Title Studies on Characterization of Variable Cell Responses to PrionAgent( 本文(Fulltext) )
Author(s) ELHELALY, Abdelazim Elsayed Abdelazim
Report No.(Doctoral Degree) 博士(獣医学) 甲第397号 Issue Date 2013-09-24 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/47361 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
1
Studies on Characterization of Variable Cell
Responses to Prion Agent
(プリオン因子に対する種々の細胞の反応特性に関する研究)
2013
The United Graduate School of Veterinary Sciences, Gifu University
(Gifu University)
2
TABLE OF CONTENTS
CONTENTS
2
GENERAL INTRODUCTION
6
CHAPTER 1: Responses to PrP
ScDuring Prolonged Cell
Culture and Its Effect on Transmission of PrP
Scto Neural
Cells
9
INTRODUCTION
10
MATERIALS AND METHODS
12
Cell lines
12
Prion strains
12
Infection of cells with prion
12
Co-cultures
13
PrPC expression in cells
13
PK treatment
14
Western blot analysis
14
Imaging of prion in cells by immunofluorescent microscopy
15
3
RESULTS
17
Characterization of initial cell responses to prion exposure
17
Endocytosis of PrPSc by cell lines
17
PrPC expression in different cell lines
18
Proteasome activity in the cell lines as an indicator of degradation ability
18
Characterization of the late responses to PrPSc exposure during prolonged incubation
19
Late cellular accumulation of PrPSc in relation to scrapie strain
20
Transfer of PrPSc from infected cells to neural cells after the long incubation
21
Establishing persistent PrPSc infection in the cell lines after short and prolonged infection
21
DISCUSSION
23
SUMMARY
28
FIGURES AND TABLES
29
CHAPTER 2: Early Transient Accumulation of PrP
Scin
Immune Cells; Characterization and Affecting Factors
36
4
MATERIALS AND METHODS
39
Cell lines
39
E. coli cell lysate and microspheres
39
Prion strains
39
Infection of cells with prion
40
Cholesterol depletion
40
Proteinase K treatment
40
Western blot analysis
41
Proteasome assay
41
RNA extraction and Quantitative real-time PCR
42
RESULTS
42
The early transient accumulation of PrPSc in macrophages and its origin
43
Characterization of PrPSc degradation in macrophages at variable incubation temperatures
44
Effect of additional PrPC on the accumulation of PrPSc in Raw cells
44
Characterization of the specificity of early prion accumulation
45
5 macrophages
DISCUSSION
47
SUMMARY
50
FIGURES AND TABLES
51
GENERAL DISCUSSION AND CONCLUSIONS
55
ACKNOWLEDGMENTS
57
6
GENERAL INTRODUCTION
Prion, a fatal neurodegenerative disease of human and animals, is the archetype of disorders known as transmissible spongiform encephalopathies (TSEs). It typically takes long incubation periods prior to the onset of progressive clinical symptoms. Diseases in this group include Creutzfeldt-Jakob disease (CJD), Gerstmnnn-Straussler-Scheinker syndrome (GSS), Kuru and fatal familial insomnia (FFI) in humans, scrapie in sheep and goat, chronic wasting disease (CWD) in deer and elk, and bovine spongiform encephalopathy (BSE) in cattle. PrPSc, an abnormal, protease-resistant isoform of the host encoded prion protein (PrP), plays a central role in the pathogenesis of these diseases. PrPSc is derived from the cellular prion protein (PrPC) by a post-translational process involving profound changes in the secondary and tertiary structure of PrP. These conformational changes appear to be critical to the propagation of the causative agent and the pathogenesis of these diseases [59-61].
Public health concerns that BSE may be transmissible to humans via dietary exposure of BSE-contaminated foods have been raised after the appearance of variant CJD (vCJD) [17]. The worldwide epidemic of BSE in cattle is thought to have been caused by the oral administration of PrPSc-contaminated meat and bone meal. However, the vast majority of cattle fed PrPSc-contaminated meals have not been diagnosed with BSE and failed to accumulate PrPSc [3]. The reasons underlying why the cattle fed the same food did not develop BSE remain unclear.
Ingested PrPSc particles may enter the host through the gut before invasion of the gut-associated lymphoid tissues, where the first PrPSc amplification takes place [1, 4, 32, 74]. Following ingestion, it is widely believed that the PrPSc first cross the intestinal
7
epithelium through the microfold cells (M cells) which are specialized for trans-epithelial transport of antigens [54]. Several experimental studies have suggested that PrPSc first accumulates within the follicular dendritic cells (FDC) located in B cell follicles within the gut-associated lymphoid tissue (GALT) [28, 48, 58]. Phagocytic cells including macrophages and dendritic cells (DC) reside within the M cell pockets [54], where they are thought to function as initial host responders against PrPSc. It is currently believed that the translocation of PrPSc agents from the lymphoid tissue to the central nervous system (CNS) occur via the peripheral nervous system [8]. PrPSc infection of the enteric nervous system has been reported to occur within fine nerve fibers directly underneath the villous or crypts of epithelium [37]. PrPSc migrates asymptomatically along the peripheral nervous system by axonal transport mechanisms via the vagus and splanchnic nerve [74]. Upon finally reaching the CNS, PrPSc aggregates and accumulates as the neurodegeneration process advances [2, 49]. The sequential detection of PrPSc in lymphoreticular tissues and subsequently along neural projections to the CNS has led to the conclusion that these tissues are involved in the spread of PrPSc to the CNS [41].
Transgenic (Tg) mice expressing PrP genes of hamster, human, and cattle have been used to study prion diseases, and especially to investigate the species barrier and susceptibility to different prion strains [18, 67, 72]. However, experiments using Tg mice were proved to be hard and time consuming. The most common and fastest ex vivo method is to incubate PrPSc with cell lines and check for its accumulation. From these studies, some cell culture models supporting PrPSc replication have been established [7, 55, 69, 76]. These present various advantages including: (i) the ability to analyze the biological properties of PrPC and PrPSc; (ii) the possibility of determining the nature and
8
factors controlling the propagation of PrPSc; (iii) evaluation of the effect of therapeutic agents at the cell level [69]. These models are important for the understanding of the cell biology of PrPSc isoforms.
As immune responses typical of acute infection do not occur in prion diseases, the potential role of the immune system in the development of prion diseases has become under the spotlight and is being focused by researchers. It has become increasingly apparent that the immune system potentially participates in the development of the disease. The role of some cells like macrophages, mast cells or peripheral neural cells in accumulating, establishing or transferring prions remains unclear and the responses of these cells to PrPSc over time require investigation. For instance, although the initial function of macrophages is to combat against exogenous antigens, phagocytic cells may catch PrPSc molecules and keep for a while, they may potentially transfer it to other pathway before complete degradation of it [15, 34].
The overall objective of this study is to investigate the potential involvement of the examined cell types in accumulation and transfer of PrPSc as well as the characters that may affect the cell-PrPSc interaction. In chapter I of this thesis, the early and prolonged cellular responses to PrPSc for up to 28 days were investigated using human and murine cell lines of immune, neural, and intestinal cell types. Chapter 2 describes further detailed investigation to characterize a transient and early PrPSc accumulation in macrophages as well as the relationship between PrPSc formation in these cells and some related factors.
9
CHAPTER 1
Responses to PrP
ScDuring Prolonged Cell Culture and Its Effect on
Transmission of PrP
Scto the Neural Cells
10
INTRODUCTION
Particles of ingested scrapie protein (PrPSc) may enter the host organism through the gut before invasion of the gut-associated lymphoid tissues (GALT), where the first PrPSc amplification takes place [1, 4, 32, 74]. PrPSc infection of the enteric nervous system has been reported to occur within fine nerve fibers directly underneath the villous or crypts of epithelium [37]. PrPSc migrates asymptomatically along the peripheral nervous system by axonal transport mechanisms via the vagus and splanchnic nerve [74]. Upon finally reaching the central nervous system (CNS), PrPSc aggregates and accumulates as the neurodegeneration process advances [2, 49]. The sequential detection of PrPSc in lymphoreticular tissues and subsequently along neural projections to the CNS has led to the conclusion that these tissues are involved in the spread of PrPSc to the CNS [41].
Much effort has been directed towards determining whether follicular dendritic cells (FDC) and dendritic cells (DC) are main players in the transfer of PrPSc to the neural cells [4, 12, 13, 75]. Accumulation of PrPSc was confirmed in the DCs following infection [34, 40] and co-culture of the DCs with the primary neural cells resulted in transfer of infection [20, 21, 43]. Mice lacking or having temporarily inactivated FDCs showed reduced or impaired PrPSc accumulation in the lymphoid organs and there was a delay in neuroinvasion [47, 57]. However, the role of other cells such as macrophages, mast cells or peripheral neural cells in accumulating, establishing or transferring prions remains unclear. The responses of these cells to PrPSc over time require further investigation.
11
This study was carried out to investigate the involvement of various cell types in maintenance or accumulation and transfer PrPSc from the intestine to the enteric or central neurons. The rapid and prolonged cellular responses to PrPSc for up to 28 days were investigated using 15 human and murine cell lines including immune, neural, intestinal cell types. After exposure to PrPSc, these cells were analyzed for their ability to sustain, accumulate or degrade PrPSc. Potential transfer of PrPSc from infected cell lines to a neural cell line was also investigated by co-culture experiments. The cell lines showed different responses, ranging from maintenance to transient or persistent accumulation or degradation of PrPSc. The present findings showed that longer incubation of the PrPSc-exposed immune, intestinal and neural cells revealed altered interactions between PrPSc and these cells. PrPSc accumulations were observed in the later stages following incubation with either Chandler or Obihiro strains. When the PrPSc-infected cells were co-cultured with neuroblastoma cells, the transfer of PrPSc to the neural cells was confirmed. Our results suggest that some of these cell types may have a possible role in the maintenance, accumulation and intercellular spread of PrPSc infection to neural cells in vivo.
12
MATERIALS AND METHODS
Cell lines
Fifteen cell lines of different types (six immune, five neuronal, one intestinal, one fibroblast and one epithelial) were used in this study (Table 1). The cells were maintained at 37C in 5% CO2 in Dulbecco's modified Eagle's medium (DMEM;
WAKO, Osaka, Japan) supplemented with 10% fetal bovine serum (FBS; PAA Laboratories, GmbH, Haidmenweg, Austria), 100 U ml−1 penicillin and 100 μg ml−1 streptomycin. A neuroblastoma cell line, N2a-3/EGFP [73], was used as recipient cells for the co-culture experiment. It is a highly PrPSc-susceptible subclone of mouse neuroblastoma N2a cells [14] and carries an Enhanced Green Flurescent Protein gene. N2a-3/EGFP cells were established by transfection of N2a-3 cells with pEGFP-C1 (Clontech, CA, USA) which contains a neomycin resistance gene. N2a-3/EGFP cells were then selected by addition of G418 at final concentration of 1 mg ml−1 and then cells resistant to G418 for 50 passages were selected.
Prion strains
Brain homogenates were prepared from the brains of mice terminally affected with mouse-adapted scrapie strains, Chandler [39] or Obihiro [68]. The infected brains were mechanically homogenized in phosphate-buffered saline (PBS) and diluted to a final concentration of 10% (w/v) in PBS, sonicated and stored at -20 °C until use.
Infection of cells with prion
Cell lines were cultured in 60-mm culture dishes at the optimum cell numbers to provide 60-70 % confluence after overnight incubation. Chandler- or Obihiro-infected brain homogenates equivalent to 1 mg of infected brain tissue were
13
added to the cell dishes and incubated for 24 h before rinsing to remove any unbound PrPSc particles. Negative control cells were prepared in the same way, but without brain homogenate, and used to measure the growth curves of the cells. The cells were then incubated until they were harvested. The cells and supernatants were collected and centrifuged at 1,000 × g for 5 min. Pellets were then stored at -20°C until proteinase K (PK) treatment and analysis by Western blotting.
Co-cultures
At the terminal stage of incubation, the PrPSc-exposed cells were washed, trypsinized and harvested into 15-ml tube. The cells were directly seeded into 10-cm dishes at a high density with N2a-3/EGFP cells at a ratio of one infected cell per one target cell. After 4 days of incubation, the cells were treated with G418 in each passage at a concentration 1 mg ml−1 to remove the donor cells. The recipient N2a-3/EGFP cells were split at 1:5 every 4 days. At each passage, 4/5 of the cells in each dish were collected, PK-treated and analyzed by Western blotting for PrPSc accumulation.
PrPC expression in cells
PrPC expression in the 15 cell lines was examined by collection of duplicates of 5 × 105 of each cell line into 2-ml tubes. Then, 200 µl of lysis buffer (5 mM EDTA, 0.5% Triton X-100, 0.5% sodium deoxycholate, 150 mM NaCl, and 10 mM Tris–HCl, pH 7.5) was added to each tube and kept for 30 min at 4C. Cell debris was removed by centrifugation for 5 min at 1,000 × g. The PrPC was precipitated by adding 800 µl of methanol at -20°C for 15 min before centrifugation at 20,000 × g for 30 min. The cell pellets were boiled in sample buffer (62.5 mM Tris–HCl, pH 6.8, 5% glycerol, 3 mM
14
EDTA, 5% SDS, 4 M urea, 4% β-mercaptoethanol, and 0.04% bromophenol blue) and separated by Western blotting.
PK treatment
Cells were lysed with 300 µl of lysis buffer and kept at 4 °C for 30 min. Cell debris was removed by centrifugation for 5 min at 500 × g. Twenty micrograms per milliliter of PK (Roche Diagnostics, USA) was added to each sample and incubated for 20 min at 37°C. Proteolysis was terminated by the addition of 1 mM Pefabloc (Roche Diagnostics). The samples were then incubated with 0.3% sodium phosphotungstic acid at 37°C for 30 minutes instead of ultracentrifugation as described previously [79]. PK-treated samples were centrifuged at 20,000 × g for 45 min and the resulting pellets were dissolved in sample buffer, boiled and stored at -20°C until loading.
Western blot analysis
After denaturation with sample buffer, proteins were separated using 12% SDS-polyacrylamide gel electrophoresis (Bio-Rad,Tokyo ,Japan). The separated protein bands in the gel were transferred to Immobilon P (Millipore, Billerica, MA, USA), blocked with 5% non-fat milk in 0.1 % Tween-20 Tris-buffered saline, pH 7.5, probed with anti-prion monoclonal antibody (mAb) 31C6 [39] at 1:4,000 followed by incubation with a peroxidase-conjugated anti-mouse antibody (GE Healthcare, Buckinghamshire, UK). Immunodetection was visualized using enhanced chemiluminescence (ECL kit; GE Healthcare Bucks, UK) and exposure to X-ray film. Analysis of ECL images was performed using the public domain ImageJ program (developed at the National Institutes of Health, Bethesda, Maryland, USA) according to the manufacturer’s instructions (URL: http://rsbweb.nih.gov/ij). The PK-treated brain
15
homogenate at terminal stage of Chandler strain-infected mouse was used as control in Western blot analysis.
Imaging of prion in cells by immunofluorescent microscopy
In order to visualize and confirm the cells ability to internalize PrPSc particles, an indirect immunofluorescent assay was used. The cells were seeded in Lab-Tek II chamber slides (2-well slides) (Nalge Nunc International, Naperville, IL, USA) and grown to approximately 50 % density. Cells were exposed to 0.25 mg of Chandler-brain homogenate for 4 or 12 h and then rinsed thoroughly with PBS. Some cell chambers were not exposed to prion and used as negative controls to confirm the efficiency of PrPC removal. Cells were fixed in 200 µl of 4% paraformaldehyde in 4% sucrose for 30 min, followed by neutralization in 0.1 M glycine in PBS for 10 min. The cells were permeabilized by adding 200 μl of 0.1% saponin for 10 min. The wells were then rinsed with 500 μl of PBS and treated with 3 M guanidine thiocyanate for 10 min to denature of PrPC. The cells were blocked with 5% FBS in PBS for 30 min and incubated with 200 μl of 1 µg ml−1 mAb anti-PrP 132 [39] at 4°C overnight. On the next day, these slides were rinsed and incubated with anti-mouse IgG conjugated to Alexa Fluor 488 (Invitrogen, Carlsbad, CA, USA) at a 1:400 dilution for 1 h. Slides were then thoroughly rinsed with PBS and coverslip mounted with 50% glycerol in PBS and examined for the endocytosis of PrPSc aggregates using a fluorescence microscope (Nikon corporation, Tokyo, Japan). Five images were captured for each cell line. The images were then evaluated by blind scoring for the amounts of internalized PrPSc. Scores obtained were averaged and summarized in Table 2.
16
The proteasome activity in the cells was measured using a 20S Proteasome Assay Kit (Cayman Chemical Co., MI, USA) according to the manufacturer's protocol. Briefly, triplicates of 5 × 104 cells per well in 100 µl of culture medium were seeded in a 96-well plate and incubated overnight. The next day, cells were treated with Chandler-infected brain homogenate at 1 mg ml−1 for 6 h and the plate was centrifuged at 500 × g for 5 min. The culture medium was aspirated and 200 µl of 20S Proteasome Assay Buffer was added to each well. The plate was centrifuged again at 500 × g for 5 min. Supernatants were aspirated and 100 µl of the 20S Proteasome Lysis Buffer was added to each well. The plate was incubated with gentle shaking for 30 min at room temperature. The plate was again centrifuged at 1,000 × g for 10 min and 90 µl of the supernatant was transferred from each well to a corresponding well in a black 96-well plate and mixed with 10 µl of Assay Buffer or 20S Inhibitor Solution. Following addition of 100 µl of positive control to the corresponding wells in the black plate, 10 µl of the substrate solution was also added to each well and the plate was incubated at 37°C for 1 h. Fluorescent intensity of each well was measured (excitation 360 nm, emission 480 nm) using Wallac 1420 ARVOsx-1 Fluoroscan (Perkin Elmer Life Sciences, Tokyo, Japan). Differences between proteasome activity in infected and normal cells were analyzed using Student’s t-test and were considered statistically significant at p < 0.05.
17
RESULTS
Characterization of initial cell responses to prion exposure
The short-term cell responses to early PrPSc infection in 15 cell lines (Table 1) exposed to 1 mg of the Chandler-infected brain homogenate were investigated. Treated cells were harvested at 1, 2, 3 and 5 days of exposure, treated with Proteinase K (PK) and assayed by Western blotting (Fig. 1). The cell lines varied greatly in their response to infection with PrPSc. The neuroblastoma N2a-3 cells and the hippocampus GT1-7 cells showed increasing accumulation of PrPSc over the 5 days of incubation (Fig. 1). On the other hand, two other neural cells, TR6Bc1 and RT4-D6P2T, showed strong catabolic effect on the PrPSc (Fig. 1). Only the neuroblastoma NB41A3 cells could sustain PrPSc, but it was at a very low level. Western blotting results showed a trend mainly towards the degradation of PrPSc (RAW, J774, J588L and P1.HTR), although two immune cell lines (P388-D1 and EL4) maintained moderate levels of PrPSc over the 5 days. P388-D1(IL-1) macrophages showed a gradual accumulation of PrPSc until day
5. A similar trend was also observed in the IEC-18 intestinal epithelial cell line. These results prompted us to investigate the cellular characters that may affect these responses and the later phases of the cell response to PrPSc.
Endocytosis of PrPSc by cell lines
To characterize factors related to the cell responses, author compared the cells in different ways. The ability of cells to endocytose PrPSc was investigated using indirect immunofluorescence. Chandler-infected brain homogenate was added to each cell line for 4 or 12 h, rinsed, fixed and labeled with the anti-PrP 132 antibody. Microscopic analysis revealed that PrPSc was present as punctate, small aggregates
18
generally localized to both cytoplasmic and perinuclear regions (Fig. 2). In general, all cell lines internalized PrPSc particles to some degree. However, only 30%–50% of any of the cell types were positive for PrPSc, suggesting that endocytosis only occurs in a subpopulation of cells. In regard to cell type, most neural cell lines except NB41A3 showed high levels of PrPSc endocytosis (Fig. 2 & Table 2). Most of the immune cell lines showed high levels of internalized PrPSc particles (Fig.2; RAW, J447, P388-D1(IL-1) & EL4). The intestinal cell line IEC-18 showed also high uptake ability
at 12 h, but NIH-3T3 and HeLa cells showed only moderate levels of PrPSc (Fig. 2).
PrPC expression in different cell lines
To evaluate the relative expression of PrPC in the cell lines and its relationship to PrPSc reactions, the author analyzed 5 × 105 cells of each cell line by Western blotting (Fig. 3 & Table 2). Three of the 5 neural cell lines expressed significant levels of PrPC while the other 3 neural cell lines and most immune cells showed low to undetectable levels of PrPC (Fig. 3a & Table 2). The graphical representation showed that the intestinal IEC-18 cells had the highest level of expression among the cell lines (Fig. 3b). When compared to the cell responses to PrPSc (Fig. 1), the expression of PrPC by each cell line may reflect its ability to maintain or accumulate PrPSc, as observed in some of the neural and intestinal cells.
Proteasome activity in the cell lines as an indicator of degradation ability
To evaluate the relationship between cellular response to PrPSc and degradation ability, the proteasome activity was measured in eight cell lines. Interestingly, the highest proteasome activity was measured in the neural cells (N2a-3, RT4-D6P2T & TR6Bc1), followed by moderate activity in the intestinal IEC-18 and the fibroblast
19
NIH-3T3 cells (Fig. 3c & Table 2). The immune cell lines (RAW, P388-D1(IL-1) and
P1.HTR) showed low proteasome activities that may indicate dependency of these cells on a different system for degradation of PrPSc. Most of the cell lines (except N2a-3) lost some level of proteasome activity when infected with PrPSc.
Characterization of the late responses to PrPSc exposure during prolonged incubation
Prion diseases progress slowly in vivo and many cells are long lived. Therefore, the author next monitored cell responses to PrPSc following incubation for up to 28 days. The responses of eight cell lines (N2a-3, GT1-7, TR6Bc1, P1.HTR, P388-D1(IL-1), J588L, IEC-18 & NIH-3T3) to exposure to Chandler-infected brain
homogenates were analyzed. The cells were exposed to 1 mg of Chandler-infected brain homogenate for 12 h and then rinsed and continuously incubated for 28 days without passage. However, they were supplemented with 1.5 ml of fresh medium every 2 days. Dishes of the treated cells were harvested at successive days until 28th day post exposure and treated with PK. Western blotting revealed that six cell lines (TR6Bc1, P1.HTR, P388-D1(IL-1), J588L, IEC-18 & NIH-3T3) showed similar patterns of
reaction to PrPSc with little variation. Most of the cell lines were found to have a biphasic response (Fig. 4 & Table 3). In the first phase, a gradual disappearance of PrPSc occurred (TR6Bc1, J588L & NIH-3T3), which was preceded by a transient increase in the level of PrPSc in the first few days after treatment in some cell lines (P1.HTR, P388-D1(IL-1) & IEC-18). In the second phase, all of these cell lines showed a
reappearance and gradual accumulation of the PrPSc bands at the terminal stage of incubation (Fig. 4a & b). The N2a-3 and GT1-7 neural cells were the only cells that constantly maintained the same pattern of PrPSc accumulation until the day 18 of the
20
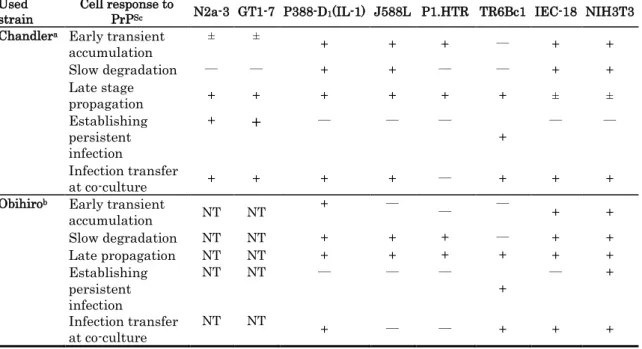
experiment (data not shown). A comparative summary of the long-term responses of these cell lines to the Chandler strain of PrPSc is shown in Table 3. Moreover, six out of the eight Chandler-infected cell lines showed a slow rate of degradation in the first phase that led to continued detectability of PrPSc for relatively long times. This may increase the chance of in vivo transfer of infection to other cells, including the neural cells. These results also revealed that one neural cell line (TR6Bc1) established persistent infection when continuously passaged after the long incubation period (Table 3).
Late cellular accumulation of PrPSc in relation to scrapie strain
To determine whether the PrPSc cell responses were strain specific, autor repeated the experiment using the Obihiro strain. Six cell lines (TR6Bc1, P1.HTR, P388-D1(IL-1), J588L IEC-18 & NIH-3T3) were treated in the same way using 1 mg of
Obihiro-infected brain homogenate of a mouse at the terminal stage of the disease. Three (TR6Bc1, IEC-18 & NIH-3T3) out of the six cell lines showed very similar results to those obtained with the Chandler strain (Fig. 4) with two main phases of responses. A transient early increase in the levels of PrPSc was observed (NIH-3T3 & IEC-18) followed by a gradual decrease of PrPSc (TR6Bc1, NIH-3T3 & IEC-18) and a second phase of reappearance of PrPSc bands by the end of the experiment (Fig. 4c & d). In IEC-18 cells, degradation was slow, allowing detection of PrPSc for relatively longer times (Fig. 4d & Table 3). Three other cell lines (P1.HTR, P388-D1(IL-1) & J588L)
were found to maintain high levels of PrPSc throughout the incubation period, without a degradation phase, leading to accumulation during the later phase (Fig. 4d). These results show that accumulation of PrPSc at the late stage of incubation may not be strain specific.
21
Transfer of PrPSc from infected cells to neural cells after the long incubation
To investigate the potential cell-to-cell transfer of accumulated PrPSc to neural cells, the author co-cultured PrPSc–loaded cells with neural cells in vitro. Cells at the terminal stage of incubation were co-cultured with an equal number of N2a-3/EGFP cells and passed up to 15 times. The transfer of PrPSc from most of the donor cell lines (N2a3, GT1-7, P388-D1(IL-1), J588L, TR6Bc1, IEC-18 & NIH-3T3) to the
N2a-3/EGFP recipient cells was successful (Table. 3). After a number of passages, PrPSc accumulated in N2a-3 cells as shown in the representative Western blots (Fig. 4). Obihiro-infected cells were also co-cultured with N2a-3/EGFP and the transfer of PrPSc was successful (Table 3).
Establishing persistent PrPSc infection in the cell lines after short and prolonged infection
After the end of incubation periods, one dish of each cell line infected with the PrPSc was subjected to 15 successive times of passage to check cells ability to establish persistent PrPSc infection. The cell lines which are able to establish persistently PrPSc infection are those which continuously self-replicate the PrPSc after a number of splits into new dishes. While the cell lines which lose their PrPSc load permanently after a number of splits and do not restore PrPSc replication till the 15th passage are considered unable to establish persistent PrPSc infection. In the short-term incubation experiment, two neural cell lines N2a-3 and GT1-7, established persistent infection at passages 3 and 6, respectively. On the other hand, persistent PrPSc infection was not detected in any of the other cell lines for up to 15 passages (Table 2). In case of the prolonged incubation experiment, three (N2a-3, GT1-7 & TR6Bc1) out of the eight
22
Chandler-infected cell lines became persistently infected after 2, 5 and 7 passages, respectively. Only two (TR6Bc1 & NIH-3T3) out of the six Obihiro-infected cell lines were persistently infected after 6 and 2 passages, respectively (Table 3).
23
DISCUSSION
In this study, the author investigated the potential role of a variety of cell types in the biological processing, maintenance and transfer of PrPSc to neural cells. Early and late responses to PrPSc were investigated in cell lines from several origins. Short-term incubation of cell lines with PrPSc resulted in a variety of responses with three main patterns identified using Western blotting. These were accumulation, maintenance and degradation of PrPSc. Consistent with previous reports [65, 70, 73], N2a-3 and GT1-7 neural cells supported the accumulation of PrPSc. The P338D1(IL-1) macrophage and
IEC-18 intestinal cell lines also accumulated PrPSc during the first few days of incubation, but this was later proved transient. Some other cell lines preserved steady amounts of PrPSc without significant fluctuation during the 5 days of incubation. The third pattern of response was the catabolism of the endocytosed PrPSc which was mostly found in immune cell lines but at variable rates. This type of response was also reported in a variety of cell line models that degrade PrPSc upon exposure [34, 45, 52, 63, 64].
The variation in the responses to PrPSc suggests the involvement of some cellular factors. The present results indicate that the ability and speed of uptake of PrPSc, the normal level of PrPC expression, and the cell’s ability to produce sufficient PrPC for conversion to PrPSc may be important for accumulation of PrPSc. The level of proteasome activity in each cell line may also be an important factor in the degradation of PrPSc, especially the ubiquitin-proteasome system (UPS), which is believed to be a primary cellular control system for degrading the misfolded proteins and toxic aggregates in cells [29]. According to the present results, this may be most applicable to the non-immune cells that showed high proteasome activity (Fig. 3c). However, the immune cells showed significant degradation of PrPSc despite their low proteasome
24
activity. This suggests that different degradation systems are being used by these cells. The 20S proteasome activity was significantly suppressed in PrPSc-infected cells, supporting a previous observation that PrPSc impairs the cell UPS by interaction with the 20S proteasome [19, 42].
Taken together, data suggest an important relationship between these cell characteristics and responses to PrPSc. However, these characteristics may be unrelated to their effect on the cell response. The cell’s ability to take up PrPSc
does not require high expression of PrPC, although it is necessary for persistent PrPSc formation. This is consistent with a previous study [31] that confirmed no interactive effect between these observations. However, PrPSc-infected brain was found to have a significant adverse effect on the 20S proteasome degradation system of the cell. The balance between the degradation and accumulation rate of PrPSc may also be an important factor for the final form of cellular reaction. Similarly, a dynamic equilibrium between synthesis and degradation of PrPSc in the infected cells is thought to determine the fate of the PrPSc in infected cell [53]. However, these may not be the only factors affecting the cell responses to prion infection. Other micro-environmental and as yet unidentified factors may be involved [31].
The author investigated the cellular responses to PrPSc exposure over a prolonged incubation period because prion diseases generally require long incubation periods to reach the brain and cause symptoms in vivo. Accumulation of PrPSc in the terminal stages of incubation was found in all cell types tested in this study by long-term incubation after 12 h exposure to Chandler PrPSc. When Obihiro-infected brain homogenate was used in the same experiment, similar responses were found in half of the cell lines and some variations were observed in the other half. This shows
25
that some variations in the cell maintenance or accumulation of PrPSc during infection may be found when different cell types or scrapie strains are used. However, a previous study [77] confirmed that the acute conversion of cellular PrPC to PrPSc is independent of the cell type and scrapie strain. There are two possible explanations for this phenomenon. The first is presence of subpopulation of cells that accumulate some undetectable level of PrPSc. The observed increase in PrPSc could be a result of these cell subpopulations dividing, and this may be important in establishing persistent infection [24, 31, 82]. This theory may be applicable to the cell lines in which continuous multiplication was observed throughout the incubation time course. However, one cell line (IEC-18) stopped dividing within a few days of reaching complete confluence in the culture dish. The second possible explanation is that there may be a gradual inhibition of the cell’s degradation systems. This may be caused by the PrPSc
itself, which the author and others have shown can impair the cellular 20S proteasome system [19, 42].
The development of persistent infection was examined by continuously splitting cells at the terminal stage of incubation. Persistent PrPSc infection was established in three cells lines (N2a-3, GT1-7 & TR6Bc1). The N2a-3 neuroblastoma and GT1-7 hippocampus cells have previously been shown to develop persistent infectivity [14, 70]. Here, the author introduce a new neural cell line (TR6Bc1) that can also become persistently infected with either Chandler or Obihiro strains of PrPSc when incubated with the infected brain homogenate for long periods. This is the first record of NIH-3T3 cells developing persistent PrPSc infection with the Obihiro strain. This cell line has previously become persistently infected with only the 22L strain of scrapie [78].
26
Late-stage PrPSc accumulation or persistent infection increases the potential transfer of infection from PrPSc-loaded cells to adjacent cells or neurons in vivo. In addition, the slow rate of early PrPSc degradation and continued detection of PrPSc for up to 15 days in some cells may also increase the likelihood that PrPSc is presented to other cells. These findings give insight into the later stages of prion invasion to lymphoreticular, intestinal and neural cells and suggest that a wider range of cell types are involved in disease development. It is possible that, following uptake of PrPSc, immune cells move inside the body and make contact with nerve endings, thereby transferring PrPSc to neural cells. However, it is more likely that lymphoreticular and intestinal cells are subject to uptake and transfer of exogenous agents, including prions. In support of this hypothesis, we have previously shown that the absorption of various exogenous particles, including recombinant mouse prion protein, from the small intestinal lumen occurred at Peyer’s Patches [46]. Transport of these particles via blood, mesenteric lymph nodes and liver was also observed in this study.
The transfer of infection from PrPSc-loaded cells was investigated by co-culturing neural cells with the cells of interest at the terminal stage of incubation. Several cell lines were found to transfer infection confirming our hypothesis. The transfer of PrPSc occurred via cell-to-cell direct contact, close contact with dendrites or direct link to neurons via tunneling nanotubes (TNT) as previously described [30, 43]. The transfer of PrPSc from DCs to neural cells has also been investigated [21] and the results suggested that prion transfer occurred through TNT-like structures shown to connect donor and recipient cells. Taken together, these results suggest that the accumulation of PrPSc in the recipient cells may be in vitro evidence of a possible involvement of various cell types, including immune and intestinal cells, in sustaining,
27
accumulating and transfer of the infection to the CNS in vivo. Some of these cells may act as reservoirs for PrPSc particles until they have the opportunity to transfer to a neural cell, which in turn, transfers it to another neural cell until the target organ is reached.
28
SUMMARY
The mechanisms and processes of the uptake, intracellular trafficking and intercellular spread of PrPSc, and its transfer to the neural cells are not clearly defined. The involvement of the immune, intestinal, mast or peripheral neural cells in this process also remains unclear. The role of these cell types in the accumulation and transfer of PrPSc to neural cells was investigated following short and prolonged exposure to PrPSc Chandler and Obihiro strains of scrapie for up to 28 days. Fifteen cell lines of human and murine immune, neural, intestinal and other cell types were investigated. In addition, the potential transfer of PrPSc from these infected cells to a neuroblastoma cell line was investigated using co-culture experiments. Certain immune, intestinal and neural cells accumulated PrPSc for up to 28 days post infection after transient degradation phases. When co-cultured with the N2a-3/EGFP neuroblastoma cells for 4 days followed by several passages, the immune, intestinal and the neural cell lines were able to transfer infection to neural cells. The present results suggest that some of these cell types may have a role in accumulating and intercellular spread of PrPSc infection to the neural cells in vivo.
33
Table 1. List of cell lines used in this study
Cell Type Name Origin Species
Neural cells N2a-3 Neuroblastoma Mouse
NB41A3 Neuroblastoma Mouse
GT1-7 Hypothalamic nerve cells Mouse
RT4-D6P2T Schwann cell, peripheral nerves Rat
TR6Bc1 Trigeminal nerve, Schwannoma Mouse
Immune cells
RAW Macrophage Mouse
J774 Monocyte-macrophage Mouse
P388-D1 Macrophages of lymphoid neoplasma Mouse
P388-D1(IL-1) IL-1 producing P388-D1 derivative Mouse
EL4 Lymphoma of T-cell Mouse
J588L Bone marrow (Myeloma) Mouse
P1.HTR Mastocytoma Mouse
Intestinal cell IEC-18 Small intestinal epithelium Rat
Other cells NIH-3T3 Embryonic fibroblast Mouse
34
Table 2. Cell line characters and their responses to prion exposure
++++; Very high, +++; High, ++; Moderate to high, +; Moderate, ±; Low, -; Negative NT; Not tested
a; Relative PrPC expression (Fig. 3a,b)
b; Relative uptake ability as estimated by IFA (Fig. 2)
c; Relative proteasome activity (Fig. 3c)
d; Degradation of PrPSc during 5 days of incubation(Fig. 1)
e; Maintaining of PrPSc level during 5 days of incubation(Fig. 1)
f; Propagation of PrPSc level during 5 days of incubation(Fig. 1)
Characters
Immune cells Neural cells Intestinal Other cells
J774 RAW P388-D1 P388-D1(IL-1) J588L EL4 P1.HTR N2a-3 NB4A3 RT4 TR6Bc1 GT1-7 IEC-18 NIH3T3 HeLa
PrPC Expressiona ± ± ± ± ± + ± +++ ± + +++ ++ ++++ + ± Uptake abilityb ++ +++ ++ ++ + ++ ++ + ± ++++ +++ ++ ++ ++ + Proteasome Activityc + + NT + NT NT ++ ++ NT ++++ +++ NT +++ ++ NT Degrading PrPSc d ++ +++ ± ― + ± + ― ± ++ +++ ― ― + ± Maintaining PrPSc level e ― ― + + ― + ± + ― ― ― + + + ± Propagation of PrPSc f ― ― ― + ― ― ― + ― ― ― + + ― ― Persistent infection ― ― ― ― ― ― ― + ― ― ― + ― ― ―
35
Table 3. Summary of cell responses to prion obtained during long-term incubation with two strains
NT; not tested
+; Positive, ±; Weak Positive, -; Negative a ; Cell responses to Chandler PrPSc (Fig. 4a,b) b; Cell responses to Obihiro PrPSc (Fig. 5a,b) Used
strain Cell response to PrPSc N2a-3 GT1-7 P388-D1(IL-1) J588L P1.HTR TR6Bc1 IEC-18 NIH3T3
Chandlera Early transient
accumulation ± ± + + + ― + + Slow degradation ― ― + + ― ― + + Late stage propagation + + + + + + ± ± Establishing persistent infection + + ― ― ― + ― ― Infection transfer at co-culture + + + + ― + + +
Obihirob Early transient
accumulation NT NT + ― ― ― + + Slow degradation NT NT + + + ― + + Late propagation NT NT + + + + + + Establishing persistent infection NT NT ― ― ― + ― + Infection transfer at co-culture NT NT + ― ― + + +
36
CHAPTER 2
Early Transient Accumulation of PrP
Scin Immune Cells;
Characterization and Affecting Factors
37
INTRODUCTION
Transmissible spongiform encephalopathies (TSE, prion diseases) are invariably fatal neurodegenerative diseases affecting animals and humans. These diseases develop following conformational changes in the cellular prion protein (PrPC) which result in the misfolded isoform of prion protein (PrPSc) at post-translational modification [22]. Animal TSE infections have arisen through feeding with PrPSc-contaminated animal food. After oral uptake, PrPSc first accumulates in gut-associated lymphoid tissues (GALT) such as Peyer’s patches in the intestines before neuroinvasion occurs [48, 58].
Unlike other infectious diseases, TSEs do not elicit specific immune responses because the infectious agent is composed of a protein with a primary structure identical to a host encoded protein [10, 56]. Moreover, the immune system appears to support rather than impair the propagation of prions or acts as a protective fortification in the process of prion infection [5, 9]. Although, comprehensively studied, the roles of mononuclear phagocytes in prion pathogenesis remain a matter of ongoing debate [81]. Several studies suggest that cells of the immune system, including macrophages, support the replication and spread of prions to the central nervous system [11, 62]. Peripheral macrophages have demonstrated both accumulation [11, 26, 33, 35, 44, 57] and proteolysis of PrPSc [11, 33]. Macrophages in lymphoid follicles have been shown to contain PrPSc at early stages of TSE infection [36]. A study of the in vitro effects of prion on mouse macrophages suggested that macrophages play a role in the transportation and replication of the infectious agent [84].
Cell culture models are important for in vitro investigation of PrPSc and nowadays, they are the fastest and most common ex vivo approach for studying PrPSc. Forms of accumulation or proteolysis of PrPSc can be observed in these cells following exposure to PrPSc. Studying these forms of cellular interaction with PrPSc may also help to elucidate the mechanism of PrPSc uptake, propagation and transmission between cells, which is a crucial step towards understanding the pathogenesis of prion diseases. Cell culture models also present various advantages in studying the disease agent itself, including the ability to analyze its biological properties and the factors controlling its propagation or degradation . Upon studying PrPSc using this tool, some cell culture
38
models supporting PrPSc replication have been established [7, 55, 69, 76] and are now used widely for further studying this agent.
In chapter 1, the author investigated the responses of various cell types including immune, neural, intestinal and fibroblast cells to PrPSc for up to 28 days. Different patterns of response to PrPSc exposure were identified. These forms varied between accumulation, degradation or mixed accumulation and degradation of PrPSc. Some of the cell types showed a characteristic cellular response in the form of transient propagation of PrPSc which preceded its clearance. The aim of our study was to characterize the transient PrPSc accumulation and its significance in disease development. Murine macrophage cells were exposed to PrPSc and showed a characteristic and specific cellular reaction in the form of a unique transient accumulation of PrPSc in the first 8-12 h post-exposure before its clearance at a later stage. Further investigation of this phenomenon provided insights into the relationship between PrPSc formation in cells and some related factors that may influence its transient accumulation. These factors include cholesterol homeostasis, temperature, the degradation power of the cell and availability of sufficient PrPC. My in vitro results suggest that immune cells, especially macrophages, despite their important role as main protective shield against prion disease, they may potentially be involved in the accumulation and intercellular spread of PrPSc infection. This may be possible if their proteolytic function was impaired due to extracellular factors or cell death before the complete clearance of PrPSc could be fulfilled.
39
MATERIALS AND METHODS
Cell lines
Two mouse macrophage cell lines (Raw and J774) and primary macrophage cells of mice bone marrow were used in this study. The bone marrow–derived macrophages (BMMs) were prepared from BALB/c mice by the method described previously [80]. After culture in L-cell–conditioned medium, BMMs were resuspended in RPMI 1640 containing 10% FBS (PAA Laboratories, GmbH, Haidmenweg, Austria). The cells were cultured at 37 C and 24 C in 5% CO2 in DMEM (WAKO, Osaka,
Japan), supplemented with 10% FBS, 100 U ml−1 penicillin and 100 μg ml−1 streptomycin.
E. coli cell lysate and microspheres
Escherichia coli (E. coli) ATCC11775 was incubated in L-broth (0.5g of yeast extract, 1g of tryptone and 0.5g of NaCl) overnight. The cell pellets were collected and 1 × 109 cells were resuspended in 1ml of PBS and boiled for 10 min. The E. coli cell lysate and polybead polystyrene microspheres (2.5% Solids-Latex, PA: 1.0 µm, Polysciences, Inc, Warrington, UK) were used as stimulants in the quantitative real-time PCR and Western blot analysis experiments and they were added to cells at a rate of 10µl/dish.
Prion strains
Brain homogenates were prepared from mice terminally affected with mouse-adapted scrapie strains, Chandler [39], MHM2/Chandler [83] or Obihiro [68]. The MHM2/Chandler strain is able to infect a transgenic mouse strain which expresses a chimeric PrP derived from mouse and hamster. The MHM2/Chandler brain samples were kindly provided by Dr. T. Yokoyama (Prion Disease Research Center, National Institute of Animal Health, Ibaraki, Japan). Infected and normal mouse brains were mechanically homogenized in PBS and diluted to a final concentration of 10% (w/v) in PBS, sonicated and stored at -20°C until use.
40
Macrophage cells were cultured in 60-mm culture dishes at the optimum cell numbers to provide 60 - 70% confluence after overnight incubation. Chandler-, MHM2/Chandler- or Obihiro-infected brain homogenates equivalent to 0.5 mg of infected brain tissue were added to the cell dishes and incubated. After 2 hours, dishes were rinsed twice to remove excessive and unbounded brain homogenates. Negative control cells without brain homogenate were prepared in the same way to measure the growth curves of the cells. The cells were then incubated until they were harvested at the indicated intervals. The cells and supernatants were collected and centrifuged at 1,000 × g for 5 min. Pellets were then stored at -20°C until Proteinase K treatment and Western blotting analysis.
Cholesterol depletion
To evaluate the influence of cholesterol metabolism on PrPSc infected-cells, U18666A (Sigma, St. Louis, MO) was used as a cholesterol synthesis and intracellular transport inhibiting compound [27]. The raw cells were treated with 5 µg of U18666A/dish and kept overnight before the addition of 0.5 mg of Chandler brain homogenate. After 2 hours, dishes were rinsed twice to remove excessive and unbounded brain homogenates. The cells were incubated with 5 µg of U18666A/dish at 37 °C for 0, 4, 8, 12, 24, 48, 72 and 120 h, lysed with PK and assayed for the presence of PrPSc by Western blotting.
Proteinase K treatment
Cells were lysed with 300 µl of lysis buffer (10 mM Tris/HCl, pH 7.5, 150 mM NaCl, 0.5% Triton X-100, 0.5% sodium deoxycholate, and 5 mM EDTA, pH 8.0) on ice for 30 min. The lysate was centrifuged at 500 × g for 5 min and the supernatant was recovered into a 2-ml tube. Proteinase K (Roche Diagnostics) was added to each sample at a concentration of 20 µg ml−1 and incubated for 20 min at 37 °C. Proteolysis was terminated by the addition of 1 mM Pefabloc (Roche Diagnostics). The samples were then incubated with 0.3% sodium phosphotungstic acid at 37 °C for 30 min instead of ultracentrifugation as described previously [79]. PK-treated samples were centrifuged at 20,000 × g for 45 min and the resulting pellets were dissolved in sample buffer, boiled for 10 min, and stored at -20°C until loading.
41
Western blot analysis
After denaturation with the sample buffer, proteins were separated using 12% SDS-polyacrylamide gel electrophoresis. The separated protein bands in the gel were transferred to Immobilon P (Millipore), blocked with 5% non-fat milk in 0.1% Tween-20 Tris-buffered saline, pH 7.5, probed with anti-prion monoclonal antibody (mAb) 31C6 [39] at 1:4,000 or 3F4 [38] at 1:4,000 followed by incubation with a peroxidase-conjugated anti-mouse antibody (GE Healthcare). Immunodetection was visualized using an enhanced chemiluminescence kit (ECL; GE Healthcare) and exposure to X-ray film. Analysis of ECL images was performed using the public domain Image-J program (developed at the National Institutes of Health, Bethesda, MD, USA) according to the manufacturer’s instructions.
Proteasome assay
The 96-well culture plates of Raw cells incubated overnight were treated with PrPSc brain homogenate and incubated at 37 °C or 24 °C for 12 h. The proteasome activity in the infected and uninfected cells incubated at different temperatures was measured using a 20S Proteasome Assay Kit (Cayman Chemical Co., MI, USA) according to the manufacturer's protocol. Briefly, triplicates of 5 × 104 cells per well in 100 µl of culture medium were seeded in a 96-well plate and incubated overnight. The next day, cells were treated with 100, 30 or 0 µg Chandler-infected brain homogenate and the plates were incubated at 37 °C or 24 °C for 12 h. The plates were then centrifuged at 500 × g for 5 min. The culture medium was aspirated and 200 µl of 20S Proteasome Assay Buffer was added to each well. The plate was centrifuged again at 500 × g for 5 min. Supernatants were aspirated and 100 µl of the 20S Proteasome Lysis Buffer was added to each well. The plate was incubated with gentle shaking for 30 min at room temperature and again centrifuged at 1000 × g for 10 min before 90 µl of the supernatant was transferred from each well to a corresponding well in a black 96-well plate and mixed with 10 µl of assay buffer or 20S inhibitor solution. Following the addition of 100 µl of positive control to corresponding wells in the black plate, 10 µl of the substrate solution was also added to each well and the plate was incubated at 37 °C for 1 h. Fluorescent intensity of each well was measured (excitation 360 nm, emission 480 nm) using Wallac 1420 ARVOsx-1 Fluoroscan (Perkin Elmer Life Sciences,
42
Tokyo, Japan). Differences between proteasome activity in infected and normal cells were analyzed using Student’s t-test and were considered statistically significant at p < 0.05.
RNA extraction and Quantitative real-time PCR
Total RNA was extracted from treated and untreated cell pellets using the Qiagen RNeasy Kit protocol (Qiagen, Tokyo, Japan). RNA was quantified using a NanoVue spectrophotometer (GE Healthcare, Tokyo, Japan) at wavelengths of A230/A260 nm and cDNA was then synthesized using the SuperScript III First-Strand
Synthesis SuperMix Kit (Invitrogen) according to the manufacturer’s protocol. As negative controls, 1 mg of each RNA sample was taken through the reverse transcription procedure but without the addition of reverse transcriptase. Quantitative RT-PCR was performed using Fast SYBR Green Master Mix (Applied Biosystems) according to the manufacturer’s instructions. PCR was carried out in 48-well plates on cDNA equivalent to 0.5 µg of total RNA. Thermal cycling conditions were 2 min at 50 °C and 10 min at 95 °C followed with 45 cycles at 95 °C for 3 s and 60 °C for 30 s. Data were collected using the StepOne analytical thermal cycler (Applied Biosystems, Tokyo, Japan). A no template negative control was included in the reaction plate to confirm that no products were synthesized. Each reaction was run on ethidium-stained gels to confirm that a specific product was being measured. The PRNP primers used
were mPrion-F: ACGACTGCGTCAATATCACCAT and mPrion-R:
GGTACTGGGTGACGCACATCT. Primers for the internal standard control gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were AmGAPDH-F: TGCACCACCAACTGCTTAG3 and AmGAPDH-R: GGATGCAGGGATGATGTTC. Chandler-PrPSc, normal brain homogenate, heat-inactivated E. coli cell lysate and polybead polystyrene microspheres were used as different antigen, stimulants for the macrophage cells.
43
RESULTS
The early transient accumulation of PrPSc in macrophages and its origin
The early cellular responses to 0.5 mg of Chandler-infected brain homogenate were investigated in PK-treated and PK-untreated Raw cells. Both cell groups were harvested at 0, 4, 8, 12, 24, 48, 72 and 120 h after PrPSc exposure and assayed by Western blot analysis. An incremental accumulation of PrPSc over the first few hours of incubation was observed with maximum levels of both PrPC and PrPSc at 4 h post-exposure in the group assayed without PK treatment (Fig. 5a). However, in the PK-treated group, the peak of PrPSc was recorded at 12 h. The time interval between the two peaks indicates conversion of PrPC into PrPSc in macrophages. To investigate whether these results were specific to the Raw cell line and to the Chandler strain, the author conducted similar experiments using J774 macrophages and primary macrophages from the mouse bone marrow. In addition, Obihiro and MHM2/Chandler strains of PrPSc were used to infect these cells. Almost identical patterns of the biphasic response to Chandler PrPSc seen in the Raw cell line were also observed with J774 and bone marrow macrophages and with the MHM2/Chandler strain of PrPSc (Fig. 5b). However, when the Obihiro strain was used, the peak of PrPSc was recorded at 24 h post-exposure and the clearance rate was slower than that of the Chandler strain (data not shown). These results confirm that this phenomenon of early transient accumulation of PrPSc is not specific to the Raw cells or to Chandler PrPSc.
The accumulated PrPSc was further investigated to elucidate whether it was a de novo product of the macrophage cells or if it originated from the PrPSc inoculum of the mouse brain homogenate. A similar experiment to that shown previously was conducted using a mouse brain homogenate of MHM2/Chandler strain which can easily be distinguished from the other PrPSc strains using a specific mAb. Raw macrophage cells were treated with Chandler or MHM2/Chandler infected brain homogenates. The 3F4 mAb was used to identify the MHM2/Chandler strain, while 31C6 mAb was used to identify both the Chandler and MHM2/Chandler strains either introduced from outside the cells or converted inside them as de novo products. Western blot analysis revealed a correlation between the curves of PrPSc accumulation of both mAb with the peak recorded at similar times post-exposure (Fig. 5b). This result shows that the
44
accumulated PrPSc is mostly the same as the inoculated PrPSc of the brain homogenate and not de novo produced by the cells.
Characterization of PrPSc degradation in macrophages at variable incubation temperatures
As a factor that may affect cell viability, metabolism or total activity, the effect of temperature on PrPSc cellular accumulation and degradation in Raw cells was investigated at different incubation temperatures. After infection with 0.5 mg of Chandler brain homogenate, cells were incubated at 37 °C or 24 °C for 5 days. After cell harvest, PK-treatment and Western blot analysis, differences in the PrPSc accumulation curves between the two groups were identified. The cells incubated at 37 °C gradually cleared PrPSc after 24 h, while the cells incubated at 24 °C showed no degradation of PrPSc up to the 5th day post-exposure (Fig. 6a). Interestingly, when the cells were moved from the 24 °C to 37 °C after the 5th day post exposure, the cellular degradation power was restored and PrPSc was completely degraded by the 9th day (data not shown). These data show that the ability of the macrophages to degrade PrPSc was impaired at the lower than optimum temperature.
To confirm this observation, one of the main PrPSc degradation systems of cells, 20S proteasome, was assayed in the Raw cells at different incubation temperatures. The proteasome activity was assayed in the infected and normal cells and the results revealed that it was significantly decreased in both infected and normal cells that were incubated at 24 °C compared to those incubated at 37 °C (Fig. 6b). These data confirm that the impaired macrophage degradation ability promotes further and prolonged accumulation of PrPSc in these cells. This high and prolonged accumulation of PrPSc increase the chance of transfer of the infection from these cells to other immune or neural cells in vivo.
Effect of additional PrPC on the accumulation of PrPSc in Raw cells
To investigate if additional PrPC from an external source may affect the pattern of PrPSc accumulation and clearance, the Raw cells were treated with PrPSc brain homogenates with or without additional amounts of normal brain homogenate. Treated cells with PrPSc-brain were incubated for 2 h to allow engulfment of PrPSc before
45
washing to remove any unbound PrPSc. Normal mouse brain homogenate were then added at 2 mg/dish as an additional source of PrPC and the dishes were incubated at 37 °C for 5 days. The cells were harvested, treated with PK and analyzed by Western blotting. Results showed that PrPSc remained for relatively longer time in the cells that were treated with additional normal brain homogenate compared to the complete disappearance after 3 days in the untreated cells (Fig. 7a). This data indicates that PrPC availability may be important to support the process of conversion of PrPC intoPrPSc in these cells. However, because of the saturation of phagocytic capacity of the cells, the PrPSc propagation rate could not be supported by enough PrPC to go on higher rate than the degradation.
To further characterize the accumulation of PrPSc in the macrophages, the effect of the cholesterol synthesis and intracellular transport inhibitor, U18666A, on the uptake and accumulation of PrPSc in macrophages was investigated. Raw cells were incubated with U18666A compound and exposed to PrPSc for 2 h. The Western blot analysis revealed a significant decrease in the endocytosed and accumulated PrPSc levels in the cells treated with U18666A (Fig. 7b). This decrease is probably attributed to the compromised cellular phagocytic activity as a result of impaired cholesterol synthesis and PrPC trafficking caused by U18666A. These data confirm the importance of cholesterol synthesis and the transport cycle as well as the PrPC trafficking in the process of PrPSc uptake and propagation in cells.
Characterization of the specificity of early prion accumulation
Prion accumulation in the macrophages was further investigated to elucidate whether it is specific to Raw cells and to PrPSc agent. To investigate specificity of this phenomenon to the Raw cells, similar experiment was conducted using J774 and BMMs cell cultures. Furthermore, to verify that PrPSc accumulation was specific to the infected brain homogenate and not due to other pathogens or antigens, normal brain homogenate, E. coli cell lysate and microspheres were added to cell cultures. After treatment of the cells with these agents, they were lysed and analyzed by Western blotting. Results revealed that PrPSc accumulation occurred only in the cells treated with PrPSc-brain homogenates (Fig. 8a). However, a transient weak accumulation of PrPC was noted in the cells treated with the normal brain (Fig. 8a).
46
mRNA expression of prion-encoding gene, PRNP, in macrophages
To characterize the potential relationship between prion protein gene (PRNP) activation and the observed accumulation of PrPSc in the macrophages, PRNP mRNA expression during exposure to PrPSc was investigated in the Raw cells by real-time PCR. A gradual up-regulation of PRNP gene, up to 2.5 times the normal level, was observed at 4 h of exposure (Fig. 8b). However, there was no direct correlation with changes in protein levels except in the case of treatment with normal brain homogenate where a transient weak accumulation of PrPC was observed.
The specificity of the observed increase of mRNA expression in the Raw cells to PrPSc exposure was further investigated by analysis of mRNA expression in Raw, J774 and bone marrow macrophages. These cells were treated with PrPSc-brain, normal brain, E. coli cell lysate and microspheres. A non-specific variable PRNP up-regulation in the Raw cells was observed with all of the treatments (Fig. 8b). Similarly, variable up-regulation of PRNP was also observed in the J774 cells and bone marrow primary macrophages with each of the treatment agents (data not shown). These results suggest that the mRNA expression of the PrP gene was not a specific response to PrPSc exposure but is potentially a normal response that occurs in immune cells exposed to various kinds of antigens.
47
DISCUSSION
In the first chapter, the author have observed that after 24 h of PrPSc exposure there have been three variable forms of cellular response in the following 5 days. These forms vary between accumulation and degradation of PrPSc in the cells. In some cell types, degradation was preceded by a transient PrPSc accumulation at around the 5th day of exposure. In this study, the author further investigated a similar characteristic phenomenon of a prominent and transient PrPSc accumulation in three macrophage cell types up to the 5th day of exposure. The source of the propagated PrPSc, the factors influencing the conversion of PrPC into PrPSc and its clearance in these cells were investigated. Data showed that the accumulated PrPSc was derived from the inoculated brain homogenate and was not a de novo product of the PrPC in cells.
In the present study, imitations of unfavorable conditions for cell viability or metabolism that may affect PrPSc clearance were fulfilled by incubating the cells at low temperature than appropriate. The results revealed that such unfavorable conditions might have affected the cells total activity leading to unsuccessful clearance of PrPSc. Moreover, higher PrPSc accumulation than previously observed was noticed. When the temperature around the cells is not optimum, cellular biophysiological mechanisms may be directed to enhance the PrPC-PrPSc conversion process resulting in propagation of PrPSc. This continuous PrPSc propagation combined with a low clearance rate may lead to its transfer to the neighboring cells, which could potentially be neural cells. The effect of temperature on the cellular degradation systems was proved by a significant decrease in proteasome activity in the infected and non-infected cells incubated at a lower temperature. This suggests that the accumulation of PrPSc is enhanced when degradation systems are impaired. PrPSc itself was found to impair the cellular 20S proteasome system even at optimum temperatures, confirming other studies [19, 42]. This may suggest that the risk of impairing PrPSc degradation could be higher at lower temperatures. Another factor that was found to relatively enhance PrPSc accumulation in the cells was the availability of excessive PrPC. The cells treated with the PrPSc-brain were later exposed to normal brain homogenate that helped maintaining PrPSc relatively longer time. The obtained results come in consensus with other studies that have shown the importance of PrPC in disease permissibility and progression [16, 70]. Here, the