低酸素誘導因子の阻害によるホウ素中性子捕捉療法
の低酸素細胞に対する治療効果の改善
著者
原田 崇臣
学位授与機関
Tohoku University
学位授与番号
11301甲第19167号
URL
http://hdl.handle.net/10097/00129308
博士論文
低酸素誘導因子の阻害による
ホウ素中性子捕捉療法の
低酸素細胞に対する治療効果の改善
東北大学大学院医学系研究科保健学専攻 生体応用技術科学講座放射線治療学分野 原田 崇臣1 目次 略語一覧 ... 2 1. 要約... 3 2. 研究背景... 6 3. 研究目的... 13 4. 研究方法... 14 5. 研究結果... 23 6. 考察... 27 7. 結論... 34 8. 文献... 35 9. 図 ... 41 10. 表 ... 51 謝辞 ... 52
2
略語一覧
本文中および図中で使用した略語は以下の通りである。
・ BNCT boron neutron capture therapy (ホウ素中性子捕捉療法) ・ BPA p-boronophenylalanine (ボロノフェニルアラニン)
・ DFO deferoxamine (デフェロキサミン)
・ 18F-FBPA 18F-fluoro-boronophenylalanine (フルオロボロノフェニルアラニン)
・ HIF-1α hypoxia-inducible factor 1α (低酸素誘導因子)
・ ICP-AES inductively coupled plasma atomic emission spectroscopy (誘導結合プラズマ発光分光分析法)
・ LAT1 L-type amino acid transporter 1 (アミノ酸トランスポーター) ・ LET Linear Energy Transfer (線エネルギー付与)
・ siRNA small interfering RNA (低分子干渉 RNA)
3
1. 要約
【背景】ホウ素中性子捕捉療法 (boron neutron capture therapy; BNCT) の生物効果は、 ホウ素10B と中性子との反応に基づくホウ素線量に依存するため、腫瘍細胞のホウ素 の取り込みは重要な因子となる。ホウ素薬剤として臨床利用されている 10B‐ボロノ フェニルアラニン (10B-p-boronophenylalanine; 10B-BPA) を用いた BNCT において、低 酸素細胞ではホウ素の取り込み能が減少し、BNCT の抗腫瘍効果が減弱することが報 告されている。しかしながら、低酸素細胞において BNCT の効果が減弱するメカニ ズムについては解明されていない。 【目的】低酸素細胞における低酸素誘導変化に関する分子メカニズムを解明し、低酸 素細胞に対するBNCT の治療効果を改善する因子を見出すことを目的とした。 【方法】本研究では、ヒト膠芽腫細胞株T98G、ヒト口腔扁平上皮癌由来細胞株 HSC-3、 ヒト乳腺癌由来細胞株 MCF-7 を使用した。まず、低酸素環境で細胞内に蓄積される 低酸素誘導因子 (hypoxia-inducible factor 1α; HIF-1α) を誘導するため、1% O2の低酸素
負荷、および疑似低酸素薬であるデフェロキサミン (deferoxiamine; DFO) の投与によ って HIF-1α を誘導刺激した状態にて、10B-BPA の取り込みに関与するアミノ酸トラ
ンスポーター (L-type amino acid transporter 1; LAT1) の遺伝子発現量、および10B-BPA
を3.00 mM の終濃度 (30 μg10B/mL) で投与後の細胞内ホウ素量を評価した。次に、
HIF-1α の蓄積と LAT1 の発現量との関係を明らかにするために、HIF-1α の遺伝子発 現をノックダウンしたときのLAT1 の遺伝子発現量を評価した。最後に、HIF 阻害剤
4 が低酸素細胞におけるBNCT の効果を増感するかどうかを検討するため、HIF 阻害薬 を添加して低酸素条件下で 24 時間培養した後に 10B-BPA を投与し、2 時間後に中性 子ビームを照射して細胞の生残率を評価した。 【結果】1% O2の低酸素環境で培養された細胞におけるLAT1 の遺伝子発現量は、通 常酸素比でそれぞれ、T98G で 0.637±0.082 (P = 0.008)、HSC-3 で 0.687±0.061 (P = 0.012)、 MCF-7 で 0.519±0.083 (P = 0.004) であり、いずれの細胞も LAT1 の遺伝子発現量が通 常酸素条件に比べて低下した。10B-BPA 投与 2 時間後のホウ素の取り込み量は、通常 酸素比でそれぞれ、T98G で 0.770±0.110 (P = 0.077)、HSC-3 で 0.663±0.074 (P = 0.010)、 MCF-7 で 0.881±0.096 (P = 0.137) であった。DFO 処理細胞における LAT1 の遺伝子発 現量は、通常酸素比でそれぞれ、T98G で 0.748±0.149 (P = 0.180)、HSC-3 で 0.360±0.014 (P = 0.0003)、MCF-7 で 0.551±0.034 (P = 0.008) であった。また、細胞内ホウ素10B の 取り込み量は、通常酸素比でそれぞれ、T98G で 0.799±0.033 (P = 0.003)、HSC-3 で 0.903±0.043 (P = 0.025)、MCF-7 で 0.940±0.037 (P = 0.066) であった。DFO 投与下にお いて HIF-1α の発現をノックダウンしたところ、細胞内の LAT1 の遺伝子発現量は、 HIF-1α をノックダウンしていないコントロール比でそれぞれ、T98G で 1.256±0.101 (P = 0.102)、HSC-3 で 2.354±0.093 (P = 0.004)、MCF-7 で 1.298±0.056 (P = 0.030) となり、 いずれの細胞においても HIF-1α のノックダウンにより LAT1 の発現量が回復する傾 向が確認された。このことから、低酸素細胞における LAT1 の発現低下に対して、 HIF-1α が関与している可能性が示唆された。さらに、HIF 阻害剤を添加して 1% O2
5 の低酸素負荷条件にて細胞を 24 時間培養し、10B-BPA を投与して 2 時間後に中性子 照射を実施したところ、HIF 阻害剤非添加群および添加群における BNCT 後の細胞生 残率は、T98G で 0.473±0.137 および 0.283±0.071 (P = 0.193)、HSC-3 で 0.424±0.062 お よび0.299±0.054 (P = 0.101) であり、HIF 阻害剤の添加は中性子照射後の細胞生残率 を低下させ、低酸素細胞に対するBNCT の効果を増感し得ることが示された。 【結論】低酸素細胞では、低酸素応答により蓄積された HIF-1α により LAT1 の遺伝 子発現量が抑制され得ることが示された。BNCT における HIF 阻害剤の併用は、低酸 素細胞に対するBNCT の治療効果を改善する可能性がある。
6
2. 研究背景
ホウ素中性子捕捉療法の原理
ホウ素中性子捕捉療法 (Boron Neutron Capture Therapy; BNCT) は、ホウ素原子10B
と熱中性子との核変換反応によって生じる、高いLET を有するヘリウム原子核 (α 線) とリチウム原子核を用いた放射線治療である (図 1)。中性子照射に先立ち、腫瘍細胞 内に選択的に取り込まれる10B を含んだ化合物をあらかじめ投与しておき、原子炉ま たは加速器より取り出した、熱中性子、熱外中性子が主体となるエネルギーの低い中 性子線を照射する。すると10B 原子核はエネルギーの低い熱中性子を捕捉し、核変換 反応を生じる。この核変換反応10B(n,α)7Li により、高エネルギーを有する α 線とリチ ウム原子核が発生する。重要な点は、発生した粒子の生体内での飛程が、α 線で 9 µm、 リチウム原子核で5 µm しか有していないことである。これら 2 つの粒子の飛程は細 胞ほぼ1 個分の大きさに相当しているため、がん細胞内に選択的にホウ素原子が集積 することで、集積のない近接する正常細胞を傷つけることなくがん細胞のみを死滅さ せることが理論上可能となる[1]。 BNCT の歴史と世界の動向 BNCT の基礎原理は、1936 年に Locher によって提唱された。BNCT による治療は、 1951‐1961 年にかけて米国の研究用原子炉を用いて実施されたのが起源であり、日 本では 1968 年に畠中らによって日立炉 (日立製作所) を用いた医療照射が行われた。 1970 年代には、京大炉 (京都大学) および武蔵工大炉 (武蔵工業大学) での治療が実 施できるようになった。1990 年には、JRR-2 (日本原子力研究開発機構) で医療照射
7 が開始され、その後 1999 年からは JRR-4 での照射に切り替えられ、これまでに 107 例の医療照射が実施された。米国では当初、望むような治療効果が得られなかったこ とから、BNCT に対して消極的となっていた。しかしながら、日本の治療成果に影響 を受ける形で、1994 年に臨床研究を再開した。その後は、EU 諸国でも次々と臨床治 療研究が開始された。BNCT の中性子源として原子炉を利用する場合、定期的な安全 点検が必要であり、停止期間が長く安定的に運用しづらいといった欠点があった。と ころが、近年では加速器を用いた BNCT が注目され、多くの加速器ベースの BNCT プロジェクトが進行している。国内では、京都大学が世界をリードして研究を続け、 中性子源として世界初の加速器を開発した。これに基づいた2 号機は、南東北 BNCT 研究センター (福島)、3 号機は 2018 年 6 月に竣工した大阪医科大学関西 BNCT 共同 医療センター (大阪) に設置されている。現在では他に、国立がん研究センター中央 病院 (東京)、いばらき中性子医療センター (茨城)、名古屋大学 (愛知)、江戸川病院 (東京) が加速器 BNCT の導入を進めている。また、国外では、フィンランド、アル ゼンチン、イタリア、ロシア、韓国、中国、イスラエルが加速器ベースの中性子発生 装置の導入を進めている。現在、国内では加速器ベースの BNCT の臨床稼働を目指 し、様々な治験および臨床試験が開始されている。南東北BNCT 研究センターでは、 2016 年 2 月から再発悪性神経膠腫、同年 10 月から再発・局所進行頭頸部癌の治験を 実施した。関西 BNCT 共同医療センターでは、2019 年 8 月から悪性髄膜腫の治験が 開始されている。国外の施設においては、フィンランドで 1999‐2011 年にかけて脳
8 腫瘍および頭頸部癌を対象とした臨床試験が実施され[2,3]、台湾では2010 年から頭頸 部癌を対象とした臨床試験が開始された。しかしながら、いずれも原子炉を利用した 実施報告であり、国外における加速器ベースでのBNCT の臨床報告は未だない。 BNCT の特長と課題 BNCT において、照射野内の腫瘍に隣接する正常組織の損傷はホウ素薬剤の腫瘍選 択性に依存しており、その損傷は X 線治療や粒子線治療といった放射線治療に比べ て低い[4–6]。したがって、BNCT は治療歴のある患者に対して負担の小さい治療であ り、浸潤性の高い悪性神経膠腫や再発頭頸部癌はBNCT のよい適用である。例えば、 耳下腺癌の治療において、X 線治療や粒子線治療では腫瘍近傍の正常組織は腫瘍と同 等の損傷を受ける一方で、BNCT では機能が保持されている正常な耳下腺組織を温存 することが可能である[7]。 BNCT の治療成績を上げるためには 2 つの重要なポイントが挙げられる。まずは、 腫瘍細胞に対して効率的に処方線量を投与することである。そのためには、腫瘍細胞 により選択的に取り込まれるホウ素化合物を開発することが重要であるといえる。も うひとつは、α 線以外の放射線を出来るかぎり減らすことである。なかでも最も影響 が大きいのはガンマ線である。ガンマ線は、加速器から中性子を取り出す際にターゲ ット部を中心に発生する以外に、生体内に存在する各種元素と中性子が反応すること によっても発生する。このガンマ線による被曝をいかに減らすことができるかが重要 なポイントである。
9 ホウ素薬剤 ホウ素薬剤としては、10B‐ドデカボレート (10B-mercaptoundecahydrododecaborate; 10B-BSH) と 10B‐ボロノフェニルアラニン (10B-p-boronophenylalanine; 10B-BPA) が、 脳腫瘍、頭頸部癌、そして悪性黒色腫に用いられてきた (図 2)。10B-BSH は 1 分子に 12 個の10B 原子を持ち、10B の輸送性に優れ、かつ水溶性が高い。正常脳では血液脳 関門によって脳組織への浸透が阻止される一方、関門機能が破綻している悪性脳腫瘍 の組織には浸透し、滞留することによって大きな濃度差を生じることから、脳腫瘍の 治療に用いられてきた[8]。しかしながら、10B-BSH はがん選択性や細胞内集積性が低 く、がん細胞に積極的に取り込まれる性質は備わっていないことから、現在はほとん ど使用されていない。10B-BPA は、三島らが行った 1987 年の悪性黒色腫患者の BNCT において世界で最初に臨床使用された[9]。その結果、悪性黒色腫細胞と正常細胞での 取り込みには大きな差があり、その後の研究で、悪性黒色腫だけでなく多岐にわたる がんに取り込まれることが確認された。10B-BPA はフェニルアラニンがホウ酸残基で 化学修飾されたもので、アミノ酸の細胞膜透過を可能にする膜タンパク質であるアミ ノ酸トランスポーター (L-type amino acid transporter 1; LAT1) を介し、他のアミノ酸 との逆輸送によってがん細胞に取り込まれると考えられており[10]、今日では多くのが
んに対するBNCT で効果が研究されている。BNCT では、抗腫瘍効果を得るためにが ん細胞に対して20 μg10B/mL 以上のホウ素原子10B を集積させる必要があり[11]、患者
10
用や毒性がほとんどないため、一般的な薬剤と比べて比較的多量に投与することが可 能である。実際に、BNCT の治療において、10B-BPA は中性子照射の 2 時間前に投与
が始まり、LAT1 を介して腫瘍細胞に選択的に取り込まれる。そして、10B-BPA 投与 2
時間後、血中のホウ素濃度が20-40 μg10B/mL に保たれた状態で中性子が照射される。
近年では、10B-BPA のポジトロン標識化合物である 18F-FBPA を用いた PET 診断によ
り、患者の腫瘍組織に対する 10B-BPA の集積を治療前に評価することが可能である [12,13]。 がん細胞と低酸素応答 一般的に、がん細胞が異常増殖し血管新生がその増殖に追いつかなくなると、腫瘍 組織内部には酸素が届きにくくなり、その結果低酸素領域が生じる[14]。血管新生が起 こった際にも、その血管網は無秩序で脆弱であるため、腫瘍内部の細胞は低酸素とな りやすい[15]。低酸素細胞では、低酸素に対する適応応答として低酸素誘導 因子
(hypoxia-inducible factor 1α; HIF-1α) が蓄積する。HIF は helixloop-helix (HLH) と Per-Arnt-Sim (PAS) ドメインをもつ α サブユニット (HIF-α) と β サブユニット (HIF-1β) が結合し、1 つの機能的なたんぱく質を構成する転写因子である。α サブユ ニットにはそれぞれ 1α、2α、3α の 3 種類が存在し、その中でも HIF-1α は 1995 年 に Semenza らによって発見され[16]、低酸素応答のメカニズムを解明したことで 2019
年のノーベル医学・生理学賞が授与された。HIF-1α は通常酸素下ではプロリン水酸 化酵素により水酸化され、さらにvon Hippel-lindau (pVHL) によってユビキチン化後、
11 プロテオソームによって速やかに分解される。逆に低酸素下ではこのプロリン水酸化 酵素による水酸化が起こらないため、その先のユビキチン化からの分解反応が起こら ないことによってHIF-1α のタンパクは安定化し、核内に移行し HIF-1β と結合して活 性化された転写因子HIF-1 となり、様々な標的遺伝子の発現領域に結合する。その結 果、エネルギー代謝応答、血管新生、浸潤能の向上といった腫瘍の成長を促す変化が 生じ[17,18]、低酸素細胞は化学療法や分子標的薬といった薬剤に対する抵抗性を獲得す る。 BNCT と低酸素細胞 10B-BSH や10B-BPA などのホウ素薬剤、とりわけ腫瘍細胞のホウ素取り込み能に依 存して細胞内に移行し分布する10B-BPA は、腫瘍内の低酸素状態にある腫瘍細胞には 非常に分布しにくく、結果として BNCT の治療効果が低下し、低酸素状態にある腫 瘍細胞への殺細胞効果が低下することが懸念される。実際に、10B-BPA を用いた BNCT において、慢性的な低酸素条件で培養された細胞では、10B-BPA の取り込み能が酸素 濃度に依存して有意に低下することが明らかにされた[19]。また、低酸素細胞に対する BNCT 後の細胞生残率の結果から、低酸素細胞における BNCT の効果の減弱が報告さ れている[20]。低酸素細胞におけるBNCT の治療効果の減弱への対策として、10B-BSH においては、BNCT 施行時に照射される中性子線ビームに含まれるガンマ線を利用し て、ガンマ線照射による殺細胞効果を向上させ得る放射線増感剤を併用した BNCT が動物実験により評価された[21]。しかしながら、これは BNCT による効果を直接増
12 強するものではなく、低酸素細胞に対する BNCT の効果の改善とは言い難いと考え る。ホウ素化合物の腫瘍選択性の向上を目的として、低酸素細胞に選択的に集積する 2‐ニトロイミダゾールにボロン酸エステル基を導入した B381 を分子設計し、その 生物学的評価を10B-BPA と比較した報告があり、その低酸素細胞への取り込みと保持 は優れていたものの、皮下移植腫瘍モデルマウスにおける腫瘍内注射後の薬剤血中濃 度の評価から、体内動態に改善すべき点があることが問題として挙げられている[22]。 よって、低酸素細胞に対する BNCT の治療効果の減弱が様々な報告から明らかにさ れつつあるものの、現状ではその具体的な改善策が明確になっていない。 そこで本研究では、10B-BPA を用いた BNCT における低酸素細胞に対する治療効果 の減弱に関するメカニズムを解明するために、低酸素環境で細胞内に蓄積するHIF-1α に着目した。HIF-1α の蓄積が、10B-BPA の取り込みに関与するアミノ酸トランスポー ターLAT1 の発現量にどのような影響を及ぼすのかについて評価し、LAT1 の発現の 低酸素誘導変化に関する分子メカニズムが明らかになれば、BNCT の治療効果を高め る新たな治療標的を見出すことが可能となると考える。
13 3. 研究目的 低酸素細胞における低酸素誘導変化に関する分子メカニズムを解明し、低酸素細胞 に対するBNCT の治療効果を改善する因子を見出すことを本研究の目的とした。 まず、低酸素環境で細胞内に蓄積される HIF-1α を誘導するため、低酸素の負荷、 および疑似低酸素薬であるデフェロキサミン (deferoxiamine; DFO) の投与によって HIF-1α を誘導刺激したときの LAT1 の遺伝子発現、細胞内ホウ素の取り込み量を評価 した。次に、HIF-1α の蓄積と LAT1 の発現量との関係を明らかにするために、HIF-1α の発現を抑制した細胞におけるLAT1 の遺伝子発現量を評価した。さらに、低酸素細 胞に対して、HIF 阻害剤と BNCT を併用することで、低酸素細胞に対する BNCT の 効果を増強することができるかどうかを評価した。
14
4. 研究方法 i) 試薬の調製
HIF-1α の誘導刺激として、疑似低酸素薬であるデフェロキサミン (deferoxamine; DFO; Abcam, Cambridge) を使用し、疑似低酸素環境を構築した。DFO の粉末 8.7 mg を蒸留水132.4 μL に溶かして 100 mM のストック溶液を作成し、培養培地で希釈す ることでDFO 含有培地を調製した。低酸素細胞内で蓄積される HIF-1α を阻害する目 的で、YC-1 (3-(5ʹ-ヒドロキシメチル-2ʹ-フリル)-1-ベンジリンダゾール; Abcam) が使用 された。YC-1 の粉末 8.8 mg をジメチルスルホキシド (dimethyl sulfoxide; DMSO; Sigma-Aldrich, Saint Louis) 57.8 μL に溶かして 500 mM のストック溶液を作成し、培 養培地で希釈することでYC-1 含有培地を調製した。
ii) 細胞培養と低酸素処理
ヒト膠芽腫細胞株T98G、ヒト口腔扁平上皮癌由来細胞株 HSC-3、ヒト乳腺癌由来 細胞株MCF-7 は、東北大学加齢医学研究所医用細胞資源センター (仙台) から購入し た。T98G 細胞は serum-free Dulbecco’s modified Eagle medium/Ham’s nutrient mixture F-12 (DMEM/F12 1:1; 富士フィルム和光純薬, 大阪)、HSC-3 細胞および MCF-7 細胞 はserum-free Dulbecco’s modified Eagle medium (DMEM; 富士フィルム和光純薬) でそ れぞれ培養した。すべての培養培地には、10%非働化ウシ胎児血清 (Sigma-Aldrich) と 1% ペニシリン/ストレプトマイシン (Gibco Life Technologies, Waltham) を添加し、
15 37°C 、 5% CO2 条件下で各細胞を培養した。なお、本研究では低酸素環境下で 24 時間の細胞培養を行うため、培養細胞に対するコンタミネーション防止の目的で 1% ペニシリン/ストレプトマイシンの抗生剤をそれぞれ、終濃度 100 U/mL および 100 μg/mL で培地に添加して使用した。なお、本濃度において抗生剤が培養細胞の細胞増 殖に影響を及ぼさないことは確認されている。培養細胞に対する低酸素環境の構築に は、低酸素チャンバー (Billups-Rothenberg Inc., San Diego) を用いた。チャンバー内に 細胞培養ディッシュ、および乾燥防止用に蒸留水を入れて密閉し、チャンバー内に 95% N2と 5% CO2の混合ガスを流して1% O2の低酸素環境を作成した。混合ガスを
流す際には、ガスの流路に酸素濃度計 (JKO-02 Ver. III monitor; JIKCO, 東京) を設置 することでチャンバー内の酸素濃度を確認した。各細胞をそれぞれの細胞の増殖率に 応じて3.0-5.0 × 105 個で 35 mm ディッシュに播種し、12 時間後に細胞のディッシュ
への定着を確認して、通常酸素および低酸素条件下で24 時間培養した。DFO を用い た疑似低酸素条件における評価では、播種後12 時間で培養培地中に DFO を投与し、 通常酸素条件下で24 時間培養した。本研究の流れを図 3 に示した。
iii) DFO および YC-1 の細胞毒性評価
DFO の細胞に対する毒性評価は、T98G、HSC-3 および MCF-7 細胞に対して行った。 調製済みの100 mM の DFO ストック溶液を培養培地で希釈することで、0, 1, 2, 5, 10, 20 μM の DFO 含有培地を調製した。24 ウェルプレートに各濃度の DFO 含有培地を
16
入れ、各細胞を24 ウェルプレートに 1.0 × 105 個で播種した。24 時間および 48 時間
培養後に培地を吸引除去して、Dulbecco's phosphate-buffered saline (D-PBS; 富士フィ ルム和光純薬) で洗浄し、トリプシン処理により細胞を剥がして回収し、トリパンブ ルーアッセイにより生細胞数をカウントした。YC-1 の細胞に対する毒性評価は、 T98G および HSC-3 細胞に対して行った。調製済みの 500 mM の YC-1 ストック溶液 を培養培地で希釈することで、0, 0.25, 0.5, 1, 2, 4 μM の DFO 含有培地を調製した。各 細胞の播種および評価は、DFO の毒性評価と同様の方法にて実施した。
iv) total RNA の抽出と定量的リアルタイム reverse transcription polymerase chain reaction (RT-PCR)による遺伝子発現の定量評価
NucleoSpin® RNA (タカラバイオ, 大津) の抽出プロトコルに基づき、スピンカラム 法によって培養細胞からtotal RNA を抽出した。抽出した total RNA の濃度は分光光 度計 (NanoDrop One; ThermoFisher Scientific, Inc., Waltham) を用いて測定し、最も濃 度の低かったサンプルにおけるtotal RNA を基準にして、各サンプルの濃度をヌクレ アーゼフリー水により希釈した。濃度調整後の各サンプルのtotal RNA 2.5 μL に対し、 iScript RT Supermix for RT-qPCR® (Bio-Rad, Hercules) 1.5 μL、ヌクレアーゼフリー水 6 μL を加えて 10 μL とし、サーマルサイクラーで 25°C で 5 分間、42°C で 30 分間、85°C で 5 分間処置することで、complementary DNA (cDNA) を合成した。目的遺伝子の mRNA 発現量は、リアルタイム PCR 解析システム (CFX connect™ real-time PCR
17
detection system; Bio-Rad) を 用 い た 、 定 量 的 リ ア ル タ イ ム reverse transcription polymerase chain reaction (RT-PCR)により定量した。各サンプルの cDNA 1 μL に対し、 SsoAdvanced Universal SYBR Green Supermix® (Bio-Rad) 5 μL、ヌクレアーゼフリー水 4 μL、目的遺伝子のプライマー (Forward primer: 0.1μL, Reverse primer: 0.1μL) を加え て、サーマルサイクラーで90°C で 30 秒間処理した後、98°C で 10 秒間、60°C で 30 秒間の処理を1 サイクルとして 60 サイクル反応させた。目的遺伝子の mRNA 発現量 は検量線法を用いて定量した。また、ハウスキーピング遺伝子としてGAPDH を使用 した。PCR に用いたプライマー配列を表 1 に示した。
v) small interfering RNA (siRNA) による HIF-1α のノックダウン
各細胞をそれぞれの細胞の増殖率に応じて3.0-5.0 × 105 個で 35 mm ディッシュに
播種し、12 時間後に細胞のディッシュへの定着を確認して、siRNA 試薬の調製を行 った。HIF1A 標的 siRNA (MISSION® siRNA; Sigma-Aldrich; Merck KGaA) 1 μL を Opti-MEM® I Reduced Serum Medium (Gibco Life Technologies) 50 μL で希釈調製した 溶液に対し、Lipofectamine® RNAiMAX Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) 3.5 μL を Opti-MEM® I Reduced Serum Medium 50 μL で希釈調製した溶液を混注 して20 分間室温でインキュベートした。なお、siRNA 非導入群に対する試薬として、 HIF1A 標的 siRNA 1 μL に代わり BLOCK-iT™ Alexa Fluor® Red Fluorescent Oligo 20 μM (Invitrogen) 1 μL を使用し、HIF1A 標的 siRNA と同様の調製を行った。20 分後、
18 各試薬をボルテックスにかけて軽く撹拌し、各細胞の培養培地に100 μL 添加して 24 時間培養した。24 時間後、培地を吸引除去して D-PBS で洗浄し、トリプシン処理に より細胞をディッシュから剥がしてセルカウントした後、各細胞の増殖率に応じて 35 mm ディッシュに 3.0-5.0 × 105 個で播種した。24 時間後、培養培地中に DFO を 5 μM の終濃度で投与し、通常酸素条件下でさらに24 時間培養した。siRNA の HIF-1α に対 するノックダウンの効果は、iv) に記載した PCR の方法に準じて遺伝子発現解析を行 うことで評価した。siRNA の実験プロトコルを図 4 に示した。また、siRNA の配列を 表2 に示した。 vi) ウェスタンブロッティング法によるタンパク発現評価
培養細胞に対して培地を吸引除去した後、Cell Lysis Buffer (Cell Signaling Technology, Inc., Danvers) 30 μL、蒸留水 270 µL からなる細胞溶解液と、プロテアーゼ阻害カクテ ル (タカラバイオ) 3 μL を加えて、氷上で 5 分間インキュベーションした後、セルス クレーパーで細胞を剥ぎ取り、1.5 mL チューブにした。氷上でさらに 20 分間インキ ュベーションした後、ホモジナイザーで60 秒間処理して細胞を破砕し、回転数 15,000 rpm で 20 分間遠心した後、上清のみを回収した。ここで得られた上清をタンパク質 試料として、タンパク質試料 270 μL に Laemmli sample buffer (4X) (Bio-Rad) 90 μL と、 2-メルカプトエタノール (Bio-Rad) 7.5 µL (終濃度 2.1%) を加えて電気泳動試料とし、 泳動処理まで冷凍庫で保存した。電気泳動前、泳動試料を沸騰水で5 分間加熱処理し
19
た後、12%のポリアクリルアミドゲルを使用して電気泳動法でタンパク質を分離し、 転写装置を用いてポリフッ化ビニリデン膜 (Trans-Blot® Turbo™ ; Bio-Rad) に転写し た。ブロックエース粉末 (Block ACE®; DS Pharma Biomedical Co., Ltd., 大阪) より作 製した4%ブロックエース水溶液で、転写したフッ化ビニリデン膜を 1 時間ブロッキ ングした。ブロッキング液を吸引除去後、トリス緩衝生理的食塩水 (TBS; Bio-Rad) に 対し酵素イムノアッセイグレードポリソルベート表面活性剤 (Tween-20; Bio-Rad) に よる1% Tween-20 溶液を調製し、蒸留水で 10 倍希釈した溶液 (TBST) で膜を洗浄し た 。 洗 浄 後 、 一 次 抗 体 反 応 液 (Can Get Signal® solution 1; Can Get Signal® Immunoreaction Enhancer Solution; 東洋紡, 大阪) で 1000 倍希釈したマウスモノクロ ーナル抗アクチン抗体 (Clone: C4) (Santa Cruz Biotechnology, Dallas)、および 500 倍希 釈したマウスモノクローナル抗 HIF-1α 抗体 (Clone: 514/HIF-1α) (BD Biosciences, Franklin Lakes) に膜を浸し 1 時間処理した。TBST で 3 回洗浄後、二次抗体反応液 (Can Get Signal® solution 2; Can Get Signal® Immunoreaction Enhancer Solution; 東洋紡) で 2000 倍希釈したマウス IgG に対する西洋ワサビペルオキシターゼ標識二次抗体 (Cell Signaling Technology) に膜を浸し 1 時間処理した。その後、TBST で 3 回洗浄し、化 学発光基質 (Clarity Western ECL Substrate; Bio-Rad) を添加した後、画像解析装置 (ChemiDoc MP; Bio-Rad) にてバンドの検出を行った。
20
高純度の10B からなる L-p-BPA (10B-BPA) 溶液、標準ホウ素溶液、イットリウム標
準液は、ステラファーマ株式会社 (大阪) から提供された。各条件で 24 時間培養さ れた細胞に対し、培養培地に10B-BPA を 3.00 mM の終濃度 (30 μg10B/mL) で投与し、
最大2 時間まで10B-BPA 曝露処理を行った。10B-BPA の処理が完了した後10B-BPA を
含んだ培養培地を吸引除去し、過塩素酸と過酸化水素水を1:2 の混合比で混ぜた溶液 700 μL を加えて、50℃で 2 時間処理した。2 時間後、各サンプルの処理液 500 μL に 対して、イットリウム標準液 500 μL、蒸留水 4 mL を加えた全量 5 mL を測定試料と した。各測定試料中の10B の濃度は、ICPE-9000 (島津, 京都) を用いた誘導結合プラ
ズ マ 発 光 分 光 分 析 法 (inductively coupled plasma atomic emission spectroscopy; ICP-AES) により測定された。10B の測定に使用する波長は 208.959 nm とした。なお、 各サンプルの測定前には、標準ホウ素溶液の希釈系列から検量線を作成し、0.05-5.0 μg10B/mL の範囲で直線性が得られていることを確認した。 viii) 中性子照射後の細胞生残率の評価 各細胞を通常酸素および低酸素条件下で24 時間培養した後、10B-BPA を投与して 2 時間処理した。2 時間後、10B-BPA を含んだ培地を培養ディッシュから吸引除去し、 中性子照射専用のアクリルファントムにディッシュを設置して中性子を照射した。中 性子照射は、南東北BNCT 研究センターの加速器中性子発生装置 (HM-30; 住友重機 械工業, 東京) を用い、照射ビーム電荷量を 0.6 C、平均ビーム電流を 1.0 mA として
21 照射を実施した。中性子照射後、トリプシン処理により細胞をディッシュから剥がし、 セルカウントした後、60 mm ディッシュに 300-2000 個/ディッシュとなるように播種 した。それぞれの細胞の増殖率に応じて、播種してから 10-12 日後に細胞を 100%メ タノールで固定処理し、ギムザ染色液 (ナカライテスク, 京都) とリン酸緩衝液を 1:2 の混合比で混ぜた溶液を蒸留水で 2 倍希釈した後にディッシュに添加し、30 分間静 置して細胞を染色した。コロニーアッセイに関する先行研究の方法[23–27]に則り、50 個以上の細胞で形成された細胞群を1 コロニーとしてカウントした。通常酸素および 低酸素条件で24 時間培養後の細胞に対する BNCT 実施後の細胞生残率評価において は、非照射サンプルのコロニー数に対する照射サンプルのコロニー数の比として、ま た、YC-1 の添加による BNCT 実施後の細胞生残率評価においては、10B-BPA 非投与 の中性子照射サンプルのコロニー数に対する各照射サンプルのコロニー数の比とし て、細胞生残率を算出した。 ix) 統計解析
DFO および YC-1 の細胞毒性評価に関する実験は 2 回、T98G 細胞における YC-1 添加の照射後細胞生残率の評価に関する実験は4 回、それ以外のすべての実験は 3 回 実施し、結果は2 回の実験では平均値±標準偏差、3 回以上の実験では平均値±標準誤 差として表した。統計解析にはマイクロソフトエクセルのStatcel 3 (OMS Publishing, 埼玉) を使用した。LAT1 の遺伝子発現評価、細胞内ホウ素量評価、照射後の細胞生
22 残率評価はスチューデントの t 検定、HIF-1α ノックダウンサンプルにおける HIF-1α およびLAT1 の遺伝子発現評価はスチューデントの t 検定またはウェルチの t 検定、 YC-1 添加の照射後細胞生残率の評価はスチューデントの t 検定またはマン・ホイッ トニーU 検定により検定し、統計的有意性を評価した。P < 0.05 をもって統計的に有 意とした。
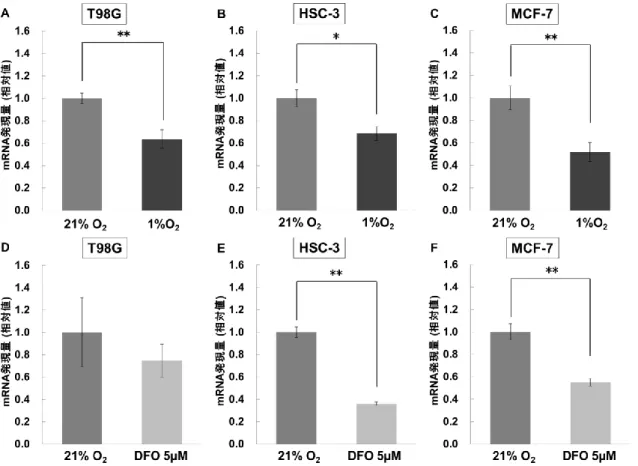
23 5. 研究結果 低酸素および疑似低酸素環境においてBNCT の効果は減弱する DFO の細胞毒性評価に関して、T98G 細胞は、24 時間培養においては DFO の濃度 変化に対する細胞数の変化は確認されなかった。しかしながら、48 時間培養では 10 μM より高い濃度において細胞数の減少が確認された (図 5A)。HSC-3 細胞において は、24、48 時間培養ともに DFO の濃度変化に対する細胞数の減少が確認された (図 5B)。MCF-7 細胞では、24 時間培養において DFO の濃度変化に対する細胞数の変化 は確認されなかったものの、48 時間培養では 5 μM より高い濃度において細胞数の減 少が確認された (図 5C)。1% O2の低酸素状態およびDFO 投与による疑似低酸素状態 は、ウェスタンブロッティングによるHIF-1α のタンパク発現量により評価した。T98G 細胞において、1% O2の低酸素負荷条件および5 μM の DFO 投与条件での HIF-1α の タンパク発現量は21% O2の通常酸素条件に比べて強く発現しており、定量結果は通 常酸素比でそれぞれ、6.25、2.75 であった (図 6)。 10B-BPA の取り込みに関与する LAT1 に関して、1% O 2の低酸素環境で培養された 細胞におけるLAT1 の遺伝子発現量は、通常酸素比でそれぞれ、T98G で 0.637±0.082 (P = 0.008)、HSC-3 で 0.687±0.061 (P = 0.012)、MCF-7 で 0.519±0.083 (P = 0.004) であ り、いずれの細胞も LAT1 の遺伝子発現量が通常酸素条件に比べて有意に低下した (図 7A-C)。また、5 μM の DFO を投与された細胞における LAT1 の遺伝子発現量は、 通常酸素比でそれぞれ、T98G で 0.748±0.149 (P = 0.180)、HSC-3 で 0.360±0.014 (P =
24
0.0003)、MCF-7 で 0.551±0.034 (P = 0.008) であった (図 7D-F)。T98G 細胞におけるホ ウ素の取り込みは、5 μM の DFO 投与によって10B-BPA 投与 2 時間後まで継続的に抑
制された。しかしながら、HSC-3、MCF-7 細胞では有意な差が確認されなかった (図 8A)。10B-BPA 投与 2 時間後のホウ素の取り込み量は、DFO 非投与群比でそれぞれ、
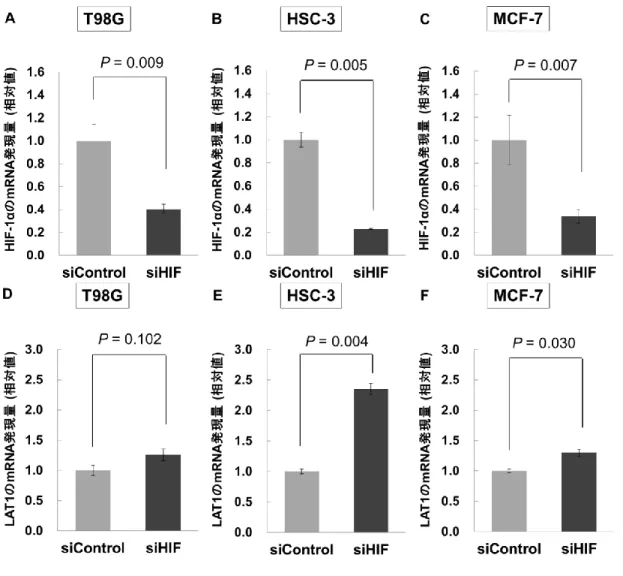
T98G で 0.799±0.033 (P = 0.003) 、HSC-3 で 0.903±0.043 (P = 0.025)、MCF-7 で 0.940±0.037 (P = 0.066) であった (図 8B-D)。また、1% O2の低酸素負荷条件下にお ける 10B-BPA 投与 2 時間後のホウ素の取り込み量は、通常酸素比でそれぞれ、T98G で0.770±0.110 (P = 0.077)、HSC-3 で 0.663±0.074 (P = 0.010)、MCF-7 で 0.881±0.096 (P = 0.137) であった (図 8E)。 HIF-1α の蓄積は LAT1 の遺伝子発現に影響する 低酸素環境において細胞内に蓄積する HIF-1α と LAT1 との関係の有無を明らかに するため、siRNA により HIF-1α の発現をノックダウンしたときの LAT1 の mRNA 発 現量をRT-PCR にて評価した (図 9A-F)。siRNA により HIF-1α の発現をノックダウン したときの細胞内のHIF-1α の遺伝子発現量は、siRNA 非導入群 (siControl) 比でそれ ぞれ、T98G で 0.406±0.041 (P = 0.009)、HSC-3 で 0.228±0.006 (P = 0.005)、MCF-7 で 0.339±0.058 (P = 0.007) となり、いずれの細胞においても siHIF 導入群で HIF-1α の mRNA 発現量は有意に低下し、HIF-1α がノックダウンされていることが確認された。 5 μM の DFO 投与下において、siRNA により HIF-1α の発現をノックダウンしたとこ
25
ろ、細胞内の LAT1 の遺伝子発現量は、siRNA 非導入群 (siControl) 比でそれぞれ、 T98G で 1.256±0.101 (P = 0.102)、HSC-3 で 2.354±0.093 (P = 0.004)、MCF-7 で 1.298±0.056 (P = 0.030) となった。 HIF 阻害剤 YC-1 は低酸素細胞に対する BNCT の抗腫瘍効果を増感する まず、各細胞を通常酸素および低酸素条件下で24 時間培養した後、培地に10B-BPA を3.00 mM の終濃度 (30 μg10B/mL) で投与し、2 時間後に中性子ビームを照射した後 の細胞生残率を評価した。通常酸素および低酸素条件で培養された細胞における BNCT 後の細胞生残率は、10B-BPA 投与の中性子非照射群比でそれぞれ、T98G で 0.414±0.049 および 0.600±0.048 (P = 0.027)、HSC-3 で 0.062±0.011 および 0.107±0.012 (P = 0.024)、MCF-7 で 0.036±0.016 および 0.045±0.025 (P = 0.396) となり、T98G、HSC-3 細胞においては通常酸素条件より低酸素条件における細胞生残率が有意に高くなっ た (図 10A-C)。次に、HIF 阻害剤である YC-1 によって、低酸素細胞に対する BNCT の効果が増強されるかどうかを T98G、HSC-3 細胞にて評価した。各細胞に対する YC-1 の毒性評価より、0.5 μM 以下の濃度の YC-1 においては細胞毒性が確認されな かった (図 11A, B)。T98G および HSC-3 細胞に対して、0.5 μM の YC-1 を添加して 低酸素条件下で 24 時間培養した後に 10B-BPA を投与し、2 時間後に中性子ビームを 照射して細胞の生残率を評価した結果を図11C, D に示す。3.00 mM の10B-BPA (30 μg10B/mL) 投与において、YC-1 非添加群および添加群の細胞生残率は、10B-BPA 非投
26 与の中性子照射群比でそれぞれ、T98G で 0.473±0.137 および 0.283±0.071 (P = 0.193)、 HSC-3 で 0.424±0.062 および 0.299±0.054 (P = 0.101) であった。1.00 mM の10B-BPA 投与 (10 μg10B/mL) においても、YC-1 非添加群および添加群の細胞生残率はそれぞ れ、T98G で 0.925±0.140 および 0.517±0.016 (P = 0.021)、HSC-3 で 0.844±0.108 および 0.357±0.048 (P = 0.007) となり、YC-1 非添加群に比べて添加群で細胞生残率は低い値 を示した。
27
6. 考察
本研究では、1% O2の低酸素負荷、および疑似低酸素薬である DFO の投与によっ
て、低酸素環境で細胞内に蓄積される HIF-1α を誘導刺激した。疑似低酸素薬に使用 した DFO は、鉄過剰病に対する鉄のキレート剤として臨床使用されている薬剤であ る。DFO は HIF-1α の転写活性を抑制するプロリン水酸化酵素の活性を阻害し、HIF-1α を安定化させる[28]。DFO を使用することのメリットは、培養中の細胞に DFO を投与 するだけで、比較的容易に疑似的な低酸素状態を作り出すことができる点である。ま た、in vitro において DFO を用いた疑似低酸素環境の構築はすでに多くの先行研究で 行われていることから、本研究における培養細胞に対する DFO の使用は、疑似低酸 素環境の構築に妥当であるといえる。[29,30] T98G および MCF-7 細胞では 5 μM の DFO までは細胞毒性を示さなかったのに対 し、HSC-3 細胞では DFO の濃度変化に対する生細胞数の減少が確認された。しかし ながら、5 μM の DFO 投与条件では 48 時間培養でも死細胞の顕著な増加は確認され ず、細胞の絶対数に関しては経時的に増加していたことから、DFO の濃度増加に伴 ってより多くの細胞死が引き起こされたのではなく、細胞に対する増殖抑制が引き起 こされたと考察される (図 5D-F)。DFO 投与 24 時間後の細胞毒性の結果から、本研 究におけるDFO の投与濃度を 5 μM とし、1% O2の低酸素負荷条件と5 μM の DFO 投与条件における HIF-1α のタンパク発現の評価を行ったところ、両条件ともに通常 酸素条件と比較してHIF-1α が十分に誘導されていることが確認された (図 6)。
28 10B-BPA の取り込みに関与する LAT1 の遺伝子発現について、1% O 2の低酸素負荷 により HIF-1α が誘導された条件下では、いずれの細胞においても通常酸素条件と比 較してLAT1 の発現量の有意な低下が確認された (図 7)。5 μM の DFO 投与条件では、 HSC-3、MCF-7 細胞にて有意な差が示されたものの、T98G 細胞では有意差が示され なかった。また、各細胞において、とりわけ HSC-3 細胞において、低酸素負荷条件 とDFO 投与条件で反応性に相違がみられた。これらの要因は、1%O2の低酸素負荷と 5 μM の DFO 投与で到達できる細胞内の酸素状態が異なることにあると考える。それ に加え、DFO によるキレート作用と低酸素負荷では、各細胞小器官に与える影響が 異なる可能性がある。今後は、生細胞での低酸素イメージング試薬を用いた細胞内イ メージングを行い、低酸素負荷条件と DFO 投与条件における細胞内の低酸素状態を 評価する必要があると考えている。 ホウ素の取り込み評価において、DFO を用いた T98G 細胞では有意な差が示された。 しかしながら、HSC-3、MCF-7 細胞では有意差は示されなかった (図 8)。ICP-AES に よる元素分析においては、装置に起因する測定誤差のほか、実試料溶液の粘性や表面 張力が標準溶液とは異なるために生じる、噴霧効率やミスト輸送効率の変化による発 光強度の変化が少なからず結果のばらつきに影響する。そのため、HSC-3、MCF-7 細 胞における 2 時間後のホウ素の取り込み量は、DFO 非投与群に比べて減少したもの の、有意な差が得られなかった可能性がある。1% O2の低酸素負荷条件では、T98G、 MCF-7 細胞においてホウ素の取り込み量に有意な差がみられなかった。低酸素条件
29 では、アミノ酸代謝が低下することが知られている[31,32]。したがって、10B-BPA の腫 瘍細胞への集積が低下した原因として、HIF-1α を介する低酸素応答に基づく変化の みならず、代謝の低下に基づいた変化が影響している可能性があるが、本研究におい ては前者に焦点を当てて評価を行ったため、後者の影響については評価できていない。 今後は、低酸素条件におけるアミノ酸の代謝低下に起因する10B-BPA の集積に関する さらなる評価が必要であると考えられる。ホウ素の取り込みに関与するLAT1 は、ア ミノ酸トランスポーターとして、遺伝子より翻訳を受けた後に細胞膜上へ移送されて 初めてその機能を発揮する。したがって、細胞膜自体のターンオーバーや膜タンパク 代謝によっても機能発現に影響が生じると考えられる。本研究では、DFO 投与時の LAT1 発現とホウ素摂取量に乖離がみられたが、これは DFO が膜代謝に対して何らか の影響を与えたことで生じた可能性があると考察される。 低酸素環境において細胞内に蓄積する HIF-1α と、ホウ素の取り込みに関与する LAT1 との関係を明らかにするため、siRNA により HIF-1α の発現を阻害したときの LAT1 の遺伝子発現量を評価した。siHIF を用いた HIF-1α の発現阻害の結果より、い ずれの細胞においてもHIF-1α の mRNA 発現量は有意に低下した (図 9A-C)。このこ とは、本研究にて使用したsiRNA が細胞の HIF-1α の発現を効果的にノックダウンし ていることを示している。さらに、5 μM の DFO を用いた疑似低酸素環境では、siHIF 非導入群に比べてsiHIF 導入群において LAT1 の発現量が増加し、HSC-3、MCF-7 細 胞では有意な差が確認された。図9 より、HIF-1α の遺伝子発現量が siHIF 導入群で低
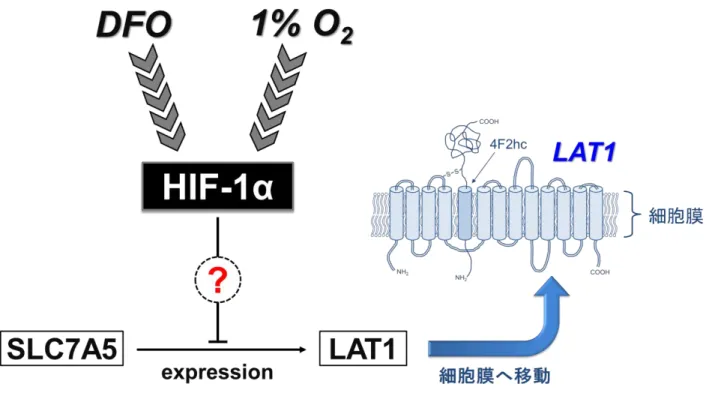
30 下しているものほど、LAT1 の発現回復が大きいことがわかることから、siRNA の導 入効率の違いが各細胞間におけるLAT1 の発現量の違いに関係していると考察される。 本研究により、低酸素細胞におけるLAT1 の発現に対して、HIF-1α が抑制的に働い ていることが明らかとなった。HIF-1α の蓄積と LAT1 の発現低下のメカニズムを図 12 に示した。低酸素により蓄積する HIF-1α のノックダウンに関しては、HIF-1α ノッ クダウン腫瘍移植マウスにおける放射線治療の再発抑制の効果が報告されている [33,34]が、BNCT において、HIF-1α のノックダウンと LAT1 の遺伝子発現評価に関する 報告は今までになく、本研究から得られた新たな知見であると考える。低酸素細胞に おけるLAT1 の発現量の回復は、細胞のホウ素の取り込み量の増加や中性子照射後の 細胞生残率の低下につながる可能性があり、HIF-1α は低酸素細胞における BNCT の 効果を改善するための要因となり得ることが示された。 低酸素細胞に対する BNCT 後の細胞生残率より、先行研究の報告と同様に低酸素 細胞におけるBNCT の効果の減弱が確認された (図 10)。T98G および HSC-3 細胞に おいては有意な差が得られたが、MCF-7 では有意差がみられなかった。今回評価し た細胞はすべて同一の照射条件で照射したが、MCF-7 における BNCT 後の細胞生残 率が通常酸素条件で0.036±0.016、低酸素条件で 0.045±0.025 と、T98G および HSC-3 細胞に比べて非常に低いため、有意な差が得られなかったと考える。 通常酸素および低酸素条件における照射後の細胞生残率に有意な差がみられた T98G および HSC-3 細胞に対して、低酸素条件下で YC-1 を添加し、BNCT 後の細胞
31 生残率を評価することで、低酸素細胞に対する HIF 阻害剤の併用による BNCT の治 療効果の増感を検討した。HIF 阻害剤として本研究で使用した YC-1 は、血小板凝集 に対する抑制効果を持ち、薬理学的に利用されている[35,36]。YC-1 の機序に関しては 詳細不明な部分が多いが、がん細胞においてはYC-1 により HIF 阻害因子を刺激する ことで、HIF-1α の活性化が抑制されるといった報告がある[37]。また、YC-1 により HIF を阻害することで、放射線治療後の腫瘍増殖を有意に抑制可能であることがマウ スを用いた実験により確認されており、放射線増感剤としての効果が期待されている [33,34,38,39]。ゆえに、BNCT においても低酸素細胞に対する治療効果を改善させるため、 慢性低酸素分画に対する増感を想定して YC-1 を用いた。YC-1 や低酸素毒といわれ るtirapazamine (TPZ) や TH-302 といった一連の薬剤、低酸素のマーカーとして低酸 素組織に蓄積するpimonidazole、低酸素増感剤である misonidazole、doranidazole とい った薬剤はいずれも、全身投与されると壊死周囲の低酸素組織に蓄積することが組織 学的に明らかにされている[40,41]。頭頸部癌に対するシスプラチンと TPZ を併用した
化学放射線療法の第III 相臨床試験では、事前に低酸素 PET である18F-FMISO-PET/CT
によって腫瘍内に低酸素分画が確認された症例のサブスタディ解析において、TPZ 全 身投与の併用が局所領域制御に寄与したことが報告されている[42]。したがって、YC-1
を全身投与しても低酸素領域に十分届くと考えられる。しかしながら、BNCT ではホ ウ素薬剤を治療の2 時間前から点滴にて持続投与するため、今後はホウ素薬剤の投与 との関係も考慮して、投与方法や投与量、投与のタイミングに関する評価をする必要
32 がある。 本研究では、HIF-1α と LAT1 の評価からその直接的な関係を示したが、低酸素環境 ではHIF-1α の影響を受けて様々な因子がその発現を亢進・低下させる可能性がある。 また、通常酸素条件下にて、LAT1 の阻害によるホウ素摂取量の低下が報告されてお り、HIF-1α の蓄積が起こらなくとも LAT1 が阻害されることでホウ素の摂取量が低下 することは明らかにされているが[10,43,44]、逆に低酸素環境のような HIF-1α が蓄積し ている条件下においてLAT1 を強制発現したときに、BNCT 抵抗性を解除できるかど うかについては評価できていない。通常酸素環境において構造的に安定な HIF-1α を 強制発現したり、HIF-1α の分解を制御する pVHL やプロリン水酸化酵素をノックダ ウンしたりすることで、LAT1 の発現低下や BNCT 抵抗性が生じるかどうかについて も検討が十分ではない。さらには、BNCT 実施後の細胞に対する HIF-1α による直接 的な作用や、X 線に対する酸素効果に類似した効果が寄与している可能性についても 評価すべきであり、この点が本研究の限界である。今後は、HIF-1α の影響を受けて 発現が変化する他の様々な因子について評価し、より詳細な分子メカニズムの解明が 必要であると考える。 また、本研究では低酸素細胞におけるBNCT の効果の減弱に HIF-1α が普遍的に関 与しているかどうかを確認するため、3 種類の細胞株を用いた。細胞ごとの反応の違 いに焦点を当ててメカニズムを解明するためには、それぞれの細胞株の有するデータ ベースにあるエピゲノミクスの違いのみならず、低酸素の刺激による変化を多角的に
33
評価しなければならず、本研究においては十分な検討ができなかった。例えば、DFO に対する細胞間の反応性の差異は、DFO の酸素キレート作用に依存した細胞内の酸 素状態や分布の違い、好気性代謝を行うミトコンドリアの細胞内密度によっても生じ うる可能性が考えられるため、今後のさらなる評価が必要であると考えられる。
34
7. 結論
低酸素細胞における BNCT の治療効果の低下には、低酸素応答により蓄積された HIF-1α による LAT1 遺伝子発現の低下が関与することが示唆された。HIF 阻害剤 YC-1 は、低酸素腫瘍細胞において10B-BPA を用いた BNCT の抗腫瘍効果を増強した。BNCT
における HIF 阻害剤の併用は、低酸素細胞に対する BNCT の治療効果を増感させる 可能性がある。
35
8. 文献
1. 日本中性子捕捉療法学会 著:BNCT基礎から臨床応用まで ‐BNCTを用いて治療
にかかわる人のためのテキスト‐, 大阪公立大学共同出版会, 大阪, 2018 2. Joensuu H, Kankaanranta L, Seppälä T et al. Boron neutron capture therapy of brain
tumors: Clinical trials at the Finnish facility using boronophenylalanine. J Neurooncol
2003;62:123–34.
3. Kankaanranta L, Seppälä T, Koivunoro H et al. L-boronophenylalanine-mediated boron
neutron capture therapy for malignant glioma progressing after external beam radiation
therapy: A phase i study. Int J Radiat Oncol Biol Phys 2011;80:369–76.
4. Minohara S, Endo M, Kanai T et al. Estimating uncertainties of the geometrical range of
particle radiotherapy during respiration. Int J Radiat Oncol Biol Phys 2003;56:121–5.
5. Ariyoshi Y, Miyatake S, Kimura Y et al. Boron neuron capture therapy using epithermal
neutrons for recurrent cancer in the oral cavity and cervical lymph node metastasis. Oncol Rep
2007:861–6.
6. Kankaanranta L, Seppälä T, Koivunoro H et al. Boron Neutron Capture Therapy in the
Treatment of Locally Recurred Head and Neck Cancer. Int J Radiat Oncol Biol Phys
2007;69:475–82.
7. Aihara T, Morita N, Kamitani N et al. Boron neutron capture therapy for advanced salivary
36
8. Soloway AH, Hatanaka H, Davis MA. Penetration of Brain and Brain Tumor. V II.
Tumor-Binding Sulfhydryl Boron Compounds. J Med Chem 1967;10:714–7.
9. Mishima Y, Honda C, Ichihashi M. Treatment of malignant melanoma by single thermal
neutron capture therapy with melanoma-seeking 10b-compound. Lancet 1989;334:388–9.
10. Detta A, Cruickshank GS. L-Amino Acid Transporter-1 and Boronophenylalanine-Based
Boron Neutron Capture Therapy of Human Brain Tumors. Cancer Res 2009;69:2126–32.
11. Soloway AH, Tjarks W, Barnum BA et al. The Chemistry of Neutron Capture Therapy.
Chem Rev 1998;98:1515–62.
12. Ishiwata K, Ido T, Mejia AA et al. Synthesis and radiation dosimetry of
4-borono-2-[18F]fluoro-d,l-phenylalanine: A target compound for PET and boron neutron
capture therapy. Int J Radiat Appl Instrumentation Part 1991;42:325–8.
13. Evangelista L, Jori G, Martini D et al. Boron neutron capture therapy and 18F-labelled
borophenylalanine positron emission tomography: A critical and clinical overview of
theliterature. Appl Radiat Isot 2013;74:91–101.
14. Höckel M, Vaupel P. Tumor hypoxia: Definitions and current clinical, biologic, and
molecular aspects. J Natl Cancer Inst 2001;93:266–76.
15. Chapman JD. Measurement of tumor hypoxia by invasive and non-invasive procedures: a
review of recent clinical studies. Radiother Oncol 1991;20:13–9.
37
hypoxia-inducible factor 1. Kidney Int 1997;51:553–5.
17. Pennacchietti S, Michieli P, Galluzzo M et al. Hypoxia promotes invasive growth by
transcriptional activation of the met protooncogene. Cancer Cell 2003;3:347–61.
18. Rofstad EK. Microenvironment-induced cancer metastasis. Int J Radiat Biol
2000;76:589–605.
19. Wada Y, Hirose K, Harada T et al. Impact of oxygen status on 10B-BPA uptake into
human glioblastoma cells, referring to significance in boron neutron capture therapy. J Radiat
Res 2018;59:122–8.
20. Masunaga SI, Tatebe H, Nishimura Y et al. Effect of oxygen pressure during incubation
with a 10B-carrier on 10B uptake capacity of cultured p53 wild-type and mutated tumor cells:
Dependency on p53 status of tumor cells and types of 10B-carriers. Int J Radiat Biol
2016;92:187–94.
21. Masunaga SI, Nagasawa H, Gotoh K et al. Evaluation of hypoxia-specific cytotoxic
bioreductive agent-sodium borocaptate- 10B conjugates as 10B-carriers in boron neutron
capture therapy. Radiat Med - Med Imaging Radiat Oncol 2006;24:98–107.
22. Luderer MJ, Muz B, de la Puente P et al. A Hypoxia-Targeted Boron Neutron Capture
Therapy Agent for the Treatment of Glioma. Pharm Res 2016;33:2530–9.
23. Hiramatsu R, Kawabata S, Tanaka H et al.
38
therapy and boron neutron capture therapy. J Pharm Sci 2015;104:962–70.
24. Ichikawa G, Tsuchida K, Kiyanagi Y et al. Development of thermal neutron moderator for
testing boron agents for Boron Neutron Capture Therapy (BNCT). J Instrum
2019;14:T06010–T06010.
25. Mucci A, Kunkiel J, Suzuki T et al. Murine iPSC-Derived Macrophages as a Tool for
Disease Modeling of Hereditary Pulmonary Alveolar Proteinosis due to Csf2rb Deficiency.
Stem Cell Reports 2016;7:292–305.
26. Nakamura M, Kyo S, Zhang B et al. Prognostic impact of CD133 expression as a
tumor-initiating cell marker in endometrial cancer. Hum Pathol 2010;41:1516–29.
27. Kim RK, Kaushik N, Suh Y et al. Radiation driven epithelial-mesenchymal transition is
mediated by Notch signaling in breast cancer. Oncotarget 2016;7:53430–42.
28. Brittenham GM. Iron-Chelating Therapy for Transfusional Iron Overload. Th e new engl J
o f Med Clin 2011;364:146–56.
29. Wang X, Zhao R, Geranpayehvaghei M et al. Targeted Co-delivery of the Iron Chelator
Deferoxamine and a HIF1α Inhibitor Impairs Pancreatic Tumor Growth. ACS Nano 2019;13:2176–89.
30. Yao Q, Liu Y, Selvaratnam B et al. Mesoporous silicate nanoparticles/3D nanofibrous
scaffold-mediated dual-drug delivery for bone tissue engineering. J Control Release
39
31. Rajpurohit R, Koch CJ, Tao Z et al. Adaptation of chondrocytes to low oxygen tension:
Relationship between hypoxia and cellular metabolism. J Cell Physiol 1996;168:424–32.
32. De Sousa CAF, Sodek L. Alanine metabolism and alanine aminotransferase activity in
soybean (Glycine max) during hypoxia of the root system and subsequent return to normoxia.
Environ Exp Bot 2003;50:1–8.
33. Harada H. Hypoxia-inducible factor 1-mediated characteristic features of cancer cells for
tumor radioresistance. J Radiat Res 2016;57:i99–105.
34. Harada H, Inoue M, Itasaka S et al. Cancer cells that survive radiation therapy acquire
HIF-1 activity and translocate towards tumour blood vessels. Nat Commun 2012;3, DOI:
10.1038/ncomms1786.
35. Friebe A, Müllershausen F, Smolenski A et al. YC-1 Potentiates Nitric Oxide- and Carbon
Monoxide-Induced Cyclic GMP Effects in Human Platelets. Mol Pharmacol 1998;54:962–7.
36. Teng C. YC-1 , a nitric oxide-independent activator of soluble guanylate cyclase , inhibits
platelet-rich thrombosis in mice. 1997:161–6.
37. Li SH, Shin DH, Chun Y-S et al. A novel mode of action of YC-1 in HIF inhibition:
stimulation of FIH-dependent p300 dissociation from HIF-1 . Mol Cancer Ther 2008;7:3729–
38.
38. Moon SY, Chang HW, Roh JL et al. Using YC-1 to overcome the radioresistance of
40
39. Harada H, Itasaka S, Zhu Y et al. Treatment regimen determines whether an HIF-1
inhibitor enhances or inhibits the effect of radiation therapy. Br J Cancer 2009;100:747–57.
40. Chapman JD. The detection and measurement of hypoxic cells in solid tumors. Cancer
1984;54:2441–9.
41. Troost EGC, Laverman P, Philippens MEP et al. Correlation of [18F]FMISO
autoradiography and pimonodazole immunohistochemistry in human head and neck
carcinoma xenografts. Eur J Nucl Med Mol Imaging 2008;35:1803–11.
42. Rischin D, Hicks RJ, Fisher R et al. Prognostic significance of [18F]-misonidazole
positron emission tomography-detected tumor hypoxia in patients with advanced head and
neck cancer randomly assigned to chemoradiation with or without tirapazamine: A substudy
of Trans-Tasman Radiation Oncolog. J Clin Oncol 2006;24:2098–104.
43. Wittig A, Sauerwein WA, Coderre JA. Mechanisms of transport of
p-borono-phenylalanine through the cell membrane in vitro. Radiat Res 2000;153:173–80.
44. Yoshimoto M, Honda N, Kurihara H et al. Non-invasive estimation of
10B-4-borono-L-phenylalanine-derived boron concentration in tumors by PET using
4-borono-2-18F-fluoro-phenylalanine. Cancer Sci 2018;109:1617–26.
45. Hayashi K, Anzai N. Novel therapeutic approaches targeting L-type amino acid
41 9. 図 図1. ホウ素中性子捕捉療法の原理 熱中性子とホウ素原子との核変換反応によって生じる、ヘリウム原子核 (α 線) とリチウム原子核を用いた放射線治療。発生する α 線およびリチウム原 子核は高LET を有し、飛程がそれぞれ 9 μm および 5 μm と短いため、がん 細胞内に選択的にホウ素原子が集積することで、近接する正常細胞を傷つけ ることなくがん細胞のみを傷害することが可能となる。 図2. ホウ素薬剤 (左: 10B-BSH、右:10B-BPA )
42 図3. 研究プロトコル (A) 通常酸素および低酸素条件における評価 (B) 通常酸素条件下での DFO による擬似低酸素条件における評価 図4. siRNA の実験プロトコル 10 cm ディッシュで培養中の細胞が 70%のコンフルエント状態に達した ところで細胞を回収し、各細胞の増殖率に応じて 10 cm ディッシュに 3.0-5.0×105個で播種した。DFO 投与群には、終濃度で 5 μM の DFO を投 与した。
43 図5. DFO の細胞毒性評価 (A)T98G、(B) HSC-3、(C) MCF-7 細胞の DFO 非投与サンプルに対する 細胞数の割合。(D)T98G、(E) HSC-3、(F) MCF-7 細胞の 24 時間および 48 時間培養後の各 DFO 濃度における生細胞数の割合。各結果は、異な る2 回の測定の平均値で表されており、グラフ中のバーは標準偏差を表 している。
44 図6. ウェスタンブロッティングによるタンパク発現評価 T98G 細胞における HIF-1α のタンパク発現評価。通常酸素 (21% O2)、 低酸素 (1% O2)、および 5 μM の DFO 投与条件下で 24 時間培養した後、 ウェスタンブロッティングにより HIF-1α および β-アクチンのバンドを 検出した。内因性コントロールとして、β-アクチンを使用した。各バン ドの数値は通常酸素を基準とした定量値であり、画像解析ソフト内の解 析ツールを用いて求めた。
45
図7. 通常酸素、低酸素、および 5 μM の DFO 投与条件下における LAT1 の遺伝子発現量
(A)T98G、(B) HSC-3、(C) MCF-7 細胞における、通常酸素、および低酸 素条件下で24 時間培養後の LAT1 の mRNA 発現量。(D)T98G、(E) HSC-3、 (F) MCF-7 細胞における、通常酸素、および 5 μM の DFO 投与条件下で 24 時間培養後の LAT1 の mRNA 発現量。グラフのカラムおよびバーは 平均値±標準誤差を表している。目的遺伝子の mRNA 発現量は検量線 法を用いて定量した。また、ハウスキーピング遺伝子としてGAPDH を 使用した。統計的有意差は、通常酸素条件下で培養したサンプルに対し て、*P < 0.05、**P < 0.01 で示した。
46 図8. 通常酸素および 5 μM の DFO 投与条件下における細胞のホウ素取り込み量 (A)T98G、(B)HSC-3、(C)MCF-7 細胞を通常酸素 (21% O2)、および 5 μM の DFO 投 与条件下で24 時間培養した後、10B-BPA を 3.00 mM (30 μg10B/mL) の終濃度で投与 して10、30、60、120 分後の細胞内ホウ素量を ICP-AES で測定した。各細胞の結果 は、異なる3 回の測定の平均値で表されており、グラフ中のバーは標準誤差を表し ている。統計的有意差は、10B-BPA の各曝露時間において、通常酸素条件 (DFO 非 投与群) に対して、*P < 0.05、**P < 0.01 で示した。 (D)各細胞における10B-BPA 投 与 2 時間後のホウ素取り込み量の結果。(E)低酸素条件で 24 時間培養した細胞に 10B-BPA を 3.00 mM (30 μg10B/mL) の終濃度で投与して 2 時間後のホウ素取り込み量 の結果。グラフのカラムおよびバーは異なる3 回の測定結果から得られた平均値± 標準誤差で表されている。
47
図9. siRNA による HIF-1αのノックダウン効果
(A)T98G、(B)HSC-3、(C)MCF-7 細胞における HIF-1αの mRNA 発現量。 (D)T98G、(E)HSC-3、(F)MCF-7 細胞における LAT1 の mRNA 発現量。 siRNA を導入して 24 時間後に各細胞を再播種し、24 時間後に通常酸素 (21% O2)、および 5 μM の DFO 投与条件を導入した。24 時間培養後、定 量的リアルタイム RT-PCR にて mRNA を測定した。グラフのカラムお よびバーは異なる3 回の測定結果から得られた平均値±標準誤差で表さ れている。目的遺伝子の mRNA 発現量は検量線法を用いて定量した。 また、ハウスキーピング遺伝子としてGAPDH を使用した。統計的有意 差はsiRNA のコントロール群 (siControl) に対して、グラフ中に P 値と して示している。
48 図10. 低酸素細胞における BNCT 後の細胞生残率 (A)T98G、(B)HSC-3、(C)MCF-7 細胞を通常酸素 (21% O2)、および低酸 素 (1% O2) 条件下で 24 時間培養した後、10B-BPA を 3.00 mM (30 μg10B/mL) の終濃度で投与して 2 時間後に中性子を照射した。照射から 10-12 日後にコロニーカウントを行った。グラフのカラムおよびバーは 異なる3 回の測定結果から得られた平均値±標準誤差で表されている。 統計的有意差は、通常酸素条件で培養されたサンプルに対して、グラフ 中にP 値として示している。
49 図11. YC-1 による低酸素細胞に対する BNCT の効果の増感評価 (A)T98G、(B)HSC-3 細胞における YC-1 の細胞毒性評価。各細胞の結果は、異なる 2 回の測定の平均値で表されており、グラフ中のバーは標準偏差を表している。 (C)T98G、(D)HSC-3 細胞における YC-1 併用 BNCT 後の細胞生残率。0.5 μM の YC-1 を投与して低酸素 (1% O2) 条件下で 24 時間培養し、10B-BPA を 0, 1.00, 3.00 mM (そ れぞれ0, 10, 30 μg10B/mL) の終濃度で投与して 2 時間後に中性子を照射した。照射 から10-12 日後にコロニーカウントを行った。結果は、YC-1 非投与群、投与群とも に、10B-BPA 非投与の照射群のコロニー数で正規化して表している。T98G の結果は 異なる4 回の測定の平均値、HSC-3 の結果は異なる 3 回の測定の平均値でそれぞれ 表されており、グラフ中のバーは標準誤差を表している。
50
図12. HIF-1α の蓄積と LAT1 の発現低下のメカニズム
細胞膜上のLAT1 の構造は、参考文献[45] Hayashi Ket al. World J Gastrointest
51 10. 表 表1 PCR に使用したプライマー配列 遺伝子 プライマー配列 GAPDH SLC7A5 (LAT1) HIF1A F: 5’-GCACCGTCAAGGCTGAGAAC-3’ R: 5’-TGGTGAAGACGCCAGTGGA-3’ F: 5’-GCATCGGCTTCACCATCATC-3’ R: 5’-ACCACCTGCATGAGCTTCTGAC-3’ F: 5'-CTCATCAGTTGCCACTTCCACATA-3' R: 5'-AGCAATTCATCTGTGCTTTCATGTC-3' 表2 使用した siRNA の配列 遺伝子 配列 HIF1A_s HIF1A_as 5’-rCrArArArGUUrCrArCrCUrGrArGrCrCUrATT-3’ 5’-UrArGrGrCUrCrArGrGUrGrArArCUUUrGTT-3’ * s :センス鎖, as :アンチセンス鎖