lesion.". change occurs in the SNR at 7 to 14 days after. ischemia.4. This pathological. It has also been reported. change. is also observed. that glucose utilization. is elevated. in the SNR over the same period.5,13,14 Many. investigators. ergic striato-nigral and that. to. the. inhibitory. the neurons. considered. have speculated pathway. mechanism. causing. proposed idea is summarized. lesion.". neuronal. death. in excitatory. amino. is. and is similar acids.4 ' 5 The. prevented. to rats after striatal. of glucose use in the SNR was. of muscimol. after. the lesion.". These. in this neuronal death.. above depends on indirect evidence. both glucose. after GABA-ergic deafferentation.. intraventricularly. that elevation. administration. However, the idea outlined has shown that. and die. The mechanism. that the GABA agonist, muscimol,. data suggest some GABA participation. which. of SNR neurons,. death due to disinhibition. death when it was administered. by intravenous. of the GABA-. in Figure 7.. Kelly et al. also reported. attenuated. hyperexcited. neuronal. Saji et al. have further reported SNR neuronal. the deafferentation. causes a disinhibition. finally become. to be transsynaptic. that. utilization. and cerebral blood. only, evidence flow increase. There is no direct evidence to account for this. proposal. In the present directly obtain. study, we evaluated. by using an extracellular. the change in SNR neuronal. unit recording technique,. activity more. and we were able to. evidence that, after striatal ischemia, SNR neurons failed to hyperexcite. accordance. with this hypothesis.. —14—. in.
(17) GABA—ERGIC °e.g. DEAFFERENTATION. DISINH1BITION. ?. EXCITATION. ?. NEURONAL. DEATH. Fig. 7 Diagram showinga proposed disinhibition mechanism For explanation, see text. Abbreviations: CPut, caudate putamen (striatum); GP, globus pallidus; SNC,substantia nigra pars compacta; SNR, substantia nigra pars reticulate.. 3.. Evaluation. of our results. The existence has. been. flows.. of both. well-established. 7 - 2 o These. locomotion, pathways.. and SNR. striato-nigro-thalamic by. experiments. extrapyramidal SNR. neurons. neurons. systems play. discharge. and striato-nigro-collicular on. antero-. participate. an important spontaneously. — 15 —. and. retrograde. in postural role. pathways. as relay. at a relatively. axoplasmic symmetry. stations higher. and. in these rate. than.
(18) neurons. in various. neurons.1'2,7. regions. of the. brain,. Our data for the control. and. they. tonically. inhibit. efferent. group agreed with previous reports with. respect to the discharge rate and the firing pattern.' The striatal also Substance. neurons. P-ergic excitatory. some other projections nucleus'''28. send not only GABA-ergic axons23-26. inhibitory. to the SNR. The globus pallidus3,22,27 and the subthalamic. extend to the SNR and form synapses with SNR neurons!°. of SNC neurons. SNR and SNC neurons. influence on each other, so that there are multiple inputs regulat-. ing SNR neurons. the SNR following. From. the evidence. lesions. that. irregular. neuronal. firing appeared in. of the SNC,9 it is suggested that the striatal outputs. regulate other efferent neurons. in a manner. consider that striatal regulation. maintains. relatively. but. to the SNR. There are, in addition,. also send their axons to the SNR. The dendrites. exert a reciprocal. axons3'21'22. similar to that of the SNC. We also. a regular discharge of SNR neurons at a. high rate. After striatal ischemia, in fact, TIH in the 24h group and the. 7d group showed firing patterns we speculate. without. such striatal regulation.. that the irregular firing of SNR neurons. result from the impairment. In conclusion,. after striatal ischemia may. of striatal regulation.. our data indicate that the remote and delayed neuronal. that occur in the SNR accompanied on hyperexcitation. From our results,. due. to. with striatal ischemia. disinhibition. mechanism involved in neuronal change.. mechanism,. but. changes. do not depend simply depend. on another.
(19) ACKNOWLEDGMENTS We thank sity. School. this study.. Prof.. Shiko. of Medicine,. Chichibu, for. First. his helpful. Department advice. and. of Physiolosy, encouragement. Kinki. Univer-. throughout.
(20) REFERENCES. 1. McNair JL, Sutin J, Tsubokawa T. Suppression of cell firing in the substantia nigra by caudate nucleus stimulation. Exp Neurol 1972;37:395-411. 2. Dray A, Gonye TJ, Oakley NR. Caudate stimulation and substantia nigra activity in the rat. J Physiol 1976;259:825-849 . 3. Ribak CE, Vaughn JE, Roberts E. GABA-ergicnerveterminals decreasein the substantia nigra following hemitransections of the striatonigraland pallidonigralpathways. BrainRes 1980;192:413-420 . 4. Tamura A, Kirino T, Sano K, Takagi K, Oka H. Atrophy of the ipsilateral substantia nigra following middle cerebral artery occlusion in the rat. Brain Res 1990;510:154-157 . 5. Tamura A, Sano K. Effect of striatal infarction on the ipsilateral substantia nigra. Adv Neurol Sci 1990; 34:968-975 (in Japanese) 6. Paxinos G, Watson C. The rat brain in stereotaxic coordinates, 2nd ed: Sydney: Academic Press, 1986. 7. Guyenet PG, Aghajanian GK. Antidromic identification of dopaminergic and other output neurons of the rat substantia nigra. Brain Res 1978;150:69-84 . 8. Bederson JB, Pitts LH, Germano SM, Nishimura MC, Davis RL, Bartkowski HM. Evaluation of 2,3,5triphenyltetrazolium chloride as a stain for detection and quantification of experimental cerebral infarction in rats. Stroke 1986;17:1304-1308 . 9. Sanderson P, Mavoungou R, Albe-FessardD. Changesin substantia nigrapars reticulata activity following lesions of the substantia nigra pars compacta. NeurosciLett 1986;67:25-30. 10. Juraska JM, WilsonCJ, GrovesPM. The substantia nigra of the rat: a Golgi study. J Comp Neurol 1977; 172:585-594. 11. Robledo P, Foger J. Excitatory influence of rat subthalamic nucleus to substantia nigra pars reticulata and the pallidal complex: electrophysiologicaldata. Brain Res 1990;518:47-54. 12. Pearson RCA, Neal JW, Powell TPS. Bilateral morphological changes in the substantia nigra of the rat following unilateral damageof the striatum. BrainRes 1987;400:127-132 . 13. Kelly PAT, Graham DI, McCullochJ. Specific alterations in local cerebral glucose utilization following striatal lesions.Brain Res 1982;233:157-172 . 14. Hosokawa S, Kato M, Shima F, Tobimatsu S, Kuroiwa Y. Local cerebral glucose utilization altered in rats with unilateral electrolytic striatal lesions and modification by apomorphine. Brain Res 1984;324: 59-68. 15. Saji M, Reis DJ. Delayed transneuronal death of substantia nigra neurons prevented by 7-aminobutyric acid agonist. Science 1987;235:66-69 16. Kelly PAT, McCulloch J. Cerebralglucose utilization following striatal lesions: the effects of the GABA agonist, muscimol,and the dopaminergicagonist, apomorphine. Brain Res 1987;425:290-300 . 17. Bunney BS, Aghajanian GK. The precise localization of nigral afferents in the rat as determined by a retrograde tracing technique. Brain Res 1976;117:423-435. 18. Faull RLM, MehlerWR. The cells of origin of nigrotectal, nigrothalamic and nigrostriatal projections in the rat. Neuroscience 1978;3:989-1002 . 19. Tulloch IF, Arbuthnott GW, Wright AK. Topographical organization of the striatonigral pathway revealedby anterograde neuroanatomicaltracing techniques.J Anat 1978;127:425-441. 20. Williams MN, Faull RLM. The striatonigral projection and nigrotectal neurons in the rat. A correlated light and electron microscopic study demonstrating a monosynaptic striatal input to identified nigrotectal neurons using a combined degeneration and horseradish peroxidase procedure. Neuroscience 1985;14: 991-1010 . 21. Hattori T, McGeer PL, Fibiger HC, McGeer EG. On the source of GABA-containing terminals in the substantia nigra. Electron microscopic autoradiographic and biochemical studies. Brain Res 1973;54:. —1;8 —.
(21) 103-114. Araki M, McGeer PL, McGeer EG. Striatonigral and pallidonigral pathways studied by a combination of retrograde horseradish peroxidase tracing and a pharmacohistochemical method for 7-aminobutyric acid transaminase. Brain Res 1985;331:17-24 . 23. Walker RJ, Kemp JA, Yajima H, Kitagawa K, Woodruff GN. The action of substance P on mesencephalic reticular and substantia nigral neurons of the rat. Experientia 1976;32:214-215 . 24. Davies J, Dray A. Substance P in the substantia nigra. Brain Res 1976;107:623-627 . 25. Gale K, Hong JS, Guidotti A. Presence of substance P and GABA in separate striatonigral neurons. Brain Res 1977;136:371-375. 26. Hong JS, Yang H-YT, Costa E. Projections of substance P containing neurons from neostriatum to substantia nigra. Brain Res 1977;122:541-544 . 27. Smith Y, Bolam JP. Neurons of the substantia nigra reticulata receive a dense GABA-containing input from the globus pallidus in the rat. Brain Res 1989;493:160-167 28. Hammond C, Deniau JM, Rizk A, Feger J. Electrophysiological demonstration of an excitatory subthalamonigral pathway in the rat. Brain Res 1978;151:235-244 . 22..
(22)
図

+4



関連したドキュメント
The re- sults presented in Table 3, showing that total lipase activity (measured in the absence of 1 M NaCl) was similar to HL activity (measured in the presence of 1 M NaCl) in
The pharmacokinetic profiles of clenbuterol enantiomers following intravenous and intraduodenal administration of clenbuterol racemate (2 mg/kg) in rats were
This study was therefore designed to examine the induc- tion of CYP2E1 in genetically obese Zucker rats fed a normal diet (OB) and its effect on the disposition kinetics of CZX and
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
[Mag3] , Painlev´ e-type differential equations for the recurrence coefficients of semi- classical orthogonal polynomials, J. Zaslavsky , Asymptotic expansions of ratios of
Cheetah Max may be applied as a preplant surface or preemergence burndown application or as a postemergence application with hooded spray equipment in cotton and as a preplant
“Indian Camp” has been generally sought in the author’s experience in the Greco- Turkish War: Nick Adams, the implied author and the semi-autobiographical pro- tagonist of the series
: Associations betwwen spectral repre sentation of the surface electromyo gram and fibre type distribution and size in human masseter muscle.. : Motor unit activity and
