ペクチンオリゴ糖連続生産のための
回一、、応クロマ卜装置の開発 課題番号:07555548 平成7年度∼平成8年度科学研究費補助金基盤糊(1研究成果報告書
平成10年3月23日 研究代表者 米本年邦東北大学工学部教授
はしがき
研究組織
研究代表者
研究分担者
研究分担者
研究協力者
研究協力者
米本年邦(東北大学工学部教授) ] u I l明生(宮城工業高等専門学校材料工学科講師)櫛引正剛(青森県産業技術開発センター技師)
]urr尚美(東北大学工学部助手) 岩村賢一郎(東北大学工学研究科院生)研究経費
平成7年度 4,(X氾千円 平成8年度 1,500千円 計 5,500千円研究発表
(1)学会誌等(N.Shibasaki-Kitakawa etal., hetic Modelfor Oligosaccharide
Hydrolysis using Suspendedand ImmobiliZX:d Enzymes.,
Biochem Eng. J. (inp血ting) ,平成10年3月23日)
(2)口頭発表(KIwamm et aL, Hydrolysis hetks of Ohgosaccharideusing
Immobilized Dextranase.,化学工学会第30回秋季大会, 平成9年9月8日) ■■■llll ■■■■■l ■■■■■_ ■■■■■■一ll ■■■■一 ■■■11 ■■■llll ■■llll ■■lllll ■■■■■l ■■■l ■■■■■■■■ ■■l1一■l ■l ■■■■■ll -00010134173
目次
第1章 緒論 1.1本研究の背景および目的 1.2 既往の研究 1.3 本研究の概要 第2章 回分反応システム 2.1 均相系における酵素加水分解実験 2.1.1実験 2.1.2速度論モデルの構築 2.1.3結果と考察 2.1.3.1モデルの適用 2.1.3.2適用結果 2.2 固定化系における酵素加水分解実験 2.2.1実験 2.2.1.1固定化酵素の調製 2.2.1.2加水分解実験 2.2.1.3分配平衡実験 2.2.2速度論モデルの構築 2.2.2.1基礎式の導出 2.2.2.2無次元化 2.2.3結果と考察 2.2.3.1分配平衡実験 2.2.3.2速度論モデルの適用 2.2.3.3溶液中の各糖濃度の経時変化 2.2.3.4担体内の各糖濃度分布と反応速度分布…‥‥…‥…… 2.2.3.5モデル定数の推算値 2.2.3.6反応速度に及ぼす各種操作因子の影響 . 第3章 連続反応システム 3.1速度論モデル 3.1.1基礎式の導出 3.1.2無次元化 3.1.3計算方法 l 1 2 4 1 4 4 4 5 ∠U 7 7 qノ l 1 3 5 7 0 3 5 5 5 7 0ノ 0ノ l 1 1 1 1 1 1 1 l つム つ一 2 2 つん 3 3 ′hU ′hU ′hU nB 1 3 3 3 3 43.2 実験 3.2.1実験装置 3.2.2連続加水分解実験 3.3 結果と考察 3.3.1リアクターからの流出液中の各糖濃度 3.3.2反応速度に及ぼす各種操作因子の検討..………‥…… 第4章 総括 使用記号 引用文献 Appendix :担体内の物質移動抵抗を考慮しない速度論モデル………--・・ 43 43 44 46 46 50 53 54 56 57
第1章 緒言
(1.1本研究の背景および目的
近年、農業加工残浮に含まれる多糖類を酵素加水分解することにより、生理活性を有す る付加価値の高いオリゴ糖が得られることが報告されている1)。特に、リンゴジュースの 生産工程で副成されるリンゴ搾汁残梓に含まれる多糖ペクチンを加水分解して得られる重 合度4程度のペクチンオリゴ糖には、植物を雑菌類の感染から防御したり、植物培養細胞 による有用物質生産を促進するなどの働きを有することが知られている1)。このようなペ クチンオリゴ糖を工業的規模で生産するプロセスが構築されれば、膨大な量に達する農業 加工残梓を有効利用することが可能となるため、環境問題解決のための一方策になると考 えられる。 酵素は一般に、生体から抽出・精製して調製されるために非常に高価なものであり、ま た、 pHや熟に対して不安定なため失括しやすい。従って、酵素を工業規模で利用するた めには、多孔性ゲルや樹脂、マイクロカプセル、膜などの不溶性の担体に固定化すること により、安定化するとともに、反応後に回収・再利用する技術が必要となる。さらに、プ ロセスの工業化には、このような固定化酵素を充填したカラム型リアクターによる連続生 産プロセスの構築、ならびに反応の解析や予測、最適操作条件の探索などを目的とした速 度論モデルの構築も重要となる。 これまでに、酵素の固定化法やそのための最適条件の探索に関しては様々な研究が行わ れており、数多くの知見が蓄積されている2).これに対し、酵素反応に関する速度論的研 究は数少なく、その大半は均相酵素反応におけるMichaelisIMenten型の速度定数を求めた ものである3)。特に、固定化酵素反応系では、担体内外の物質移動抵抗が存在するので、 酵素固有の速度定数を実験的に決定することが困難であり、見かけの速度定数を求めるた めの種々の方法が検討されているにすぎない4)■)0 一方、高重合度の糖類の酵素加水分解は、種々の大きさをもつ糖の生成やその糖のさら なる分解を含む逐次的で複雑な反応であるため、反応機構の解析が極めて困難である。与 れまで多糖類の酵素加水分解反応に関していくつかの速度論モデルが報告されているが、 そのほとんどは回分反応器を用いた均相酵素反応に関するものである7)Ill)。また、これら の研究では極度に簡略化した反応機構に基づいて速度論モデルを構築しているにすぎず、 多糖の加水分解によって生じる種々の分子量の糖を定量的に分析することが困難であるため、実験的な検証もほとんど行われていない。 本研究では、多糖ペクチンの酵素加水分解プロセス設計のための基礎的研究として、ま ず、糖琴の逐次的な酵素加水分解プロセスの解析や予測を可能とする、より現夷的な反応 機構に基づいた速度論モデルを構築する。そして、モデル系として、基質に逐次加水分解 が起こる最も小さな糖であり、かつ、分析の容易な重合度6のオリゴ糖(イソヘキサマル トース)を、酵素にこの基質を加水分解するエンドデキストラナ-ゼを取り上げ、回分反 応器を用いて種々の条件下で均相酵素加水分解実験を行い、モデルの安当性を実験的に検 証する。次に、この酵素を固定化し、同様に回分反応器を用いて種々の条件下で固定化酵 素加水分解実験を行い、固定化系における速度論モデルを構築する。さらに、この酵素加 水分解プロセスの連続化を目的として、固定化酵素を充填したカラム型リアクターを設 計・製作し、種々の条件下で連続加水分解実験を行う。そして、本連続反応プロセスに関 する数学モデルを構築し、実験結果と合わせてプロセスの最適な運転条件を探索する。
1.2 既往の研究
これまでに多糖類の酵素加水分解に関していくつかの速度論モデルが報告されている7)-ll)。 Fujiiらは、エンド型活性を有するa-アミラーゼとユキソ型活性を有するグルコアミ ラーゼを用いたデンプンの加水分解に関する速度論モデルを構築している8)。エンド型活 性を有する酵素は、基質となる多糖の内側のα-1,6結合をランダムに切断する性質を持 つため、その分解生成物は広い分子量分布を示すが、エキソ型活性を有する酵素は、多糖 の末端基のα-1, 6結合から順に切断する性質を持つため、その分解生成物は単糖あるい は二糖であることが知られている。彼らは、基質の分子量が充分に大きい場合、エンド型 酵素とユキソ型酵素の反応性は等しいが、エンド型酵素による加水分解が進行するにつれ てユキソ型酵素に分解される基質濃度が増えると仮定し、 Michaelis-Menten型の反応機構 に基づき、還元糖の生成速度を次式で表している。dCR V-TCs(0)en . V-rcs(0)
dt KLnxo+Cs(0)ed ■ KLn血+cs(o) (2-1) ここで、右辺の第一項はユキソ型酵素に分解されて生じる単糖の生成速度であり、第二項 はエンド型酵素に分解されて生じる還元糖の生成速度である. Kn.exD 、 vEnSはユキソ型酵素に関するMichaelis定数と最大反応速度、 K;ndo 、 vn.e禁はエンド型酵素に関するMichaelis
定数と最大反応速度、 cRは還元糖の濃度、 cs(o)は初期基質濃度、 Kはユキソ型酵素に関
は"lumped model"と呼ばれており、この系で生成する種々の分子量を有する糖をすべて1 つの基質としてまとめて考慮している。彼らは、実際に前述した2種類の酵素を用いて加 水分解琴験を行い、分解生成物として還元糖の総量のみを分析し、 Lineweaver-Burk plotに より求めた各酵素の最大反応速度およびMichaelis定数を用いて(2-1)式により還元糖の生 成速度を求めている。このモデルによる計算値は、反応の初期段階でのみ実験結果と一致 しているが、極度に簡略化されているため、生成物の分子量分布などを予測することがで きない。 これに対してSugaらは、多糖の逐次的な加水分解反応を表現するためには"lumped model"では限界があることを認識し、基質や生成物の分子量分布を考慮した''distributed model"を提案している,).彼らは、重合度i(i=1,2,3, --)の糖の生成や消費をそれぞれ Mich・aelis-Menten型の反応速度式で表現しており、エンド型とユキソ型酵素が混在してい る場合の一般化されたモデル式として次式を提案している。 dCsL VneQ(i-1)Csl dt CO 2Vmenaxdo ∑ CSL )・=1 7' V芸Csi + V.nS Cs.I. 1
KSn血+∑(i-1)Csi K;n血+∑(i-1)CsL Kmw +∑csI Kmw+∑cs.・
i=2 i=2 i=Z i=2 (2-2) ここで、右辺の第一、二項はエンド型酵素による基質の消費や生成を、第三、四項はユキ ソ型酵素による基質の消費や生成を示している. cs`は重合度iの糖濃度である。このモ デルでは、 "lumped model"よりも実際の複雑な現象に近い反応機構を考えているが、反応 生成物の分子量分布を実験的に測定することが困難であるため、実験によるモデルの検証 は行われていない。さらにokazakiらは、 sugaらのモデルを拡張することにより、反応活 性が異なる3種類の酵素セルラーゼを用いたセルロースの加水分解に関する速度論モデル を構築している10)。彼らは、実験的に観察される各酵素同士の相互作用を述べる基本的な モデル定数を定量化して論じているが、 sugaらの場合と同様に、基質や生成物の分子量 分布に関する実験的な測定が困難であることから、実験的な検証は行っていない。 Deanらは、sugaらのモデルを改良することにより新たな速度論モデルを構築している11)0 彼らは、基質である多糖の主鎖にある程度の間隔で枝分かれ部分があり、その部分は酵素 によって加水分解されない、また、 3糖以下はエンド型酵素によって分解されないという さらに現実的な事象を取り入れた。モデル式は、 Micbaelis-Menten型の反応機構を考慮し、 次式で表される。
dCsi = dt vmT (i I 3)Cs.・ CO 2Vmeio ∑ csi )・=2 1' vnF:Csi + Vz芸C5,.1 co co I KLndo・∑(i-3)Csi K;nab.∑(il3)Cs` Kmw'∑cs` Kmw'∑Csi
i=4 i=4 i=2 i=2 (2-3) sugaらの場合と同様に、右辺の第一、二項はエンド型酵素による基質の消費や生成を、 第三、四項はユキソ型酵素による基質の消費や生成を示している。彼らは、エンド型およ びユキソ型デキストラナ-ゼを用いてデキストランの加水分解実験を行っている。彼らの モデルによる計算値は、反応時間が短い場合では実験結果と一致しているが、長い場合で は一致しなかった。彼らは、この原因を実際の反応では酵素阻害などのより複雑な現象が 起こっているためであると考えている。このように現在のところ、多糖の酵素加水分解を 良好に表現できる速度論モデルは構築されていない。
1. 3本研究の概要
本研究の概要を以下に示す。 第1章では、本研究の背景および目的について述べるとともに、本研究の概要を示す。 第2章では、回分反応器を用いて、種々の条件下で均相酵素加水分解実験を行い、この系 に関する速度論モデルを構築する。次に、この酵素を固定化し、同様に回分反応器を用い て種々の条件下で固定化酵素加水分解実験を行い、固定化系における速度論モデルを構築 する。 第3章では、固定化系に関する速度論モデルを拡張することにより連続酵素加水分解シス テムに関する速度論モデルを構築し、このモデルに基づき、固定化酵素を充填したカラム 型リアクターを設計・製作する。また、このリアクターを用いて、種々の条件下で連続加 水分解実験を行い、モデルの妥当性を検討するとともに最適操作条件を探索する。 第4章では、本研究を総括する。第2章 回分反応システム
(2.1均相系における酵素加水分解実験
均相系における酵素加水分解実験を種々の条件下で回分反応器を用いて行
い、本酵素加水分解システムを表現するための速度論モデルを構築する。 2.1.1実験 一基質には、逐次的な分解経路が比較的簡単で分析が容易なデキストラン由来のイソマ ルトヘキサオース(6糖)を用いた.酵素には、 Penl'cl'111'um sp.由来のエンドデキスト ラナ-ゼを用いた。回分反応器には500cm3の三角フラスコを用い、反応液体境は100cm3 とした。均相酵素反応は、37℃の恒温槽中、毎分60往復の速度でフラスコを振畳させなが ら行った.初期基質濃度および遊離酵素濃度を変化させた。反応条件をTable 2-1に示す. 反応は所定濃度の基質溶液に所定量の酵素溶液を滴下することにより開始した。サンプリ ングは適当な時間間隔で1.0cm3の反応液をマイクロピペットを用いて採取することによ り行った。採取したサンプルは0.02N NaOH 0.5cm3を加えて速やかに反応を停止させた後、 0.02N HCI 0.5cm3を加えて中和した。さらに、サンプルから酵素を取り除くため、分画分子 量が10000である微量試料用限外波過膜(日本ミリポア(秩)モルカットⅡ)を用いて波過し た。ただし、この限外波過膜には保存剤としてグリセリンが含まれており、糖濃度分析の際 に不純物として検出されるため、予め一晩超純水に浸すことによりグリセリンを取り除い た。 分析は、埴過後のサンプルの各糖濃度を、示差屈折計を検出器とした分配吸着式高速 液体クロマトグラフィー(HPLC)を用いて測定した。Table 2-2と2-3に分析装置の仕様と分 析条件を示す。Table 2ll Reac'tion conditionsinsuspended enzyme system・
反応温度 : 37℃ 反応液体積 : 100 cm3
初期基質濃度 : 1.3, 1.7 × 104mol・dm3lliq
遊離酵素濃度 : 1.16, 2.1 mg・dni3lliq
Table 2-2 Specification of analyzer・
示差屈折計 :島村製作所(秩) YRU-880mdget
インテグレータ- :日立製作所(秩) D-7500
オートサンプラ- :日立製作所(秩)し7200 インテリジェントポンプ:日立製作所(秩) L-7100
カラム :日立製作所(秩) NH-C18
Table 2-3Analysis conditions・ Ehent :超純水100%
Flow Rate : 1cm3・min-1 column Temp・ : 60℃
2.1.2速度論モデルの構築
エンドデキストラナ-ゼによるイソマルトヘキサオース(6糖)の加水分解反応機構を Fig.2-1に示す。図の黒丸はグルコース1つを、 Eは酵素を、k.・は反応速度定数(i=1,2,3)を表 す.デキストラナ∼ゼはエンド型活性を示すため、基質末端以外のα-1,6グリコシド結合 に対する加水分解反応を触媒する。従って、 6楯は図の点線の示す3通りの位置(Ⅱ, Ⅰ ,Ⅱ) で酵素によりα11,6グリコシド結合を切断され、 3糖あるいは2糖と4糖に分解され、さ らに4糖は2糖に分解されることになる。本加水分解反応に関して、基質が酵素と複合体 を形成して、生成物を生成するというMichaelis-Menten型の反応機構を考えている。 E;H20llH2HHH2○N M○N○HM○○N○HM○NNN○H○HN○N○MH○N○M読;:_・_.I_き
Ⅰ ⅡⅠⅡ @ klllk2[α( ㈹㌫⑳)
tk, ●-●-● 〇一〇一〇&"=@
01t胸
1 【Ⅶロ + J.rT
一し旦j ㊨-⑳ ⑳-⑳耳魯1診klJl k2
IJ-}_二
tk, ⑳-⑳ ㊨-⑳Fig・2-1 MichaelisIMenten type hydrolysis process of isomaltohexaose usmg endo-dextranase. Deanら11)は、本反応のような多糖類のエンド型酵素加水分解に対して数学モデルを構 築した。彼らのモデルでは、酵素が基質末端以外のどのα-1,6グリコシド結合も同じ確率で 切断するという仮定が用いられている11)。しかし、そのモデルでは実験結果との一致が得 られなかったことが報告されている。そこで本研究では、Fig.2-1に示すように酵素が6鹿 のⅠの位置でα-1,6グリコシド結合を切断する(3糖が生成する)確率をαとして考慮するこ とにより新たな速度論モデルを構築する。さらに、モデルの簡略化のために6糖および4 糖の反応速度定数k,,kbk,がそれぞれ等しいと仮定するo
これより、各糖の反応速度は(2-1)-(2-4)式のように表される。
V6 -些立ニー3klCs6CE 'k2CES6 dt
V4 -告- (1- α)k3CES6 -klCs4CE ・k2CES4
V, -鷲と- 2碑,CES6 V2 -警- (1-α)k3CES ・2k3CES. ( (211) (2-2) (2-3) (2-4) ここで、 Cslは重合度i(i=2,3,4,6)の糖濃度を、 cESEは基質と酵素の複合体を表す。 次に、 6糖と酵素、および4糖と酵素の各複合体に対して擬定常状態近似を適用すると、 (2-5)、(2-6)式が得られる。 dCES6 dt dCES4 dt
= 3k.Cs6CE -k2CES6 lk3CES6 = 0
= klCs4CE -k2CES4 lk3CES4 = 0
(2-1卜(2-4)式に(2-5), (2-6)式を代入して整理すると、 (2-7卜(2-10)式となる。 dCs6 _ -3Vn)axCs6 dt Km+3Cs6+Cs4 dCs. _ Vmaxl3(1- α)Cs6 - Cs.] dt K,. +3Cs6 +Cs4 dCs3 _ 6VznaxaCs6 dt Km+3Cs6+Cs4 dCs2 _ Vnax l3(1- α)Cs6 + 2Cs4 I dt Km +3Cs6 +Cs4 (2-5) (2-6) (2-7) (2-8) (2-9) (2-10)
ここで、 Km = (k2 +k,)/kl,Vznax =k,CE(0)であり、 cE(0) 【mg・dm 3-1iquid】は反応溶液中の初 期遊離酵素濃度である.各糖濃度cs` (i-2,3,4,6) lmol・dm 3-1iquid】の経時変化は実験的に求 まるため、本モデルの未知定数は最大反応速度vw lmol・dm 3-1iquid・min-1]、Michaelis定数Km 【mol・dm 3-1iquid】、6糖が3糖に分解される確率αの3つとなる。ここで、 α= 1βとした場合
の本モデル式は、 3つのα-1,6グリコシド結合の切断される確率が等しいとしたDeanらの モデル式11)と一致することになる。
2.1.3 結果と考察 2.1.3.1 モデルの適用 前節で構築した速度論モデルの未知定数はvn,.Lr、 RL、 aの3つである。ここで、 vwは 初期酵素濃度cE(0) 【mg・dm-3-1iq'uid】によって異なる値となるが、このVwをcE(0)で割っ た値、すなわち単位酵素量あたりの最大反応速度V,W'(=V,nw /cE(0))は酵素自身の固有の 値であり、一定であると考えられる.そこで、新たにvmA.'、 Kmおよびaを未知定数として、 実験値とのフィッティングを行った。 Fig.2-2に計算手順のフローチャートを示す。これ ら3つの未知定数は、 (2-ll)式で定義される各糖濃度の計算値と異なる3つの条件下での 実験値との相対誤差の二乗和である∫を最小にするようにS血plex法により推算された。 糖濃度の計算値は、連立微分方程式(2-7)∼(2110)式をRunge-Kutta法を用いて数値的に解く ことにより求めた。この際の時間刻みは60secとした。 ∫=∑ Csi,caEc - Csi,exp
No
2.1.3.2 適用結果 Fig.2-3に均相酵素反応での実験値と計算値を示す。プロットは実験値を、夷線は本モ デルによるフィッティング結果を表す。参考のために、 Dean ら11)のモデルによるフィッ ティング結果も点線で示す。実験結果のプロットから、どの条件においても反応時間の経 過とともに6糖の濃度は単調に減少しており、 3,2糖の濃度はそれぞれ単調に増加してい る。一方、 4糖の濃度は始め増加しているが途中で減少に転じており、生成した4糖がさ らに2糖にまで加水分解されていることがわかる。従って、このシステムには最終的に2 糖と3糖のみが存在しているo初期基質濃度をFig2-3(a)の1.7× 104mol・dm-3からFig.2-3(b) の1.3×104mol・dm13へと減少させた場合、 6糖の消費や2, 3糖の生成の傾向にあまり違い は見られない。一方、初期酵素濃度をFig.2-3(a)の2.1mg・dm 3-1iquidからFig2-3(C)の1.16mg・ dm 3-1iquidへと減少させた場合、 6糖の消費速度や2, 3糖の生成速度は、各糖濃度の経時 変化の傾きからわかるようにかなり小さくなっている。また、実線で示された本モデルに よるフィッティング結果は、 Deanら11)のモデルによるフィッティング結果と比べて、ど の条件下でも、実験値と良好に一致している。従って、基質のα-1,6グリコシド結合の切 断に関する酵素の選択性を考慮することにより、本酵素反応を表現することができたと考 えられる。
[C_ ∈p・l.∈PX]♂ [?∈p二。∈も妄】♂ 5 4 3 2 1 0 中" 耳耳耳耳耳耳耳耳耳耳耳爾リ耳耳耳耳耳自?ィ而 一一)U ∫ ′ ′ ′▲人▲▲▲ .._∠■ゝ__fj■●J=壬__ーん÷_I________ ..A.l..- 50YooV1Yoー200V25 0 50 100 150 Time lmin] 200 250
Fig・213 Experimentaland calculated resultsinsuspended enzyme system・ (a) C6(0)=1・7X IOl4 mol'dm-3, cE(0) =2.1mg・dm'3.
(b) C6(0)=1・3× 104 mol・dm 3, cE(0) =2.1mg・dml3. (C) C6(0)=1・7× 104 mol・dm'3, cE(0) =1.16mg・dm-3・
Table 2-4にフィッティングにより推算された各モデル定数値を示す。本モデルの新た な定数である6糖の分解により3糖が生成する確率αは0.411と推算された。この値は、 実験結果に基づき6糖および4糖が完全に分解された後の2糖に対する3糖の執合からも 求めることができる。各実験結果から算出したαの値もTable 2-4に示す. aの推算値は 実験値とほぼ一致しており、本モデルで新たに導入した酵素の選択性の概念が安当である と考えれる。また、この値は了3つのα-1,6グリコシド結合の切断される確率が等しいと したDeanらのモデル11)での値(α=1/3)よりも大きい値となっている。これはエンド活性を 有するデキストラナ-ゼは基質分子のより内側にあるα-1,6グリコシド結合に配位しやす いためであると考えられる。 また、 Table 2-5に以上のフィッティングによる各定数の推算値の妥当性を検討するた め一に行ったシミュレーション結果を示す。シミュレーションの方法は、各定数値を推算値 で一定とし、 1つの定数値のみを0.5-2.0倍に変化させた場合の実験値との相対誤差の二 乗和Sの値を求めた.定数値を変化させることで得られたSの値は最小値sm加よりも少な くとも16%以上大きい値となっている。これより、本研究でフィッティングにより推算 されたモデル定数値の信頼性は比較的高いものであると考えられる。
Table 214 Estlmated constantsinsuspended enzyme system.
constants estimated values experimental values
Km lmol●dm13-1iquid] 2.01 × 1014 * V.nax 'lmol●g-11enZyme・min-1】 2.90 × 10 3 *
a l-] 0.411 0.41-0.42 * uno bt ainable
Table 215 Sensitivity of model constantsinsuspended enzyme system.
constant s value S deviation
S - S,nin)偲,Jp X 100 +31.3% mmlmum 十16.7% Km 1.01 × 104 106.2 2.01 × 10`4 80.9 4.02 × 104 94.5 1.45 × 10 3 123.6 +52.7% Vznax ' 2.90 × 10-3 80.9 mlnlmum 5.80 X 10-3 160.2 +97.9% 0.206 101.8 +25.8% α 0.411 80.9 mmlmum O.822 173.4 114.3%
2.2 固定化系における酵素加水分解実験
固定化酵素を作成し、均相系と同様に回分反応器を用いて種々の条件下で酵素加水分解実験を行い、本固定化酵素加水分解システムを表現するための
速度論モデルを構築する。 2.2.1実験 2.2.1.1固定化静素の調製 基質および酵素は均相系と同様である。担体にはN-サクシルイミドエステルにより活 性化された多孔性メタクリル酸共重合体ビーズであるAffi-PreplO(バイオラット社製)を用′いた。この担体の公称の粒径は50pm、孔径はo・lpmであるo Affi-PreplOは、 Fig・214に示
すような共有結合反応により酵素のアミノ基と迅速に反応し、安定なアミド結合を形成す ることにより、酵素を固定化することができる。
Affi-PreplO Enzyme Immobilized Enzyme
弓gi;]. R-NH2急弓』R I HON.0(I;h]
Fig・2-4 ℃ovalently reaction of enzyme and support・
A臥PreplOへの酵素の固定化手順を以下に示す。 1) AfEi-PreplOはイソプロパノールで膨潤されているため、これを除去するため一晩超 純水に浸す。 2) 5・O gのAffi-PreplOを所定濃度(2.32 mg・cm-3, 1.6 mg・cm-3)の酵素溶液に加え、4℃で おだやかに12時間摸拝しながら共有結合反応させる。 3)酵素を固定化したAffi-PreplOを取り出し、超純水で洗浄した後、反応に用いる. 4)残った酵素溶液中の残存酵素濃度を595nmでの吸光度測定により求め、Affi-PreplO に固定化された酵素量を算出する。 2.2.1.2 加水分解実験 固定化酵素を用いた加水分解実験は、均相系の場合と同様に、回分反応器に500cm3の 三角フラスコを用い、反応液体穂を100cm3とし、 37℃の恒温槽中、毎分60往復の速度で 振塗させて行った。初期基質濃度、固定化担体量および担体あたりの酵素濃度を変化させ た。反応条件をTable 216に示す.予備実験として、振畳速度を変化させた条件下でも同 様に実験を行い、その振畳条件が加水分解反応速度に影響を与えないことが確認されてい る。反応は所定濃度の基質溶液に、所定量の固定化酵素を加えることにより開始した。サ ンプリングは適当な時間間隔で1.0cm3の反応液をマイクロピペットを用いて採取するこ とにより行った。採取したサンプルは0.02N NaOH 0.5cm3を加えて速やかに反応を停止さ
せた後、o・02N HCI O・5cm3を加えて中和し、さらに、酵素を取り除くため、分画分子量が10900
である微量試料用限外波過膜(日本ミリポア(秩)モルカットⅡ )を用いて波過した。この場合 にも、予め一晩超純水に浸すことによりグリセリンを取り除いた限外波過膜を用いた。そ して、各サンプル中の各糖濃度の分析をHPLCを用いて行った。分析条件も均相系と同様
Table 2-6 Reaction conditions in immobilized enzyme system. 反応温度 反応液体積 初期基質濃度 固定化担体量 酵素濃度 37℃ 100 cm-3 1・3-2・O X 104 mol・dm-3lliquid 1.0, 1.5 g 9・5, 13・5 mg ・ cm-3-particle 2.2.1.3 分配平衡実験 固定化酵素を用いて加水分解実験を行う際、液相中の糖濃度と固定化担体中の糖濃度と は、厳密には一致していないと考えられる。従って、液相と担体間の糖濃度の関係を知る ために、両者間の分配平衡実験を以下の手順で行った。実験条件をTable 2-7に示す. 1)所定濃度の基質溶液cs,(o)に所定量wpの酵素を固定化していない担体を加え、固定 化酵素反応と同様の条件下で恒温槽中で振塗する。 2)平衡に達した時点(24時間経過後)での溶液中の糖濃度cs,をHPLCにより分析する. 3)この溶液中の糖濃度に基づき、次式により担体中の糖濃度を求める。この際、実際 の担体は多孔性のポリマーであるが、これを均一なゲルであると仮定し、 Fig.2-5に 示すように担体中の糖濃度は一定であると考えている。 百- (Cs甥'p; pCsi ht (i-2・3・4,6, ここで、 V, ldm3-1iquid】は溶液体積、 p lg-particle・dm-3-particle]は担体密度であるo
Table 2-7 Experlmental conditions of distribution equilibrium experlment.
基質 反応温度 反応液体積 6糖、2糖 37℃ 50 cm3 初期基質濃度 1.5, 2.0 × 104 mol・dm-3-liquid 担体量 3.0, 4.0 g (2- 12)
Fig.2-5 Schematic皿Iustration of equilibrium saccharide concentration
inlmmobilized el1Zyme System.
2. 2. 2速度論モデルの構築 2.2.2. 1基礎式の導出 固定化酵素による糖類の加水分解に関する速度論モデルを構築するにあたり、モデルの 簡略化のため、以下のような仮定を施す。 1)液相は完全混合である。 2)担体粒子は均一かつ球形で、その半径は一定である。 3)酵素は担体内に均一に固定化されている。 4)反応器が十分に混合された状態であるため、外部物質移動抵抗は存在しない。 5)各糖濃度に関して、液相と担体表面の間には、 (2-13)式で与えられる線形の分配平衡 が成り立っている。 C-S・・し=Rp -HCs・・ (i-2・3・4・6) (2-13) ここで、 H ldm3lParticle・dm-3-1iquid】は分配平衡定数である。 6)担体内部の各糖の移動速度はFickの第一法則に従う。各糖の有効拡散係数D,FHは (2-14)式で与えられる。 D,g - A x (Mm・ )-B (i-2,3,4,6) (2-14) ここで、 Alm2・S-l], Bl-]は回帰係数である。 これらの仮定に基くと、担体近傍の糖濃度分布はFig.2-6のようになる。
Rp r
Fig.2-6 Saccharide concentration distribution h immobilized enzyme system.
まず、各糖に関する反応器内の物質収支は、仮定1卜3)より(2-15)式で、初期条件は(2-16) 式で表される。
誓Tcls..;;(Vi;=250;,-.,6, 'i=2'3'4'6'('22:1156;
ここで、古は全反応体積に占める反応溶液の割合であり、(2-17)式のように定義される。 6 - & (2-17) vLは反応溶液体積を、vp Cま担体体積を表すoまた、担体中の平均糖濃度(CTE)および平均 反応速度(可は(2-18)式、(2-19)式で定義されるo (CTE) (vT) -RJpeE.W 2d, 0 言mRp3 Rfpv74- 2dr 4_n3 7ZRp 3 (i=2,3,4,6) (2-18) (i=2,3,4, 6) (2- 1 9) 一方、各糖の担体内における反応速度V7 1ま2.1.2で構築した均相系の速度論モデルの(2-7)∼(2-10)式に基づき、担体体積基準の最大反応速度vLmaxm lmol・dm-3lpa,ticle・min-1】、担体体積
基準のMichaelis定数K悪m 1iquid]、および担体内の各糖濃度cs.・
lmol・dm-3-particle]を用いて(2-20)∼(2-23)式のように表わされるo - 3V.nT Cs6 V3 V2 KmLm'n + 3Cs6 + Cs4 vmr b(1 I αつを二年J Kmh + 3Cs6 +Cs4 6VEnaXST αb"m cs6 Kn,m +3Cs6 +Cs4 耳1. - αLmm )Cs6 + 2Cs4 KnIJm + 3Cs6 + Cs4 (2-20) (2-21) (2- 22) (2-23) ここで、 abml-]は固定化系における6糖が3糖に分解される確率である。 また、各糖に関する担体内の物質収支は、仮定6)に基づき、式(2-24)式で表される。 誓-DleH(;雪原)・vT (i-2・3・4,6, (2124, ここで、初期条件は(2-25)式、境界条件は仮定4), 5)より(2-26)、 (2-27)式で表される。
i.c O≦r≦R, ; CTm-o (i-2,3,4,6) (2125)
b・C・ r-0; %-0 (i-2・3・4,6) (2-26) r=Rp ; CsE = HCsE (i=2,3,4,6) (2-27) 以上の(2-15)∼(2-27)の基礎式を数値的に解くことにより、溶液中および担体中の各糖濃 度を計算することができる。 2.2.2.2無次元化 以下の無次元数を定義する。 (i=2,3,4, 6) (2-28) (t'=2,3,4, 6) (2129) (2-30)
(2-31) ( (i=2,3,4, 6) (2-32) (2-33) Rp 2vLmaxm (2-34) ここで、cs6 (0)は6糖の初期濃度、D6eHは6糖の有効拡散係数である. これらの変数を用いることにより、反応器内の物質収支式(2-15)式、および初期条件 (2-16)式は以下のように無次元化される。 算音響寺4(X7-0 (i-2,3,.,6, (2-35, i・C・ T-0 ; Zi - (i(ii'0='Zi,3:.6,' (2-36, 担体内の平均糖濃度(2-18)式および平均反応速度(2-19)式は以下のように無次元化され る。 1
(zT) - 3lzTR2dR
O 1 (xT) - 3JxT;R2dR O (i=2,3,4, 6) (2137) (i=2,3,4, 6) (2-38) また、担体内の微分物質収支式(2-24)は以下のように無次元化される。霊-芳(孟夏・雷)
+ Qx` (i=2,3,4,6) (2139) 初期条件(2-25)式および境界条件(2-26),(2-27)式は以下のように無次元化される。i・C・ T=0、0≦R≦1 ; Z,I(0)=0 (i=2,3,4,6) (2-40) b・C・ R-0・・獣O
R=1 ; Zi=月Zf
各糖の反応速度式(2-20)∼(2-23)は以下のように無次元化される.
(i=2,3,4,6) (2141)
X6 X4 X3 X2 - 3Z6 Z4+3Z6+K 3(1- αh)Z6 -Z4 Z4+3Z6+K 6αL" Z6 Z4+3Z6+K 3(1 - ahn')Z6 + 2Z. Z4 +3Z6 +K (2-43) (2-44) (2145) (2146)
これより、本モデルの未知定数はvLmaxm、 KiTm、 A、 B、H、αimmの6つとなる。ここで、
TILmaxmは均相系の場合と同様に担体あたりの酵素濃度cLmm(o) 6,=依存して変化する値である。 そのため、酵素固有の値である単位酵素量あたりの最大反応速度vLmaxm '(-vLmaT/cLmm(o)) lmol●glllenZyme・min-1]として計算に用いる。 2.2.3結果と考察 2.2.3.1分配平衡実験 Fig.2-7に平衡分配実験の結果を示す。横軸は液相中の各糖濃度、縦軸は担体中の各糖 濃度を示すo白抜きのプロットは担体量3・Og、黒抜きのプロットは4.0gの場合である。 液相の平衡濃度と担体中の平衡濃度の間には担体量や糖の初期濃度の違いとは関係なく、 ほぼ線形関係が成り立っていることがわかる。また、本システム中に存在する最も分子量 の大きい6糖と最も小さな2糖を用いた実験結果が同一直線上にあることもわかる。分配 係数Hは、これらのデータの傾きから最小自乗法により求めることができ、ここではH=0.95 となった。これより、本システムにおける分配係数は、 2-6糖では分子量によらず一定 であると考え、この値を計算に用いた。
[苫!Ved-?∈p・l.u'もLx] 2.5
2
0 0.5 1 1.5 2 2.5 3
Csi [× 10-4mol・dm 3-liq]
2.2.3.2 速度論モデルの適用
本固定化系では、前述した分配平衡実験により分配係数が求まるため、本モ≠ルの未知
定数はvLmaxm ,KBrnz,A ,B ,αimmの5つとなる。溶液中の糖濃度のみが実験的に求まるため、
均相系と同様に5つの異なる条件下で得られた溶液中の糖濃度の実験結果に対してモデル によるフィッティングを行った。 Fig.2-8に計算手順のフローチャートを示す。計算は、 モデル定数および各糖濃度の初期値を設定したのち、無次元化した担体内の物質収支式 (2-39)式をcrank-Nicolson法により離散化し、数値的に解き、担体内の各糖濃度zTを計算 する12)・13).次に、このZTを用いて、 (2-37), (2138), (2143) ∼(2-46)式より、担体内の平均糖 濃度(Z7および平均反応速度(嘉)を計算するoこれらの噂を用いて、反応器内の物質収支 式である(2-34)式を解くことにより、液相中の各糖濃度zfを計算する。そして、得られ た液相中の各糖濃度の計算値と実験値の相対誤差の二乗の和を(2-47)式より求め、それが 最小になるようなモデル定数、 vLmaxm',KBrm,A , B ,αimmをsimplex法により推算する.ま た、数値計算に用いた担体内の差分格子はR方向に50であり、時間刻みは18secである。 ∫=∑ Csi,calc - Csi,exp
2. 2. 3. 3 溶液中の各塘濃度の簾時変化 Fig.2-9(a)∼(e)に種々の条件下で得られた溶液中の各糖濃度の経時変化を示す。 (プロット は実験値であり、太線は本モデルによる計算値である。破線は、参考のために行った担体 内の物質移動抵抗を考慮しないモデル(Appendix参照)による計算結果である。 (a)∼(C) は、初期基質濃度を1.3-2.0× rO4mol・dm・3と変化させた結果であり、各糖の消費や生成の 傾向には大きな違いは見られなかった。これに対し、 (b)と(d)は固定化担体量を1.0-1.5g と変化させた結果であり、固定化担体量を増加させることにより各糖の消費や生成の速度 も大きく増加していることがわかる。また、 (b)と(ら)は担体あたりの酵素濃度を 13.5-9.51mg・cm13-particleと変化させた結果である。このように酵素濃度を減少させても、 各糖の消費や生成の傾向には大きな違いは見られなかった。これらの結果から、糖の消費 や生成に最も大きな影響を与える因子は固定化担体量であると考えられる。一方、破線で 示した担体内の物質移動抵抗を考慮しないモデルでは、どの条件においても計算値は実験 値の大まかな傾向を示してはいるが、あまり一致していない。特に4糖の実験値は計算値 よりもかなり小さくなっており、これは担体内の物質移動抵抗が存在するため、担体内で 生じた4糖が溶液中に出てくる前に素早く2糖に分解されてしまうことが原因であると考 えられる。これに対し、物質移動抵抗を考慮した本モデルによる計算値は、どの条件にお いても実験値と良好に一致しており、種々の条件下での固定化酵素反応を良好に表現する ことができると考えられる。
[cI LLJP・l.uもLx]duOOOPでeL499t2S [?uJP・l。uJもLx]+。U。C)。PてeL49Ot?S 2 1 0 3 2 1 0 5432-0 [cO ∈P・).∈もLx]du。OOPてttLJOOt2S 0 50 1 00 1 50 200 250 Time lmin] 0 50 1 00 1 50 200 250 Time lmin] 100 150 Time lmin] 200 250 543210 【C. LLJP・B.∈もLx]'9U.OaPでeL499eS ∩?uJP・]。uJもLx].9U.OaPでt!LJ99t2S 0 50 100 1 50 200 250 Time lmin] 200 250 0 50 100 150 Time lmin】
Fig.2-9 Experimental and calculated resultsin血mobilized enzyme system・
(a)cs6(0) -1.3 × 104 mol・dml3, W,-1・O g, CBh (o) -13・86mg●cm-3-particle (b)Cs6 (0) -1.7 × 104 mol・dml3, W,-1・O g, CEbw (o) -13・86mg●cm 3-particle (C)cs6(0) -2.0 × 104 mol・dm 3, W,-1・O g, CBhm (o) -13・86mg'cm-3-particle
(d)Cs6(0) -1.7 × 104 mol・dml3, W,-1・5 g, CEh(o) -13・86mg'cm 3particle (e)cs6(0) -1.7 × 104 mol・dmー3, W,-1・O g, C㌢t (0) - 9・51mg●cm13-particle
2. 2. 3. 4 担体内の各糖温度分布と反応速度分布
(
Fig. 2110(a)∼(C), (a)'-(C)'に担体内の半径方向の各糖濃度分布を示す.これは、初期基質 濃度1.7×104 mol・dm 3、担体量1.0 gの条件下における、反応開始後、 10, 60, 200分後の 担体内の各糖濃度分布である。 (a)∼(C)は担体あたりの酵素濃度13.86mg・cm-3-particleとし た場合、 (a)'∼(C)'は担体あたりの酵素濃度9.51mg・cm-3-particleとした場合である。グラフ の横軸は、担体中心から半径方向への距離である。反応開始後10 mhであるFig.2-10(a)に おいて、全ての糖濃度勾配が担体表面近傍にのみ存在していることがわかる。そして、反 応時間が経過するにつれて、各糖の濃度勾配が小さくなり、反応開始後200 minである Fig.2-10(C)においては、濃度勾配はほとんど消滅して、各糖濃度は担体内のどの場所でも ほぼ一定となっていることがわかる。酵素濃度を減少させたFig.2-10 (a),-(C),の場合では、
Fig・2110 (C)'でわずかに担体表面近傍に濃度勾配が存在しているが、 Fig.2110 (a)∼(C)とほぼ 同様の傾向がみられる.次に、 Fig.2-ll(a)∼(C),(a)'-(C)'に同条件下での担体内の半径方向の 反応速度分布の計算結果を示す。グラフの横軸は前図と同様に担体中心から半径方向への 距離であり、縦軸は反応速度の大きさである。反応速度の値が負の場合、糖が反応によっ て消費されていることを、正の場合は生成していることをそれぞれ意味している。反応開 始後10 minであるFig.2-ll(a)において、反応速度は、濃度分布と同様に担体表面近傍にお いて大きな値を持っていることから、反応が担体表面近傍でのみ起こっていることがわか る。また、4糖の反応速度はFig.2-ll(a)においては正の値をとっているが、反応開始後60 mh であるFig.2-ll(b)においては負の値をとっており、生成から消費に移行したこともわかる。 各糖の反応速度は反応時間が経過するにつれて減少し、反応開始後200 minであるFig.2-11(C)においては、ほぼoとなっていることから、全ての反応が終了していると考えられる. 担体あたりの酵素濃度を減少させたFig.2-ll (a)'∼(C)'の場合でもこの傾向は同様であるが、 担体内のより内側の部分でも反応が起こっていることがわかる。このように速度論モデル を構築することにより、観察することが困難な担体内の糖濃度分布や反応速度分布に関す る情報を得ることができる。また、これらの情報から、本固定化酵素反応システムでは、 担体表面近傍に固定化された酵素のみが、糖の加水分解反応に対して有効に働いていると 考えられる。これは、この系での糖の加水分解速度が担体内での糖の拡散速度よりも著し く大きいためであると考えられる。
[op!vt2d-C・∈p・LOuJ昌ニス].OuOOaP!JeLPOE!S PPPJedA・ufP・lOub占Lx].DuOOaP!Lt2LJ8eSPP!VedJ・LuP・IOu>占tx]+OuOOaP!JeLPOeS 5 4 3 2 1 (b)t=60lm 末袵
iiiiiZI
______.__.!_.1=_& (C)t=200[min] 5 10 15 r[〟m] 5432-0543210 [むPモedi・∈P・lOuP占Lx]'OuOOGIPてt!LJ00t!S[〇一ひでed・C・Ep・lOき占Lx]duOOOPてeLJ99eS 543210 【opでedJ・∈P・lOub占Lx].9uOOaP!JeLJDDeS ( (a)ーt=10[min] ____.●■..-)._./_.I.JT一●- D510152025 (b)'t=60lmin]iiiiiiia
._____".一...i._ここiヲ一 (¢)'t=200[min] 0510152025Fig.2-10 Calculated distribution of saccharide concentrationinsupport particle・ Cs6(0) -1・7 × 104 mol●dm-3, wp=1・O g
[LJJ!uJ・。P!Ved-C・up・一〇∈昌Lx]F L -1 ) 2 1 0 1 2 0 ll [Lトup・Ol9nJedi・uP・l.u昌Lx】F 2 1 0 1 2 ll 【L・u!Lu,。J。モedi・∈P・10∈昌Lx]F 0 5 (b)t=60lm 末袵 _一′ ーr-l'Lq、..1 (C)t=200[min] 0510rl〟m]15 25 2 1 0 1 2 lll ∩LL u!u"一oモed-C_ uJP・J.∈もLX]i 【 llll (a)■t=10[min]l I ■ ■ i-■ --■■■ -- FL-.Lir - ヽ 一 ヽ I 一 ● t ll.ll 0 2 1 0 1 2 l】 [TuPJ・ePrUed-C・uJp・[o∈>・oLx]け 2 1 0 1 2 ll [L・u!uJ・〇一〇でed-?∈p・10∈昌Lx]A (b)'t=60[min】 ′ _-ノ●● 一一-Il'l'I>--..-.1' ヽ (C).t=200[min] 0510152025
Fig・211l Calculated distribution of hydrolysis rateinsupport particle・
cs6(0) -1・7 × 104 mol・dm-3, wp-1・O g
2.2.3.5 モデル定数の推算値 本モデルのフィッティングにより推算された各定数値をTable 2_8に示す.血chaelis定 数Kn,bmの推算値は、固定化酵素を用いた種々の反応に関して報告されている文献値と等 しいオーダーとなっている6)。また、本固定化系における速度定数、 K㌘、 VnT,は均相 系で推算された値よりもかなり大きな値となっている。この傾向は、他の研究者たちによ っても報告されており、一般的に見られるものである14)。本モデルでは、担体内に酵素が 均一に固定化されていると仮定している。しかし、実際には、担体表面近傍の酵素濃度が 担体中心よりもかなり高くなっているという報告がある3)・15)・16)。このような場合、加水分 解に効果的に関与している酵素の総量が本モデルで考えるよりもかなり多くなると考えら れ、その場合の速度定数は本モデルにより推算された値よりも小さな値になると推測され る。このことが、本研究で得られた速度定数の推算値が均相系と固定化系で異なった理由 の一つと考えられる。また、均相系と固定化系では微視的な環境や酵素の立体構造が変化 すると考えられるため、酵素活性も変化していると推測され、このことも理由の一つとし て考えられる. α加の推算値は、固定化系でも実験値とほぼ等しくなっているが、均相系 の値よりも若干大きな値となっている。この理由については明らかではないが、固定化に よる酵素の立体構造の変化が関係していると考えられる。
Table 2-8 Estimated constantsinlmmobilized enzyme system・
constant s estimated values experimental values
K㌘ 【mol・ dm-3-particle] 8.67 × 10-2 VnT '【mol・g-1-enzyme・min111 5.86 × 10-2 A lm2・S-1] 1.72X 10-10 月H o.808 αbw H 0.455 辛 unobta 加able
Table2-9に本モデルによる担体内の有効拡散係数DieHの推算値を示す。参考のために、 Nakaoらが報告した経験式(2-48)式17)より計算した水溶液中での拡散係数の値も示す. DieH =8.76×10 9(M訂o・48 (i-2,3,4,6) ! (2-48) 固定化系に関する多くの研究において、担体内の基質や生成物のDieHの値は水溶液中の DieHとほぼ等しいと仮定されている18)-20).しかし、本研究でフィッティングにより得られ た担体内のDleHの値は、水溶液中の値よりも数オーダー小さな値となった.一般的に、こ の傾向はイオン交換樹脂などの高分子媒体中でよく観察される21)。本研究で用いた担体粒 子は、分配平衡定数〝がほぼ1に等しいことから、水で十分に膨潤している多孔性物質 であると考えられるが、担体内の拡散は、粒子内の綱目構造による立体障害により制限さ れるため、水溶液中の拡散係数の値よりも小さな値となったと考えられる。 -また、 Table2-10にフィッティングによる各定数の推算値の安当性を検討するために行 ったシミュレーション結果を示すo シミュレーションの方法は、前述した均相系の場合と 同様である。定数値を推算値の0.5-2.0倍まで変化させることで得られた∫の値は、最小 値smhよりも少なくとも6%以上大きい値となっているo従って、本研究で推算されたモ デル定数値の信頼性は、均相系と同様に比較的高いものであると考えられる。
Table 2-9 Estimated values of effective diffusivitiesinSupport particle and calculated values
from empirical equation reported byKimura and Nakao17)inaqueous solution・
effective diffuSivities lm2・S-1] insupport particle inaqueous solution
好6 54 好3 好2 8 8 8 か 6.53 × 10-13 8.99 × 10 13 ll.3 × 10-13 15.4 × 10-13 3.19 × 10-10 3.86 × 10 10 4.42 × 10 10 5.32 × 10 10
Table 2-10 Sensitivity of model constantsinlmmobilized enzyme system・ l
const ants value S deviation
(S - Smin )/ Smか 4.33 × 10 2 196.7 K㌘ 8.67 × 1012 180.8 17.3 X 10-2 196.0 8.8% mmlmum 8.4% VznT 2.93 × 10 2 196.1 5.86 × 10 2 180.8 ll.7 × 10 2 196.8 8.5% mmlmum . 8.8% 0.86 × 10 10 * * 1.72 × 10 10 180.8 mmlmum 3.44 X 10-10 191.6 6.0% 0.404 307.2 69.9% 0.808 180.8 mmlmum 1.62 * * αLM 0.228 220. 1 21.7% 0.445 1 80. 8 mmlmum 0.911 391.8 116.7% * calculation impossible
2. 2. 3. 6反応速度に及ぼす各種操作国子の影苧 2.2.2で構築した固定化酵素システムに関する速度論モデルの安当性が実験的た示され たため、このモデルによりシミュレーションを行い、加水分解速度に及ぼす各種操作因子 の影響を検討する。シミュレーションには、前節のTable 218に示したフィッティングに よって推算したモデル定数の値を用いた。シミュレーションを行う条件をTable 2_11に示 す。 caselからcase3の各条件は、初期基質濃度、反応系内の総酵素量を一定としている.
Table 2-llList ofo erational factors for simulation.
基質濃度 酵素濃度 担体量担体粒径総表面墳総酵素量 lmol'dm13-1iq・] lmg・cm 3-particle] lg] lpm] lm2] lmg] easel 1.7 × 104 13.86 1.0 50 0.12 12.08 case2 1.7 × 104 6.93 2.0 50 0.24 12.08 case3 1.7 X IOl4 13.86 1.0 25 0.24 12.08 Fig・2-12(a) ∼ (C)に液相中の各糖濃度の経時変化に関するシミュレーション結果を、 Fig.2-13(all)イa-3), (b-1)∼(b-3), (C-1)∼(C-3)に担体内の半径方向の糖濃度分布に関するシミュレー
ション結果を示す. Fig・ 2-12(a)イC)で6糖の消費に注目した場合、 (a)すなわちcaselでは
150min, (b)すなわちcase2では100min, (C)すなわちcase3では70min程度でほとんど完了
していることがわかる。 2,3糖の生成に関しても、 (a)では250minでもまだ若干生成傾向 が見られるが、 (りでは250minで、 (C)では200min程度で、ほぼ生成が完了していること がわかる。 caselと比較して総表面積が2倍であるcas62では、各糖濃度の変化が著しい。 このことから、担体の絵表面積が大きいほど反応速度が大きいと考えられる。これは、反 応が担体近傍でのみ起こっているため、総表面積が大きいほど関与する酵素量が多くなり 反応速度が大きくなると考えられる。 case2と比較して担体の総表面積が等しく担体粒径 が1/2倍であるcas63では、各糖濃度がcas62よりもさらに速く変化している。この理由 をFig・2-13(a)∼(C)に示す担体内の各糖濃度分布で説明する. (b)すなわちcase2では担体中 の糖濃度分布が反応開始10分後では担体表面近傍にのみ勾配が存在しているのに対し、(C) すなわちcase3では担体中心付近まで濃度勾配が存在していることがわかるo担体粒径を 小さくした場合、 (2-34)式で定義されるThiele数が担体半径の2乗に比例するため小さく なり、より担体内部にまで各糖が拡散し、担体内部に固定化された酵素も反応に関与する
ことができる。このため、反応系内の総酵素量が等しくても粒径を小さくした場合に反応 速度が大きくなったと考えられる。 ( 5432-0. FLLJP・(。LLlもLx】.9uOOOPTeLJ8t!S [C・uJP・IO∈●・OLx].9uOOOP!JeLJ00eS 5 4 3 2 1 0 50 1 00 1 50 200 Timo [min] (ら) 一′ ヽノ ヽ ''-、ヾ.:_./_:__._,.一.___.__ 1 00 1 50 200 250 Time lmin] 【C・uJp・lOLLJ昌Lx].OuOOOPてtPL18eS 5 4 3 2
Fig・2-12 Calculated results inlmmobilized enzyme system・
(a)easel ; CBh (o) =13・86 mg・cm-3-particle, Wp-1・O g, Dp=50 pm・ (b)case2 ; CEh (o) -6・93 mg・cml3-particle, Wp=2・O g, Dp=50 pm・ (C)case3 ; CEh(o) -13・86 mg・cm-3-particle, Wp-1・O g, Dp-25 pm.
case 1 case2 case3 [opで邑IC・∈P・tOuJ昌Lx]'OuOOOPてeLJ0PeS[。Pモed・?up・l〇uJ>・OLx】.OuOOGIP!LeLPOeS 5 4 3 2 1 0 (a-2)t=60lmin] 0510152025 0 543210 [〇一〇!VedTe・uJP・lOE寸占Lx]'OuOOaPてt2LJ00eS (a-3)t=200【min] 5 2 0 2 15rf 〃 o[ 1 r 5 (b-1)t=10[min] ′ __=‥..._i..:).- ' 510152025 15 20 25 5 10 1 5rf 〟 I.; 5 2 4 6 8 10 12 2 4 6 8 10 12 r[〟m]
Fig.2-13 Calculated distribution of saccharide concentration in support particle・
(a) easel ;CEiw(o) -13・86 mg●cml3-particle, Wp=1・O g, Dp=50 pm・ (b) case2 ; CEb"m (o) =6・93 mg●cml31Particle, Wp=2・O g, Dp=50 pm・ (C) case3 ; CEb"m (o) =13・86 mg●cml3-particle, Wp-1・O g, Dp-25 pm・
第3章 連続反応システム
( 前章で構築した回分反応システムに関する速度論モデルを拡張することにより連続加水 分解反応に関する速度論モデルを構築する。そして、モデルによるシミュレーションに基 づき、前章で調製した固定化酵素を充填するカラム型リアクターを設計、製作するととも に、このリアクターを用いて種々の条件下で連続加水分解実験を行い、最適操作条件の検 討を行う。3. 1速度論モデル
3. 1. 1基礎式の導出 連続反応システムに関する速度論モデルを構築するにあたり、モデルの簡略化のため、 以下のように仮定する。 1)担体粒子は均一かつ球形で、その半径は一定である。 2)酵素は担体内に均一に固定化されている。 3)反応器の半径方向の濃度分布は存在しない。 4)外部物質移動抵抗は存在しない。 5)各糖濃度に関して、液相と担体表面の間には(3-1)式で与えられる線形の平衡関係が 成立している。 clsiIFRp -HCsi (i-2・3・4,6) (3-1) ここでHは分配平衡定数である. 6)担体内部の各糖の移動速度はFickの第-法則に従う。各糖の有効拡散係数は分子量 Mwiに基づき(312)式で与えられる。DieH = A x (Mwi) B (i=2,3,4, 6) (3-2)
反応器内の各糖の物質収支は、 (3-3)式で与えられる。
ここで、左辺は非定常項であり、右辺の各項はそれぞれ分散、対流および反応による消失・ 生成を表す。 E lm2・S-1】は反応器内における分散係数であり、 (3-4)式に示される経験式22)よ り求められる. ( E = 2uRp (3-4) ここでRp lm]は担体粒子半径である。 u lm・sll】は線流速であるo 初期条件は(315),(316)式で与えられる。境界条件は入口側をステップ入力として(317)式、 出口側を閉鎖系の境界条件に基づき(3-8)式で与えられる。 i・C・ t=0 0くだ≦L;Csi=0 X=0; Cs.・ b.C. X=0; Csi i く Cs6(0)(i = 6) 0(i = 2,3,4) Cs6(0)(i = 6) 0(i - 2,3,4)
X-L・・% - 0
(i=2,3,4,6) (3-5) (3-6) (3-7) (i--2,3 ,4,6) (3-8) ここで、 Llm]はカラム長さである. また、担体内の平均糖濃度(CTE)および平均反応速度(vT)は回分反応と同様に、 (319),(3-10) 式で定義される。 (CTE) -RfpcTt4- 2dr (i=2,3,4, 6) (3 -9)・vT,婁sRfpvTr2dr (i-2・3・4,6, (3-10,
-万、各糖の担体内における反応速度V71ま、 2.2.2と同様に均相系の速度論モデルであ る(2-7)∼(2110)に基づき、担体体積基準の最大反応速度vLmaxm lmol・dml3lParticle・min-1ト担体体積基準のMichaelis定数KBrm lmol・dm'3-pa,ticle]および担体内の各糖濃度esL
lmol・dm-3-particle]を用いて(3-ll)∼(3-14)式のように表わされる. V6 V4 - 3V.nT Cs6 K,.'mn'+ 3Cs6 + Cs4 vmr b(1 I αhJC7jE] Kn,imm + 3Cs6 + Cs4 (3-ll) (3-12)
V3 V2 6VTnTαzmnz Cs6 KmLm +3Cs6 +Cs4 堤桓トαhlCTEL2CTEJ Kmzmnl + 3Cs6 + Cs4 (3-13) (3-14) 担体内の各糖の物質収支は、回分システムと同様に仮定6)より(3-15)式で表される。 誓-Died(各雷)・vT (i-2,3,4,6, ( ここで、初期条件は(3-16)式、境界条件は仮定4), 5)より(3-17), (3-18)式で与えられる。 3.1.2無次元化 以下の無次元数を定義する。 + vi (i=2,3,4,6) (3-15) i・c t=0 0≦r≦R,; CsE(0)=O b・C・ r-0 ;%-0 r=Rp ; Cs.・ = HCs.I Csi Z・ = csI6'('o) - Csi zl = -C5-6'('。)
X-i
・-gt
AL等
pe=些 か㌘ Kmh K = cs61(0) (i=2,3,4,6) (3-16) (i=2,3,4,6) (3-17) (i=2,3,4,6) (3- 1 8) (i=2,3,4,6) (3-19) (i=2,3,4, 6) (3-20) (3-21) (3-22) (3-23) (3-24) (3-25)(i=2,3,4,6) (3-26) ( (3-27) (3-28) (3-29) ここで、C∫6(0)は6糖の初期濃度、8㌢は6糖の有効拡散係数であるo これらの変数を用いることにより、反応器内の物質収支式(3-3)、初期および境界条件(3-5) - (3-8)式は以下のように無次元化される。 算音響- A2揺IAPe%・TQ(i) 'i-2,3,4,6''3-30' i.C. T=0 0く方≦1;Zi=0 X=0; Zi = b.C. X=0;Zi = i i 1(i = 6) 0(i = 2,3,4) 1(i = 6) 0(i = 2,3,4)
X-1;獣0
(i=2,3,4,6) (3-31) (3-32) (3-33) (i=2,3,4,6) (3-34) 担体内の平均糖濃度(3-9)式および平均反応速度(3-10)式は以下のように無次元化される。 1 (zT;) - 3IzIR2dR O I(i) - 3IiR2dR
0 また、担体内の物質収支式(3-15)は以下のように無次元化される。 (i=2,3,4,6) (3135) (i=2,3,4,6) (3 -36)霊歌孟夏・雷〕・¢嘉(i-2・3・4・6, (3-,7,
初期条件および境界条件(3-16)∼(3-18)式は以下のように無次元化される。 i.C. T=0、0≦R≦1 ; Zi(0)=0 (i=2,3,4,6) (3-38)b・C・ R-0 ・獣0 (i-2,3,4,6) R=1 ; Zi = HZl (i=2,3,4,6) 各糖の反応速度式(3-ll)∼(3-14)は以下のように無次元化される. X6 X4 X3 X2 - 3Z6 Z4 +3Z6+K 3(1 I αimnr )Z6 - Z. Z4+3Z6+K 6αimm z6 Z4 +3Z6 +K 耳11 αbnm)Z6 + 2Z. Z4 +3Z6 +K (3-41) (3 -42) (3-43) (3-44)
3.1.3計算方法
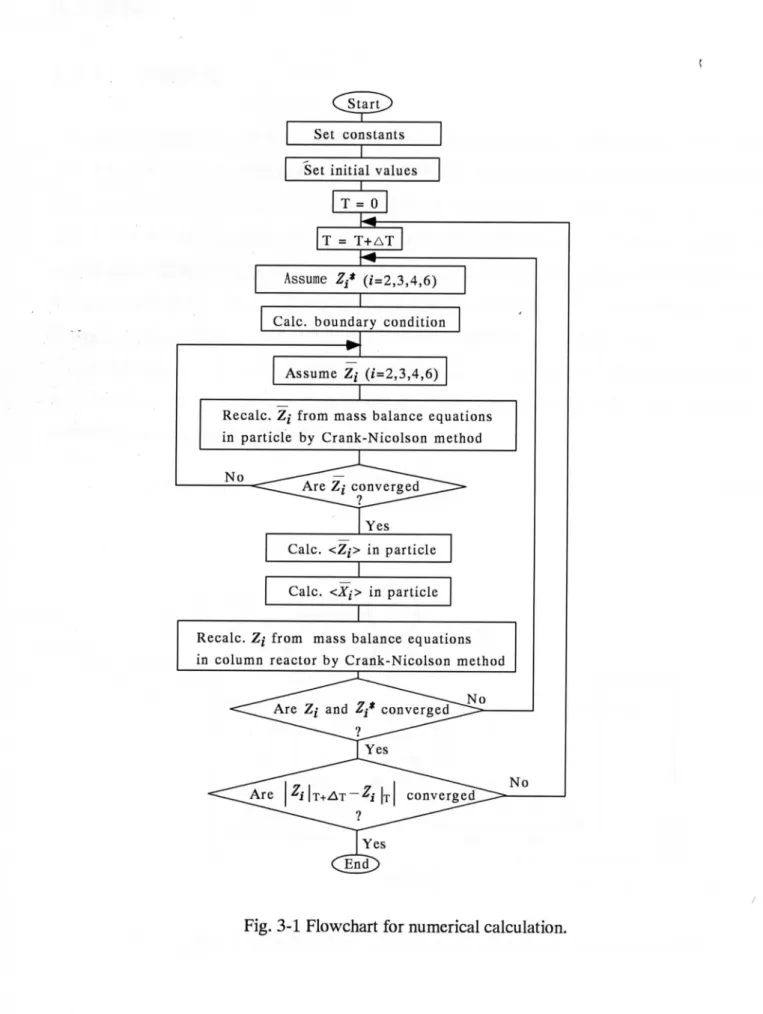
計算に用いたモデル定数値をTable 313に示す。これらの値は2.2.3で述べた向分反応シ
ステムでのフィッティングにより推算されたものである。
Table 3-3List of constantsfor numerical calculation.
Constant s Valu e K㌘ 【mol・dm 3-particle] 8.67 × 1012 VznT '【mol・g-1-enzyme・min-1] 5.86 × 10 2 A lm2・sl1] 1.72X 10-10 β日 0.808 αbw l-] 0.455 計算手順のフローチャートをFig.3-1に示す。計算は、モデル定数および各糖濃度の初 期値を設定したのち、無次元化した担体内の物質収支式(3-37)をGank-Nicolson法により 離散化し、数値的に解き、担体内の各糖濃度zTを計算する。次に、このZTを用いて、 (3-35), (3136), (3-41) ∼ (3-44)式より、担体内の平均糖濃度(Z7、および平均反応速度(司を 計算する。これらの値を用いて、反応器内の物質収支式である(3-30)式をcrank-Nicolson 法により離散化して数値的に解くことにより、液相中の各糖濃度ziを計算する12)・13).計算 に用いた差分格子数は担体内はR方向に50、 X方向に1000、時間刻みは7.2 secとした。 連続反応システムでは、定常解のみを求めればよいが、本モデルでは、計算を安定に進め るために(3-30)式による非定常計算を行い、漸近定常解を求める手法を用いている。
3.2実験
3.2.1 実験装置 Fig.3-2に連続反応システムの概略図を示す。本システムは、試料供給部、カラム型リ アクター、サンプリング部および恒温槽からなる。 500cm3の三角フラスコ中の基質溶液 はチュービングポンプにより、一定流量でカラム型リアクターへ供給した。フラスコ、お よびリアクターは一定温度に保たれた恒温槽内(37℃)に設置した。リアクターに供給され る基質溶液の温度が一定温度(37℃)になるように余熱部を設けた。用いた実験装置の仕様 をTable3-4に示す。カラム型リアクターは市販のものを即、、サイズは内径(Ld);5mm、 長さ(Lc) ; 10cmであるoこのサイズは前節で述べた速度論モデルを用いたシミュレーショ ンにより決定した。カラムに充填する固定化酵素は、 2.2.1.1で述べた方法で同様に作成し たものである。また、カラムへの固定化酵素の充填は自然沈降により行った。この結果、 空隙率を(3-45)式に基づき計算すると8 =0.40になった.8=1_a
(L5 /4)Le 恒温槽 サンプリング部Fig. 3-2 Schematic illustraion of column reactor system.
Table 3-4 Specification of column and tube pump・ カラム :柴田科学(秩) 高圧ガラスカラムGC型(内径:5rrm、長さ: 10cm、) チュービングポンプ: EYELA(秩) MICROmBEPUMPMP-3 3. 2. 2連続加水分解実験 前述した装置を用いて、 6糖の連続加水分解実験を行った。反応は、供給液の初期基質 濃度および担体あたりの酵素濃度を変化させ、各々の条件下で流量を1.0-9.2 cm3・min 1ま で変化させて行った。また、活性の変化についても検討するため、全ての供給液流量条件 での実験が終了した後、 1回目と同じ固定化酵素を充填したカラムを用いて、同条件下で 2回目の実験を繰り返し行った.反応条件をTable 3-5に示す.また、実験手順を以下に示 す。 1)所定量の固定化酵素をカラム型リアクターに充填するとともに、十分に脱気した基質 溶液および超純水それぞれ300cm3程度用意し、恒温槽中に設置する。 2)リアクターや流路中の気泡を除くため、超純水をチュービングポンプにより1時間程 度流した後、流量を実験での最も大きい値に設置する。 3)ポンプによる供給液を超純水から基質溶液に変えることにより反応を開始させ、所定 の時間間隔で3.0cm3ずつリアクターからの流出溶液をサンプリングする。 4)所定時間経過後、再び供給液を超純水に変え、リアクター内に残っている糖を洗い流 すため流量3.0cm3・min-1で1時間程度流す。 5)ポンプを次の実験流量に合わせた後、再び供給液を基質溶液に変え、反応を開始させ、 サンプリングを行う。 6) 4), 5)の操作を繰り返して、全ての流量条件下で反応を行う。 採取したサンプルは、回分反応の場合と同様に、微量試料用限外波過膜を用いて波過し た。ただし、連続反応の場合は、担体からの酵素の漏出によるサンプル中への固定化酵素
の混入がないことが確認されているため、反応停止などの操作は行わなかった。サンプル 中の各糖濃度の分析は、 HPLCを用いて測定した。分析条件は回分反応系と同様である。
また、申ラム型リアクターに充填する固定化酵素は、供給液中の初期基質濃度や酸素濃度 条件を変えるごとに、新たに調製したものを用いた。
Table 3-5 Reaction conditionsincolumn reactor system・
初期基質濃度 1.0, 2.0, 3.0× 104 lmol・dml3] 酵素濃度 9.5, 13.5 【mg ・ cm 3-particle】 供給液流量 1.0, 3.0, 5.0, 9.2 [9m3・minll】
3. 3結果と考察
3.3.1リアクターからの流出液中の各糖濃度
Fig.3-3(a)∼(d)に、リアクターに基質を供給し始めた時点をoとした、各サンプリング時 間での流出液中の各糖濃度をプロットで示す。白抜きのプロットは1回目の実験結果であ り、黒抜きのプロットは2回目の実験結果である。供給液の初期基質濃度は2.0×104mol・ dm-3、酵素濃度を13.5mg・cm 3-particleとし、供給液流量は9.2, 5.0, 3.0, 1.0 cm3・min11と変化 させたものである。どの条件においても、若干のばらつきはあるが、定常状態に達してい ることがわかる。また、 1回目と2回目の結果がほぼ一致していることから、これら2回 の実験での酵素の活性に低下はなく、本固定化酵素は繰り返しの利用が可能であると考え られる。 FuJP・l.LLJもLx].9LJOOOPTt!tJ00tPS FLLJPこ.LLJもLx]dLJ.O。PてetJ00eS 5 4 5 4 1 0 1 00 200 300 400 500Sampling time lsec]
墓 皇 皇
0 200
Sampling timo 【sec]
800 FLLJP・l。uJもLx]duOOOPTt?LJ00tPS FuJP・I.uJもLx]duOOOPTE!LJ00eS 5 4 3 2 1 0
Hii
0 200 400 600 800 1 000 Sampling timo [$○¢] ● ● ㊤ 8 皇 皇 △ ▲ 500Sampling time lsoc]
Fig・ 3-3 The saccharide concentrationinefnuent solution at each sampling tlme・ (a)Q=9・2 cm3'min11, (b)Q=5・O cm3・min-1, (C)Q-3.0 cm3・min-1, (d)Q=1.0 cm3・min-1
Fig.3-4に同条件下でのリアクター内の管軸方向の各糖濃度分布を示す。グラフの横軸 はカラム入口から出口方向への距離である。プロットはリアクター出口における実験値で あり、ラインはモデルによるシミュレーション結果である。(a卜(d)は供給液流量を9.2,5.0, 3.0, 1.0 cm3・min'1と種々変化させた場合であり、リアクター内の滞在時間が長くなること に対応する。どの流量条件下においても、リアクター出口でのプロットとラインはほぼ一 致しており、 3.1で構築した速度論モデルにより、本連続反応システムを表現することが できると考えられる。また、実験ではリアクター出口の溶液中の糖濃度しか測定すること ができないが、このようなモデルを構築することにより、リアクター内の糖濃度分布に関 する情報を得ることができる。これより、供給溶液中の6糖がリアクター内を進むにつれ て消費され、代わりに4, 3, 2糖が生成し、 4糖はさらに分解され2糖になるという様子を 管の長さ方向に見ることができる。 Fup・l.LLJTOLx]JOu.OOP!JetJ8tPS [C・∈P・10LLJTOLx].OuOOOP!JeLPOt!S 5 4 3 1 0 5 4 6 7 8 2 3 4 5 L [0m] 6 7 8 rc・∈p・lOuJTOLx1'ouooopてt2tJ00eS [C・uP・lOLLlTOLx】.OuOOOPてeLJ8t2S 8 7 6 5] m O 【 4L 3 2
Fig. 3-4 Experimental and calculated of saccharide concentrationinefnuent solution.
Fig3-5(a)∼(d)に初期基質濃度および担体あたりの酵素濃度を変化させた場合の試料溶液 のリアクター内滞在時間と各糖濃度の関係を示す。本リアクターでは、実験条件で示した 供給液流量9.2, 5.0, 3.0, 1.0 cm3・min-1がそれぞれ滞在時間、 4.5, 8.2, 13.7, 4lsectC=あたるo プロットは実験値であり、ラインは本モデルによる計算値である。どの条件下においても、 滞在時間が長くなるにつれて、 6糖の出口濃度は単調に減少しているが、 4,3,2糖の出口濃 度はそれぞれ増加しており、 4糖はさらに2糖にまで加水分解されていることがわかる。 滞在時間40secとした場合は、出口溶液中に、 6,4糖はほとんど存在していないことがわ かる。 (a)∼(C)は、供給液基質濃度を1.0-3.0 mol・dm-3-1iq.と変化させた結果である.当然の ことながら、供給液中の糖濃度が高いほど、 4,6糖の出口濃度も高くなっている。 (b)と(d) は、担体あたりの酵素濃度が13.5mg・cm-3-particleと9.51mg・cm 3-particleの結果である。こ め場合も滞在時間の変化による各糖の出口濃度に、大きな違いはなかった。一方、どの条 件下においてもシミュレーション結果と実験結果は良好に一致している。これより、本モ デルにより、種々の条件下における本連続酵素加水分解反応を良好に表現することができ ると考えられる。
[C・uJp・]ouJTOLx】.OuOOaP!Jt2LJ99eS 5 4 人■ 2 1 0 ∩?LLJP・rOu寸占Lx】duOC)i-P!JeL199t}S 5 4 3 2 1 543210 FuJP・r.uTOLx].9uOOOP!JeLJ8eS
Fig・3-5 Experimentaland calculated resultsincolumn reactor system・
(a) Cs6 (0) =1・0 × 104mol・ dm13-liquid, CEbn'(o) =13.5mg・cm 3-particle (b)Cs6 (0) =2・0 × 104mol'dm 3-liquid, CEimm (o) =13.5mg・cml3-particle (C)Cs6 (0) =3・0 × 1014mol●dm-3-liquid, CEbw(o) =13.5mg・cm13-particle (d)Cs6 (0) =2・0 × 104mol●dm-3-liquid, CEbw (o) -9.51mg・cm-3-particle