先天性インプリント異常症と生殖補助医療との関連
性についての検討
著者
服部 裕充
学位授与機関
Tohoku University
学位授与番号
11301甲第19125号
URL
http://hdl.handle.net/10097/00129220
博士論文
先天性インプリント異常症と
生殖補助医療との関連性についての検討
東北大学大学院医学系研究科
医科学専攻
環境遺伝医学総合研究センター
情報遺伝学分野
服部 裕充
2
目
次
要 約 ... P.3-6 研究背景と目的... P.7-12 研究方法 ... P.13-19 研究結果 ... P.20-25 考 察 ... P.26-33 結 論 ... P.34 謝 辞 ... P.35 参考文献 ... P.36-46 図表の説明... P.47-54 図 表 ... P.55-72 略 語 ... P.73-743
<要約>
【研究の背景・目的】 近年の初婚年齢の上昇(晩婚化)の社会情勢に伴い、生殖補助医療(Assisted reproductive technologies:ART)の需要が国内外で益々高まってきている。ART によ る出生数は、年々増加傾向を示し、本邦では 2015 年の日本産科婦人科学会データで は年間出生児数の5.07%、つまり 20 人に 1 人の割合にまで増加している。ART では、排卵誘発、体外受精(In vitro fertilization:IVF)、顕微授精(Intracytoplasmic sperm injection:ICSI)、体外培養、受精卵凍結などの操作が行われ、一定の効果を上げてい る。しかし、ART による妊娠では、本来は発症が非常に稀である先天性インプリント 異常症の発症頻度が、増加しているという数多くの報告がみられる。ゲノムインプリ ンティングとは、父親または母親由来のどちらか一方の親由来の遺伝子のみが発現す るように、あらかじめ発現の記憶が刷り込まれた現象のことである。また、ゲノムイ ンプリンティングは、おもに、DNA のメチル化によって遺伝子発現が制御されてい る。DNA のメチル化は、受精から初期胚までの発生過程において変動する。この初期 発生の時期は、環境変化に対して最も脆弱かつ不安定であるため、ゲノムインプリン ティングの異常が起こりやすいと考えられている。本研究では、本邦におけるART と 先天性インプリント異常症の関連について実態調査を行うとともに、ART 由来患者の 臨床像とDNA メチル化変異の特徴を明らかにすることを目的とした。
4
【研究方法】
全国の小児科および産婦人科施設の協力下に、先天性インプリント異常症である Beckwith-Wiedemann 症候群(BWS)、Angelman 症候群(AS)、Prader-Willi 症候群(PWS)、 Silver-Russell 症候群(SRS)の 4 疾患を対象とし、患者数、臨床像、ART 治療内容お
よび遺伝子診断の有無などについて調査を実施した(期間:2015 年 8 月から 2017 年
7 月)。次に、ART 由来の SRS 患者(ART-SRS) 5 名、自然妊娠由来の SRS 患者(非 ART-SRS)5 名、および自然妊娠由来の健常者 10 名の末梢血 DNA を用い、RRBS (Reduced representation bisulfite sequencing)法による網羅的 DNA メチル化(メチロ
ーム)解析を行い、ART-SRS のメチル化変異の特徴について統計学的な解析を行った。 【研究結果】 全国調査では、2777 施設の調査対象機関のうち、1957 施設から有効回答が得られ た(回答率70.5%)。先天性インプリント異常症 4 疾患の総患者数は 931 名であり、そ の内訳は、BWS が 117 名、AS が 227 名、PWS が 520 名、SRS が 67 名であった。こ のうちART 由来患者の割合は、BWS が 6.0%(7/117)、AS が 1.8%(4/227)、PWS が 4.6%(24/520)、SRS が 11.9%(8/67)であり、AS 以外の疾患は、本邦の総出生数(1985 年から2015 年)における ART 由来児の出生率(1.34%)よりもその頻度が高かった。
5 めていた[BWS:42.9%(3/7)、AS:25.0%(1/4)、PWS:58.3%(14/24)、SRS:75.0% (6/8)]。ART 由来患者の臨床像は、非 ART 由来患者と類似していたが、若干の相違 点も見られた。たとえば、AS では、ART 群の不眠症の割合が 100%(4/4)に対し、 非ART 群では 46.2%(103/223)であった(P =0.048)。また、PWS では、ART 群では 色素異常症と小肢端症の割合が、それぞれ91.7%(22/24)、87.5%(21/24)と高く、非 ART 群は 69.0%(342/496)、58.3%(289/496)であった(P =0.020、P=0.005)。SRS で は、ART 群の心奇形の割合が 25.0%(2/8)に対し、非 ART 群では 3.4%(2/59)であ った(P=0.036)。また、腫瘍形成および糖尿病の発生は、ART 群のみにしか認められ なかった。ART-BWS および ART-SRS では、疾患発症の原因は、全例が責任遺伝子領 域のDNA メチル化の異常(4/4 および 5/5)であった。
ART-SRS、非 ART-SRS を対象としたメチローム解析の結果では、DNA メチル化変 異(DNA methylation variation:DMV)を 7.5%以上のメチル化レベル変化かつ false discovery rate(FDR)< 0.05 と定義した場合、ART-SRS 群では、ほとんどの領域にお
いて非 ART-SRS 群より、DMV の割合が有意に高かった(FDR<0.001)。また、DMV
が多く認められた領域は、精子由来ゲノムで高メチル化を示し、受精直後に脱メチル 化を受ける領域であった。
6 【考察および結論】 本研究により、BWS、SRS、PWS の先天性インプリント異常症の発症頻度の増加が ART と関連していることが推測された。これらの疾患のうち、BWS と SRS の発症原 因は、DNA メチル化の異常であった。メチローム解析の結果、ART-SRS 群の DMV は、ゲノム全体にランダムに広がり、脱メチル化よりもメチル化されている割合が多 かった。このことは、ART 由来児の先天性インプリント異常症が、多彩な臨床像を示 すことと関連するのかもしれない。また、ART-SRS 群では、精子由来ゲノムの脱メチ ル化を受ける領域で、DMV が起こりやすかったことから、DMV が生じた時期は、受 精直後に影響を受けた可能性が高いことが推測された。すなわち、ART 操作のうち、 受精直後の体外操作や体外培養が、先天性インプリント異常症の発生リスクと関連す る可能性が高いことが示唆された。最近、胎児期、乳幼児期のエピゲノム変異が、成 人期の生活習慣病(Developmental Origins of Health and Disease︓DOHaD 学説)と関連
する数多くの報告がみられる。今後、ART 由来児を対象とした大規模かつ長期的な健
7
<研究の背景と目的>
1.初期胚発生過程におけるダイナミックなメチル化の変調 通常、体細胞におけるエピゲノムは、細胞分裂後も安定に維持される。しかし、生 殖細胞では、それぞれ特異的なエピゲノムを有する卵子および精子のゲノムは、受精 後の初期胚発生過程において、エピゲノムがダイナミックに変化し、ゲノムの「初期 化」が起こる。このゲノムの初期化は、リプログラミングと呼ばれている。リプログ ラミングでは、DNA のメチル化はゲノム全体にわたって消去される(脱メチル化) が、卵子と精子由来のゲノムでは異なるメチル化の変化を示すことが報告されている 1-3)(図1)。マウスの場合、受精後、精子由来ゲノムは、TET3(Ten-eleven translocation-3)酵素により 5-メチルシトシン(5-methlcytosine:5mC)から 5-ヒドロキシメチルシ トシン(5-hydroxymethylcytosine:5hmC)に変換され、脱メチル化される(能動的脱 メチル化)4)。一方、卵子由来ゲノムは、母性タンパク質によってTET3 からの脱メチ ル化から保護されている 5)。ただし、DNA 複製の際に、DNA 鎖をメチル化させる維持型DNA メチル基転移酵素 DNMT1(DNA methyltransferase 1)の大部分が、細胞質
内に存在するので、細胞核内に存在する DNA のメチル基を維持できず、細胞分裂の
たびにDNA メチル化が失われる(受動的脱メチル化)6)。ヒトの場合もマウスと同様
に、精子由来ゲノムは能動的脱メチル化を受け、卵子由来ゲノムは不完全ながら、受
8 2.ゲノムインプリンティング 受精後のゲノム全体に起こる脱メチル化に対し、抵抗性を示す領域がある。その一 つが、ゲノムインプリンティングを受ける領域である。哺乳類のゲノムには、父親ま たは母親由来のどちらか一方の親由来の遺伝子のみが発現するように、あらかじめ発 現の記憶が刷り込まれた対立遺伝子が存在し、その記憶をもとに子での遺伝子発現が 制御される。この現象は、『ゲノムインプリンティング』と呼ばれる。また、この片親 性発現を示す遺伝子は、インプリント遺伝子と呼ばれ、その発現は生殖細胞系列特異 的メチル化領域(Germline-derived differentially methylated region:gDMR)の DNA メ
チル化によって調節されている1, 11)。マウスの研究から、卵子や精子のもとの細胞で ある始原生殖細胞は、生殖隆起に到達する胎齢 11.5-12.5 日目までに gDMR のメチル 化の消去が起こる12)。卵子では、第一減数分裂前期の一次卵母細胞の成長期で、メチ ル化が生じ、インプリンティングが確立する。一方、精子では、出生前後の胎齢 14.5-19.5 日までの減数分裂前の前精祖細胞で、メチル化が生じ、インプリンティングが確 立する13, 14)。つまり、卵子および精子の生殖細胞では、受精前にインプリンティング が確立され、受精後の胚では、初期発生過程におけるゲノム全体におこる脱メチル化 の影響を受けずに、インプリンティングは保持される15)。
9
3.生殖補助医療と先天性インプリント異常症
近年、晩婚化の社会情勢と生殖医療技術の進歩により、体外受精(In vitro fertilization: IVF)や顕微授精(Intracytoplasmic sperm injection:ICSI)を用いた生殖補助医療(Assisted reproductive technologies:ART)が急速に普及している。本邦では、治療件数が、増加 の一途をたどり、2015 年には年間 51,001 人の ART 由来の出生児(ART 由来児)が誕 生している16)。ART が普及した要因として、培養液の改良による受精卵の長期培養技 術の開発や、ガラス化による受精卵の凍結技術の開発などにより、妊娠率が飛躍的に 向上したことが挙げられる。 一方、ART の普及に伴い、本来発症が非常に稀である Beckwith-Wiedemann 症候群
(Beckwith-Wiedemann syndrome:BWS)、 Angelman 症候群(Angelman syndrome:AS)、 Prader-Willi 症候群(Prader-Willi syndrome:PWS)、Silver-Russell 症候群(Silver-Russell syndrome:SRS)といった先天性インプリント異常症の発生頻度の増加が報告されて いる17-22)。しかしながら、ART とこれらの疾患発症の関係については、否定的な報告 23, 24)もあり、未だ不明な点が多い。しかし、マウス、ウシ、ヒツジの動物実験による 研究では、ART がインプリンティングに影響を及ぼすことが、多数報告されている 25-27)。ART による排卵誘発、配偶子操作、受精操作、体外培養、受精卵凍結などは、イ ンプリンティングが確立および維持される時期に行われるため、DNA メチル化の異 常を引き起こすことが推測されている28)。ART による増加が懸念されている、BWS、
10 SRS、AS、PWS の発症頻度は、1 万人から 10 万人に 1 人であり、BWS では巨舌、過 成長、SRS では成長障害、特徴的な身体所見、AS では精神運動発達遅延、てんかん、 PWS では筋緊張低下、精神運動発達遅延などの臨床像が認められ診断しやすい29)(表 1)。特に SRS については、1.子宮内胎児発育不全、2.成長障害、3.相対的頭囲拡 大、4.前額部突出、5.左右非対称、6.摂食障害の 6 つの症状のうち 4 つ以上が認 められると臨床的に SRS と診断される、臨床診断スコアリングシステムが確立され ている30)。これらの疾患は臨床像や遺伝子診断による鑑別が比較的容易であることか ら、代表的な先天性インプリント異常症とされており、疫学調査の対象として患者数 の推定や臨床像の比較に適している。また、それぞれの発症原因は、1)片親性ダイソ ミー(Uniparental disomy:UPD) 2)遺伝子(領域)の欠失 3)遺伝子変異 4)メチ ル化異常に分類される(表1)。このうち、ART 由来患者の場合、インプリント遺伝子 の gDMR のメチル化異常を病因とした症例が多いという報告もある(BWS では KCNQ1OT1-gDMR の低メチル化、SRS では H19-gDMR の低メチル化、AS では SNRPN-gDMR の低メチル化)31-37)。以前、Hiura らは、ART 由来 BWS および SRS 患者では、 インプリント異常症の責任領域のgDMR 以外に、複数のインプリント遺伝子の gDMR
のメチル化異常(Multi-locus imprinting disturbances:MLID)が多く認められることを
11
メチル化と低メチル化のモザイクパターンを示し、メチル化異常が複雑で不完全であ
12 4.研究目的 現在、本邦の ART に由来する先天性インプリント異常症は、出生児の追跡調査が 容易ではないため、その患者数や病態などの実態は明らかではない。本研究では、ART とインプリンティング異常との関連性を調べるため、先天性インプリント異常症の実 態に関する全国多施設による調査を実施し、ART により出生した先天性インプリント 異常症の患者数や臨床像を明らかにした。また、ART と自然妊娠(非 ART)由来の SRS 患者を対象とした、RRBS 法による網羅的な DNA メチル化解析を行い、ART が もたらすDNA メチル化の変異の特徴について検討した。これらの成果は、ART によ り出生した児の健康リスクと安全性の評価に繋がるものと考える。
13
<研究方法>
1. 倫理審査 東北大学医学系研究科倫理委員会に本研究計画を提出し、承認を得てから研究を開 始した。また、適宜計画の変更や修正を行い、疫学研究と実験研究の両方について倫 理委員会の承認を得た(受付番号 2015-1-130, 2015-1-735, 2017-1-880, 2018-1-81)。疫 学調査における個々の症例に関する情報は、匿名化して通し番号をつけて収集した。 患者名と通し番号の対応表は、それぞれの医療機関にて厳重に保管してもらった。す べての研究は、ヘルシンキ宣言に基づいて実施した。 2. 先天性インプリント異常症の全国規模の実態調査 全国疫学調査は、日本医療研究開発機構(AMED)難治性疾患実用化研究事業「先 天性インプリント異常症の診断と生殖補助医療の安全性評価」研究開発代表者名:有 馬隆博(AMED: JP17ek0109132h0003)から助成を受け、厚生労働省難治性疾患克服研 究事業特定疾患の疫学に関する研究班の刊行による「難病の患者数と臨床疫学像把握 のための全国疫学調査マニュアル第 2 版(厚労省、永井班)」に則って実施した 38)。 調査は、2015 年 8 月から 2017 年 7 月に行い、対象は、病院要覧、医療機関名簿を参 考に、全国の国公立病院の小児科および産婦人科とした。調査は二段階に分け、一次 調査では4 つの先天性インプリント異常症(BWS、SRS、PWS、AS)の患者数を集計14
し、二次調査では臨床像の調査を行った。BWS の臨床診断は国際基準 39)、SRS の臨
床診断はNetchine-Harbison clinical scoring system30)を用いて行い、PWS、AS について は染色体検査、遺伝子検査、臨床的表現型を基に、臨床医が診断した。二次調査では、 出産時の両親年齢、患者年齢、性別、発育状況、臨床症状、遺伝子検査の有無、診断
方法、不妊治療の有無と不妊治療の方法(排卵誘発/タイミング法、人工授精、IVF、
ICSI)について質問票を作成し、主治医に回答をお願いし、郵送にて回収した。質問
票で得られた患者は、不妊治療の有無で、ART 由来の患者(ART-BWS、-SRS、-PWS、
-AS)と非 ART 由来の患者(非 ART-BWS、-SRS、-PWS、-AS)に分類した。
3. 全ゲノム DNA メチル化(メチローム)解析 (1)対象患者 二次調査でSRS の受診の回答が得られた施設に血液検体提供の依頼をし、ART-SRS (0-5 歳、男 3 名、女 2 名)、非 ART-SRS(2-5 歳、男 2 名、女 3 名)、自然妊娠由来の 健常児(3-11 歳、男 5 名、女 5 名)(対照群)を対象としてメチローム解析を行った。 担当主治医は、家族および本人に、本研究の内容について説明を行い、同意または代 諾を得てから採血を行った。採血は3-5ml 採取し、血液は東北大学に移送した。
15
血液から、DNA Extractor WB-Rapid Kit(富士フィルム和光純薬工業社:293-54803) を用いて白血球の抽出を行った。白血球のペレットに、Lysis buffer(50 mM Tris-HCl, 20 mM EDTA,100 mM NaCl,1% SDS)と Proteinase K(New England Biolabs 社:P8107S) を加え、ゲノムDNA を溶解させた後、PureLink RNase A (Thermo Fisher Scientific 社: 12091039)による RNase 処理、フェノール/クロロホルム処理、エタノール沈降によ り精製し、適量のTE buffer (10 mM Tris [pH8.0],1 mM EDTA [pH8.0])に溶解した。 得られたゲノムDNA 溶液は、NanoDrop ND-1000(LMS 社:NanoDrop1000)を用い て吸光度を測定し、濃度50 ng/μl 以上、吸光度 260/280 値が 1.8 以上、260/230 値が 2.0 以上であることを確認し、Qubit ds DNA HS Kit(Life Technologies 社:Q32851)に
よりDNA の定量を行った。精製したゲノム DNA は、-80℃で保存した。
RRBS ライブラリーの調整法は、既報を基に一部変更して行った40)(図2)。具体
的には、20ng のゲノム DNA に、制限酵素 MspI(New England Biolabs 社:R0106S) と10x NEBuffer2(New England Biolabs 社:B7002S)を加え、37℃で 16 時間インキ
ュベート後、80℃で 20 分間のインキュベートにより酵素を失活させた。この DNA
断片溶液に、10x NEBuffer2 と Nucleotide End-Repair mix (1 mM dATP, 0.1 mM dCTP, 0.1 mM dGTP) 、Klenow fragment(3’→5’ exo−, New England Biolabs 社:M0212)を 加え、末端平滑化及び3’末端アデニン付加反応(end-repaired/dA-tailing reaction)を 30℃ 20 分間、次いで 37℃ 20 分間のインキュベートを行い、その後 75℃ 20 分間の
16
インキュベートにより酵素を失活させた。平滑化したDNA 断片と 1 µM pre-annealed
methylated adaptors に、T4 DNA Ligase(New England Biolabs 社:M0202)と Ligase Reaction Buffer(New England Biolabs 社:B0202S)を加え、さらに 16℃にて 16 時間
インキュベートによりライゲーションを行った後、65℃にて 20 分間インキュベート
して酵素を失活させた。次に、この反応液をNuSieve 3:1(Lonza 社:50090)で作成
した3%アガロースゲルにて 50V の電圧で電気泳動を行い、150-350bp の DNA 断片
を切り出し、MinElute Gel Extraction Kit(Qiagen 社:28604)を用いて DNA を精製 し、EB buffer(Qiagen 社:19086)で溶出した。精製した DNA 断片は、EpiTect Bisulfite Kit(Qiagen 社:59104)を用いて、バイサルファイト反応に供した。インデ ックス導入およびライブラリー増幅のためのPCR は、KAPA HiFi HotStart Uracil+ ReadyMix(Kapa Biosystems 社:KK2800)を用いて行った。PCR の条件は、95℃で 2
分間の熱変性後、98℃ 20 秒間の熱変性、65℃ 30 秒間のアニーリング、72℃ 30 秒
間の伸長反応を14 サイクル行い、最後に 72℃ 1 分間の伸長反応を行った。その
後、RRBS ライブラリーは、Agencourt AMPure XP beads(Beckman Coulter 社: A63880)にて 2 度精製し、Qubit および KAPA Library Quantification Kit(Kapa Biosystems 社:KK4828)にて定量した。シークエンスは TruSeq SR Cluster Kit v3-cBot-HS(Illumina 社:GD-401-3001)および TruSeq SBS Kit v3-HS(Illumina 社:FC-401-3001)を用い、HiSeq 2500 platform(Illumina 社)による 101-bp single-end reads
17
を行った。シークエンスリードは、Illumina standard base-calling pipeline(v1.8.2)に
より処理後、cutadapt(1.18)を用いてアダプター配列を除去した。さらに、FASTX
Trimmer(v0.0.13.2)にてリードの最初および最後の 4 塩基を除去後、Bismark (v.0.9.0)を用いてリファレンスゲノムである Human Genome Assembly
(GRCh37/hg19)の常染色体にマッピングした 41)。不完全なバイサルファイト変換
リードを除くため、リード内のnon-CpG シトシンのバイサルファイト変換効率を算
出し、変換効率90%以上のリードを解析対象とした。それぞれの CpG シトシンのメ
チル化レベルは、Bismark methylation extractor にて算出し、5 リード以上のシークエ
ンスリードがカバーされたCpG シトシンを解析した。
(3)ゲノム領域のアノテーション
シークエンスされたリードをリファレンスゲノムにマッピングするアノテーショ ンを行った。既知の配列であるRefseq gene とリピート配列は、UCSC Genome Browser (https://genome.ucsc.edu/)からダウンロードした。ただし、300bp 以下の Refseq genes (microRNAs、small nucleolar RNAs など)は、解析から除外した。プロモーター領域
は、Refseq 転写産物の転写開始点から上流および下流のそれぞれ 500bp の計 1kb と定
義した。平均メチル化レベルの計算は、10 以上の CpG シトシンが含まれ、5 リード
以上のシークエンスリードが含まれるゲノム領域にて行った。対象とするゲノム領域 は、プロモーター領域、遺伝子領域、エクソン領域、イントロン領域、遺伝子間領域、
18 インプリントgDMR、CpG アイランド領域(CpG サイトが集中して存在する領域で、 低メチル化状態にある)、CpG アイランドショア領域(CpG アイランド近傍の約 2kb 以内の領域)および CpG アイランドシェルフ領域(CpG アイランドショアの外側の 約2kb 以内の領域)と、短鎖散在反復配列(SINE:レトロトランスポゾンの一種で、 300bp 以下の短い塩基配列を持つ散在性の反復配列)、長鎖散在反復配列(LINE:レ トロトランスポゾンの一種で、長い塩基配列を持つ散在性の反復配列)、長鎖末端反 復配列(LTR:レトロトランスポゾンの一種で、配列の両末端に長い繰り返し配列を 持つ)、DNA 反復配列、複合型レトロトランスポゾン(SVA)および単純反復配列と した。21 ヵ所のインプリント gDMR の位置と領域名は、既報に従って定義した 42)。 4. 統計解析
R または JMP Pro 13.1.0(SAS Institute 社)を用いて統計解析を行った。疫学調査に おける臨床的特徴の解析は、Fisher’s exact test または Mann–Whitney U test(P<0.05) を用いた。DNA メチル化解析は、Mann–Whitney U test with the Benjamini-Hochberg correction を用い、7.5%以上のメチル化レベルの変化を示し、かつ false discovery rate (FDR)< 0.05 を示す領域を DNA メチル化変異(DNA methylation variation:DMV) とした。DMV の数の比較は、Fisher’s exact test を用いた。DMV が認められた遺伝子 は 、The Database for Annotation, Visualization and Integrated Discovery ( DAVID )
19
(https://david.ncifcrf.gov/)43)を用いて遺伝子オントロジー(Gene ontology:GO)解析 を行った。また、Fisher’s exact test with the Benjamini-Hochberg correction により多重比
較補正を行い、生物学的プロセスの GO term のうち、P<0.05 を有意な結果として抽
出した。
5. シークエンスデータの登録
本研究で得られたデータは、DDBJ Japanese Genotype-phenotype Archive データベー
20
<研究結果>
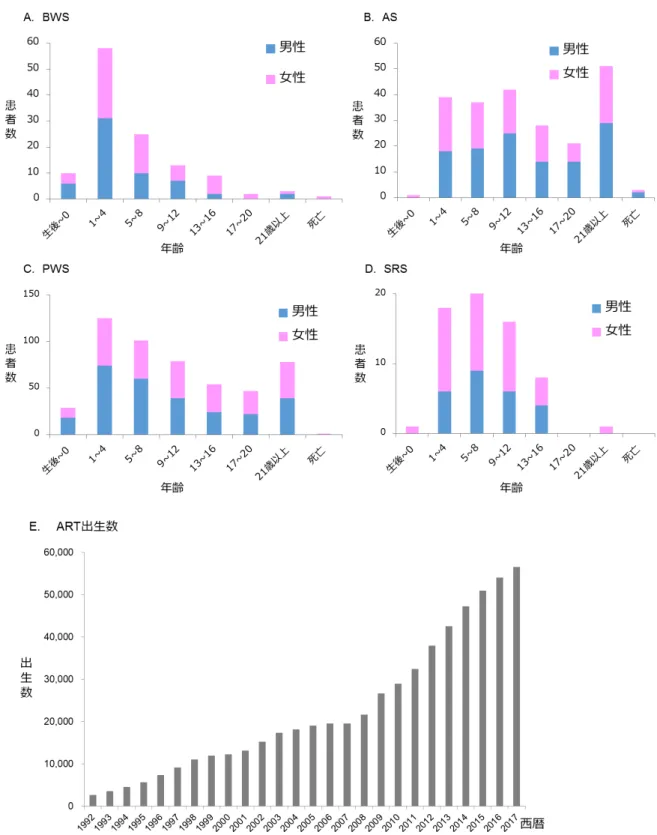
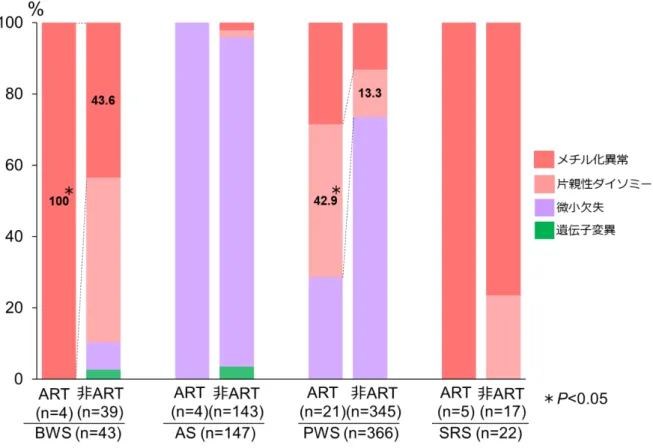
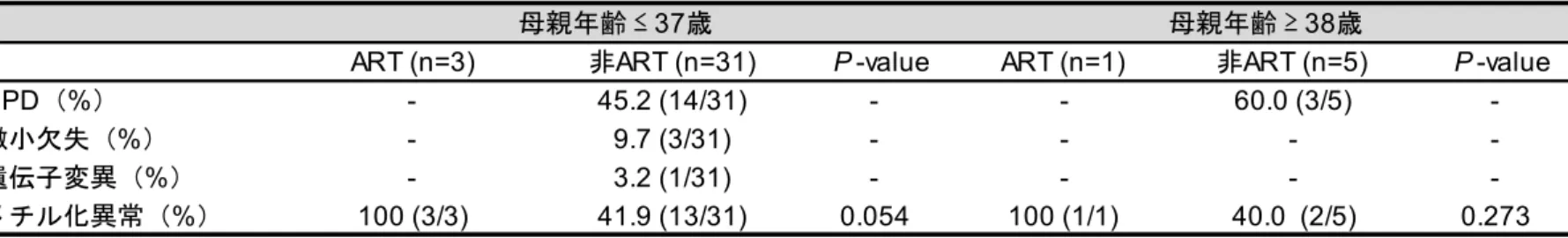
1. 先天性インプリント異常症と ART との関連性 まず、わが国の先天性インプリント異常症と ART との関連に関する実態調査を行 った。全国の国公立病院(療育センター、重症心身障害施設を含む)を 2777 の施設 を対象に、調査票による4 疾患(BWS、AS、PWS、SRS)の受療患者数の実態の把握 を行った。一次調査では、先天性インプリント異常症 4 疾患の有無について質問し、 1957 施設から回答を得た(回答率 70.5%)(表 2)。一次調査にて先天性インプリント 異常症の患者が受診していると回答したのは476 施設で、二次調査では、これらの施 設を対象に患者の年齢、性別などの質問票調査を行った。その結果、273 施設から回 答を得た(回答率57.4%)。患者の重複や欠測を除き、4 つの疾患の総計は、931 人で、 その内訳はBWS が 117 人、AS が 227 人、PWS が 520 人、SRS が 67 人であった。疾 患数の地域差については地域の施設数と回答率に偏りがあり、不明であった。それぞ れの疾患の患者数、年齢、性別と日本産科婦人科学会が公表している年別の ART 由 来児数を、図3 にまとめた。その結果、BWS、PWS、SRS の患者は、増加傾向がみら れ、特にBWS は増加が顕著であった。さらに、これらの疾患の増加は、ART 出生数 の増加率と類似していた。また、性差はいずれの疾患も有意な違いは認められなかっ た。不妊治療を受けた患者の割合は、BWS が 6.0%(7/117)、AS が 1.8%(4/277)、 PWS が 4.6%(24/520)、SRS が 11.9%(8/67)であった(表 3)。疾患別に不妊治療由21 来患者の年齢、性別、出生時両親年齢、ART の治療内容を表 4 にまとめた。ART の治 療内容のうち、IVF または ICSI により出生した患者の割合は、SRS が 75.0%(6/8)で 最も多く、次にPWS が 58.3%(14/24)、BWS が 42.9%(3/7)で、AS は 25.0%(1/4) と少なかった。厚生労働省が公表している1985 年から 2015 年までの日本の年間出生 児数(36,122,499 名)(https://www.mhlw.go.jp/index.html)と日本産科婦人科学会が公表 しているART 由来児数(482,627 名)(http://plaza.umin.ac.jp/~jsog-art/)から、ART 由 来児の割合を算出した結果、1.34%であった。実際に観察された ART 由来患者数と予 測ART 由来患者数の比は、それぞれ BWS が 4.46、AS が 1.32、PWS が 3.44、SRS が 8.91 であった。以上の結果より、AS 以外の 3 疾患は ART と関連がある可能性が示唆 された。 2. ART 由来先天性インプリント異常症の発症原因について 次に、先天性インプリント異常症の発症原因について、ART による特徴が認められ るか検討した。二次調査より、各疾患の診断に用いた遺伝子検査と DNA メチル化検 査の結果を図4 と表 5 にまとめた。BWS では、ART-BWS で 100%(4/4)、非 ART-BWS で43.6%(17/39)に DNA メチル化異常が認められ、ART 由来患者でメチル化異常の 頻度が有意に高いことが明らかとなった(P<0.05)。SRS では、ART-SRS で 100%(5/5)、 非ART-SRS で 76.5%(13/17)に DNA メチル化異常が認められた。また、PWS では、
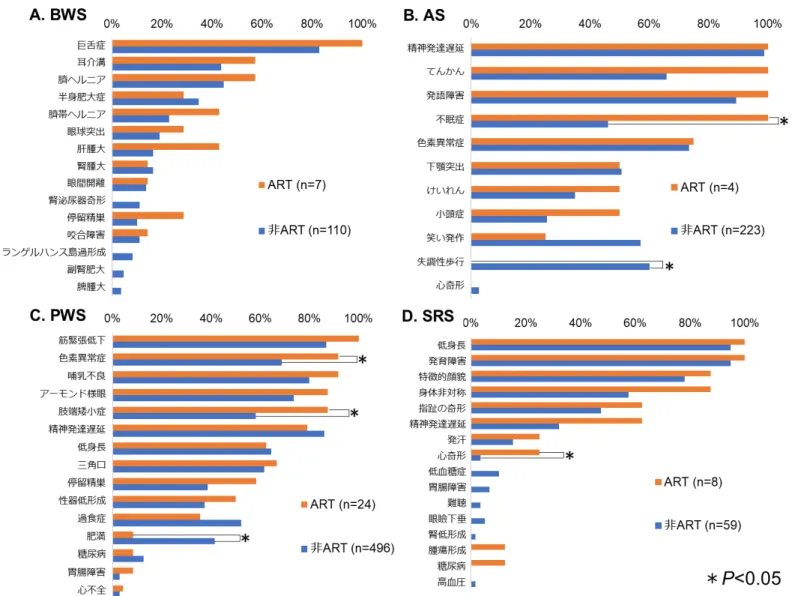
22 ART-PWS の場合、15 番染色体母親性ダイソミー(UPD(15)mat)の割合が有意に高か った(P=0.001)。ヒトの受精卵は、母体年齢が 38 歳以上になると染色体異数性の割 合が著しく増加するという報告44)がある。そこで、PWS の出生時の母体年齢を 37 歳 以下と38 歳以上の 2 群に区分し、PWS の病因の分析を行った。その結果、37 歳以下 では、ART-PWS で DNA メチル化異常の割合が、有意に高い結果(P=0.041)となっ た。しかし、38 歳以上では、ART-PWS で UPD(15)mat の割合が、有意に高かった (P=0.021)(表 6)。他の 3 疾患では、同様の母体年齢の区分を行い、病因を分析した が、年齢による違いは認められなかった(表7)。 3. ART 由来先天性インプリント異常症患者の臨床症状の特徴について 先天性インプリント異常症の臨床症状について、ART 群と非 ART 群で比較検討し た(図5)。その結果、いずれの疾患も 2 群間に大きな臨床像の違いは認められなかっ たが、いくつかの臨床症状においては、有意な違いを認めた。たとえば、AS では、 ART-AS の不眠症の割合が 100%(4/4)で、非 ART-AS の 46.2%(103/223)より有意 に高かった(P =0.048)。失調性歩行の割合は、AS は 0%(0/4)であり、非 ART-AS の 60.1%(134/223)よりも有意に低い結果となった(P =0.027)。PWS では、ART-PWS の色素異常症と小肢端症の割合が、それぞれ 91.7%(22/24)、87.5%(21/24)で あり、非ART-PWS の 69.0%(342/496)、58.3%(289/496)よりも有意に高かった(P
23 =0.020、P=0.005)。反対に、PWS の肥満の割合は 14.3%(2/14)であり、非 ART-PWS の 49.4%(205/415)よりも有意に低かった(P =0.012)。さらに、非 ART-SRS で は、心奇形の頻度は3.4%(2/59)であったが、ART-SRS では 25.0%(2/8)で、ART 群 で有意に高かった(P=0.036)。また、腫瘍形成および糖尿病の発生は、ART-SRS のみ で、ともに12.5%(1/8)に認められた。 4. ART 由来 SRS 患者における DNA メチル化の変化(DMV)の特徴
DNA メチル化異常を原因とし、発症した SRS 患者において、ART 由来患者の DNA
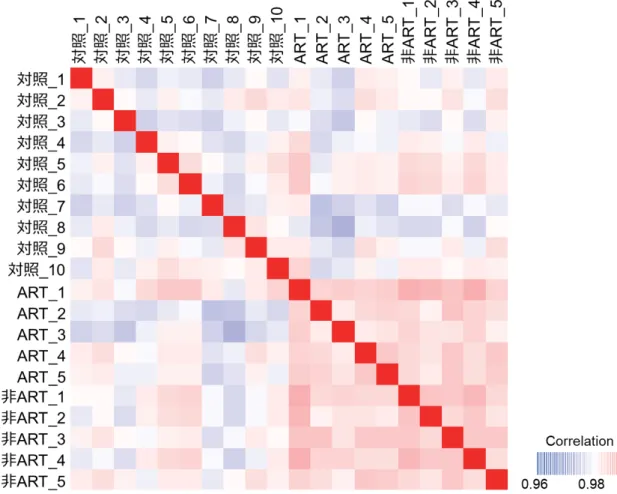
メチル化の特徴について検討してみた。ART-SRS(5 名)、非 ART-SRS(5 名)と対照 として無作為に選ばれた自然妊娠由来健常児(10 名)の末梢血 DNA を用い、RRBS 法にてメチローム解析を行った。一検体当たり平均3,242,341 CpG シトシンが検出さ れ、全ての検体で常染色体上に共通して存在する2,069,685 CpG シトシンについて解 析を行った(表8)。全遺伝子領域の平均メチル化レベルは、各群の個体間で強い類似 性を示した(R>0.975)。ART-SRS 群と非 ART-SRS 群の個体間の相関は、対照群の個 体間の相関よりもさらに強かった(R>0.980)(図 6)。次に、ゲノム領域毎にメチル化 レベルを詳細に比較した。その結果、ART-SRS 群と非 ART-SRS 群では、多くの領域 (遺伝子領域、イントロン領域、遺伝子間領域、CpG アイランドショア領域、CpG ア イランドシェルフ領域、SINE、LINE、DNA 反復配列、単純反復配列)で平均メチル
24
化レベルが、対照群と比較して有意に高かった(表9)。次に、対照群と比較して、メ
チル化レベルの差が 7.5%以上、かつ統計的な有意差(FDR<0.05)が認められた領域
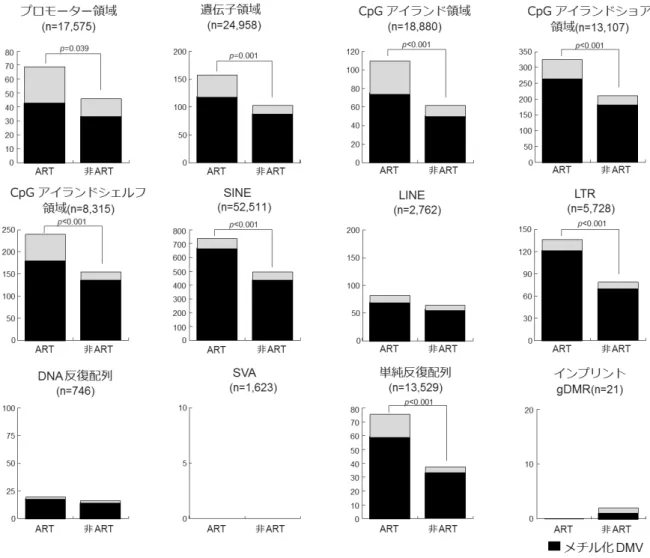
をDNA メチル化変異(DMV)と定義し、ART-SRS 群と非 ART-SRS 群にみられる、
DMV の数と領域について検討した(図 7)。その結果、プロモーター領域、遺伝子領
域、CpG アイランド領域、CpG アイランドショア領域、CpG アイランドシェルフ領
域、SINE、LTR および単純反復配列において、非 ART-SRS 群よりも ART-SRS 群で
は、DMV の数が有意に増加していた(P<0.05)。また、ART-SRS 群、非 ART-SRS 群 のDMV は、脱メチル化よりもメチル化の方が多かった。 プロモーター領域の DNA メチル化は、遺伝子の転写制御に関与することから、プ ロモーター領域について、さらに詳細な解析を進めた。SRS では、全体で 83 の領域 (79 遺伝子)に DMV が認められ、そのうち ART-SRS 群では 37 領域(36 遺伝子)、 非ART-SRS 群では 14 領域(13 遺伝子)が特異的であった(図 8A)。また、これらの DMV は、ほぼ全ての常染色体上に分布していた(図 8B)。さらに、DAVID データベ ースを使用して、DMV が認められた全ての遺伝子について、生物学的プロセスを抽 出したが、いずれも有意なGO term は認められなかった。DMV が認められた 19 遺伝 子は、SRS の一般的な臨床症状である成長遅延や、ART-SRS のみで認められた腫瘍形 成、心奇形などに関係していた。
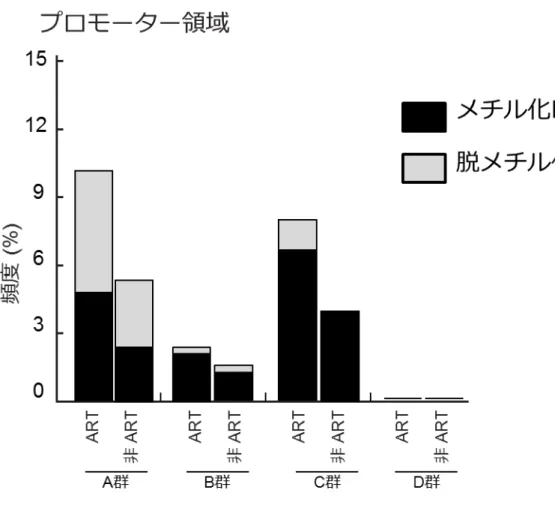
25 5. ART 由来 SRS 患者の DMV の特徴について ART-SRS の DMV の由来は、受精後の特徴を維持していることが考えられるため、 DMV の特徴について、正常な生殖細胞のメチル化パターンを 4 つに分類し、どのカ テゴリーに多く認められるか調べてみた。4 つの分類とは、精子で特異的な高メチル 化を示す領域(精子で80%以上の高メチル化、卵子で 20%以下の低メチル化:A 群)、 卵子で特異的な高メチル化を示す領域(卵子で80%以上の高メチル化、精子で 20%以 下の低メチル化:B 群)、精子および卵子の両方で高メチル化を示す領域(80%以上の 高メチル化:C 群)、精子および卵子の両方で低メチル化を示す領域(20%以下の低メ チル化:D 群)である8)。プロモーター領域では、A 群において DMV の頻度が多く、 また、ART と非 ART の差も大きかった(図 9)。
26
<考察>
1. 先天性インプリント異常症の発症原因と ART との関連性について Lazaraviciute らの総説では、自然妊娠と比較して ICSI/IVF 出生児における先天性イ ンプリント異常症の発症オッズ比は、3.67 倍であると報告されている45)。今回、我々 が実施した全国多施設共同研究では、先天性インプリント異常症の発症頻度は4 疾患 のうち、BWS、PWS、SRS は、ART と関連があると考えられ、それぞれ、4.48 倍(6.0/1.34)、 8.88 倍(11.9/1.34)、3.43 倍(4.6/1.34)、自然妊娠より高いと推測される。 一般に、BWS、SRS の発症原因である DNA メチル化異常の頻度は、それぞれ 50-60%、35-45%と報告されている46, 47)。本研究では、ART-BWS、ART-SRS の、DNA メ チル化異常を原因とする頻度は100%であった。この結果より、BWS、SRS はメチル 化異常により発症する頻度は高く、ART による DNA メチル化異常により、BWS、SRS の発症頻度が増加しているのではないかと予想される。また、PWS では、一般に、遺 伝子欠失の頻度が原因として最も多いと報告されている 46, 47)。しかし、本研究では、 ART-PWS の発症原因として、UPD(15)mat である頻度が 42.9%(9/21)であり、遺伝子欠 失の頻度の 28.6%(6/21)よりも高い結果となった。この要因として、ART だけではな く、母体年齢の上昇も考えられる。松原らは、母体年齢の上昇により卵子の第一減数 分裂における染色体不分離が増加し、その結果としてUPD(15)mat を原因とする PWS の発症が起こると報告している48)。本研究においても、母体年齢が38 歳以上の場合、27
ART-PWS の原因は UPD(15)mat の割合が有意に高く、37 歳以下では DNA メチル化異
常の割合が、有意に高い結果 (P=0.041)を示した。この結果より、ART-PWS の発症 頻度の増加は、母体年齢の上昇に加え、ART による DNA メチル化異常が関連してい るのかもしれない。 これまでに ART と DNA メチル化異常の関係については、数多くの報告がある 45, 49)。しかし、症例数が少ないため、ART と先天性インプリント異常症の関連は十分評 価できていない。さらに、本研究では、父親の年齢と DNA メチル化異常に関連は認 められなかったが、高齢男性を父親にもつ子供において遺伝性疾患、自閉症、統合失 調症および発達障害などを発症するリスクが高いことが報告されている50)。この原因 として、男性の加齢に伴う、精子ゲノムのメチル化変異が関連していることが指摘さ れている51)。このように、先天異常の原因が両親の年齢や遺伝的影響であるのか、あ るいは、ART 技術そのものに由来するのか、現時点では未だ課題となる。仮に、不妊 ではない対象が ART 技術を利用し、わずかではあるが、児の予後を医原性に変化さ せる結果を招くのであれば、さらなる ART 技術の改良が必要であり、改良によりそ の影響が小さくなる可能性があると考える。 2. メチル化変異と臨床症状との関連性
28 ART 由来児には、特異的な症状が認められるという報告がある。Lim らの報告では、 ART-BWS の場合、臍帯ヘルニアの頻度が低く、非ウィルムス性腎腫瘍の頻度が高い ことが報告されている 52)。本研究では、ART-BWS と非 ART-BWS 間において臨床症 状に有意な差は認められなかったが、 ART-SRS で心奇形、腫瘍形成および糖尿病な どの症状が、非 ART-SRS より多く、典型的な臨床症状といくつか異なるものが認め られた。 また、ART-SRS と非 ART-SRS のメチローム解析の比較では、メチル化の変異を示 す ART-SRS に特異的な DMV が 5 遺伝子領域で認められ、それらが、臨床症状の違 いに関与している可能性が示された。例えば、TCF7L2 は、低血糖に関与している可 能性がある。Wnt シグナル伝達経路調節因子である TCF7L2 は、2 型糖尿病のリスク 増加および肝臓の糖新生における負の調節因子と関連している53)。RIPOR2(FAM65B としても知られている)は、聴覚に必須である有毛細胞の原形質膜関連タンパク質を コードしており、難聴と関係している可能性がある54)。PRKAG2 は、心奇形に関係し ている可能性があり、糖原病性心筋症との関連55)が指摘されている。これら複数の遺 伝子におけるDMV は、ART-SRS で認められた糖尿病や心奇形などの特徴的な臨床症 状に関係しているかもしれない。 また、転移能力の高いレトロポゾン(自身を RNA に複写した後、逆転写反応によ り DNA コピーをつくり、ゲノムの他の部位に挿入されるタイプの分散型反復配列)
29 は、インプリントgDMR と同様に、受精後の DNA 脱メチル化に対し抵抗性である1)。 レトロポゾンの転写は、ゲノムに変異を引き起こし発生異常や疾患の原因にもなる。 DNA メチル化にはレトロポゾンの転写を抑制する役割がある56)。ART-SRS の場合、 レトロポゾンにおいてもDMV が認められたことから、ゲノムインプリンティングだ けではなく、これらの反復配列におけるメチル化変異が、ART-SRS の特徴的な臨床像 に影響しているかもしれない。今後、RRBS 法よりもゲノムのカバー率の高い、全ゲ ノムバイサルファイトシーケンシング解析によるゲノム領域の詳細なメチル化解析 が期待される。 最近、ART 由来児に新たな症状を示す先天性インプリント異常症の報告がみられる 57)。この疾患は、先天性インプリント異常症の責任領域以外のgDMR で DNA メチル 化の異常を認め、非特異的な臨床症状を呈し、診断基準を満たしていない。さらに、 ART 由来児では、診断基準を満たさない BWS や SRS 類似疾患の報告も散見されてお り、これらの症例では、これまで明らかになっていないメチル化変異が原因となって 新たな疾患を発症している可能性も考えられる35, 58, 59)。 また、ART 由来児と周産期疾患の関連について、いくつかのメタアナリシスが報告 されている。これらの論文では、ART 由来児では、早産、低出生体重児、低身長、帝 王切開、NICU 入院率、死亡率のリスクが高い傾向があると報告している 60-62)。さら に、ART 由来児では、食道閉鎖、泌尿器疾患、四肢短縮、神経管異常、心血管奇形な
30
どの先天奇形のリスクの上昇が認められたという報告63)や、血圧および乳児期の身体
発育速度が高いまたは早いという報告64)もある。胎児期、乳幼児期のエピゲノム変異
が、成人期の生活習慣病と関連すること(Developmental Origins of Health and Disease:
DOHaD 学説)が報告されている65, 66)。ART 由来児でみられる児の予後の違いは、ART
によって引き起こされたメチル化変異に起因する可能性があるため、ART とエピゲノ ム変異の関連性、それに基づくリスク要因の特定が、今後の ART 技術の向上に繋が り、出生児の健康問題に不案を抱える不妊患者の身体的・精神的負担の軽減となるこ とから、その社会的意義は大きいと考えられる。 3. ゲノムインプリンティングによるモニタリングの重要性 ゲノムインプリンティングを受ける遺伝子は、ヒトでは約100 種類あると報告され ている42, 67)。ほとんどのインプリント遺伝子は、gDMR の DNA メチル化により調節 されている。このメチル化は、受精後のゲノム全体に起こる脱メチル化の影響を受け ずに安定に維持されなければならない。配偶子や初期胚を操作する ART では、DNA メチル化に外的な影響を及ぼすかもしれない。本研究では、ART 由来患者のメチル化 変異が認められた領域に共通性は認められず、メチル化レベルは個体間で異なってい た。ART の手技がもたらす DNA メチル化変異については、凍結胚移植で出生した児 の胎盤で、MEG3-DMR の高メチル化が認められ、遺伝子発現に関係する miRNA の発
31
現に影響していたという報告がある68)。メチル化変異を受ける領域は、ART の手技に
よって異なる可能性があるため、さらなる研究が必要であると考える。
動物胚の体外培養では、子宮内で胎仔の過剰発育が起こり、出生後の死亡率や疾患
罹患率が高くなることが報告され、過大仔症候群(Large Offspring Syndrome:LOS)
と呼ばれている 27, 69)。過大仔症候群の産仔では、母親由来アレルの gDMR における メチル化が低下することで、Airn の発現上昇および Igf2r の発現低下が起こり、その 結果、IGF2 が過剰産生となり過大仔となることが示されている。ヒトでは、BWS は、 巨大児を示すことが特徴で、LOS とその病態は類似する70)。同様に、マウスの胚培養 液の組成の違いにより、インプリント遺伝子 H19、IGF2 のメチル化に異常が生じる との報告もある 71, 72)。さらに、体外培養により受精後 3 日目までのヒト初期胚では 76%、5 日目の胚盤胞では 50%と高頻度にメチル化インプリントの異常が起こること も報告されている73)。このように、ゲノムインプリンティングのメチル化は、初期胚 において変化を受けやすく、表現型として現れやすい。ART による影響を評価する上 で、ゲノムインプリンティングのメチル化の評価は適していると考えられる。近年、 ヒト初期胚の DNA メチル化解析が行われ、ヒトとマウスの初期胚のゲノムの初期化 過程が異なることが報告されている。ヒト、マウスとも、精子由来ゲノムは、能動的 脱メチル化を受ける。一方、卵子由来ゲノムでは、マウスは細胞分裂のたびに受動的 脱メチル化を受けるが、ヒトの場合は脱メチル化が不完全であることが明らかとなっ
32 ている 7-10)。さらに、ヒトの精子由来ゲノムでは、移動能力が高く疾患の原因となる レトロトランスポゾンがメチル化されていることがわかっている8)。このように、ヒ トとマウスでは異なる現象がみられることから、ヒト初期胚を用いた研究は倫理的観 点から制限があるが、ヒトのエピジェネティクスの理解にはヒトでの研究が重要であ ると考えられる。 4.メチル化変異の特徴から見た疾患発症のリスク要因 以前、Hiura らは、ART-BWS、ART-SRS 患者における、インプリント遺伝子 22 個 のgDMR のメチル化状態は、1)複数のインプリント遺伝子領域における異常が認め られること、2)父親および母親由来両方の gDMR における異常が認められること、 3)低メチル化および高メチル化両方の異常が存在すること、4)モザイクメチル化パ ターンが示されることを報告した35)。本研究では、メチローム解析により、ART-SRS 患者の場合、1)DMV の数が多く、広範囲に認められること、2)DMV は、低メチル 化よりも高メチル化されている頻度が高いこと、3)DMV は、精子特異的な高メチル 化領域で起こりやすいことを明らかとした。Okae らや他のグループの報告では、通 常、受精後にヒト精子由来ゲノムは、能動的脱メチル化を受けるが、卵子由来ゲノム のメチル化の脱メチル化は、受動的で遅れる7-10)。ART 由来児の場合、精子特異的な 高メチル化を示す領域で、DMV が多く認められたことは、受精以降の初期胚で、比
33 較的早期の脱メチル化の時期に変異が生じたものと推測される。マウス受精卵の凍結 融解操作により、DNA 脱メチル化酵素である TET3 の活性が変化すると報告されて いることから、ART によって TET3 酵素活性の変化が起こる可能性も考えられる 74)。 すなわち、ART 技術では、IVF/ICSI などの受精操作、受精卵の長期培養、凍結融解操 作および胚移植などが DNA メチル化変異のリスク要因となっているかもしれない。 これらの技術は、最近開発され、妊娠率の向上に関与していることから、今後も注意 深い観察と、対象症例数を増やした基礎的な研究が必要と思われる。
34
<結論>
本研究では、ART と関連する先天性インプリント異常症について全国調査を行い、 ART 由来患者に特有な臨床症状や DNA メチル化変異の特徴について検討した。 受精時のエピゲノム変異は、胎児に遺残し成人期の生活習慣病の発症に関連 (DOHaD 学説)することも報告されているため、今後、ART 由来児を対象とした大 規模かつ中長期にわたる健康調査が必要であると考える。ART リスク要因の特定は、 今後の ART 技術の向上に繋がり、出生児に安全な ART 医療を提供できると考える。35
<謝辞>
本研究に御協力頂きました患者様、ご家族の方々および全国多施設の医療機関関係 者の方々に、深謝致します。本研究を通じて、終始懇切な御指導と御鞭撻を賜りまし た東北大学大学院医学系研究科環境遺伝医学総合研究センター情報遺伝学分野教授 有馬隆博博士ならびに同分野前助教 樋浦仁博士(東京農業大学 生命科学部バイオ サイエンス学科現准教授)に甚謝の誠意を表します。また、本研究の遂行にあたり、 御助言および御協力を賜りました浜松医科大学小児科教授 緒方勤博士、国立成育医 療研究センター研究所分子内分泌研究部臨床内分泌研究室室長 鏡雅代博士、東北大 学大学院医学系研究科情報遺伝学分野准教授 岡江寛明博士、同分野助教 小林記緒 博士、同分野技術補佐員 北村茜修士ならびに宮内尚子さんに深謝致します。 本研究で用いた次世代シーケンサー解析につきまして、御助力を賜りました東北大 学大学院医学系研究科創生応用医学研究センター細胞増殖制御分野教授 中山啓子 博士、同分野助教 舟山亮博士、同分野技術補佐員 津田美由紀さん、菊地麻未さん、 中川真喜子さんならびに黒田清隆さんに深く感謝致します。 さらに、博士課程進学の機会を与えて下さいました、医療法人社団レディースクリ ニック京野理事長 京野廣一先生、京野アートクリニック院長 五十嵐秀樹先生なら びに職員の皆様にも、深い感謝の意を表します。36
<参考文献>
1) Messerschmidt DM, Knowles BB, Solter D. DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos. Genes Dev. 2014;28(8):812-28.
2) Saitou M, Kagiwada S, Kurimoto K. Epigenetic reprogramming in mouse pre-implantation development and primordial germ cells. Development. 2012;139(1):15-31. 3) Smith ZD, Meissner A. DNA methylation: roles in mammalian development. Nat Rev Genet. 2013;14(3):204-20.
4) Kohli RM, Zhang Y. TET enzymes, TDG and the dynamics of DNA demethylation. Nature. 2013;502(7472):472-9.
5) Nakamura T, Liu YJ, Nakashima H, et al. PGC7 binds histone H3K9me2 to protect against conversion of 5mC to 5hmC in early embryos. Nature. 2012;486(7403):415-9. 6) Cardoso MC, Leonhardt H. DNA methyltransferase is actively retained in the cytoplasm during early development. J Cell Biol. 1999;147(1):25-32.
7) Guo H, Zhu P, Yan L, et al. The DNA methylation landscape of human early embryos. Nature. 2014;511(7511):606-10.
8) Okae H, Chiba H, Hiura H, et al. Genome-wide analysis of DNA methylation dynamics during early human development. PLoS Genet. 2014;10(12):e1004868.
37
9) Smith ZD, Chan MM, Humm KC, et al. DNA methylation dynamics of the human preimplantation embryo. Nature. 2014;511(7511):611-5.
10) Zhu P, Guo H, Ren Y, et al. Single-cell DNA methylome sequencing of human preimplantation embryos. Nat Genet. 2018;50(1):12-9.
11) Li Y, Sasaki H. Genomic imprinting in mammals: its life cycle, molecular mechanisms and reprogramming. Cell Res. 2011;21(3):466-73.
12) Szabo PE, Mann JR. Biallelic expression of imprinted genes in the mouse germ line: implications for erasure, establishment, and mechanisms of genomic imprinting. Genes Dev. 1995;9(15):1857-68.
13) Hata K, Okano M, Lei H, et al. Dnmt3L cooperates with the Dnmt3 family of de novo DNA methyltransferases to establish maternal imprints in mice. Development. 2002;129(8):1983-93.
14) Kaneda M, Okano M, Hata K, et al. Essential role for de novo DNA
methyltransferase Dnmt3a in paternal and maternal imprinting. Nature. 2004;429(6994):900-3.
15) Sasaki H, Matsui Y. Epigenetic events in mammalian germ-cell development: reprogramming and beyond. Nat Rev Genet. 2008;9(2):129-40.
38
16) 日本産科婦人科学会. ART データブック 2016 2018 [Available from: http://plaza.umin.ac.jp/~jsog-art/2016data_20180930.pdf.
17) Chiba H, Hiura H, Okae H, et al. DNA methylation errors in imprinting disorders and assisted reproductive technology. Pediatr Int. 2013;55(5):542-9.
18) Cortessis VK, Azadian M, Buxbaum J, et al. Comprehensive meta-analysis reveals association between multiple imprinting disorders and conception by assisted reproductive technology. J Assist Reprod Genet. 2018;35(6):943-52.
19) Lucifero D, Chaillet JR, Trasler JM. Potential significance of genomic imprinting defects for reproduction and assisted reproductive technology. Hum Reprod Update. 2004;10(1):3-18.
20) Matsubara K, Murakami N, Fukami M, et al. Risk assessment of medically assisted reproduction and advanced maternal ages in the development of Prader-Willi syndrome due to UPD(15)mat. Clin Genet. 2016;89(5):614-9.
21) Mussa A, Molinatto C, Cerrato F, et al. Assisted Reproductive Techniques and Risk of Beckwith-Wiedemann Syndrome. Pediatrics. 2017;140(1).
22) Uk A, Collardeau-Frachon S, Scanvion Q, et al. Assisted Reproductive Technologies and imprinting disorders: Results of a study from a French congenital malformations registry. Eur J Med Genet. 2018;61(9):518-23.
39
23) Doornbos ME, Maas SM, McDonnell J, et al. Infertility, assisted reproduction technologies and imprinting disturbances: a Dutch study. Hum Reprod. 2007;22(9):2476-80. 24) Lidegaard O, Pinborg A, Andersen AN. Imprinting diseases and IVF: Danish National IVF cohort study. Hum Reprod. 2005;20(4):950-4.
25) Market-Velker BA, Zhang L, Magri LS, et al. Dual effects of superovulation: loss of maternal and paternal imprinted methylation in a dose-dependent manner. Hum Mol Genet. 2010;19(1):36-51.
26) Urrego R, Rodriguez-Osorio N, Niemann H. Epigenetic disorders and altered gene expression after use of Assisted Reproductive Technologies in domestic cattle. Epigenetics. 2014;9(6):803-15.
27) Young LE, Fernandes K, McEvoy TG, et al. Epigenetic change in IGF2R is associated with fetal overgrowth after sheep embryo culture. Nat Genet. 2001;27(2):153-4. 28) Kitamura A, Miyauchi N, Hamada H, et al. Epigenetic alterations in sperm associated with male infertility. Congenit Anom (Kyoto). 2015;55(3):133-44.
29) Lim DH, Maher ER. Human imprinting syndromes. Epigenomics. 2009;1(2):347-69. 30) Wakeling EL, Brioude F, Lokulo-Sodipe O, et al. Diagnosis and management of Silver-Russell syndrome: first international consensus statement. Nat Rev Endocrinol. 2017;13(2):105-24.
40
31) Cox GF, Burger J, Lip V, et al. Intracytoplasmic sperm injection may increase the risk of imprinting defects. Am J Hum Genet. 2002;71(1):162-4.
32) DeBaun MR, Niemitz EL, Feinberg AP. Association of in vitro fertilization with Beckwith-Wiedemann syndrome and epigenetic alterations of LIT1 and H19. Am J Hum Genet. 2003;72(1):156-60.
33) Gicquel C, Gaston V, Mandelbaum J, et al. In vitro fertilization may increase the risk of Beckwith-Wiedemann syndrome related to the abnormal imprinting of the KCN1OT gene. Am J Hum Genet. 2003;72(5):1338-41.
34) Hiura H, Okae H, Chiba H, et al. Imprinting methylation errors in ART. Reprod Med Biol. 2014;13(4):193-202.
35) Hiura H, Okae H, Miyauchi N, et al. Characterization of DNA methylation errors in patients with imprinting disorders conceived by assisted reproduction technologies. Hum Reprod. 2012;27(8):2541-8.
36) Maher ER, Brueton LA, Bowdin SC, et al. Beckwith-Wiedemann syndrome and assisted reproduction technology (ART). J Med Genet. 2003;40(1):62-4.
37) Orstavik KH, Eiklid K, van der Hagen CB, et al. Another case of imprinting defect in a girl with Angelman syndrome who was conceived by intracytoplasmic semen injection. Am J Hum Genet. 2003;72(1):218-9.
41
38) Kawamura T NM, Tamakoshi A, Hashimoto S, Ohno Y, Nakamura K. The nationwide epidemiological survey manual for investigating the number of patients and clinico-epidemiological features of intractable diseases, 2nd. edition (in Japanese). Tokyo: Japanese Ministry of Health and Welfare. 2006.
39) Brioude F, Kalish JM, Mussa A, et al. Expert consensus document: Clinical and molecular diagnosis, screening and management of Beckwith-Wiedemann syndrome: an international consensus statement. Nat Rev Endocrinol. 2018;14(4):229-49.
40) Kobayashi N, Okae H, Hiura H, et al. Genome-Scale Assessment of Age-Related DNA Methylation Changes in Mouse Spermatozoa. PLoS ONE. 2016;11(11):e0167127. 41) Krueger F, Andrews SR. Bismark: a flexible aligner and methylation caller for Bisulfite-Seq applications. Bioinformatics. 2011;27(11):1571-2.
42) Court F, Tayama C, Romanelli V, et al. Genome-wide parent-of-origin DNA methylation analysis reveals the intricacies of human imprinting and suggests a germline methylation-independent mechanism of establishment. Genome Res. 2014;24(4):554-69. 43) Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44-57. 44) Fragouli E, Alfarawati S, Spath K, et al. The origin and impact of embryonic aneuploidy. Hum Genet. 2013;132(9):1001-13.
42
45) Lazaraviciute G, Kauser M, Bhattacharya S, et al. A systematic review and meta-analysis of DNA methylation levels and imprinting disorders in children conceived by IVF/ICSI compared with children conceived spontaneously. Hum Reprod Update. 2015;21(4):555-7.
46) Nicholls RD, Knepper JL. Genome organization, function, and imprinting in Prader-Willi and Angelman syndromes. Annu Rev Genomics Hum Genet. 2001;2:153-75.
47) Ounap K. Silver-Russell Syndrome and Beckwith-Wiedemann Syndrome: Opposite Phenotypes with Heterogeneous Molecular Etiology. Mol Syndromol. 2016;7(3):110-21. 48) Matsubara K, Murakami N, Nagai T, et al. Maternal age effect on the development of Prader-Willi syndrome resulting from upd(15)mat through meiosis 1 errors. J Hum Genet. 2011;56(8):566-71.
49) van Montfoort AP, Hanssen LL, de Sutter P, et al. Assisted reproduction treatment and epigenetic inheritance. Hum Reprod Update. 2012;18(2):171-97.
50) Milekic MH, Xin Y, ODonnell A, et al. Age -related sperm DNA methylation changes are transmitted to offspring and associated with abnormal behavior and dysregulated gene expression. Mol Psychiatry. 2015;20(8):995-1001.
43
51) Jenkins TG, Aston KI, Pflueger C, et al. Age-associated sperm DNA methylation alterations: possible implications in offspring disease susceptibility. PLoS Genet.
2014;10(7):e1004458.
52) Lim D, Bowdin SC, Tee L, et al. Clinical and molecular genetic features of Beckwith-Wiedemann syndrome associated with assisted reproductive technologies. Hum Reprod. 2009;24(3):741-7.
53) Ip W, Shao W, Chiang YT, et al. The Wnt signaling pathway effector TCF7L2 is upregulated by insulin and represses hepatic gluconeogenesis. Am J Physiol Endocrinol Metab. 2012;303(9):E1166-76.
54) Diaz-Horta O, Subasioglu-Uzak A, Grati M, et al. FAM65B is a membrane-associated protein of hair cell stereocilia required for hearing. Proc Natl Acad Sci U S A. 2014;111(27):9864-8.
55) Arad M, Maron BJ, Gorham JM, et al. Glycogen storage diseases presenting as hypertrophic cardiomyopathy. N Engl J Med. 2005;352(4):362-72.
56) Hancks DC, Kazazian HH, Jr. Roles for retrotransposon insertions in human disease. Mob DNA. 2016;7:9.
57) Sanchez-Delgado M, Riccio A, Eggermann T, et al. Causes and Consequences of Multi-Locus Imprinting Disturbances in Humans. Trends Genet. 2016;32(7):444-55.
44
58) Bliek J, Verde G, Callaway J, et al. Hypomethylation at multiple maternally methylated imprinted regions including PLAGL1 and GNAS loci in Beckwith-Wiedemann syndrome. Eur J Hum Genet. 2009;17(5):611-9.
59) Shah PS, Weksberg R, Chitayat D. Overgrowth with severe developmental delay following IVF/ICSI: A newly recognized syndrome? Am J Med Genet A. 2006;140(12):1312-5.
60) Helmerhorst FM, Keirse M. Assisted reproductive technology and pregnancy outcomes. BJOG. 2016;123(8):1329.
61) Jackson RA, Gibson KA, Wu YW, et al. Perinatal outcomes in singletons following in vitro fertilization: a meta-analysis. Obstet Gynecol. 2004;103(3):551-63.
62) McDonald SD, Han Z, Mulla S, et al. Preterm birth and low birth weight among in vitro fertilization singletons: a systematic review and meta-analyses. Eur J Obstet Gynecol Reprod Biol. 2009;146(2):138-48.
63) Kallen B, Finnstrom O, Lindam A, et al. Congenital malformations in infants born after in vitro fertilization in Sweden. Birth Defects Res A Clin Mol Teratol. 2010;88(3):137-43.
64) Ceelen M, van Weissenbruch MM, Prein J, et al. Growth during infancy and early childhood in relation to blood pressure and body fat measures at age 8-18 years of IVF
45
children and spontaneously conceived controls born to subfertile parents. Hum Reprod. 2009;24(11):2788-95.
65) Barker DJ, Winter PD, Osmond C, et al. Weight in infancy and death from ischaemic heart disease. Lancet. 1989;2(8663):577-80.
66) Godfrey KM, Barker DJ. Fetal nutrition and adult disease. Am J Clin Nutr. 2000;71(5 Suppl):1344s-52s.
67) Hamada H, Okae H, Toh H, et al. Allele-Specific Methylome and Transcriptome Analysis Reveals Widespread Imprinting in the Human Placenta. Am J Hum Genet. 2016;99(5):1045-58.
68) Hiura H, Hattori H, Kobayashi N, et al. Genome-wide microRNA expression profiling in placentae from frozen-thawed blastocyst transfer. Clin Epigenetics. 2017;9:79. 69) Young LE, Sinclair KD, Wilmut I. Large offspring syndrome in cattle and sheep. Rev Reprod. 1998;3(3):155-63.
70) Weng EY, Moeschler JB, Graham JM, Jr. Longitudinal observations on 15 children with Wiedemann-Beckwith syndrome. Am J Med Genet. 1995;56(4):366-73.
71) Fauque P, Jouannet P, Lesaffre C, et al. Assisted Reproductive Technology affects developmental kinetics, H19 Imprinting Control Region methylation and H19 gene
46
72) Market-Velker BA, Fernandes AD, Mann MR. Side-by-side comparison of five commercial media systems in a mouse model: suboptimal in vitro culture interferes with imprint maintenance. Biol Reprod. 2010;83(6):938-50.
73) White CR, Denomme MM, Tekpetey FR, et al. High Frequency of Imprinted Methylation Errors in Human Preimplantation Embryos. Sci Rep. 2015;5:17311. 74) Fu L, Chang H, Wang Z, et al. The effects of TETs on DNA methylation and hydroxymethylation of mouse oocytes after vitrification and warming. Cryobiology. 2019;90:41-6.
47
<図表の説明>
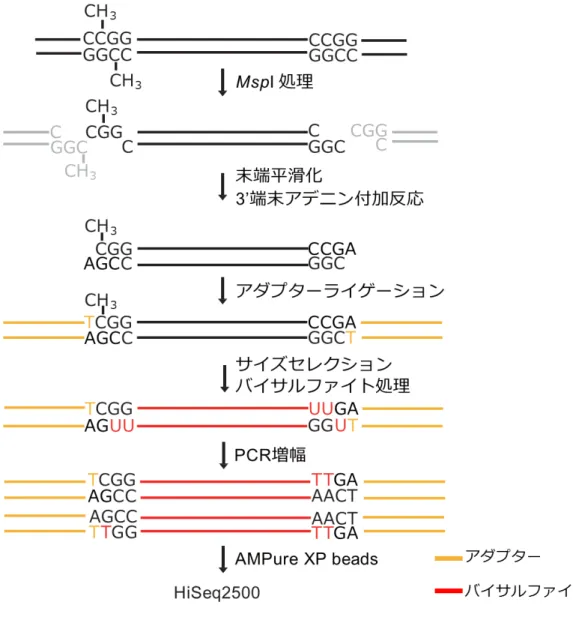
図1 マウスとヒトの初期胚発生のメチル化の変化 A.マウスの場合、精子由来ゲノムは、TET3 酵素により 5mC から 5hmC に変換された 後、塩基除去修復を経てすみやかに脱メチル化される(能動的脱メチル化)。一方、卵 子由来ゲノムでは、母性タンパク質によってTET3 からの脱メチル化から保護されて いるが、DNA 鎖をメチル化させる維持型 DNA メチル基転移酵素 DNMT1 ないため、 DNA メチル基を維持できず、細胞分裂のたびに DNA メチル化が失われる(受動的脱 メチル化)(Kohil and Zhang. Nature. 2013 より改変)。B.ヒトの場合、マウスと同様に、 精子ゲノムは能動的脱メチル化を受けるが、卵子由来ゲノムではメチル化状態に変化 が少なく、ゲノムの複製による受動的脱メチル化が不完全である。 図2 RRBS 法の概略 RRBS 法の概略を示す。まず、ゲノム DNA をメチル化非感受性制限酵素 MspI により 切断した。 その後、DNA 断片に末端平滑化修復、3’末端アデニン付加反応およびア ダプターライゲーションを行い、バイサルファイト処理を行った。反応物を PCR に より増幅し、インデックスを導入後、ライブラリーを AMPure XP beads にて精製し、 次世代シーケンサーを用いて解析した。黄はアダプター、赤はバイサルファイト処理 後の配列を示す。48
図3 先天性インプリント異常症の患者数と年齢の推移
A. Beckwith-Wiedemann 症候群(BWS)、B. Angelman 症候群(AS)、C. Prader-Willi 症
候群(PWS)、D. Silver-Russell 症候群(SRS)。縦軸は患者数、横軸は年齢を示し、青 は男性、赤は女性を示す。E.は本邦における ART 出生児数を示す(日本産科婦人科学 会ART データブックより引用)。BWS、PWS、SRS では、ART 出生児数の増加と類似 して年々増加傾向が認められ、特にBWS および SRS は、増加が顕著であった。 図4 ART 由来先天性インプリント異常症の病因の特徴 4 つの先天性インプリント異常症の病因を、メチル化異常、片親性ダイソミー(UPD)、 遺伝子微小欠失、遺伝子変異に分類した。ART-BWS、ART-SRS では DNA メチル化異 常が全例で認められ、ART 由来患者でメチル化異常の頻度が有意に高かった(P<0.05)。 また、ART-PWS では、15 番染色体母親性ダイソミー(UPD(15)mat)の割合が有意に 高かった(P=0.001)。 図5 ART 由来先天性インプリント異常症患者の臨床像の特徴
ART と非 ART 由来の先天性インプリント異常症にみられる症状を比較した。A. BWS、 B. AS、C. PWS、D. SRS。尚、PWS における過食と肥満の割合は、食事における両親
49 または患者本人の意思が影響することから、3 歳以上を対象として算出した。結果、 ART-AS では、不眠症の割合が有意に高かったが(P =0.048)、失調性歩行の割合は有 意に低い結果となった(P =0.027)。PWS の ART 由来患者では、色素異常症および小 肢端症の割合が有意に高かった(P =0.020 および P=0.005)が、肥満の割合は有意に 低かった(P =0.012)。SRS では、心奇形の頻度が、ART 由来患者で有意に高かった (P=0.036)。また、腫瘍形成および糖尿病の発生が ART 由来患者のみで認められた。 BWS では差が認められなかった。 図6 SRS 患者の全ゲノムのメチル化レベルの比較
ART-SRS 患者(ART)5 名、非 ART-SRS 患者(非 ART)5 名、自然妊娠由来健常児
(対照)10 名の末梢血 DNA を用い、RRBS 法によるメチローム解析を行い、比較し た。その結果、ART-SRS 群および非 ART-SRS 群の個体内の相関は、対照群の個体内 の相関よりも強かった(R>0.980)。 図7 ART-SRS と非 ART-SRS 間の DMV 数の比較 各ゲノム領域における DNA メチル化変異(DMV)の数を比較した。DMV は、対照 群と比較して、メチル化レベルの差が、7.5%以上、かつ統計的な有意差(FDR<0.05) が認められる領域と定義した。縦軸はDMV の数、横軸は ART-SRS 患者(ART)と非
50 ART-SRS 患者(非 ART)を示す。黒と白のバーはそれぞれメチル化された DMV と脱 メチル化されたDMV の数を示す。プロモーター領域、遺伝子領域、CpG アイランド 領域、CpG アイランドショア領域、CpG アイランドシェルフ領域、SINE、LTR、単純 反復配列では、非 ART-SRS 群よりも ART-SRS 群において、DMV の数が有意に増加 していた(P<0.05)。また、これらのすべての領域では、脱メチル化よりもメチル化さ
れたDMV の数が多かった。SINE:Short interspersed elements(短鎖散在反復配列)、 LINE:Long interspersed elements(長鎖散在反復配列)、LTR:Long terminal repeat(長 鎖末端反復配列)、SVA:SINE-VNTR(Variable number of tandem repeat)- Alu(複合型
レトロトランスポゾン)、インプリントDMR では、有意差を認めなかった。 図8 SRS 患者のプロモーター領域の DMV の数と分布 A. ART-SRS 群と非 ART-SRS 群の DMV:合計 83 個のプロモーター領域で DMV が認 められた。83 領域のうち、37 領域が ART-SRS 群で、14 領域が非 ART-SRS 群に特異 的に認められた。B. 各染色体のプロモーター領域の DMV の頻度(左)と分布(右): 赤は、ART-SRS 群、緑は非 ART-SRS 群を示す。 図9 生殖細胞のメチル化変動パターン分類による DMV 正常な生殖細胞のメチル化パターンを4 つに分類し、どのカテゴリーに DMV が多く 認められるか調べた。A 群:精子で特異的な高メチル化を示す領域(精子 80%以上の
51
高メチル化、卵子で 20%以下の低メチル化)、B 群:卵子で特異的な高メチル化を示
す領域(卵子 80%以上の高メチル化、精子で 20%以下のメチル化)、C 群:精子およ
び卵子の両方で高メチル化を示す領域(80%以上の高メチル化)、D 群:精子および卵
子の両方で低メチル化を示す領域(20%以下の低メチル化)。プロモーター領域では、
A 群において DMV の頻度が多く、また、ART-SRS(ART)と非 ART-SRS(非 ART) の差も大きかった。
表1 先天性インプリント異常症の臨床症状と原因
4 つの先天性インプリント異常症における発症頻度、原因について示す。BWS: Beckwith-Wiedemann 症候群、AS:Angelman 症候群、PWS:Prader-Willi 症候群、SRS: Silver-Russell 症候群。BWS において、UPD(11 番染色体父親性ダイソミー)、遺伝子 変異(CDKN1C の変異)、メチル化異常(KCNQ1OT1-gDMR)の低メチル化および H19-gDMR の高メチル化を示す。AS において、UPD(15 番染色体父親性ダイソミー)、遺 伝子変異は UBE3A の点突然変異、メチル化異常は SNRPN-gDMR の低メチル化を示 す。PWS において、UPD(15 番染色体母親性ダイソミー)、メチル化異常は SNRPN-gDMR の高メチル化を示す。SRS において、UPD(7 番染色体母親性ダイソミー)、 メ チ ル 化 異 常 は H19-gDMR の低メチル化を示す。(BWS、SRS:Ounap.K. Mol