ミツバ チ科 学16(1):1-8 HoneybeeScience(1995)
スズメバチ栄養液 の運動への作用
オオスズメバチは日本 に生息す るスズメバチ の中で も最 も体が大 きく, どう猛な性質を持 っ ている.私がその採集を通 じて生態行動を観察 した限 りでは他 に類を見 ない圧倒的な強さを誇 示 して森 に君臨 し,食物連鎖 の頂点 に位置す る. その行動半径は巣か ら2km 以上の範囲に および体重のy4以上の肉団子を くわえて巣 と餌 場の間を1日に何回 とな く往復 している.その 働 きぶ りには目を見張 るものがある.夕方,薄 暗 くなって も働 き続 け,真暗になると餌場 に泊 まる.詳細 に調べると最盛期のコロニーでは多 くの場合,働 き蜂全体の 1割弱が外泊 している ようである. スズメバチの毒の研究 (阿部,1985)を始め て数年が過 ぎた1984年の秋に野田市近郊の江 戸川辺 りにスズメバチ採集へ出かけた.当時, このあたりで毎年数個のオオスズメバチの巣を 採集 してきた. この年 もオオスズメバチの飛期 を確認 し,ハチの集 まる樹液を出す クヌギを見 つけ出 し, 1日を費や し, ようや くその巣を探 し当て,夜 に巣 と- チを採集 し研究所へ戻 っ た. さて, クヌギの樹液には通常,複数の巣か ら働 き蜂が飛来す ることが多 く,他 にも巣の見 つかる可能性が高いことか ら, 3日後にこの地 を再 び訪ずれ,-チの集 まっていた餌場のクヌ ギへ行 ってみた.樹液の周 りにオオスズメバチ が2-3匹止 まっているのを見つけ, もう1つ 別の巣がありそうだと内心 はっとした. ところ が,ハチを良 く見 ると何 とな く元気がない.樹 液の周 りを這 っている.変 に思 いクヌギの根元 に目をや ると 1匹のオオスズメバチが苦 しそ うに地面の上で もがいている. さらに死骸が4
-5匹落ちていた.一般 に, オオスズメバチは阿部 岳
異 ったコロニーの間で餌場を確保するために激 しい争 いをする.双方の力が均衡 している場合 には片方が死ぬまで戦 う.麹や肢を唆み切 った り,毒針で相手を刺 し殺す.死骸を1つ 1つ調 べてみたが争 った様子 はない.そ して, いくら 待 って も巣 と餌場を通 う-チが来ない.巣は先 日取 った 1つだけだ ったのだ.そこで,巣のあ った場所へ行 き,- チが いるか ど うか確かめ た.驚いたことに-チは一匹 も営巣地の周辺 に いない.外泊 していたハチは巣には戻 っていな い.餌を取 って巣に戻 って も女王蜂 も幼虫 もい ないので餌場で自らの腹を満たす しかないのだ ろう.外泊 して捕獲を免れたハチたちはどのよ うに してその一生を終えるのだろうか.森の王 者 にふさわ しく,何の不 自由 もな く一匹で生活 して行 くに違 いない.だ とすれば巣がな くなっ て しまったあと,外泊 して取 り残 されたスズメ バチは樹液の出る木の下 に何故死んでいるのだ ろうか. Ⅰスズメバチ社会の秤
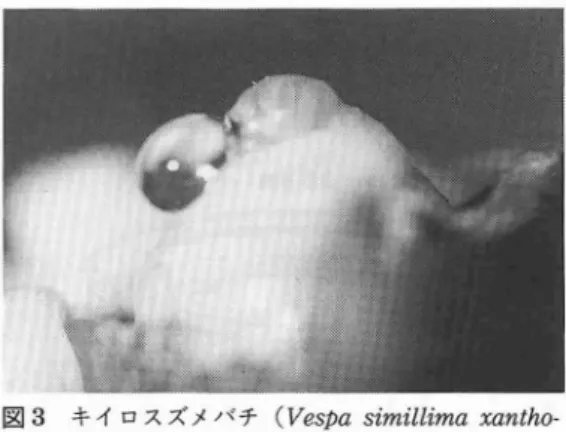
スズメバチの食性の特徴を進化の歴史を もと に振 り返 ってみれば疑問 が解 けるか も しれな い.原始的な狩蜂 は単独生活を営み,食物を捕 獲するため毒針を自在に操っれるよう細 くくび れた長 い尾を持 っている (図 1).このような基 '本的な体型,つまり, くびれた尾 は食性が変 ら ないまま社会性を獲得 したスズメバチにも受 け 継がれている (岩田,1981)(図2).口か ら入 った食べ物 は糸のように細 い尾 と胸の間を通過 し,消化器官へ運ばれる.従 って,成虫は固型 食を摂取 しに くい.つまり,スズメバチの成虫 は流動性の高 い液体食を主食 としなければな ら図 1 単独で狩生活をするシガバチ (♀)の一種 (Sceliphronfistularium) ない.流動食 は栄養のバ ランスさえ取れていれ ば消化の手間 も省け吸収 も良いので効率の良 い 食べ物の形態である.食形態の進化 と考え られ る.成虫が狩で作 る肉団子 は幼虫のための食料 で,その際,捕食 した昆虫の体液や樹液が成虫 の食料 となる. しか し,それだけで命がけの狩 と激 しい飛期行動に充分な栄養が補給できるの か.加えて, クヌギの根元であえ ぐスズメバチ を見た時,その疑問は大 きくなった. 本来,単独生活を していたハチがどのように して社会性生活を営むよ うになったのかは巣を 作 り子育てをす るようになったことが始 まりと 考え られる.そ して,そのためにいかなる仕組 を獲得 していったかが個々の-チの社会性 シス テムの特徴を示 している.草食性のハチは幼虫 に花の蜜や花粉を与えるが,それを貯蔵 して子 育てする仕組を作 り出 したのが ミツパテである (坂上,1970).この仲間は巣が最大の食物のよ りどころであるか ら,巣へ戻れば自分達の生命 は保てる.そのために必要な社会 の仕組,例え 巣内環境の制鍬 ばカス ト制やそれに伴 う機能分化が確立 された (Frisch,1967).従 って, ミツバチにとって不 可欠な食料の蓄えとカス ト制の実権を握 った女 王蜂がその社会の紳 となっている.他方,肉食 性の-チは食物を巣に貯蔵す ることが難 しいた め,子育てには特別な仕組が必要であった.そ れは栄養交換 (Trophallaxis)という成虫が幼 虫へ肉団子を与え,幼虫か ら成虫が栄養液をも らうGiveandTakeの相互依存の システムで ある.成虫のもらう栄養液がその生命維持 に不 可欠であ ると仮定すれば, この栄養交換 こそ が,肉食性の-チが社会を築 く上での紳 となっ ているに違いない (図 2).この粋が立ち切れた 時,幼虫 と成虫はそれぞれ互 に生 きて行 くこと ができな くなる. これをよりどころに して,礼 会性狩バチは基本的な社会形態を構築 し,社会 性営巣活動を維持 していると考えた. 社会性昆虫にみ られる栄養交換の現象 はすで に生 態学 的 に観 察 され て い るが (Wilson, 1971), その役割 についてはほとんど研究 され ていない. この システムが獲得 されたあと,肉 食性の社会性昆虫にとっては幼虫が食物庫の役 割を果たす ことになる. その中で もスズメバチ はこのよ うな社会性 システムを最 も発達 させ た. この幼虫の出す栄養液 は体内に発達 した分 泌腺で作 られ,頭部を刺激す ると口器より分泌 される (図3).初回の刺激に対す る分泌量が一 番多 く,連続的には分泌 されない.オオスズメ バチの幼虫で100〝lほど分泌す るが,キイロス 巣 内環境 の調整 図2 スズメバチの栄養交換と社会的秤
図3 キイロスズメバチ(Vespasimillimaxantho -♪tera)の分泌する栄養液 ズメバチ, コガタスズメバ チ, チ ャイ ロスズメ バチ, ヒメスズメバ チそ してモ ンスズメバチで はその半量 よ りも少 ない. 肉団子 を与 えないと 数時間後 には刺激 に対す る分泌を行 なわな くな る. スズメバ チ栄養液が成虫 の生命維持 に不可欠 な ものであ ることは成虫 の飼育実験 で砂糖水 を 与 え た ものが2- 3週 間 しか生 きな いの に比 べ, 栄養液 を加 えた砂糖水 を与 え ると2-3か 月生 きることか らも分か るが,寿命 とは別 に自 然 の中では成虫の行動 を制御 していると思 われ る (図2).例 えば,水分 の供給,エネルギーの 補給,飛期時の疲労予 防,狩 に伴 うス トレスの 緩和,体調 の維持,巣 内での争 い防止 などスズ メバチの生活 を支え るあ らゆることに関与 し, 3 コ ロニーの盛 衰 を も左右 して い るに違 いな い (Abeeta1.,1991). そ して,幼虫が栄養液 の 組成 を変 え ることで成虫 の様 々な行動 を コ ン ト ロールす る化学情報伝達 の役割 を も担 っている だろ う. Ⅲ ス ズ メ バ チ栄 養 液 の組 成 日本 に生息 す る5種類 の ス ズメバ チ幼虫 が 分泌す る栄養液を分析 した ところ, ア ミノ酸 を 主成分 とす る糖 や タンパ ク質 を含 む水溶液であ った(Abeata1.,1991)(表 1).特 に,ア ミノ 酸 の組成 は肉や クマ ゴそ して ミル クの タ ンパ ク 質 とは全 く異 な り, どの種 も共通 して グ リシ ン, プ ロ リンそ して ス レオ ニ ンな どを多 く含 み,同時 に全ての必須 ア ミノ酸 も含 む特異 なパ ター ンを示 した(Abeeta1.,1991)(図4).ま た グル コースや昆虫の グ リコーゲ ンに相 当す る トレ- ロース も含んでいた. このよ うな組成 の 栄養液が成虫 のどのよ うな行動 を制御 している のか, スズメバ チを用 いて明 らかにす ることは 人工飼育 を含 めて行動生態学 の基礎研究 が欠 け ている現状 では難 しい. しか し,多岐 に渡 る作 用 が その組成 の複雑 さか ら予 想 され る. 例 え ば,攻撃的 な性格 の強 いスズメバチが巣 の中に 沢 山集 まる状 況下 で, GABAの よ うな抑制性 神経 の トランス ミッターや グ リシンのよ うなア 蓑 1 スズメバチ栄養液中のアミノ酸と糖の含量 総炭水化物 D-グルコース(A) トレハロース(B) (mg/ml) (mg/ml) (mg/ml) A+B(%)b) チ チ チ チ チ パ パ バ パ パ ・5 憲 仙 オ ン メ ガ イ オ モ ヒ コ キ 3 2 5 8 5 9 1 5 9 8 1 4 3 6 9 0 0 5 3 8 0 0 7 6 0 L 4 3 5 3 18 18 が 35 02 1 . L N 2 . 9 . 3 6 6 5 3 1 2 0 1 2 1 1 1 1 1 総ア ミノ酸 生理的ア ミノ酸 プロリン u-アミノ酸') 働き蜂の体重 (mg/ml)C) (mg/ml) (%)d) (mg/ml) (%)e) bmol/ml) (mg) 4 8 4 0 9 4 5 4 0 2 5 2 6 7 3 2 1 7 2 5 9 9 7 3 8 6 3 3 9 0 8 8 8 3 9 8 6 5 7 9 0 7 9 1 6 6 2 1 3 8 7 8 0 6 5 4 7 L t 2 2 6 8 3 9 8 5 6 3 6 5 1 0 0 1 0 15.17 1365±285(∩-403) 12.98 931±365(∩-301) 10.18 1325±187(∩-84) 5.96 600±80 (∩-109) 3.08 355±53 (∩-296) a)検出不能 b)総炭水化物中に占める割合 C)栄養液の酸水解によって算出 d)全アミノ酸中に占める生理 的アミノ酸の割合 e)生理的アミノ中に占める割合 f) タウリン,グリシン,β-アラニン,GABAの合計
コ ガ タ ス ズ メバ チ 0
5
-
≦
L
T
l
G
h
i
由一・★-・=*・ ア ミノ酸 図 4 日本産5種のスズメバチ栄養液中に含まれるア ミノ酸とその含量 (Mol%) *印は必須アミノ酸 ゴニス トの働 きをす るW-ア ミノ酸 を多量 に摂 取 した場合,-チの攻撃性 は沈静す るだ ろ う (表 1). プロ リンは昆虫一般にエネルギー源 と して使 われて いるので,飛糊時 には欠かせな い. ロイシン, イソロイ シンそ してバ リンなど は筋 収縮 の エ ネルギ ー と して使 われて い る (Abeeta1.,1991).この特徴ある複雑なア ミ ノ酸組成 は行動範囲の広 い狩行動を も支えてい るだろう.具体的には疲労時の脳の活動低下を 予防 し,狩をする時の身の安全を守 り,虫を素 早 く捕 まえ る行動 に役立 って いるので はない か. また, 肉団子を2km以上離れた所か ら途 中で休む ことな く時速30kmあまりの速 さで 巣 まで持 ち帰 る. それを1日中繰 り返 して い る. もし,何 らかの理由で幼虫-の肉団子の供 給が悪 くなれば幼虫の生長 は止 まり, コロニー の増大が抑制 される.成虫が良 く働 くことが巣 の繁栄に繋がる. このようなことか ら,栄養液 が飛期,つまり筋肉運動 に伴 う疲労の改善に役 立っのではないか と考えた. 卜 = 38cm- -「 図5 マウス遊泳実験装置 水温35℃,流速5-8m/分のリバプールⅢ
スズメバ チ栄養 液 の作 用 運動に伴 う疲労を測定す るため,マウスを用 いる遊泳装置を新 たに作 り (Eg15),遊泳能力 と 運動状態に対するスズメバチ栄養液の作用を検 討 した.オオスズメバチと同 じ組成の栄養液を選 び,VespaAminoAcidMixture(VAAM) と名付け, この他にVAAMとは異なる組成の 様々なア ミノ酸栄養液, カゼイ ンア ミノ酸栄養 液 (CAAM),グルコース溶液 (Glu)そ して, 蒸留水 (D.W.)をマウスにlmlずつ飲 ませ,投 与後30分間静置 してか ら, 35℃の流水プール (流速5-8m/分)で遊泳を開始 し,溺れるまで の遊泳時間を測定 した.1.8%VAAM投与群が 最 も長 く,0.9%CAAM,10%Glu,D.W.の順 に 泳 ぐ時間が短か くな った (図 6).遊泳時間の延 長効果 は乳酸などの疲労物質の体内蓄積が少な いことを示唆 している.尾 に0.3gの重 りを負 荷 したマウスを用いて30分間遊泳後の血中乳 酸値 と血糖値を測定 した.1.8%VAAM投与群 が1.8%CAAM,10%Glu,D.W.さらには様々 な組成のア ミノ酸栄養液に比べ,極めて低い乳 酸値を示 した (図7)(Abeeta1.,1995).筋肉 中の乳酸量 もVAAM投与群で最 も低い. 同時
D.W. 1.8%CAAM 0.9%VAAM 1.8%vAAH GtLICOSe (10.0%)
0
1 0 0 200 300 iこ Ji ;R hkl.lIm(め) 図6 VAAMおよび種々の栄養液を摂取 したマウス の最長遊泳時間 に乳酸の前馬区体であるピル ビン酸 の減少 もみ ら れた (Abeeta1.,1989). これは VAAM の摂 取 によって運動時の疲労物質が体内に蓄積 しに くいことを示 している.血糖値の減少 はこれ と は逆 にVAAM 投与群で負荷運動 に もかかわ ら ず,極 めて少 なか った (Abeeta1.,1995)(図 7).この相反す る作用の同時進行 は通常の持続 運動 に伴 って起 こる乳酸 の蓄積 と血糖値 の低下 を同時に抑制 してお り,疲労の回避 に好 ま しい 状況である. オオスズメバチ栄養液その ものを 投与 した場合, さ らにこの抑制作用 は強か った (図 7). 我 々のよ うな高等動物 は糖, タンパ ク質,脂 質 などをェネルギー源 と し,炭酸 ガスと水 に変 換す ることで生命活動 を営んでいる.糖 (グ リ コーゲ ンや グルコース) は運動 に際 して最 も使 いやす いエネルギーで,糖 の不足 に伴 いア ミノ 酸や脂質 が順次使われることは良 く知 られてい る.糖 の燃 え る時に酸素が不足す ると乳酸がで きやす くなる.従 って,運動時には血糖 の減少 と乳酸 の増加が起 こる. VAAM による乳酸減 少 と血糖維持作用 は糖の利用を調節 し,同時に エネルギー源 として脂肪の利用促進 を示唆 して いる.種々の栄養液 による同様 の遊泳実験 によ って,1.8%VAAM 投 与 群 は 1.8%CAAM や 10%Gluと比 べて負荷遊 泳後 に血 中脂肪酸 の 遊 離 が増 加 した (田村 ら, 1990;飯 田 ら, 1991)(図 8).遊離 された脂肪酸 の組成 は安静 時の飽和一不飽和脂肪酸 の割合 とほぼ同 じ比率 で増加 していた (白井 ら, 1991).脂肪酸が酸 化 してで きるケ トン体 の量 も1.8%CAAM や 10%Glu投与群 に比 べて 1.8%VAAM で は増 加がみ られた (飯田 ら,1991).また,ウサギを 用 いた0.9%VAAM の静脈 内投与 で も血 中の 遊離脂肪酸 の顕著な増加が認 め られた (森下 ・ ⑳ :オカスズメバチ (V ma,'dar川Za)紫恭液★:18%V^^M @ :L8%CA^MO:DW □ :10%G)u △:10%GLuIL8%VAAM 1-20日di/Iの ア ミ/赦柴鐘液 図7 種々のアミノ酸栄養液を摂取 し,30分間負荷 遊泳したマウスの血中乳酸値と血糖値 (t\ b3 ∈ ) 撃 溢 当 聖 蓋 鞘 rh TT )10
38 60 遊泳時糊 ())) 図8 種々の栄養液を摂取 した負荷遊泳マウスの血中 遊離脂肪酸の経時変化VAAM とCAAM は 1.8%,Gluは 10%.
阿部,1993). このよ うな脂肪 の分解促進 はノルア ドレナ リ ンやア ドレナ リンなどのホルモ ンの増加が もた らしていた ことが同様の遊泳実験で明 らかにな った (表2)(稲森 ら,1992;白井 ら,1993). つまり, ホルモ ン感受性 リパ ーゼが活性化 され (土 田 ら,1991),続 いて脂肪 の分離が促進 し, 脂肪酸が増加す る. これ らの結果 はエネルギー
6 蓑 2 種々のアミノ酸栄養液を摂取 し, 30分間負荷 遊泳 したマウスの血中ノルア ドレナ リンとア ドレナリン濃度 ア ドレナ リン ノルアドレナリン 匹数 品 匹敷 詰 1.8%VAAM 15 5.06±0.93 12 6.19±1.ll 1.8%CAAM 12 3.10±0.49 11 4.52±0.56 D.W. 12 4.80±0.46 11 6.17±0.41 源 に脂 肪 を利 用 す る代 謝 経 路 の引 き役 体 を VAAM が果 していることを示唆 している. 運動 という生体 にとってのス トレスへの順応 をVAAM が促 しているよ うにも見え る. しか し, ス トレス状態を加速 しているかにみえる一 方で,運動 ス トレスに伴 う血液中のア ミノ酸, 特 に筋肉で特異的にエネルギー源 と して用 い ら れる分枝 ア ミノ酸 を中心 に多 くのア ミノ酸 の減 少 をVAAM は抑制す る.VAAM を投与 した ラ ッ トは走行運動 に伴 う血 中 ア ミノ酸変動 が CAAM やD.W.投与群 に比 べて顕著 に少 ない (図 9)(土 田 ・阿部,1993). ヒ トで も激 しい, あるいは長時間の運動 のあとは分枝 ア ミノ酸 や セ リン, ス レオニ ン, グ リシン, アラニ ン, チ ロシンなどが著 しく減少す るが (Bazzarre et a1., 1992; Blomstrand and Newsholme,
1992), こ れ ら変 動 の 大 き い ア ミノ酸 は VAAM に多 く含 まれて いる. 他方, アスパ ラ ギ ン酸, ス レオニ ン, メチオニ ンそ して グルタ ミン酸 は運動 に伴 う変化 は少 な く, VAAM 中 の含量 も少 ない.運動 によるス トレスか ら発生 す るア ミノ酸 のイ ンバ ラ ンスの改善 をVAAM が促 し, 運動時に減少す るア ミノ酸 をVAAM が補 っている.
Ⅳ
推定 されるVAAM
の作用機序 VAAM に関わ る様 々な実験結果か ら, 中間 代謝産物 の変動 や ホル モ ンの増加 を もとに し て,VAAM の最 も単純 な作用機序を考えてみ た (図10).VAAM は吸収 され る腸,あるいは 代謝 される肝臓 や脳でその組成 にもとずいたア ミノ酸の種頬 と量 のパ ター ンが認識 され, この よ うな シグナ リングを もとに した高次 ホルモ ン の合成 と分泌の促進を促す と推定 した. その結 果, カテコールア ミンなどの低次のホルモ ンを 誘導 し, さらにホルモ ン感受性 リパ ーゼを活性 化 す る.活性化 した酵素 は脂肪 の分解 を促進 し, グ リセロールと脂肪酸 を増加す る.活性脂 肪酸 (アシル CoA)は解糖系の律速酵素である ホスホフルク トキナーゼや脂肪酸合成酵素 を阻 害 し, 脂肪酸 の分解を促す. アシルCoA はβ 酸化 によってケ トン体 などを経て多量のアセチ ル CoA となる.増加 したアセチル CoA は ピル ビン酸 デ ヒ ドロゲナーゼを阻害 し, ピル ビン酸 か らアセチルCoA への合成反応 を抑制す る. また逆 に ピル ビン酸 カルボキ シラーゼを活性化 し, ピル ビン酸か らオギザロ酢酸への反応を促 進す る結果, グル コース合成反応 (糖新生)が 進み,同時 に脂肪の分解 によって増加 したグ リ セ ロール もこの反応系 に加わ り, グルコース合 成 が促進 され る. アセチルCoA は TCA サイ クルで代謝 され, クエ ン酸を経て最終的に多量 のATPを産生 す る. クエ ン酸 も ATP もホス ホフルク トキナーゼの活性を阻害 し, グルコー スの分解を抑制す る.他方, カテコールア ミン はcAMP依存性 プ ロテ ィンキナーゼ活性化機 構 に も働 き, グ リコーゲ ンの分解促進 と合成阻 害を行 い, グルコースの増加 を促す,VAAM はこのよ うな脂肪の分解 に始 まる一連 の代謝反 pro Gly AIR His _ L., VBl Lou ぎ 丁 … 宣 ,hr d cE) Met Tyr Cys Phe 図9 種々のアミノ酸栄養液を摂取 したラットの30 分走行後の血中アミノ酸組成 3.6%栄養液を4ml投与 した.応 を活 性 化 し, 多 量 の エ ネ ル ギ ー (ATP) を生産す ると同時 にグルコー スの減少 を抑制す るスムーズな代謝調 節 を行 っている. この他 に も血 中ア ミ ノ酸の運動 に伴 うイ ンバ ランスを補正 す る作用 は運動 によって起 こる筋細胞 の破壊防止 を示唆 している.
Ⅴ
ヒ トに対するVAAM の作用 この様 な作用を示すVAAM がどの ような味を しているのか興味を持っ人 も多 いと思 う. ア ミノ酸 の味 は一般 に うまみ と表現 され るが, どのア ミノ酸 も溶液に して味わ うと決 して良 い味で はない. どのような割合で混ぜれば味 が良 くなるのか,人の手で採 し当て る のは手間のかか る難 しい仕事である. 自然 が作 ったカクテルであるVAAM は美味 しいとは言えないが,飲めない はどひどい味ではない.実験 に用 いた CAAM や他 の組成 のア ミノ酸栄養液 †一一一一一 尉 肝絶 版 ● -ルアミンなど・一一-一〇 D-グリセルアルデヒドー ジヒドロキシ/ : :アセトンリン酸 〇一一一一日活性化 ▼ ●一・一・・一一抑制 一一一一代謝の流れ 図10 と比べれば格段 に飲みやす い. イヌは VAAM を好んで飲む. ヒ トで も動物 実験 と同様の作用が期待で きそ うだが,直ちに 実験 はで きないので,まず1.8%VAAM を 200 -300ml試飲 して もらうことに した.美味 しい と答えた日本人 はほとん どいなか ったが,運動 中に飲む と甘 くて美味 しLとい うヒ トはいた. ある中国人 は華南 の高級料理 に出て くるスープ の味に似て美味 しいと言 った. さらに飲みやす 7 吸収 : VAAM (スズメバチ栄妊液) 脂肪 (トリグリセリド) 11 COZ 中間代謝産物およびホルモンの変動から示唆された運動 時におけるスズメバチ栄養液(VAAM)の作用機序 0 2 4 6 8 10 ぶtrn・ Al (km) 図11 体重 55kgのランナーが走前 30分前に 1.8% VAAM を 150ml摂取 し,2-10km走った後 の体重減少 nはくり返 し回数 くす るため,多少の甘味を付 け,多数 の ヒ トに 試 して もらった. は じめて飲んだ ヒ トの半数以 上が直後 に身体のほて りや温感を訴えた.精力 剤を飲んだ時のような身体 の躍動感や興奮感 は ない. 一連 の動物実験 が示 したVAAM の効果 は運 動時に顕著 に現われる. VAAM の成分 および 作用の点か ら, ドー ピングには抵触 しない.そ こで, ラグビー, アイススケー ト, アイスホ ッ ケー,マ ラソン, アメ リカ ンフッ トボールなど の トップ アス リー トに試 合 お よ び練 習 時 に 1.8%VAAM を 300mlはど飲んでいただいた. ●ほとん どの選手 が何 らか の効果 を認 めて くれ た.「飲 まないと試合 に出ない」
,
「疲れを感 じに くい」,
「また飲みたい」,
「最後 まで シャキ ッと している」,
「運動中は飲みやすい」,
「飲んで練 習す ると試合時 に良 い」,
「疲れがたま らない」 など様々な感想が寄せ られた.一万,10km 走 および20km - - フマ ラソ ンでベテ ランラ ン8 ナ 一に よ る摂 取 テス トの結 果 ,体重 の減少 が多 数 の被験者 で小 さか った (図 11).これ は運 動 時 の発 汗 量 が少 な い こ とを示 唆 して い るの か 「給 水 を取 らな くて もす む
」
,
「空腹 感 が な い」と い う感 想 が多 か った. 同 時 に記 録 の上 で も改善 がみ られ た (本 間 ら, 1993). 現在, VAAM に関 して運動生理学 的お よび 生化学 的 な観点 か ら,動 物 実験 よ り も広範 かっ 多 角 的 に研 究 を展 開 して い る.詳 細 な結 果 が得 られ れ ばVAAM の ヒ トに対 す る作用 が さ らに 明確 にな る と期 待 して い る. (〒35ト01 埼玉県和光市広沢 2-1 理化学研究所 昆虫生体制御研究室) 引 用 文 献 阿部 岳.1985. ミツバチ科学 6(1):13-24. Abe,T. etal.1989. The 14th InternationalCongressofNutrition. Seoul,Korea. Abe,T,etal.1991.Comp. Biochem. Physiol.
99C:79-84.
Abe,T.etal.1995. Jap.J.PhysicalFitness& SportsMedicine44(2):inpress.
BazzarreT.L.etal.1992.I.Am.CollegeNutri -tion ll.501-511.
Blomstrand E. and E.A.Newsholme. 1992. ActaPhysiol. Scaれd. 146:293-298.
本間保夫 ら.1993.体力科学 42:523. 岩田久二雄. 1971.本能の進化 サイエンティス ト 社.東京.pp.503. 飯田耕司 ら,1991.生化学63:765. 稲森美穂子 ら.1992.生化学 64:965. 森下幸治,阿部 岳 1993.体力科学 42:231, 坂上曙一.1970. ミツバチのたどったみち.思索社 東京pp,327. 白井暢子 ら.1991.体力科学41:655. 白井暢子 ら.1993.生化学 65・691. 田村正啓 ら.1990.生化学 62:606. 土田 博 ら.1991.体力科学41:656. 土田 博,阿部 岳 1993.体力科学 42:708. Frisch,K.Yon.1967.TheDanceLonguageand
Orientation ofBees. Harvard Univ.Press, Combridge.pp.566.
Wilson.E.0.1971.TheInsectSociety.Harvard Univ.Press.Cambridge. pp.548.
ABE,TAKASHI. EffectofVes♪aAmi no Acid
Mixturefrom hornetlarvalsalivaonendurance exrcise.HoneybeeScience(1995)16(I)1-8.Lad. InsectToxicology and Physiology,Instituteof Physical and Chemical Research. 2-1,
Hirosawa,Wako-Sh呈,Saitama,351-01Japan, Itiswellknownthatthesocietyofhoneybee isstronglytiedbythequeenasacontrollerof castesystem and by thestored food such as honeyand pollen. Comparedtothehoneybee which isa vegetarian insect,thehornetofa canivorousinsectisnotabletostorethefood intheirnest,andalsohavenotmoresolidcaste system thanhoneybeesociety Oneofthemost importantsocialtiesisthereforeatrophallaxis,
thatisafoodexchangebetweenadultmaking meatballandlarvalsaliva.inthehornetsocie -ty. Thisnutritionalexchangewasatransfor一 nationofchemicalinformationsfrom larvaeto adu一tsforcontrolofthesocialactivity. The larvalsaliva of5 speciesinhabited in Japan was analyzed. They were contained amino acidsandsugars.Theaminoacidcompositions werecommonly shown a peculiarcomponent thatwasrichinproline,glycineandthreonine. The mixture ofamino acids (VAAM)in the salivaofVespamandariniashowedanactivity oftime elongation in swimming exercisesof mice. VAAM alsoshowedsupperssionsofthe increaseofblood lactateand thedecreaseof blood glucose during loaded swimming exeト cisesofmice. ThoseeffectsofVAAM were produced by theactivation oflipolysis. The lipolysiswasinduced by catecholamines. The hormanalinduction would occure through a recognitlOnOftheamlnOacidpatternofVAAM in the intestine orthe brain, When human took theVAAM beforeexercise,theperfor m-ancewasimproved.

![[書評] 菱田雅晴・園田茂人著『経済発展と社会変 動 シリーズ現代中国経済 8』](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)