1
Pharmacogenomics of Tamoxifen Therapy
Hiltrud Brauch1,2,a, Thomas E. Mürdter1,2, Michel Eichelbaum1,2 and Matthias Schwab1,2,3
1 Dr. Margarete Fischer-Bosch Institute of Clinical Pharmacology, Stuttgart, Germany; 2 University Tübingen,
Tübingen, Germany; 3 Department of Clinical Pharmacology, University Hospital Tübingen, Tübingen, Germany.
aAddress correspondence to this author at: Dr. Margarete Fischer-Bosch Institute of Clinical Pharmacology,
Auerbachstrasse 112, 70376 Stuttgart, Germany. Fax +49-(0)711-859295; e-mail [email protected]. Clin Chem 2009;55:1770-1782
タモキシフェン治療のゲノム薬理学
概要 背景:タモキシフェンは、ステロイドホルモン受容体陽性乳癌の予防、および治療の標準的な 内分泌治療に用いられる。 内容:タモキシフェンが薬理作用を示すためには、シトクロムP450 (CYP) 酵素による活性型代 謝産物、4-ヒドロキシタモキシフェンおよびエンドキシフェンへの変換が必要である。 代謝前 の薬剤と比べて両代謝物質は、エストロゲン受容体へのほぼ100 倍の親和性の増加、並びに細 胞増殖抑制能を有する。多様な形を持つCYP2D6 は、この生体内変換の重要な酵素であり、最 近の機能的、薬理学的、臨床的な研究結果から、遺伝子多型やCYP2D6 抑制剤による薬物相互 作用によって、活性型タモキシフェンの血漿中濃度や、タモキシフェンを服用している患者の 治療成果に影響を及ぼすことが示唆されている。特にCYP2D6 酵素活性欠損または極めて活性が低い(Poor Metabolizer)または相応に酵素活性が低い(Intermediated Metabolizer) CYP2D6 対
立遺伝子の有無は、高い再発率と関連している。
要約:従って、治療前にタモキシフェン代謝の状況を予測する目的で、CYP2D6 (シトクロム
P450、 family 2、subfamily D、polypeptide 6) の遺伝子型を決定することは、高い代謝能が期待さ れるかどうかが分かるメリットと共に、個別化された内分泌治療への新しい道を開くかもしれ
2 セチンやフルオキセチンのような強いCYP2D6 抑制剤 (顔面潮紅)は、活性型代謝産物の生成 に著しく悪影響を及ぼすので、使用を避けるべきである。 序文 タモキシフェンの生体内代謝にかかわる薬物代謝酵素のゲノム薬理学は、治療の開始前に乳癌 患者の治療効果を予測できる可能性を持っていることから関心が集まっている。もしタモキシ フェンのゲノム薬理学的パラダイムを証明することができれば、個々人に適した最も有効な治 療薬を選択したうえで、内分泌治療に適した患者にタモキシフェンを投与することができるだ ろう。多くの(特に閉経後の女性の)乳癌は、ホルモンレセプターが陽性である。したがって、
毎年世界中の何十万の女性が内分泌治療を始めている。Early Breast Cancer Trialist Collaborative
Group(EBCTCG)の結果に基づき、選択的エストロゲン受容体(Estrogen Receptor: ER)モジュ
レーターのタモキシフェンによる、5 年間の治療が標準勧奨法として採用されている(1)。現在、 タモキシフェンは早期乳癌や、ステロイドホルモン受容体陽性の転移乳癌の標準的なアジュバ ント療法の薬として、世界120 カ国以上で処方されている。早期乳癌においては、タモキシフ ェンによるアジュバント療法によって、閉経の前後にかかわらず患者の再発率および死亡率が ともに有意に減少し、この治療効果は投与後5 年でも変わらず、10 年以上経っても維持されて いる(1)。内分泌治療に反応性を示す閉経後の女性において、タモキシフェンはアロマターゼ阻 害剤(Aromatase Inhibitors:AIs)とともに有効な治療オプションであり(2)、ハイリスクな閉経 前女性患者の浸潤性乳癌および非浸潤性乳管癌(3)の予防や男性乳癌の治療(4)において標準的な 治療法であると考えられる。タモキシフェンは一般的にひろく容認されていて、顔面潮紅とい った更年期症状は最も一般的な副作用である。この薬剤による血栓塞栓症や子宮内膜癌のよう な、重篤な副作用の発生は稀である(1)。タモキシフェンの臨床的な効果については、30 年以上 のエビデンスの蓄積がある。しかし、タモキシフェンのアジュバント療法を受けた患者のなか で、最大50%の患者が癌特異的な抵抗性や患者の遺伝的素因により、再発や死亡する例がみら れる。 タモキシフェンのゲノム薬理学の領域はタモキシフェンの代謝学や代謝薬理学的知見をきっか けに一層弾みがかかった。この進展のきっかけは、タモキシフェンの主要な活性型代謝産物が
シトクロムP450(Cytochrome P450:CYP)(特に CYP2D6)によって生成され、この酵素がタ
モキシフェン代謝の遺伝型・表現型の多型の要因となることを明らかとした研究によるもので ある。 これまでの臨床研究の中には、表現型と遺伝子型の変異がタモキシフェンの臨床的効果との関 連を報告したものがあり、現在、国際的に両者の関係を明らかにする研究が進んでいる。 タモキシフェンの薬理ゲノム学的知見の臨床診療への将来的な移行の可能性の観点から、この 総説ではタモキシフェンの効果のメカニズム説明のための、基本的な薬理学的、遺伝学的、表 現型の原理を示す。
3 今回は臨床的に活性型代謝物質である4-ヒドロキシタモキシフェンおよび 4-ヒドロキシ-N-デス メチルタモキシフェン(エンドキシフェン)に重点を置いて、タモキシフェンの一次、二次代 謝体の生体内代謝に注目する。また、タモキシフェンはCYP2D6 が重要な役割を果たすことか ら、この総説ではCYP2D6 の遺伝子型と表現型の関係にも注目している。今回の考察には、 CYP2D6 阻害剤の表現型模写効果も含まれている。これはタモキシフェンの治療を受けている閉 経後の女性の顔面潮紅を緩和するためによく同時投与されるものである。これらの基礎研究に よって得られて知見により、 タモキシフェンのゲノム薬理学的研究についてのこれまでの文献への徹底した考察ができるだ けの科学的な背景が得られる。さらに、最終的にはタモキシフェンの効果が他の代謝酵素や代 謝以外の因子により影響を受ける可能性がある。これらのトピックスを踏まえて、この総説で は乳癌への内分泌治療薬の治療効果の向上を目指した新たな個別化医療の原理についての概略 を示したいと思う。
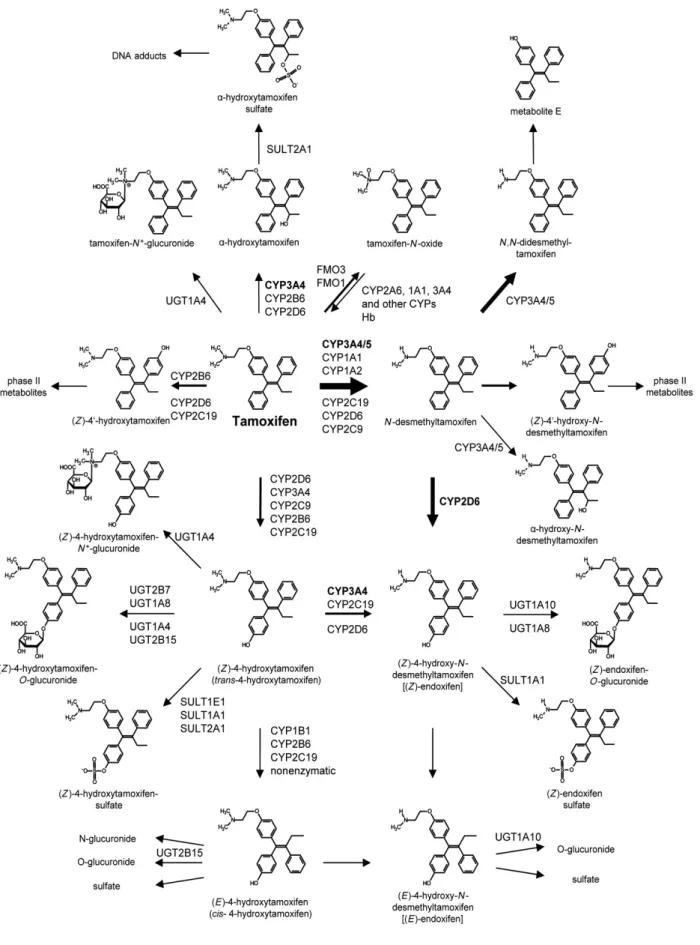
タモキシフェン代謝および活性型代謝体
tans-タモキシフェン {(Z)-2-[4-(1,2-diphenylbut-1-enyl)-phenoxy]-N,N-dimethyl-ethanamine}は、ヒ トの肝臓で第1 相反応、第 2 相反応を受ける。タモキシフェンの生体内変換には N-酸化、N-脱 メチル、水酸化反応がある。主要な代謝体であるN-デスメチルタモキシフェンは主としてCYP3A4 と CYP3A5 の触媒により形成され、CYP2D6、CYP1A1、CYP1A2、CYP2C19、CYP2B6 (5) (6) (7)といった酵素も関与する。毎日 20mg のタモキシフェンを摂取しそれを少なくとも 3 週 間継続した場合の定常状態におけるN-デスメチルタモキシフェン血漿濃度は、代謝される前の タモキシフェンの濃度の約2 倍になっている(前者が 100-290μg/L に対して後者は 72-160μ g/L)(8) (9) (10) (11) (12) (13) (14)。N-デスメチルタモキシフェンは主要な活性型代謝体であるエ ンドキシフェンの産生において水酸化(大部分はpara 位)の対象となるため、この事実は臨床 上最も重要なことである。さらに重要なことに、N-デスメチルタモキシフェンからエンドキシ フェンへの変換はほとんどがCYP2D6 よって触媒される (15) (16)。エンドキシフェンの血漿濃 度は、CYP2D6 が変異型のホモ接合体患者では平均で 8.1μg/L(n=51)、野生型のホモ接合体 の患者では20.7μg/L(n=55)の幅が観察された (17)。 他に臨床的に活性型である代謝体には4-ヒドロキシタモキシフェンがある。これはタモキシフ ェンの4 位水酸化、また、原薬のフェニル環のパラ位での水酸化によっても生成される。この
変換は、CYP2D6、CYP3A4、CYP2C9、CYP2B6 および CYP2C19 を含む多くの CYP によって触
媒される (7) (18) (19) (20) (21)。endoxifen と比較して 4-ヒドロキシタモキシフェンの定常状態の
濃度はより低く、1.15μg/L から 6.4μg/L の幅を持つ (11) (14) (22)。endoxifen および 4-ヒドロキ
4
図 1. ヒトにおけるタモキシフェンの代謝変換
主要な代謝経路は太い矢印で強調されている。それぞれの代謝段階で優位に反応する酵素は太字 で示した。Hb, hemoglobin; FMO1, flavin-containing monooxigenase 1
5 さらに水酸化が別のフェニル環の4' 位で起こることで 4'-ヒドロキシタモキシフェンになる。 この反応は主としてCYP2B6 と CYP2D6 によって触媒される (7)。そして 4'-ヒドロキシ-N-デス メチルタモキシフェンになる。他の水酸化された代謝体である - ヒドロキシタモキシフェンは 主としてCYP3A4 によって生産される (5) (6) (23) (24)。 4 位に水酸化された代謝体は、 in vitro において化学的に異性化し、反応性 E か cis 異性体へ 変換する (25)。これらは弱い ER アンタゴニストである。 さらに、4-ヒドロキシタモキシフェン
の異性化はCYP1B1、CYP2B6 および CYP2C19 によって触媒される(7)。
注目すべきは、cis-4-ヒドロキシタモキシフェンの蓄積がタモキシフェン治療に対する抵抗を示した患者の腫瘍組織 で観察されたことである (26)。しかしながら、cis 異性体の血漿濃度に関するデータがわずかで あるため、この観察は予備的なものと見なされるかもしれない。4-ヒドロキシタモキシフェンの フェニル基がCYP3A4 および CYP2D6 によってさらなに水酸化を受けると、3,4-ジヒドロキシタ モキシフェンになる (27) 。この物質はタンパク質や DNA に共有結合により結合するため、タモ キシフェン治療と毒性や発癌性との関連が報告されている (28) (29)。 タモキシフェン代謝の他のルートはflavin-containing monooxigenase 1 と 3 によるタモキシフェ ンN-酸化物の生成で、これは CYP2A6、CYP1A1、CYP3A4 や他の酵素を含む多くの CYP によ ってタモキシフェンN-オキシドが還元されてタモキシフェンが生成する反応とともに起こる (30) (31)。しかしながら、分析的な視点から見て、この代謝体は試料調製の間の N-酸化物の化学 的還元の可能性のために無視することができません。なぜならタモキシフェンN-オキシドの定 量がタモキシフェン代謝体の分析において問題と見なされるかもしれないからである。これま でのところ、この問題に関するデータはほとんどなく、患者の血漿中のN-オキシドがタモキシ フェンの15%以下を占めることが示唆されている (32)。 第II 相のタモキシフェン代謝のレベルでは、硫酸化とグルクロン酸抱合が主なメカニズムであ る。4-ヒドロキシタモキシフェンの O-グルクロン酸抱合は、主として UDP-グルクロノシルトラ ンスフェラーゼ (UGTs) UGT1A4、UGT2B15、UGT2B7、UGT1A8、そして他の 4-ヒドロキシタ モキシフェン-O-グルノロニドを生産する酵素によって触媒される (33) (34) (35)。エンドキシフ ェンは、対応するO グルノロニドへの UGT1A10 と UGT1A8 によって優位にグルクロン酸抱合 される。注目すべきことには水酸基部位で第II 相反応を受けるヒドロキシル化された代謝体に 加えて、タモキシフェン自体が対応するN+グルクロニドにUGT1A4 によって抱合されるという ことである(36)(37)。いずれの N+グルクロニドも形成しないエンドキシフェンとは対照的に、4-ヒドロキシタモキシフェンはUGT1A4 によりアミノ基にグルクロン酸抱合を受けて 4-ヒドロキ
6 シタモキシフェン-N+-グルクロニドを生成する。4-ヒドロキシタモキシフェンとエンドキシフェ ンの硫酸化はスルフォトランスフェラーゼ(Sulfotransferase; SULTs)SULT1E1、SULT1A1、 SULT2A1 によって触媒される (32) (38)。4-ヒドロキシタモキシフェンおよびエンドキシフェン のE 異性体もまたこれらの抱合反応の基質となるが、異なるアイソザイムと異なる親和性を持 つと考えられる (33)。 - ヒドロキシタモキシフェンは SULT2A1 によって硫酸化され (39)、それ により生じる - ヒドロキシタモキシフェンサルファイトは DNA と共有結合をすることで発癌効 果を及ぼす可能性がある (40) (41)。 in vitro で同定されたタモキシフェンの代謝体は数多くあるが(図 1)、タモキシフェンで治療 された患者からの血漿サンプルによる in vivo での分析的な測定ではほとんどの代謝体は定量さ れておらず、測定されているものにはN-デスメチルタモキシフェン、エンドキシフェン、4-ヒ ドロキシタモキシフェン、N,N-ジデスメチルタモキシフェン、 - ヒドロキシタモキシフェン、 タモキシフェンN-オキシドがある(表 1)。
CYP2D6 の生化学および遺伝学
CYP2D6 は、多くの臨床的に重要な薬の代謝に関与し、この中にはβ-ブロッカー、不整脈治 療剤、抗高血圧剤、抗精神病薬、抗うつ薬、オピオイド他がある。米国の「Top 200 Drug」 (http://www.rxlist.com; 最も頻繁に処方した 200 の薬、April 2008)のための排泄経路の最近の分析により、CYP3A(37%)および CYP2C(33%)とともに 15%が CYP2D6 であることが示された
(42)。
ヒトCYP2D6 座位は第 22 番染色体上に位置し、ここには CYP2D6 遺伝子、偽遺伝子 CYP2D7P1
(シトクロムP450、ファミリー2、サブファミリーD、ポリペプチド 7、偽遺伝子 1)、
CYP2D7P2(シトクロム P450、ファミリー2、サブファミリーD、ポリペプチド 7、偽遺伝子 2 )、 CYP2D8P1(シトクロム P450、ファミリー2、サブファミリーD、ポリペプチド 8、偽遺伝子 1 )、 CYP2D8P2
7 表 1 タモキシフェンと代謝体 (シトクロムP450、ファミリー2、サブファミリーD、ポリペプチド 8、偽遺伝子 2 )を含ん でいて、これら偽遺伝子はもともと偽遺伝子CYP2D7 と CYP2D8 と記述されていた (43)。 CYP2D6 遺伝子は 9 つのエクソンおよび 8 つのイントロンからなり、また、その塩基配列は高度 に多型性を示す。
8
不整脈治療薬であるスパルテイン (44) や高血圧治療薬デブリソキンの投与例 (45) などの臨床
観察によって、最初のCYP2D6 表現型変異(スパルテイン/デブリソキン多型)が Extensive
Metabolizer (EM)から区別され る形で約 30 年前に同定され、Poor Metabolizer (PM) と命名された。
現在では、4つのCYP2D6 表現型は薬の酸化容量に基づいて、コーカサス系人種の中では一般
に、EM、Intermediated Metabolizer (IM)、PM、Ultrarapid Metabolizer (UM) の 4 種が観察されてい
る (46) (47) (48)。コーカサス系人種の間では、7−10%が PM で、10-15%は IM、活性スペクトル の反対端であるUM は 10%-15%以下である。 PM ステータスは、99%以上の確実性で 2 つとも機能を持たない対立遺伝子の存在や、20 以上同 定されているnull 対立遺伝子から推定することができる。したがって、遺伝子型決定によって 酵素の触媒機能の喪失といったCYP2D6 PM 表現型は表現型を見るまでもなく、患者の DNA の ジェノタイピングにより正確に予言することは可能である (42) (46) (48) (49)。EM 表現型は、*1 あるいは*2 のような野生型の機能を備えた、1 あるいは 2 の対立遺伝子変異の存在による。こ の表現型は、ホモ接合体性かヘテロ接合体性EM かを遺伝子型によって分離することができ、 これは1 あるいは 2 の機能的な対立遺伝子をもつかどうかに依存している。IM か PM アレルと ともに、1 つの*1 あるいは*2 の対立遺伝子をもつヘテロ接合性 EM では、酵素産生や酵素の機 能低下のため、彼らはIM に分類される。これは、ヘテロ接合体 EM がホモ接合体 EM に対して、 50%しか酵素産生能や触媒活性を持たないといった遺伝子量効果の仮定に基づくものである。し かしながら、この仮定は必ずしも正しくなく、酵素量や酵素活性ともにEM のホモ接合体とヘ テロ接合体の間には、実質的な重複部分が存在する。従って、遺伝子型にはやや貧弱な予測値 である。注目すべきは、IM には機能を損なった遺伝子発現および酵素機能(これらの変異は*9、 *10 および*41 を含んでいる)および(または)機能を持たない変異を含む (47) (52) ヘテロ接合性の EM とは、異なる表現型および遺伝子型があるということである (47) (50) (51) (52)。ドイツの人 口の2%-3%は重複したまたは多重の CYP2D6 遺伝子を持っていて、その結果、非常に高い酵素 活性(UM)を示す。酵素活性のこれらの差は、薬物代謝体の血漿濃度で三環系抗うつ薬ノルトリ プチリンにおいて観察されたような意味深い結果を示す。ノルトリプチリンを標準の一日量で ある100-150 mg で処方したとき、ノルトリプチリンの定常状態での血漿濃度は PM および UM の間で30 倍以上の差が観察された (53) (54)。しかしながら、UM の表現型に関して、コーカサ ス系人種の中で観察されたUM の表現型の 20%-30%だけが、遺伝子型決定によって識別可能で ある (46) (48) (55)。 これまでのところ、多くの個人についての系統的な遺伝子分析によって、100 以上の異なる対立 遺伝子が発見された [http://www.cypalleles.ki.se, (56)]。少なくともこれらの対立遺伝子のうちの 15 は機能を喪失した遺伝子産物をコード化していて、それらは異常なスプライシング、ナンセ ンスコドン、一塩基突然変異、小規模挿入または欠失、CYP2D6 遺伝子を含む部分の染色体の欠 損、CYP2D6/CYP2D7 ハイブリッド遺伝子、ヘム取り込みを欠除したり機能を持たない完全長タ ンパク質を産生したりするような、突然変異によって起こる。
9 PM、IM および UM の頻度に関して著しい人種の違いがあり、CYP2D6 基質の薬物治療の臨床成 果は人種によって異なる可能性がある。こういった背景の内で、ヨーロッパ系の人口に対して 遺伝子重複の頻度が、北東のアフリカ[例えばエチオピアの 29% (57)]やサウジアラビア[21% (58)]ではるかに高いことを認識することは重要である (59) (60)。しかしながら、アジアでは、 CYP2D6*10 に関連する IM が優勢であり (61)、漢族では 57%の頻度で、PM は少数派である (59)。 全体として、CYP2D6 の 遺伝子型と表現型との関係に対する認識は、特に有効な代替薬が入手 可能な時に、治療決定に影響を及ぼすかもしれない。経口で投与されたコデインの場合のよう に、コーカサス系人の10%を占める PM では、モルヒネに効率的に代謝されず、したがって、 鎮痛効果をほとんど得られないし、CYP2D6 PM か IM 遺伝子型/表現型を持った女性が、活性型 の代謝物の生成が不十分なために、タモキシフェンの抗エストロゲン効果を十分に得られない 可能性がある。UM の場合、コデイン治療の場合に代謝により迅速なモルヒネを産生するために、 より重度のオピオイドの副作用になり (62) (63)、UM の女性患者がタモキシフェン治療中に顔面 潮紅により感受性が高くなる可能性があることに気をつけることは重要である。
CYP2C9、2C19、2B6、3A4 および 3A5 の遺伝学
タモキシフェンの代謝活性化に関与するファミリー2 の他の重要な CYP アイソザイムは、CYP2C9、CYP2C19 および CYP2B6 であり(15) (18)、これらの酵素は多型性を示す。CYP2C9(シ
トクロムP450、ファミリー2、サブファミリーC、ポリペプチド 9)には 30 以上の変異対立遺伝 子があり、*2 および*3 対立遺伝子の異なる対立遺伝子は、徹底的に調査されており、両者は関 連性が高いが、基質に依存してそれぞれ固有のクリアランスで減少する (64)。*3 対立遺伝子は *2 より強く影響を受け、ある特定の薬剤によって、90%以下の酵素活性の減少がおこる (65)。 両方の対立遺伝子はコーカサス系人のおよそ35%にあるが、ネグロイドとアジアの人種におい ては、それほど優位ではない (42) (66)。コーカサス系人では約 2%はホモ接合体で、24%はヘテ ロ接合体である(67)。多数の臨床研究によって、薬物有害反応のより高い発生率との関連に関し て、CYP2C9 遺伝学の臨床上の重要性が実証された。最も顕著な例は抗凝固剤のワルファリンの 例で、CYP2C9 遺伝学が重度のワルファリンによる出血事象を減らすために、ワルファリン投薬 を調節するのに臨床的に役立つことが、いくつかの後ろ向き研究や前向き研究により実証され た (68) (69)。抗凝固性反応は、さらにビタミン K エポキシド還元酵素の遺伝学にも依存する(68)。 さらに非ステロイド系抗炎症薬による消化管出血 (70) やスルホニル尿素によって引き起こされ た低血糖症のような副作用 (71) も、また CYP2C9 多型と関係がある。 CYP2C19 遺伝子 (シトクロム P450、ファミリー2、サブファミリーC、ポリペプチド 19 ) につい
ては、既知のnull 対立遺伝子 (CYP2C19*2、*3、*4、*5、*6、*7 および*8) には CYP2C19 酵素
活性がなく (PM)、*2 対立遺伝子はコーカシアンにおいて優位である。 これらの null 対立遺伝子
10 は)安定の変化によるものである (72)(http://www.cypalleles.ki.se/)。最近、いくつかの新しい CYP2C19 対立遺伝子が異なる人種グループから同定された(*9- *25)。しかしながら、これらの 変化が生体内の酵素活性の著しい変化を起こすかどうかは明らかにされていない。 CYP2C19*2 と*の 3 は、最も頻度の高い変異である。遺伝子型および表現型の決定により示される結果や CYP2D6 との相似性によれば、PM の分配は広い民族間の違いを示す。コーカシアンのヨーロッ パ人では、PM の平均頻度は 3%であるが、一方では 23%もの高い PM 頻度がアジア/オセアニア 人集団で見出された (72) (73)。しかしながら、ヘテロ接合体の変異の保有者は、白人の 32%を占 める(74)。CYP2C19*17 のプロモーター変異は最近見出され、CYP2C19 基質であるオメプラゾー ル [プロトンポンプ阻害薬 (75)] や抗うつ薬のエシタロプラム(76) において UM の in vivo での CYP2C19 活性の増加に関係していることを示された。CYP2C19*17 対立遺伝子頻度の差が報告 され、スウェーデン人およびエチオピアではともに18% (75)、ドイツ人の 25% (77)、またポー ランド人口の27% (78)であった。中国人ではより低い頻度(4%)であることが報告されている (75)。 これらの遺伝子型/表現型関係から、私たちが乳癌でタモキシフェンの薬理遺伝学の研究を報告 したように(79)、CYP2C19 の UM がタモキシフェン代謝および臨床成果に役割を果たしてもよ い可能性がある。 CYP2B6(シトクロム P450、ファミリー2、サブファミリーB、ポリペプチド 6) に関して、最も一 般的な変異対立遺伝子である*6 が、異なる人口集団を通じて 15%-60%の頻度で生じている (80)。 CYP2B6*6 の遺伝子型決定により、エファビレンツとネビラピンの血漿濃度の増加および HIV 感染者の中のエファビレンツ関連の神経毒性を予言できた (81) (82)。また、この結果により CYP2B6*6 のホモ接合体だったアフリカの患者の中で、服用量を 35%減らすことを示唆した(83)。 これらの知見により、CYP2B6*6 アイソザイムがより低い活性にあることが示された。しかし、 それは基質に依存的かもしれない。現在、タモキシフェン治療成果へのCYP2B6 変異の貢献は 未知である。
ヒトの薬物代謝に関係する最も重要なCYP アイソザイム・サブファミリーは CYP3A4 と 3A5 で
あり、それは最も頻繁に処方される薬の40%の代謝に関与している(42)。不完全 CYP3A4 突然変 異体は非常にまれな場合において毒性を説明できるかもしれないが、CYP3A4 (シトクロム P450、 ファミリー3、サブファミリーA、ポリペプチド 4) の遺伝子発現および酵素機能に関連した寄与 を立証する根拠はほとんどない(84)。対照的に、遺伝的多型により CYP3A5(シトクロム P450、 ファミリー3、サブファミリーA、ポリペプチド 5 ) 表現型における変化の多くを定義できる。コ ーカシアン(85%-95%)対アフリカ系アメリカ人 (30%-50%)において不活発な変異である CYP3A5*3 のより高い発生率により、アフリカ系アメリカ人(>30%vs50%)と比較してコーカシア ンで、より低いCYP3A5 タンパク質レベルが引き起こされることが見られた。CYP3A5*6 と*の 7 は機能的な活性を欠き、もっぱらアフリカ起源の個人にみられる。免疫抑制剤タクロリムスに 対する明瞭な影響とは別に(85)、CYP3A を介した代謝への CYP3A5 酵素多型性の影響は、論争 の的になっている。 CYP3A4 と CYP3A5 のタンパク質構造、機能および基質が非常に類似して いるので、これらの相対的寄与度を線引きするのは難しい。実際、これらの酵素のうちの1 つ
11 は、機能的に他方の不足を補うかもしれない。CYP3A4 および(または)CYP3A5 多型がタモキシ フェンの治療成果に寄与するかどうかは未知である。
タモキシフェン・ゲノム薬理学
タモキシフェンのゲノム薬理学の原理の基礎となる論理的根拠は、減少したか酵素機能が欠損 したタンパク質をコードする薬物代謝酵素のDNA 塩基配列多型は、活性型タモキシフェン代謝 体の血漿濃度が低下することに関係しているかもしれないということであり、それはタモキシ フェン治療の効能に影響を及ぼすことができた。約30 年前に、ヨルダンらは最初の有力な抗エ ストロゲン代謝体産物(4-ヒドロキシタモキシフェン) の特徴として、原薬より ER に対して 100 倍の親和性を持つことを報告した (86)。この代謝産物は、エストロゲン依存の細胞増殖の抑制が タモキシフェンより30∼100 倍強であることがその後示された (86) (87) (88) (89)。抗エストロゲ ンとしての性能にもかかわらず、その血漿濃度がタモキシフェンおよび他の代謝産物と比較し て低いため、タモキシフェンの臨床的効果へのこの代謝物質の影響は、不明瞭なままであった (86)。エンドキシフェンの性質が明らかになったのち、タモキシフェンの代謝と治療反応の間の つながりについての、私たちの知識が急速に拡張した (16)(22)。エンドキシフェンについては、 それ自体は1980 年代の終わりには同定されたが、最初はその生物活性に関して不明瞭なままで あった。遂には、これらの薬理学的特徴づけに関する一連の実験室での研究により、エンドキ シフェンはER への結合親和力の点から 4-ヒドロキシタモキシフェンと同等な性能を持っている こと (16)、乳癌細胞のエストロゲン依存の増殖の抑制 (16) (89) (90)、またエストロゲンを介した 広範な遺伝子発現の制御( 91)することが立証された。 詳細な生体外の分析により、エンドキシ フェンは主として一次代謝体であるN-デスメチルタモキシフェンの 4 位水酸化によって生成さ れ、このCYP2D6 酵素がこの律速段階を触媒することが示された (15)。エンドキシフェンの生 成にCYP2D6 が支配的な役割を果たすことから、CYP2D6 遺伝子型および表現型における変異は、 タモキシフェンの薬理遺伝学の中核にある。この概念を立証する現在利用可能な証拠は、2 レベ ルの臨床試験であり、(a) 活性型タモキシフェン代謝体と CYP2D6 遺伝子型との関連性、あるい は活性型代謝産物と臨床成果との関連性、および (b) CYP2D6 遺伝子型および臨床成果との間の 関連性の探求によるものである。後者のアプローチから、2 つの機能的な CYP2D6 対立遺伝子を 持った患者が、タモキシフェン治療で最も効果的であったことが示された。in vivo のエンドキシ フェンの血漿濃度と臨床成果との関係の一層の解明は、さらに多くの患者コホートによる追加 の詳細な調査が必要となるだろう。タモキシフェン代謝体濃度の影響
アジュバントタモキシフェン治療の前向きコホート研究では、CYP2D6 遺伝子の変異をもつ女性 において、タモキシフェン治療中にタモキシフェン代謝体の生成においての広い個人間での差、 およびエンドキシフェンの定常状態での血漿濃度の大幅な減少を示した(8)(11)(22)。さらに、説12
得力のある証拠として、選択性セロトニン再吸収阻害剤(Selective Serotonin Reuptake Inhibitors:
SSRI)で強い CYP2D6 抑制剤であることが知られているパロキセチンおよびフルオキセチンに より、血漿エンドキシフェン濃度を低下することが示された。SSRI によって引き起こされたエ ンドキシフェン血漿濃度の著しい減少の表現型の類似性は、特に野生型のCYP2D6 遺伝子型の ホモ接合体乳癌患者において観察された。しかし他の代謝産物の濃度は、CYP2D6 遺伝子型/表 現型に影響されないままであった。CYP2D6 変異および血漿エンドキシフェン濃度の関係は、 CYP2D6*4 遺伝子型を持った PM 患者で最初に示されたが (11)、PM、IM および UM の遺伝子型 を含んだ定量的アプローチにより、この関係が実証された (8) 。 しかしながら、エンドキシフェ ン濃度は遺伝子型を通じてオーバーラップしていた。このことから他の要因が血漿エンドキシ フェンを調節するかもしれない。 CYP2D6 変異とエンドキシフェンの前駆体である N-デスメチルタモキシフェンが高濃度である こととの関係は、予防を目的とした化学療法にて報告された。N-デスメチルタモキシフェンの 著しく高い血漿濃度は、タモキシフェン治療の1 年後の突然変異保持者において報告され、こ れは臨床的に活性型のエンドキシフェンへの変換が障害されている可能性を示した(92)。 より最近の研究は、定常状態治療中にタモキシフェンの薬物動態学に対するSULT1A1 コピー数
の影響を含む、CYP2D6 および SULT1A1(sulfotransferase ファミリー、細胞質性、1A、フェノー
ルを好む、メンバー 1)の遺伝子型との関係が取り組まれた(32)。CYP2D6 および SULT1A1 遺伝 子型の両方は、タモキシフェン代謝物の薬物動態に影響を及ぼしたが、SULT1A1 のコピー数に は影響を及ぼさなかった。エンドキシフェンと4 ヒドロキシタモキシフェンの生成ではより低 い代謝率を示したが、エンドキシフェンの前駆体であるN-デスメチルタモキシフェン生成の代 謝では、高い比率がCYP2D6 変異遺伝子型の保持者で観察された。これは遺伝子量の効果と一 致する結果である。対照的に、高い酵素活性が予測される CYP2D6 対立遺伝子を持つ患者では、 両方の活性型代謝物で高い代謝率を示した。このような代謝率が臨床的に妥当であるかどうか はまだ決められていない。 同様に、韓国の早期または転移性乳癌患者に対する前向きコホートに関する研究では、IM CYP2D6*10 のホモ接合体変異と、定常状態における低い 4-ヒドロキシタモキシフェンやエンド キシフェンの血漿濃度との間に関連性が見出された (17)。 また、中国の研究で、CYP2D6*10 ホ モ接合体の患者で血清4-hydroxytamoxifen 濃度が低いことが見出された(93)。東アジアの CYP2D6*10 対立遺伝子の高い頻度は、IM 活性型代謝体の生成が障害された IM の関連性ととも に、コーカシアンの中でのCYP2D6 PM での結果を確認した。
タモキシフェン治療の臨床結果および予測
CYP2D6 変異と治療効果との関連を証明する最初の根拠は、ER 陽性乳癌に罹患した閉経後女性 患者への前向き無作為化III 相試験 (North Central Cancer Treatment Group aduvant breast cancer trial)13
において、5 年間のアジュバントタモキシフェン標準処方計画に 1 年間アンドロゲンフルオキシ
メステロンを加えた効果の調査の結果により得られた。タモキシフェンのみの群からの患者の
薬理遺伝学的調査により、平均11.4 年のフォローアップの後、CYP2D6*4 変異対立遺伝子が高
再発リスクおよび低い顔面潮紅発生率の独立予測因子だったことが示された(94)。追跡調査によ
り、CYP2D6 遺伝学に加えて、CYP2D6 抑制剤(SSRI)の同時処方による表現型の類似性が、タモ
キシフェンを服用する閉経後女性の乳癌経過の独立予測因子であることが分かった(95)。最近、 CYP2D6 遺伝子型および治療結果の間の強健な関連性は、アジュバントタモキシフェン治療を受 けるER 陽性の閉経後乳癌患者の、非無作為後ろ向きコホートの結果から得られた(79)。平均 71 か月のフォローアップでは、PM および IM 遺伝子型(つまり CYP2D6*4、*5、*10 および*41 対 立遺伝子の保持者)の保持者には、機能を持つ対立遺伝子保持者よりも著しく高い乳癌再発、短 い無再発時間およびより悪い無事象生存が認められた (Figure 2) 。このような関係はタモキシフ ェンで治療されない閉経後のER 陽性の患者では観察されなかった。興味深いことには、さらに UM CYP2C19*17 変異も持つ患者ではタモキシフェン治療結果に好ましい効果があった。ホモ接 合体の*17 遺伝子型を持った患者は、非*17 保持者より著しく少ない乳癌再発、長い無再発時間 およびより無事象生存がみられた。全体として、この研究は、CYP2D6*4、*5、*10 および*41 の遺伝子型決定が、アジュバントタモキシフェン治療から治療効果が得られない患者を識別す ることができるかもしれないことを示唆した。おそらくすべての患者の約50%を占める CYP2D6 EM 表現型は、タモキシフェンから利益を得るために患者を識別できるが、利益は完全 に機能的なCYP2D6 対立遺伝子および CYP2C19 UM の組み合わせを持った個人にとって最大と なる。後者はすべての患者の3 分の 1 にあてはまるが、タモキシフェン薬理遺伝学的問題が、 内分泌治療を受ける乳癌患者の本質的な一部分に関連性があることを示す。
14 朝鮮、中国および日本からの臨床研究は、悪い臨床結果と CYP2D6 遺伝学とをつなぎ合わせた。 予想通りに、IM CYP2D6*10 対立遺伝子の高い頻度を持つ集団については、韓国のコホートでの *10 ホモ接合体遺伝子型は転移性乳癌患者の悪い臨床効果と関連が見られた。しかし、*10 ヘテ ロ接合体および野生型のホモ接合体遺伝子型は、そうではなかった(17)。同様に CYP2D6*10 対 立遺伝子のホモ接合体だった中国の患者 (93) では、好ましくない無病生存率という点で関連を 示した。後に結果が正しかったことが、タモキシフェン治療のないコントロール患者グループ との比較を通じて確立された。ただ、この際、臨床結果とCYP2D6*10 変異との間の関連性は観 察されなかった。さらに、野生型のCYP2D6 を持った患者と比較して、アジュバントタモキシ フェン単独療法を受けた日本の乳癌コホートからのCYP2D6*10 ホモ接合体の患者は、10 年間の フォローアップ内に著しく高い再発率を示した(96)。遺伝子型と効果との相関性を実証するアジ アの研究では、いくつかのサンプルサイズが小さかったが、CYP2D6 遺伝子型からのタモキシフ ェンの効果予測の臨床意味合いは、他の報告に一致するものであった。 他方では、米国からの研究は、CYP2D6 遺伝学およびタモキシフェン結果の間に関連性がないこ とが報告された(97)。また、この関係と矛盾している結果が、スウェーデンから報告された。こ れはタモキシフェンに治療された患者において、CYP2D6*4 変異とよりよい臨床成果との関連が 見いだされたというものである(98)。さらに拡大した研究では、機能的な CYP2D6 対立遺伝子の ホモ接合体がヘテロ接合である患者と比較して、CYP2D6*4 保有者でよい無病生存率を示した (99)。 乳癌のタモキシフェン治療におけるCYP2D6 の役割の問題として、乳癌予防の観点からも取り
組まれている。例えば、Italian Tamoxifen Trail のデータによれば、CYP2D6*4/*4 遺伝子型を持っ
た女性が、化学療法としての予防的な薬としての目的で、タモキシフェンからあまり効果を得
ていないことが示唆されている。この知見は、CYP2D6 がタモキシフェンの代謝活性化および効
能に重要な役割を果たすという概念を支持する(100)。さらに、顔面潮紅がタモキシフェン効能
の独立予測因子かもしれないという「演繹的な」仮説が、Women’s Health Eating and Living
図2 CYP2D6 遺伝子型からの予測された Metabolizer 表現型のための乳癌患者の無再発時間
(RFT)の Kaplan-Meier 確率
(A)アジュバントタモキシフェン療法(TAM)で治療した患者。EM は障害された表現型(PM または IM)を持った患者より、著明に良好な RFT がみられた。(B)TAM を行わない患者は、CYP2D6 で
予測された表現型と、RFT との間に相違は示されなかった[Schroth ら (79)]。 hetEM (ヘテロ接合
体のEM)。もとは、Schroth、W らにより公表された:J Clin Oncol 2007;25(33): 5187-93。許可の
15 randomized trial により検証された(101)。タモキシフェンを服用する 864 人の患者のうち、674 人 (78%)は顔面潮紅を報告し、また、これらの患者の 12.9%は 7.3 年間のフォローアップの後に乳 癌の再発を経験した。しかし、顔面潮紅をなかった患者の21%はこの期間に乳癌が再発した。 顔面潮紅は年齢やホルモン受容体の状態や診断時の腫瘍のステージよりも、乳癌の特異的治療 成果の予測因子であったため、著者は副作用とタモキシフェン代謝および効能の間の関連性を
示唆した。最後に、BRCA1(Breast Cancer 1(初期の発病))あるいは BRCA2(Breast Cancer 2(初期の
発病))のいずれかに突然変異をもつもので、タモキシフェンで治療した家族性乳癌患者を対象と した小研究では、CYP2D6 PM ステータスと家族性乳癌中のより悪い生存率の関係があることを 示唆した(102)。しかしながら、この調査では患者数が少人数である点と同様に ER 陽性と ER 陰 性患者両方が含まれているため、さらなる研究による解明にはこれらのBRCA 突然変異の保有 者における貧弱な予後の結果からの、薬理遺伝学的効果と区別することが必要になるだろう。 有効なオプションとしてAIs を使用した ER 陽性閉経後乳癌患者に対して、長期的なエストロゲ ン阻害を行う現在の治療方針を得たため、アジュバント内分泌治療のための最適な選択への薬 理遺伝学的変異の効果に対する疑問は、モデリング分析で取り組まれた(103)。Markov モデルは 野生型のCYP2D6 遺伝子を持った患者のための最適な治療戦略が、CYP2D6*4 変異の保持者の ものと異なるかどうか調べるために作成された。この研究は、BIG1-98 trial からの患者を、情報 はこのTrial の再発の危険性から、対応する遺伝子型のデータは Goets らのものを使用した(94)。
AIs 代謝が CYP2D6 から独立しているという仮定の下では、モデルによって閉経後の CYP2D6 EM 患者においてアジュバントタモキシフェン治療の 5 年間の効果が、これまで一般的に知られ ているAI 治療さえもまさるかもしれないことを示唆している。
結論
:乳癌における CYP2D6 の臨床的妥当性
モデリング・データと同様に強い機能生理学、薬理学、臨床での科学的根拠によって、タモキ シフェン効能および臨床的効果が、CYP2D6 酵素多型や薬理学の相互作用によって、制御された CYP2D6 代謝に依存することが示された。国際的な研究データによって、活性型タモキシフェン代謝物の血漿濃度が、遺伝学的に決められるCYP2D6 metabolizer 状態や、強い CYP2D6 抑制剤
による表現型の類似性や臨床的効果とのつながりを常に実証してきた。わずかの矛盾するデー タは、患者の選択基準、タモキシフェン服用量、治療の長さ、追加の化学療法処方計画、ある いは一貫したER 試験の不足による研究のばらつきとして説明されるかもしれない。重要なこと には、ほとんどの著者は、SSRIs のような処方された同時服用薬剤による CYP2D6 の抑制と同様 に、CYP2D6 遺伝子変異もタモキシフェン代謝を減少させ、タモキシフェン効能および治療結果 に悪影響を及ぼすかもしれないということには賛成している。 CYP2D6 やタモキシフェン治療の結果の新しいデータから、高い可能性を秘めた多くの臨床使用 の結論がある。
16 まず、パロキセチンまたはフルオキセチンのような有力なSSRIs は、タモキシフェンを処方さ れている乳癌患者に顔面潮紅を除くためには使用されるべきではない。SSRIs は閉経期の血管拡 張性の徴候に対する、数少ない科学的根拠に基づく治療オプションのうちの 1 つであるが(104)、 説得力のあるデータにより、これらの薬がCYP2D6 に依存するタモキシフェン代謝に対する影 響に表現型が類似することで、タモキシフェン効能を危険にさらすかもしれないことが示され ている。 しかし、タモキシフェン代謝産物の血漿濃度の差は、CYP2D6 抑制剤の強さに依存し て観察された(11)(105)。顔面紅潮の治療が示される場合、citalopram または escitalopram のような SSRI あるいは venlafaxine のような、選択的ノルエピネフリン再取り込み抑制剤のような薬剤が 使用されるべきである。その理由はこれらの薬剤が、CYP2D6 の抑制を示さないためである。 2 番目として、CYP2D6 遺伝子型と表現型および治療成果との関係によって、アジュバント内分 泌治療に関する決定に先立つ一般的なCYP2D6 遺伝子型決定によって、利益が見込まれること が指摘されている。総合的に有効で、標準化され、クオリティーコントロールされたCYP2D6 遺伝子型決定分析は、タモキシフェン代謝に影響するかもしれない遺伝変異の確認のためにテ ストする必要があるだろう。Goetz らのデータ(94)や Schroth ら (79)のデータによれば、そのよう なアッセイには一般的なPM 対立遺伝子(CYP2D6*3、*4、*5)や、IM 対立遺伝子のための試 験を含むべきであり、これは個人の民族的起源に依存する。注目すべきは、*41 はヨーロッパ人 で、おもに*17 はアフリカ人で、また、*10 はアジア人(*9 も考慮される)で、最も頻繁な IM 対 立遺伝子である(59)。臨床応用に関しての他の関心分野としては、CYP2D6 表現型の代わりとし ての血漿エンドキシフェン濃度の測定がある。 代替治療オプション(つまりタモキシフェン vs AI)を行うとき、利用可能な科学的・臨床上の証 拠を考慮して、閉経後の乳癌患者の内分泌治療への個別化のアプローチが望ましい。一つは、 タモキシフェン単独療法はCYP2D6 EM および EM 保持者には十分有効であるが、一方で CYP2D6 変異対立遺伝子を持った閉経後の患者では、一般的な AI 治療を行うのがよいと推測す るかもしれない。このアプローチはPM 患者に直接的であると見なされるかもしれないが、IM 患者にとっては最善の治療法ではないかもしれない。IM は、コーカシアン、アフリカ系アメリ カ人およびアジア人を含む多くのエスニック・グループの人々の共通の表現型である。したが って、IM 遺伝子型を持つ患者の治療の限界と効能とを判断するための臨床的に重要なデータと して、タモキシフェン服用量の調節に関して質問を十分に行うよう求めている。同様に、投薬 にとって潜在的な意味合いを持つ代謝産物血中濃度、治療効能および毒性のUM 表現型への影 響についても、さらなる調査が求められる。治療決定に向けたCYP2D6 遺伝子型の診断基準の 公式な推奨法は、国際タモキシフェン薬理遺伝学コンソーシアム (http://www.pharmgkb.org/do/serve?objId=63&objCls=Project) によって試みられているような、より 大規模な後ろ向き研究または前向き臨床研究による、これらの遺伝子型の評価を待たなければ ならない。これまでのところ、遺伝的に起こりやすくなる4-ヒドロキシタモキシフェンおよび エンドキシフェン濃度の差が、治療反応や病態の進行、および表現型の類似性を含む顔面潮紅 のような副作用にたいして、どのように関係しているかという問題に取り組んでいる研究はな
17 い。したがってこれに代わる有用な方法としての薬物治療モニタリングは、タモキシフェンの 場合にはいまだ利用可能ではない。 CYP2D6 遺伝子型の決定が、ER 陽性の閉経後乳癌患者に適 切なアジュバント内分泌治療を選ぶための診断のツールになるかどうかは、CYP2D6 遺伝子型に よるタモキシフェン対AI 治療に対する、無作為化前向き臨床試験での評価を待つことになる。 そのようなプロスペクティブな臨床試験が現在計画されている。 他の未解決の問題点としては、人種によるバラツキと同様に、それらの治療効果と関連がある 遺伝子型において、他の代謝酵素や突然変異の臨床との関連性が問題になるかもしれない。最 後に薬物動態関連の遺伝子が、部分的にのみタモキシフェンのゲノム薬理学について説明して いる可能性がある。したがって、腫瘍細胞の特徴としてエストロゲン抵抗性を評価する際や、 エストロゲンを介した細胞増殖に関連した遺伝子の役割を探すときに、さらなる薬理作用と遺 伝子の影響を調べることは重要であろう。この総説では、タモキシフェンとの反応性に影響す るアンチ・エストロゲン耐性遺伝子(107)や、その変異体と同様に ER をコードする遺伝子や、こ れらのコアクチベーターやコリプレッサー(106)を突き止めることも興味深いだろう。これらの 結果により、タモキシフェンのゲノム薬理学の全体像の把握できる可能性を高めるかもしれな い。 このような目的のために、現在の使われているほとんどの癌治療法が、経験的に確立されたこ とを認識することは重要である。しかしながら、タモキシフェンの薬理学および遺伝学につい ての私たちの理解における最近の進歩は、テーラーメイド医療によって治療法を改良する見込 みが十分に期待される。ゲノムに基づいたアプローチは、患者のMetabolizer 表現型の予測のた めのCYP2D6 の遺伝子型決定に基づいているので、倫理の問題に十分に取り組む必要がある。 最も重要なことには、アジュバント内分泌治療に関するインフォームド・チョイスや、効能を 欠く治療を回避することがもっとも興味があることに違いない。したがって、患者とその治療 に携わる人々にこれらの問題点に気づかせて、専門家と議論を始めることは重要だろう。
謝辞
Author Contributions: All authors confirmed they have contributedto the intellectual content of this paper and have met the following3 requirements: (a) significant
contributions to the conceptionand design, acquisition of data, or analysis and interpretation of data; (b) drafting or revising the article for intellectualcontent; and (c) final approval of the published article.
Authors’ Disclosures of Potential Conflicts of Interest:Upon manuscript submission, all authors completed the Disclosuresof Potential Conflict of Interest form. Potential conflictsof interest:
18 Employment or Leadership: None declared. Consultant or Advisory Role: None declared. Stock Ownership: None declared.
Honoraria: None declared.
Research Funding: Robert Bosch Foundation, Stuttgart, Germany,and Bundesministerium für Bildung und Forschung Grant No.01ZP0502.
Expert Testimony: None declared.
Role of Sponsor: The funding organizations played no role inthe design of study, choice of enrolled patients, review andinterpretation of data, or preparation or approval of manuscript.
脚注
1 Nonstandard abbreviations: ER, estrogen receptor; CYP, cytochromeP450; UGT, UDP-glucuronosyltransferase; SULT, sulfotransferase;EM, extensive metabolizer; PM, poor metabolizer; IM, intermediatemetabolizer; UM, ultrarapid metabolizer; SSRI, serotonin reuptakeinhibitor; AI, aromatase inhibitor.
2 Human genes: CYP2D6, cytochrome P450, family 2, subfamily D,polypeptide 6; CYP2D7P1, cytochrome P450, family 2, subfamilyD, polypeptide 7 pseudogene 1; CYP2D7P2, cytochrome P450, family2, subfamily D, polypeptide 7 pseudogene 2; CYP2D8P1, cytochromeP450, family 2, subfamily D, polypeptide 8 pseudogene 1; CYP2D8P2,cytochrome P450, family 2, subfamily D, polypeptide 8 pseudogene2; CYP2C9, cytochrome P450, family 2, subfamily C, polypeptide9; CYP2C19, cytochrome P450, family 2, subfamily C, polypeptide19; CYP2B6, cytochrome P450, family 2, subfamily B, polypeptide6; CYP3A4, cytochrome P450, family 2, subfamily A, polypeptide4; CYP3A5, cytochrome P450, family 2, subfamily A, polypeptide5; SULT1A1, sulfotransferase family, cytosolic, 1A, phenol-preferring,member 1; BRCA1, breast cancer 1, early onset; BRCA2, breastcancer 2, early onset.
19
References
1. . Early Breast Cancer Trialists’ Collaborative Group (EBCTCG). Effects of
chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: an overview of the randomised trials. Lancet 2005;365:1687-1717. 2. Goldhirsch A, Wood WC, Gelber RD, Coates AS, Thürlimann B, Senn HJ. Progress
and promise: highlights of the international expert consensus on the primary therapy of early breast cancer 2007. Ann Oncol 2007;18:1133-1144.
3. Fisher B, Costantino JP, Wickerham DL, Cecchini RS, Cronin WM, Robidoux A, et al. Tamoxifen for the prevention of breast cancer: current status of the National Surgical Adjuvant Breast and Bowel Project P-1 study. J Natl Cancer Inst 2005; 97:1652-1662.
4. Fentiman IS, Fourquet A, Hortobagyi GN. Male breast cancer. Lancet 2006;367: 595-604.
5. Boocock DJ, Brown K, Gibbs AH, Sanchez E, Turteltaub KW, White IN. Identification of human CYP forms involved in the activation of tamoxifen and irreversible binding to DNA. Carcinogenesis 2002;23:1897-1901.
6. Coller JK, Krebsfaenger N, Klein K, Wolbold R, Nussler A, Neuhaus P, et al. Large interindividual variability in the in vitro formation of tamoxifen metabolites related to the development of genotoxicity. Br J Clin Pharmacol 2004;57:105-111.
7. Crewe HK, Notley LM, Wunsch RM, Lennard MS, Gillam EM. Metabolism of tamoxifen by recombinant human cytochrome P450 enzymes: formation of the 4-hydroxy, 4'-hydroxy and N-desmethyl metabolites and isomerization of trans-4-hydroxytamoxifen. Drug Metab Dispos 2002;30:869-874.
8. Borges S, Desta Z, Li L, Skaar TC, Ward BA, Nguyen A, et al. Quantitative effect of CYP2D6 genotype and inhibitors on tamoxifen metabolism: implication for
optimization of breast cancer treatment. Clin Pharmacol Ther 2006;80:61-74. 9. Dowsett M, Cuzick J, Howell A, Jackson I, . the ATAC Trialists’ Group.
Pharmacokinetics of anastrozole and tamoxifen alone, and in combination, during adjuvant endocrine therapy for early breast cancer in postmenopausal women: a
20
sub-protocol of the ‘ArimidexTM and Tamoxifen Alone or in Combination’ (ATAC)
trial. Br J Cancer 2001;85:317-324.
10. Hutson PR, Love RR, Havighurst TC, Rogers E, Cleary JF. Effect of exemestane on tamoxifen pharmacokinetics in postmenopausal women treated for breast cancer. Clin Cancer Res 2005;11:8722-8727.
11. Jin Y, Desta Z, Stearns V, Ward B, Ho H, Lee KH, et al. CYP2D6 genotype,
antidepressant use, and tamoxifen metabolism during adjuvant breast cancer treatment. J Natl Cancer Inst 2005;97:30-39.
12. Lee KH, Ward BA, Desta Z, Flockhart DA, Jones DR. Quantification of tamoxifen and three metabolites in plasma by high-performance liquid chromatography with fluorescence detection: application to a clinical trial. J Chromatogr B Analyt Technol Biomed Life Sci 2003;791:245-253.
13. MacCallum J, Cummings J, Dixon JM, Miller WR. Concentrations of tamoxifen and its major metabolites in hormone responsive and resistant breast tumours. Br J Cancer 2000;82:1629-1635.
14. Sheth HR, Lord G, Tkaczuk K, Danton M, Lewis LM, Langenberg P, et al. Aging may be associated with concentrations of tamoxifen and its metabolites in breast cancer patients. J Womens Health (Larchmt) 2003;12:799-808.
15. Desta Z, Ward BA, Soukhova NV, Flockhart DA. Comprehensive evaluation of tamoxifen sequential biotransformation by the human cytochrome P450 system in vitro: prominent roles for CYP3A and CYP2D6. J Pharmacol Exp Ther 2004; 310:1062-1075.
16. Johnson MD, Zuo H, Lee KH, Trebley JP, Rae JM, Weatherman RV, et al. Pharmacological characterization of 4-hydroxy-N-desmethyl tamoxifen, a novel active metabolite of tamoxifen. Breast Cancer Res Treat 2004;85:151-159. 17. Lim HS, Ju LH, Seok LK, Sook LE, Jang IJ, Ro J. Clinical implications of CYP2D6
genotypes predictive of tamoxifen pharmacokinetics in metastatic breast cancer. J Clin Oncol 2007;25:3837-3845.
18. Coller JK, Krebsfaenger N, Klein K, Endrizzi K, Wolbold R, Lang T, et al. The influence of CYP2B6, CYP2C9 and CYP2D6 genotypes on the formation of the
21
potent antioestrogen Z-4-hydroxy-tamoxifen in human liver. Br J Clin Pharmacol 2002;54:157-167.
19. Crewe HK, Ellis SW, Lennard MS, Tucker GT. Variable contribution of cytochromes P4502D6, 2C9 and 3A4 to the 4-hydroxylation of tamoxifen by human liver
microsomes. Biochem Pharmacol 1997;53:171-178.
20. Dehal SS, Kupfer D. CYP2D6 catalyzes tamoxifen 4-hydroxylation in human liver. Cancer Res 1997;57:3402-3406.
21. Mani C, Gelboin HV, Park SS, Pearce R, Parkinson A, Kupfer D. Metabolism of the antimammary cancer antiestrogenic agent tamoxifen. I. Cytochrome P-450-catalyzed N-demethylation and 4-hydroxylation. Drug Metab Dispos 1993;21:645-656. 22. Stearns V, Johnson MD, Rae JM, Morocho A, Novielli A, Bhargava P, et al. Active
tamoxifen metabolite plasma concentrations after coadministration of tamoxifen and the selective serotonin reuptake inhibitor paroxetine. J Natl Cancer Inst 2003;95: 1758-1764.
23. Kim SY, Suzuki N, Santosh Laxmi YR, Rieger R, Shibutani S. Alpha-hydroxylation of tamoxifen and toremifene by human and rat cytochrome P450 3A subfamily enzymes. Chem Res Toxicol 2003;16:1138-1144.
24. Notley LM, Crewe KH, Taylor PJ, Lennard MS, Gillam EM. Characterization of the human cytochrome P450 forms involved in metabolism of tamoxifen to its -hydroxy and ,4-dihydroxy derivatives. Chem Res Toxicol 2005;18:1611-1618.
25. Katzenellenbogen JA, Carlson KE, Katzenellenbogen BS. Facile geometric
isomerization of phenolic non-steroidal estrogens and antiestrogens: limitations to the interpretation of experiments characterizing the activity of individual isomers. J Steroid Biochem 1985;22:589-596.
26. Osborne CK, Wiebe VJ, McGuire WL, Ciocca DR, DeGregorio MW. Tamoxifen and the isomers of 4-hydroxytamoxifen in tamoxifen-resistant tumors from breast cancer patients. J Clin Oncol 1992;10:304-310.
27. Dehal SS, Brodie AMH, Kupfer D. The aromatase inactivator
4-hydroxyandrostenedione (4-OH-A) inhibits tamoxifen metabolism by rat hepatic cytochrome P-450 3A: potential for drug-drug interaction of tamoxifen and 4-OH-A in combined anti-breast cancer therapy. Drug Metab Dispos 1999;27:389-394.
22
28. Dehal SS, Kupfer D. Evidence that the catechol 3,4-dihydroxytamoxifen is a
proximate intermediate to the reactive species binding covalently to proteins. Cancer Res 1995;56:1283-1290.
29. Liu X, Pisha E, Tonetti DA, Yao D, Li Y, Yao J, et al. Antiestrogenic and DNA damaging effects induced by tamoxifen and toremifene metabolites. Chem Res Toxicol 2003;16:832-837.
30. Hodgson E, Rose RL, Cao Y, Dehal SS, Kupfer D. Flavin-containing monooxygenase isoform specificity for the N-oxidation of tamoxifen determined by product
measurement and NADPH oxidation. J Biochem Mol Toxicol 2000;14:118-120. 31. Parte P, Kupfer D. Oxidation of tamoxifen by human flavin-containing
monooxygenase (FMO) 1 and FMO3 to tamoxifen-N-oxide and its novel reduction back to tamoxifen by human cytochromes P450 and hemoglobin. Drug Metab Dispos 2005;33:1446-1452.
32. Gjerde J, Hauglid M, Breilid H, Lundgren S, Varhaug JE, Kisanga ER, et al. Effects of CYP2D6 and SULT1A1 genotypes including SULT1A1 gene copy number on tamoxifen metabolism. Ann Oncol 2008;19:56-61.
33. Nishiyama T, Ogura K, Nakano H, Ohnuma T, Kaku T, Hiratsuka A, et al. Reverse geometrical selectivity in glucuronidation and sulfation of cis- and
trans-4-hydroxytamoxifens by human liver UDP-glucuronosyltransferases and sulfotransferases. Biochem Pharmacol 2002;63:1817-1830.
34. Ogura K, Ishikawa Y, Kaku T, Nishiyama T, Ohnuma T, Muro K, Hiratsuka A. Quaternary ammonium-linked glucuronidation of trans-4-hydroxytamoxifen, an active metabolite of tamoxifen, by human liver microsomes and
UDP-glucuronosyltransferase 1A4. Biochem Pharmacol 2006;71:1358-1369.
35. Sun D, Sharma AK, Dellinger RW, Blevins-Primeau AS, Balliet RM, Chen G, et al. Glucuronidation of active tamoxifen metabolites by the human UDP
glucuronosyltransferases. Drug Metab Dispos 2007;35:2006-2014.
36. Kaku T, Ogura K, Nishiyama T, Ohnuma T, Muro K, Hiratsuka A. Quaternary ammonium-linked glucuronidation of tamoxifen by human liver microsomes and UDP-glucuronosyltransferase 1A4. Biochem Pharmacol 2004;67:2093-2102.
23
37. Sun D, Chen G, Dellinger RW, Duncan K, Fang JL, Lazarus P. Characterization of tamoxifen and 4-hydroxytamoxifen glucuronidation by human UGT1A4 variants. Breast Cancer Res 2006;8:R50.
38. Falany JL, Pilloff DE, Leyh TS, Falany CN. Sulfation of raloxifene and
4-hydroxytamoxifen by human cytosolic sulfotransferases. Drug Metab Dispos 2006; 34:361-368.
39. Apak TI, Duffel MW. Interactions of the stereoisomers of alpha-hydroxytamoxifen with human hydroxysteroid sulfotransferase SULT2A1 and rat hydroxysteroid sulfotransferase STa. Drug Metab Dispos 2004;32:1501-1508.
40. Kim SY, Laxmi YR, Suzuki N, Ogura K, Watabe T, Duffel MW, Shibutani S. Formation of tamoxifen-DNA adducts via O-sulfonation, not O-acetylation, of alpha-hydroxytamoxifen in rat and human livers. Drug Metab Dispos 2005;33:1673-1678.
41. Osborne MR, Hewer A, Phillips DH. Resolution of alpha-hydroxytamoxifen; R-isomer forms more DNA adducts in rat liver cells. Chem Res Toxicol 2001;14: 888-893.
42. Zanger UM, Turpeinen M, Klein K, Schwab M. Functional
pharmacogenetics/genomics of human cytochromes P450 involved in drug biotransformation. Anal Bioanal Chem 2008;392:1093-1108.
43. Zanger UM. The CYP2D subfamily. Ioannides C eds. Cytochromes P450: role in the metabolism and toxicity of drugs and other xenobiotics 2008:p 241-275 Royal Chemical Society London.
44. Eichelbaum M, Spannbrucker N, Dengler HJ. Proceedings: N-oxidation of sparteine in man and its interindividual differences. Naunyn Schmiedebergs Arch Pharmacol 1975;287(Suppl):R94.
45. Mahgoub A, Idle JR, Dring LG, Lancaster R, Smith RL. Polymorphic hydroxylation of debrisoquine in man. Lancet 1977;2:584-586.
46. Griese EU, Zanger UM, Brudermanns U, Gaedigk A, Mikus G, Morike K, et al. Assessment of the predictive power of genotypes for the in-vivo catalytic function of CYP2D6 in a German population. Pharmacogenetics 1998;8:15-26.
24
47. Raimundo S, Toscano C, Klein K, Fischer J, Griese EU, Eichelbaum M, et al. A novel intronic mutation, 2988G>A, with high predictivity for impaired function of
cytochrome P450 2D6 in white subjects. Clin Pharmacol Ther 2004;76:128-138. 48. Sachse C, Brockmoller J, Bauer S, Roots I. Cytochrome P450 2D6 variants in a
Caucasian population: allele frequencies and phenotypic consequences. Am J Hum Genet 1997;60:284-295.
49. Marez D, Legrand M, Sabbagh N, Guidice JM, Spire C, Lafitte JJ, et al.
Polymorphism of the cytochrome P450 CYP2D6 gene in a European population: characterization of 48 mutations and 53 alleles, their frequencies and evolution. Pharmacogenetics 1997;7:193-202.
50. Raimundo S, Fischer J, Eichelbaum M, Griese EU, Schwab M, Zanger UM.
Elucidation of the genetic basis of the common ‘intermediate metabolizer’ phenotype for drug oxidation by CYP2D6. Pharmacogenetics 2000;10:577-581.
51. Toscano C, Klein K, Blievernicht J, Schaeffeler E, Saussele T, Raimundo S, et al. Impaired expression of CYP2D6 in intermediate metabolizers carrying the *41 allele caused by the intronic SNP 2988G>A: evidence for modulation of splicing events. Pharmacogenet Genomics 2006;16:755-766.
52. Zanger UM, Fischer J, Raimundo S, Stuven T, Evert BO, Schwab M, Eichelbaum M. Comprehensive analysis of the genetic factors determining expression and function of hepatic CYP2D6. Pharmacogenetics 2001;11:573-585.
53. Bertilsson L, Dahl ML, Sjöqvist F, Aberg-Wistedt A, Humble M, Johansson I, et al. Molecular basis for rational megaprescribing in ultrarapid hydroxylators of
debrisoquine. Lancet 1993;341:63.
54. Dalen P, Dahl ML, Bernal Ruiz ML, Nordin J, Bertilsson L. 10-Hydroxylation of nortriptyline in white persons with 0, 1, 2, 3, and 13 functional CYP2D6 genes. Clin Pharmacol Ther 1998;63:444-452.
55. Lovlie R, Daly AK, Matre GE, Molven A, Steen VM. Polymorphisms in CYP2D6 duplication-negative individuals with the ultrarapid metabolizer phenotype: a role for the CYP2D6*35 allele in ultrarapid metabolism?. Pharmacogenetics 2001;11: 45-55.
25
56. Algeciras-Schimnich A, O'Kane DJ, Snozek CL. Pharmacogenomics of tamoxifen and irinotecan therapies. Clin Lab Med 2008;28:553-567.
57. Aklillu E, Persson I, Bertilsson L, Johansson I, Rodrigues F, Ingelman-Sundberg M. Frequent distribution of ultrarapid metabolizers of debrisoquine in an Ethiopian population carrying duplicated and multiduplicated functional CYP2D6 alleles. J Pharmacol Exp Ther 1996;278:441-446.
58. McLellan RA, Oscarson M, Seidegard J, Evans DA, Ingelman-Sundberg M. Frequent occurrence of CYP2D6 gene duplication in Saudi Arabians. Pharmacogenetics
1997;7:187-191.
59. Sistonen J, Sajantila A, Lao O, Corander J, Barbujani G, Fuselli S. CYP2D6
worldwide genetic variation shows high frequency of altered activity variants and no continental structure. Pharmacogenet Genomics 2007;17:93-101.
60. Ingelman-Sundberg M. Genetic polymorphisms of cytochrome P450 2D6
(CYP2D6): clinical consequences, evolutionary aspects and functional diversity. Pharmacogenomics J 2005;5:6-13.
61. Bradford LD. CYP2D6 allele frequency in European Caucasians, Asians, Africans and their descendants. Pharmacogenomics 2002;3:229-243.
62. Gasche Y, Daali Y, Fathi M, Chiappe A, Cottini S, Dayer P, Desmeules J. Codeine intoxication associated with ultrarapid CYP2D6 metabolism. N Engl J Med 2004; 351:2827-2831.
63. Koren G, Cairns J, Chitayat D, Gaedigk A, Leeder SJ. Pharmacogenetics of morphine poisoning in a breastfed neonate of a codeine-prescribed mother. Lancet 2006; 368:704.
64. Lee CR, Goldstein JA, Pieper JA. Cytochrome P450 2C9 polymorphisms: a
comprehensive review of the in-vitro and human data. Pharmacogenetics 2002;12: 251-263.
65. King BP, Khan TI, Aithal GP, Kamali F, Daly AK. Upstream and coding region CYP2C9 polymorphisms: correlation with warfarin dose and metabolism. Pharmacogenetics 2004;14:813-822.
26
66. Garcia-Martin E, Martinez C, Ladero JM, Agundez JA. Interethnic and intraethnic variability of CYP2C8 and CYP2C9 polymorphisms in healthy individuals. Mol Diagn Ther 2006;10:29-40.
67. Xie HG, Kim RB, Wood AJ, Stein CM. Molecular basis of ethnic differences in drug disposition and response. Annu Rev Pharmacol Toxicol 2001;41:815-850.
68. Flockhart DA, O'Kane D, Williams MS, Watson MS, Flockhart DA, Gage B, et al. Pharmacogenetic testing of CYP2C9 and VKORC1 alleles for warfarin. Genet Med 2008;10:139-150.
69. Limdi NA, Veenstra DL. Warfarin pharmacogenetics. Pharmacotherapy 2008;28: 1084-1097.
70. Pilotto A, Seripa D, Franceschi M, Scarcelli C, Colaizzo D, Grandone E, et al. Genetic susceptibility to nonsteroidal anti-inflammatory drug-related gastroduodenal bleeding: role of cytochrome P450 2C9 polymorphisms. Gastroenterology 2007; 133:465-471.
71. Holstein A, Plaschke A, Ptak M, Egberts EH, El-Din J, Brockmoller J, Kirchheiner J. Association between CYP2C9 slow metabolizer genotypes and severe hypoglycaemia on medication with sulphonylurea hypoglycaemic agents. Br J Clin Pharmacol
2005;60:103-106.
72. Desta Z, Zhao X, Shin JG, Flockhart DA. Clinical significance of the cytochrome P450 2C19 genetic polymorphism. Clin Pharmacokinet 2002;41:913-958. 73. Xie HG, Stein CM, Kim RB, Wilkinson GR, Flockhart DA, Wood AJ. Allelic,
genotypic and phenotypic distributions of S-mephenytoin 4'-hydroxylase (CYP2C19) in healthy Caucasian populations of European descent throughout the world.
Pharmacogenetics 1999;9:539-549.
74. Wedlund PJ. The CYP2C19 enzyme polymorphism. Pharmacology 2000;61:174-183.
75. Sim SC, Risinger C, Dahl ML, Aklillu E, Christensen M, Bertilsson L, Ingelman-Sundberg M. A common novel CYP2C19 gene variant causes ultrarapid drug metabolism relevant for the drug response to proton pump inhibitors and antidepressants. Clin Pharmacol Ther 2006;79:103-113.
27
76. Rudberg I, Mohebi B, Hermann M, Refsum H, Molden E. Impact of the ultrarapid CYP2C19*17 allele on serum concentration of escitalopram in psychiatric patients. Clin Pharmacol Ther 2008;83:322-327.
77. Justenhoven C, Hamann U, Pierl CB, Baisch C, Harth V, Rabstein S, et al.
CYP2C19*17 is associated with decreased breast cancer risk. Breast Cancer Res Treat 2008;115:391-396.
78. Kurzawski M, Gawronska-Szklarz B, Wrzesniewska J, Siuda A, Starzynska T, Drozdzik M. Effect of CYP2C19*17 gene variant on Helicobacter pylori eradication in peptic ulcer patients. Eur J Clin Pharmacol 2006;62:877-880.
79. Schroth W, Antoniadou L, Fritz P, Schwab M, Muerdter T, Zanger UM, et al. Breast cancer treatment outcome with adjuvant tamoxifen relative to patient CYP2D6 and CYP2C19 genotypes. J Clin Oncol 2007;25:5187-5193.
80. Zanger UM, Klein K, Saussele T, Blievernicht J, Hofmann MH, Schwab M.
Polymorphic CYP2B6: molecular mechanisms and emerging clinical significance. Pharmacogenomics 2007;8:743-759.
81. Rotger M, Colombo S, Furrer H, Bleiber G, Buclin T, Lee BL, et al. Influence of CYP2B6 polymorphism on plasma and intracellular concentrations and toxicity of efavirenz and nevirapine in HIV-infected patients. Pharmacogenet Genomics 2005; 15:1-5.
82. Rotger M, Tegude H, Colombo S, Cavassini M, Furrer H, Decosterd L, et al. Predictive value of known and novel alleles of CYP2B6 for efavirenz plasma concentrations in HIV-infected individuals. Clin Pharmacol Ther 2007;81:557-566.
83. Nyakutira C, Röshammar D, Chigutsa E, Chonzi P, Ashton M, Nhachi C,
Masimirembwa C. High prevalence of the CYP2B6 516G T(*6) variant and effect on the population pharmacokinetics of efavirenz in HIV/AIDS outpatients in Zimbabwe. Eur J Clin Pharmacol 2008;64:357-365.
84. Westlind-Johnsson A, Hermann R, Huennemeyer A, Hauns B, Lahu G, Nassr N, et al. Identification and characterization of CYP3A4*20, a novel rare CYP3A4 allele without functional activity. Clin Pharmacol Ther 2006;79:339-349.