名古屋市立大学大学院薬学研究科(〒4678693 名古屋 市瑞穂区田辺通 31) e-mail: umezawa@phar.nagoya-cu.ac.jp 本総説は,日本薬学会第 127 年会シンポジウム S11 で 発表したものを中心に記述したものである. ―Reviews―
アミノ酸,ペプチドの化学的性質を利用した生体高分子の選択的認識と検出
梅澤直樹,秋田昌二,鴨東美絵,樋口恒彦Selective Recognition and Detection of Biomacromolecules Utilizing Chemical Property
of Amino Acid or Peptide
Naoki UMEZAWA, Shoji AKITA, Mie KAMOTO, and Tsunehiko HIGUCHI Graduate School of Pharmaceutical Sciences, Nagoya City University, 31 Tanabe-dori,
Mizuho-ku, Nagoya 4678603, Japan (Received July 20, 2007)
Recently, the ‰uorometric detection of biomacromolecules has attracted much attention. In this paper, we report the development of two new techniques utilizing the chemical properties of amino acids or peptides: 1) a ‰uorescence as-say for serine/threonine kinase activity; and 2) ``turn-on'' ‰uorescent probes for protein labeling, which could be useful for bioimaging. To develop the novel kinase assay, we utilized the chemical reactivity of phosphorylated serine or threo-nine. Phosphorylated peptide on resin was successfully labeled ‰uorescentlyvia base-mediatedb-elimination, followed by Michael addition with novel coumarin derivatives. Protein kinase A and casein kinase I activities were detectable with our method. Also, this method was confirmed to be applicable for kinase inhibitor screening. For the development of the novel protein labeling technique, the selective interaction between ``His-tag (His6)'' and ``metal ion nitrilotriacetic
acid (NTA) complex'' was utilized. This interaction is useful for protein puriˆcation and immobilization. We designed ‰uorescent probes composed of a ‰uorophore and Ni2+or Co2+-NTA complex. These probes were found to be weakly
‰uorescent as expected. When His-tag peptide was added, these probes became brightly ‰uorescent. On the other hand, these probes remained non ‰uorescent with the addition of angiotensin I (H-Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu-OH). These probes will be powerful tools for the bioimaging of target proteins.
Key words―‰uorescence; protein kinase; protein labeling
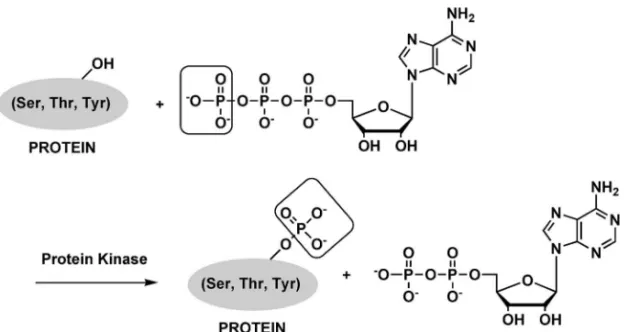
1. はじめに 近年,分析化学は目覚ましい発展を遂げている. その中で大きな役割を果たしてきたのは,様々な測 定機器ハードウェア―の進歩といえる.だが,よ り厳密な計測や時空間情報も加えた分析化学を進め るには,最新機器の性能を最大限に活用すると同時 にソフトウェアに当たる様々な新規手法の開発が不 可欠である.本誌上シンポジウムでは,ソフトウェ アとなる「アミノ酸,ペプチドの化学的性質を利用 した生体高分子の選択的認識と検出」について紹介 したい.具体的には,「新規セリン・トレオニンキ ナーゼ活性検出法」及び「ペプチドタグ導入タンパ ク質の発蛍光性蛍光試薬を用いた選択的標識法」の 2 つのトピックについて紹介する.われわれが行っ てきた研究を中心に,最近報告された新しい「ソフ トウェア」についても概観する.今回紹介する 2 つ のトピックの間には,新規検出試薬の開発が研究の 中で大きな比重を占めるという共通点はあるもの の,直接的な関係はほとんどない.各論的な紹介と なってしまうことをご容赦頂きたい. 2. アミノ酸の化学的性質を利用した生体高分子 の選択的認識と検出新規セリン・トレオニンキ ナーゼ活性検出法の開発 2-1. プロテインキナーゼ 細胞は,細胞外か らの情報を受け取ると,その内容を探知し,その情 報に応じた対応を行うシステムを有している.この ような細胞内でのシグナル伝達において最も重要な メカニズムは,タンパク質のリン酸化であると言わ れている.リン酸基はサイズも大きく,二価の陰イ オンであるため,リン酸化はタンパク質の構造を変 化させる有効な方法であると同時に,他のタンパク
Fig. 1. Protein Phosphorylation Catalyzed by a Protein Kinase 質によって認識される標識としても優れている.こ のリン酸化を行う酵素がプロテインキナーゼであ り,標的基質のセリン,トレオニン,チロシン残基 をリン酸化することで,細胞活動の幅広い制御を行 っている.キナーゼは,Fig. 1 に示すように,ATP を用いて基質タンパク質をリン酸化し,リン酸化タ ンパク質と ADP を生成する反応を触媒する.この 際,ATP の g 位のリン 酸基が基質に取 り込まれ る.ヒトには 500 種類を超えるキナーゼが存在する と言われており,キナーゼの関与する過程は複雑か つ多様性に富んでいる.これらプロテインキナーゼ は細胞内情報伝達系経路において,種々の酵素の活 性を制御し,細胞の分化,増殖,細胞死,代謝,運 動などを含む細胞の機能発現に重要な役割を果たし ているため,細胞内情報伝達におけるキナーゼの役 割の解明は,広範な興味を集めている.また,キ ナーゼの活性異常は,がん,炎症性疾患を始めとす る様々な疾病に関与することが明らかになってお り,細胞内情報伝達系におけるキナーゼの機能の解 明は,生命現象の理解はもとより,疾患の予防や治 療においても重要である.プロテインキナーゼの阻 害剤は既に医薬品として臨床で用いられているもの もあり,キナーゼは医薬品開発の重要なターゲット としても注目されている. キナーゼが関与する生命現象の解明や,キナーゼ 阻害剤のスクリーニングには,キナーゼ活性を検出 する信頼性の高い方法が不可欠である.そのため, キナーゼ活性の検出法は市販のものを含め,既に相 当数が報告されている.1)本稿では,代表的な検出 法に焦点を絞って,紹介していくこととする. 2-2. 一般的なプロテインキナーゼ活性検出法 キナーゼのアッセイには g 位のリン酸基を放射性 同位体(RI)32P で標識した ATP を用いる伝統的 な手法がよく用いられている.g 位のリン酸基はリ ン酸化反応により基質タンパク質に取り込まれるた め,32P 量を定量すればリン酸化反応の進行が定量 できる.この手法は,一般性が高いという大きな特 徴を有するが,放射性同位体を用いるため安全性に 問題がある上に,感度よく検出するためにはある程 度の露光時間が必要という問題点もある. 他に,よく用いられている方法として,抗リン酸 化アミノ酸抗体を用いる方法が挙げられる.キナー ゼによりリン酸化された基質ペプチドあるいはタン パク質に,例えば蛍光標識した抗リン酸化抗体を作 用させてリン酸化の有無を検出するというものであ る.この方法は RI を用いないため安全で,簡便な 方法であるが,抗体の特異性に問題があるとの指摘 がある.特に,リン酸化セリン,リン酸化トレオニ ンに対する抗体の特異性に問題があると言われてい る. 2-3. 新 し い プ ロ テ イ ン キ ナ ー ゼ 活 性 検 出 法 上述したキナーゼ活性検出法は,多くの重要な知 見を提供してきたが,それぞれの問題点も明らかと なってきている.そのため,最近新たな手法の報告
Scheme 1. Summary of Our Assay が相次いでいる.代表的なものを以下に挙げる. 2-3-1. リン酸基と選択的に相互作用する金属錯 体を用いる手法 Ojida らは,亜鉛の二核ジピコ リルアミン錯体がリン酸基と特異的に相互作用する ことを利用した,蛍光性の人工レセプター分子を報 告しており,SDS ポリアクリルアミドゲル上での リン酸化タンパク質の検出等に成功している.2,3)ま た , Kinoshita ら は リ ン 酸 基 選 択 的 ホ ス ト 分 子 Phos-Tag の開発に成功している.4)二核亜鉛錯体で あるこの分子にビオチンを付与した誘導体と,基質 ペプチドを金表面にアレイしたペプチドを用いて, キナーゼ活性の表面プラズモン共鳴による検出を報 告している.5)詳しくは,本誌上シンポジウムの木 下らの稿をご参照頂きたい. 2-3-2. 基質ペプチドに蛍光団を導入したキナー ゼ活性検出プローブ Higashi らは環境感受性蛍 光団(アクリロダン等)を基質ペプチドに導入し, リン酸化による環境の変化で蛍光強度が変化するプ ローブを開発し,生細胞内のキナーゼ活性の可視化 を行っている.6)Yeh らも同様のコンセプトで設計 したプローブを用いて,生細胞内のキナーゼ活性の 可視化に成功している.7)他に,リン酸基が金属配 位能を有することを利用し,金属配位性の蛍光団を 導入したプローブも開発されている.基質ペプチド のリン酸化後に,リン酸基と蛍光団がカルシウムや マグネシウムといった金属イオンと相互作用するこ と で , 大 き く 蛍 光 強 度 が 増 大 す る プ ロ ー ブ が , Chen ら8)や Shults ら9)によって報告されている. 2-3-3. MALDI-TOF MS を用いる手法 MAL-DI-TOF MS を用いる手法が Min らによって報告さ れている.チップ上に固定した基質ペプチドにキ ナーゼを作用させたのち,MALDI-TOF MS で分 子量を測定すると,リン酸化された基質ペプチド は,元の分子量より 80 大きい値(+PO3H)を示 すことを利用してキナーゼ活性を測定する方法であ る.この手法により種々のキナーゼ活性の検出に成 功している.10) ここで挙げたもの以外にも様々な検出法が開発さ れているが,多様なキナーゼに適用可能で,一般性 の高い検出法の開発は,いまだ困難かつチャレンジ ングな課題である. 2-4. 新規セリン・トレオニンキナーゼ活性検出 法の開発 筆者らは上述の検出法とは異なるアプ ローチでプロテインキナーゼアッセイ法の開発を進 めている.リン酸化アミノ酸に特異的な化学反応性 を利用し,リン酸化を受けた基質ペプチドを蛍光検 出する手法の開発に成功したので,紹介する.11) リン酸化アミノ酸のうち,リン酸化セリン・リン 酸化トレオニンは類似した化学反応性を有してい る.特に,塩基性条件下,リン酸基が b- 脱離する という反応性をいずれも有する.この反応性に着目 し,Scheme 1 に示すセリン・トレオニンキナーゼ アッセイ法を考案した. まず,固相に固定したペプチド基質にキナーゼを 作用させる.その後,塩基を作用させるとリン酸基 が b- 脱離し,炭素炭素二重結合が生成する.この 二重結合は良好なマイケル反応受容部位となるため, SH 基を有する蛍光色素を反応させることで,リン 酸化セリン・トレオニンを特異的に蛍光性に変換で きると考えた. 本検出法の作業仮説を検証するため,ペプチドを 固定する固相として,まずビーズ(レジン)を選択 した.これは,Fmoc 固相合成法を用いて,基質と
Fig. 2. a) Structure of Fluorescent Molecules Synthesized, b) Reaction of 1 with Methyl Vinyl Ketone, c) Emission Spectra (ex: 360 nm) of 1 and 3 (5 mM, 0.1% DMSO) in 100 mMGlycine/NaOH BuŠer (pH 9.4)
なるペプチドを直接ビーズ上に合成できるため基質 ペプチドの固定化が容易な上に,ビーズ上で酵素活 性を検出している例がいくつか報告されており,12) 検出原理の検討に適していると考えたためである. 検出に用いる蛍光色素には Fig. 2(a)に示す分子を デザイン,合成した.蛍光団として,強い蛍光を有 し,かつ比較的疎水性が低いクマリンを選択した. 疎水性が低いことで,固相との非特異的な相互作用 が小さくなることを期待した.このクマリンにマイ ケル付加反応に適した SH 基を導入した分子をデザ インした.まず,合成した化合物でマイケル付加反 応が進行するか,反応後に蛍光を有するかを検討し た . モ デ ル 基 質 と し て methyl vinyl ketone を 用 い,塩基性条件下反応したところ,蛍光色素 1, 2 ともに,予想通りマイケル付加反応が進行した.原 料 1 及び生成物 3 の蛍光強度を測定した結果,3 の 蛍光強度が 1 と比し 10 倍以上強くなることが確認 された(Fig. 2(b)).ここでは 1 の結果のみを示す が,2 を用いた場合にも同様の結果が得られた.こ のように反応後に蛍光強度が増大する性質は予想外 であったが,蛍光色素の非特異的吸着に起因する蛍 光を相対的に低減できるため,アッセイに適した性 質といえる. 次に,ビーズ上に合成したリン酸化ペプチドの検 出を試みた.広範な溶媒に適用可能で,酵素アッセイ にも適用例がある TentaGel 上に,Protein kinase A (PKA)の基質ペプチド(Kemptide:Ac-LRRASLG) とリン酸化されたペプチド(Ac-LRRApSLG;pS はリン酸化セリンを表す)を合成し,塩基処理後, 蛍光色素を反応させた.蛍光顕微鏡でビーズを観察 した結果,リン酸化ペプチドを有するビーズに選択 的な強い蛍光が観測され,本検出原理に基づいてリ ン酸化ペプチドを検出し得ることが示された.また 色素 1 と 2 を比較した場合,1 を用いた場合の方が
Fig. 3. a) Detection of PKA Activity, b) EŠect of Inhibitory Peptide (IP)
a): Beads were incubated with PKA (30 units ml-1) for 2 h, 4 h, or 12 h
at RT, then treated with base and 1. b): Beads were incubated with IP (0500 mM) and PKA (30 units ml-1) then treated with base and 1. All ‰uorescent
images (ex: 360 nm, em: >425 nm) were taken in 100 mMglycine/NaOH buŠer.
若干ではあるが良好な結果を与えたため,以後色素 1 を用いて検討を進めることとした.
続いて,キナーゼ活性の検出に着手した.Kem-ptide を固定した TentaGel を用いて PKA の活性検 出を試みたが,PKA を作用させたビーズ,作用さ せ な か っ た ビ ー ズ と も に 蛍 光 は ほ と ん ど な く , PKA によるリン酸化反応が進行していないことが 示唆された.そこでペプチドを固定するビーズを, より水溶液中での利用に適した PEGA800レジンに 変更し,PKA の活性検出を検討したが,TentaGel を用いた場合と同様に PKA を作用させたビーズか ら蛍光はみられなかった.筆者らは,ビーズ上の基 質ペプチドから蛍光が検出されない原因として, ビーズ内部に存在する基質が酵素に認識されず,リ ン酸化が進行していない可能性を考えた.つまり巨 大な分子である酵素はビーズ内に浸透できないた め,基質をリン酸化することができないのではない かと考えたのである.用いたレジンの性質を再検討 したところ,PEGA は TentaGel よりも水中でよく 膨潤するものの,PEGA800は分子量約 35 kDa まで の分子しかビーズ内に透過できない性質であること が明 らか となっ た.13)用いた PKA の分 子量は 40 kDa であり,ビーズ内に浸透できないことでリン 酸化が進行しなかったものと推察された. そこで,より水中で膨潤し,分子量 70 kDa まで の分子を透過可能な PEGA1900レジンを用いて, PKA の活性検出を試みた.その結果,PKA を作用 させたビーズに選択的な蛍光がみられた.酵素反応 時間を変化させたところ,酵素反応時間依存的な蛍 光強度の増大がみられ,ビーズ上で PKA 活性を検 出することができた(Fig. 3(a)).次に本アッセイ を用いて,キナーゼ阻害剤のスクリーニングができ るか検討した.酵素反応時に PKA の阻害剤となる ペプチド(Ac-GRTGRRNAI-NH2)を添加し,同 様の検出を行った.ビーズ上の蛍光は阻害剤濃度依 存的に減弱し,本検出法がキナーゼ阻害剤のスク リーニングに用い得ることが明らかとなった(Fig. 3(b)). また PKA 以外のキナーゼの活性も検出できるか 試みた.ビーズ上に Casein kinase I (CK I)の基質 ペ プ チ ド ( Ac-IGDDDDAYSDTETTEA-NH2) を 合成し,CK I の活性検出に適用したところ,CK I を作用させたビーズのみに蛍光がみられ,キナーゼ を作用させなかったビーズ,PKA を作用させた ビーズには蛍光はみられなかった.PKA,CK I と いう基質ペプチド配列が全く異なる(PKA 基質ペ プチドは正電荷を帯びているのに対して,CK I 基 質ペプチドは負電荷を帯びている)二種のキナーゼ のアッセイに成功し,本検出法が種々のキナーゼ活 性検出に適用できることが示された. 以上のようにわれわれはリン酸化アミノ酸の化学 反応性に基づき,ビーズ上でキナーゼの活性検出に 成功した.だが,ビーズを固相とするアッセイには いくつかの制限がある.最も大きな問題点は PKA, CK I の活性検出に成功した PEGA1900レジンであ っても,70 kDa 程度までのキナーゼにしか適用で きないことである.生体内には多様なキナーゼが存 在し,70 kDa 以上のキナーゼも数多く存在すると 考えられる.本アッセイ系は,酵素のサイズによっ てアッセイ対象が限られてしまう可能性が高く,問 題である.また,アッセイの効率にも制限がある. ビーズ 1 個 1 個を検出するビーズアッセイでは高い 効率が期待できない.現在,これらの問題点の克服 を目指し,検出原理をペプチドチップへ適用した研
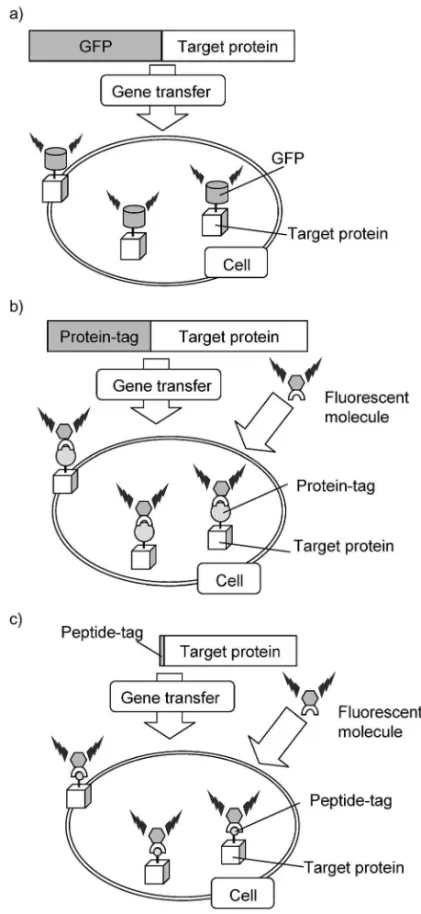
Fig. 4. Fluorescent Labeling by a) GFP, b) Protein Tag-‰uorescent Molecule System, c) Peptide Tag-Tag-‰uorescent Molecule System 究を進めている. 3 ペプチドの化学的性質を利用した生体高分子 の選択的認識と検出発蛍光性蛍光試薬を用いたペ プチドタグ導入タンパク質の選択的標識 3-1. タンパク質の蛍光イメージング タンパ ク質は,核酸や糖鎖と並ぶ重要な生体高分子であ り,生物の中で起こる化学反応の大半を担ってい る.多くのタンパク質の機能がいまだ明らかとなっ ていないが,その解明は,生命現象の理解,疾病の 発症機構解明,創薬ターゲットの発見等につながる と考えられる.現在,タンパク質機能の網羅的解析 を目指したプロテオーム研究を中心に,多くの重要 な知見が得られてきているが,この方法では,個々 のタンパク質機能に関する限定された基本情報しか 得ることができない.そのため,さらなる機能解析 には異なる戦略が必要と考えられる.その重要な戦 略 の 1 つ に , 蛍 光 イ メ ー ジ ン グ 法 が 挙 げ ら れ る.14,15) タンパク質の蛍光イメージングは,生きた細胞内 のタンパク質を蛍光性物質でラベル化し,直接観察 することで,その局在や挙動を明らかにしようとす る手法であり,タンパク質機能を研究する強力な手 段となる.しかし,細胞内に存在する数多くのタン パク質の中から「観察したいタンパク質だけを蛍光 ラベル化する」ことは,非常に困難である.さら に,目的タンパク質を選択的にラベル化することが 望まれる一方で,様々なタンパク質に適用できる一 般性も必要であり,そのハードルは高い. 3-2. タンパク質イメージング技術開発の現状 3-2-1. 蛍光性タンパク質を用いた蛍光ラベル化 法 現在,タンパク質の蛍光イメージングにおい て最もよく用いられている手法は,Green Fluores-cent Protein (GFP)を中心とした蛍光タンパク質を 利用した手法16,17)である(Fig. 4(a)).この手法は, 標的タンパク質の遺伝子に蛍光タンパク質の遺伝子 を付与することで,標的タンパク質のみを蛍光ラベ ル化するというものである.ラベル化が簡便かつ, 特異性が高いという大きな利点を有する.また,最 近様々な変異体が開発され,蛍光波長の異なる 2 種 類の蛍光性タンパク質間の FRET(蛍光共鳴エネル ギー移動)により,タンパク質の構造変化を捉える といった応用例も報告されている. この方法は非常によく用いられているが,蛍光性 タンパク質の分子サイズが約 240 アミノ酸と大きい ために,蛍光ラベル化することで,標的タンパク質 の構造や機能に影響を及ぼすという問題点が指摘さ れている.18)また,蛍光波長の異なる様々な変異タ ンパク質が開発されてはいるものの,その蛍光波長 のバリエーションは十分なものとはいえず,マルチ カラーラベリングは難しいのが現状である.
3-2-2. タンパク質タグ蛍光性有機小分子を用い た蛍光ラベル化法 上述のように,蛍光性タンパ ク質は,分子サイズが大き過ぎるという問題点があ った.そこで,タグ部分をもう少し小さいタンパク 質に変換し,それと相互作用する蛍光性有機小分子 を用いてラベル化する手法がいくつか報告されてい る(Fig. 4(b)).19)この手法は,タンパク質タグと 蛍光性分子が共有結合するものと,共有結合ではな い相互作用するものとに分類できる. 前者は,酵素反応により生成する新たな共有結合 を利用したものである.Keppler らは,ヒト O6- ア ルキルグアニントランスフェラーゼ(hAGT)によ る自己アルキル化反応を用いたラベル化法20)を報告 している.具体的には,蛍光団を有する O6- アルキ ルグアニン誘導体を用いて,hAGT タグ(207 アミ ノ酸)導入タンパク質を修飾するというものであ る.また,ホスホパンテチントランスフェラーゼ ( PPTase ) に よ る ア シ ル キ ャ リ ア タ ン パ ク 質 (ACP)への補酵素 A の転移反応を利用した例21)で は,タンパク質タグのサイズも ACP(77 アミノ酸) まで小さくすることができている. 後者は,タンパク質と基質との特異的な相互作用 を利用したものである.Miller らは,ジヒドロ葉酸 還元酵素(DHFR)とメトトレキサートの強い親和 性を利用して,蛍光団を有するメトトレキサートを 用いて,DHFR(187 アミノ酸)タグ導入タンパク 質 を 蛍 光 ラ ベ ル 化22)し て い る . ま た , Marks ら は , 免 疫 抑 制 剤 FK506 と そ の 結 合 タ ン パ ク 質 (FKBP12)の相互作用を利用して,蛍光団を有す る FK506 誘導体を用いた FKBP12 タグ(98 アミノ 酸)導入タンパク質のラベル化23)を報告している. いずれも,タンパク質と反応基質ないしはリガン ドとの特異的な相互作用を巧みに利用したものであ る.しかし,蛍光性タンパク質よりタグサイズが小 さくなっているとはいえ,タンパク質タグの大きさ は 100200 アミノ酸と大きく,標的タンパク質の機 能への影響が懸念される.また,内在性の相互作用 を利用しているので,細胞膜上のタンパク質には適 用できるが,細胞内タンパク質をラベル化すること は難しいという問題点もある. 3-2-3. ペプチドタグ蛍光性有機小分子を用いた 蛍光ラベル化法 最近注目を集めている手法は, 短いペプチドタグを標的タンパク質に導入し,ペプ チドタグと特異的に結合する蛍光性小分子でラベル 化する手法である(Fig. 4(c)).本手法は短いペプ チドタグを用いているので,標的タンパク質に与え る影響は非常に小さいと考えられる.また,蛍光性 有機小分子については,種々の蛍光団を選択し応用 可能であるという利点を有する. この先駆的な例として,テトラシステインモチー フ(-Cys-Cys-X-X-Cys-Cys-)と選択的に結合する
FlAsH,24)ReAsH25)が挙げられる.FlAsH は,フ
ルオレセインにヒ素を導入した化合物で,ペプチド タグ配列中のシステイン残基に結合して標的タンパ ク質を蛍光ラベル化する.FlAsH は蛍光ラベル化 の際に,蛍光強度が増大するという優れた特徴を有 するが,内在性チオール基との非特異的な相互作用 によるバックグラウンドが無視できない.このた め,高濃度のエタンジチオールで前処理する必要が ある. 他にも,蛍光性小分子と短いペプチド配列との新 規な相互作用を見い出したという例が数例報告され ている.Rozinov らは,ファージディスプレイによ り蛍光色素 Texas red と選択的に相互作用するペプ チド配列(13 アミノ酸)を見い出すことに成功し ている.26)また,Ojida らは,亜鉛の二核ジピコリ ルアミン錯体とオリゴアスパラギン酸配列との親和 性を利用したラベル化法を報告している.27) 以上のように,小分子と短いペプチド配列との相 互作用が最近報告されているが,最もよく知られて いる相互作用は,ヒスタグ((His)6)と金属 -NTA 錯体との相互作用であろう.28,29)この相互作用は, アフィニティークロマトグラフィーによるタンパク 質精製に広く用いられており,ヒスタグ導入に伴う 標的タンパク質の機能変化が少ないことが知られて いる.この相互作用に着目したタンパク質蛍光ラベ ル化法も,いくつか報告例がある.3032)しかし,い ずれも NTA 錯体と蛍光団を単に結合させたもの で,タグ導入タンパク質と相互作用はするものの, タグ配列と結合後に蛍光強度の増大がみられなかっ た.すなわち,ラベル化後に洗浄や精製といった煩 雑な操作が必要とされ,洗浄操作が可能な膜タンパ ク質には応用できるものの,細胞内タンパク質のラ ベル化は困難であった. 筆者らが知る限り,唯一蛍光変化がみられるもの として環境応答性蛍光団である Dansyl 基を導入し
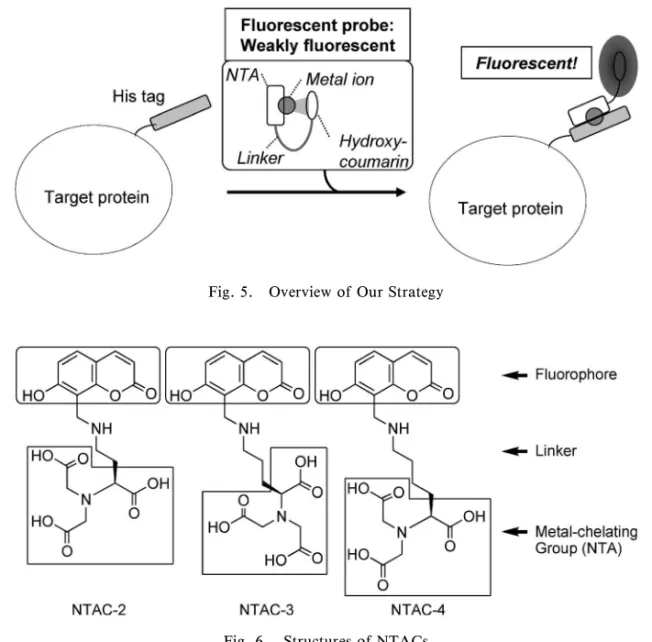
Fig. 5. Overview of Our Strategy
Fig. 6. Structures of NTACs
た Soh らの Ni-NTA プローブがある.33)これは,ヒ スタグ配列にさらに疎水性アミノ酸残基((Trp)3) を導入することで,ラベル化後に蛍光波長がシフト するようにデザインされており,洗練されたプロー ブである. 3-2-4. 発蛍光性蛍光試薬を用いたペプチドタグ 導入タンパク質の選択的標識 筆者らは,新たな 戦略として,タグ配列認識後に蛍光強度が大幅に増 大する性質を付与した蛍光プローブの開発を目指し 研究を進めている.このような性質を有する分子 は,ラベル化反応後の未反応プローブの洗浄操作が 必要ないため,細胞内タンパク質のイメージングに も応用できると考えられる. 筆者らの提案する蛍光プローブの蛍光ラベル化機 構の概略を Fig. 5 に示す.まず標的タンパク質が 存在しないとき,蛍光団は金属イオンに配位し,蛍 光は消光している.そこにヒスタグ導入タンパク質 を添加すると,蛍光団よりも強く金属イオンに配位 するヒスタグ配列が存在するため,金属錯体部位が 標的タンパク質と結合する.それに伴い,蛍光団は 遊離し蛍光が回復すると期待した. 上述の作業仮説に基づき,新規蛍光色素を設計, 合成した(Fig. 6).蛍光団としてヒドロキシクマ リンを,金属配位部位として NTA を選択した.両 者を異なる長さのリンカーで結んだ蛍光色素をデザ イン,合成し,リンカーの長さから NTAC-2, 3, 4 と名付けた.蛍光団として選択したヒドロキシクマ リンは,金属に配位することで蛍光強度が減弱する ことが知られているため選択した. 合成した NTAC 類に遷移金属を中心に,約 20 種 類の金属イオンを添加して蛍光強度変化を検討し た.その結果いずれの蛍光色素についても,ある種 の金属イオンの添加により蛍光強度の減弱がみられ た.なかでも Co2+,Ni2+の添加では,濃度依存的 な大幅な蛍光強度の減弱がみられた.Figure 7 に NTAC-4 の 結 果 を 示 す が , 他 の NTAC-2 及 び NTAC-3 でも同様の結果が得られた.続いて,金 属と蛍光色素がどのような錯体を形成しているか確
Fig. 7. Relative Fluorescence Intensity of NTAC4 (5mM) in the Presence of Various Metal Ions (Mn2+, Co2+, Ni2+,
Cu2+, Ca2+, Mg2+) in 50 mMTris-buŠer (pH 7.4)
Excitation: 365 nm, Emission: 455 nm.
Fig. 8. Job's Plot of the Fluorescence Changes upon Com-plexation of NTAC4 with Ni2+ or Co2+ in 50 mM Tris
BuŠer (pH 7.4) at 25°C.
[NTAC4]+[metal ion]=10 mM. Excitation: 365 nm, emission: 455
nm.
Fig. 9. Fluorescence Intensity of NTAC4Co2+ (5 mM) in
the Presence of a) Peptide 1 (H-(His)6-Tyr-NH2), b)
An-giotensin I (H-Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu-OH) in 50 mMTris-buŠer (pH 7.4) at 25°C Excitation: 365 nm, emission: 455 nm. 認するため,Job's plot を行ったところ,すべての NTAC 類は金属イオンと 1 対 1 の錯体を形成して いることが示唆された.Figure 8 に NTAC-4 の結 果を示す. Co2+-NTA, Ni2+-NTA 錯体は,一般にヒスタグ と強く相互作用することが知られている.そこで, ヒスタグ配列を有するモデルペプチド(H-(His)6 -Tyr-NH2)を添加して,蛍光強度に変化がみられる か検討した.モデルペプチドには,ヒスタグ配列に 加えてチロシンを導入しているが,正確なペプチド 濃度を吸光度により算出するためである.
Co2+-NTAC 錯体,Ni2+-NTAC 錯体にモデルペ
プチドを添加したところ,いずれの錯体も大幅な蛍 光回復を示すことが明らかとなった.NTAC-4 の
結果を Fig. 9(a)に示す.Ni2+-NTAC 錯体でも蛍光
強度の回復はみられたが,Co2+-NTAC 錯体と比 し,蛍光強度変化に時間がかかることが明らかとな り,Co2+-NTAC 錯体の方が優れたプローブである ことが示唆された.また,蛍光変化から,解離定数 を求めたところ約 10-610-7Mと強い相互作用をす ることが明らかとなった.一方,ネガティブコント ロールとして,連続したヒスチジン配列を持たない ペ プ チ ド ( Angiotensin I: H-Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu-OH)を添加したところ,蛍
光強度の回復は全くみられないという結果が得られ た(Fig. 9(b)). 今回,ヒスタグ配列を認識し発蛍光するような蛍 光プローブの合成を行い,その蛍光特性について検 討してきた.ペプチドレベルではあるが,当初予測 した通り,ペプチド配列を選択的に認識し大幅な蛍 光増大を示すという結果が得られた.現在,タグタ ンパク質を用いて蛍光変化がみられるか鋭意検討を 進めている. 4. おわりに アミノ酸やペプチドの持つ化学反応性に着目した 新たな検出法の開発について,筆者らの研究を中心 に述べてきた.いずれの研究も,端緒を開いたばか りといったところである.幸いなことに,今回開発 した蛍光色素はいずれも,蛍光団の変換が容易とい う特徴を持つ.研究を進めるうちに明らかになって きた問題点を解決し,より成熟した方法論を確立し たいと考えている. REFERENCES
1) von Ahsen O., Bomer U.,ChemBioChem, 6, 481490 (2005).
2) Ojida A., Mito-oka Y., Inoue M., Hamachi I., J. Am. Chem. Soc., 124, 62566258 (2002). 3) Ojida A., Mito-oka Y., Sada K., Hamachi I.,
J. Am. Chem. Soc., 126, 24542463 (2004). 4) Kinoshita E., Takahashi M., Takeda H., Shiro
M., Koike T., Dalton Trans., 11891193 (2004).
5) Inamori K., Kyo M., Nishiya Y., Inoue Y., Sonoda T., Kinoshita E., Koike T., Katayama Y.,Anal. Chem., 77, 39793985 (2005). 6) Higashi H., Sato K., Ohtake A., Omori A.,
Yoshida S., Kudo Y.,FEBS Lett., 414, 5560 (1997).
7) Yeh R.-H., Yan X., Cammer M., Bresnick A. R., Lawrence D. S., J. Biol. Chem., 277, 1152711532 (2002).
8) Chen C.-A., Yeh R.-H., Lawrence D. S., J. Am. Chem. Soc., 124, 38403841 (2002). 9) Shults M. D., Imperiali B., J. Am. Chem.
Soc., 125, 1424814249 (2003).
10) Min D.-H., Su J., Mrksich M.,Angew. Chem. Int. Ed., 43, 59735977 (2004).
11) Akita S., Umezawa N., Higuchi T., Org. Lett., 7, 55655568 (2005).
12) Wu J., Ma Q. N., Lam K. S., Biochemistry, 33, 1482514833 (1994).
13) Basso A., Braiuca P., Ebert C., Gardossi L., Linda P.,J. Chem. Technol. Biotechnol., 81, 16261640 (2006).
14) Giepmans B. N. G., Adams S. R., Ellismann M. H., Tsien R. Y., Science, 312, 217224 (2006).
15) Miyawaki A., Sawano A., Kogure T., Nat. Rev. Mol. Cell Biol., 5, S1S7 (2003).
16) Tsien R. Y.,Annu. Rev. Biochem., 67, 509 544 (1998).
17) Schwartz J. L., Patterson G. H.,Science, 300, 8791 (2003).
18) Zhang J., Campbell R. E., Ting A. Y., Tsien R. Y., Nat. Rev. Mol. Cell Biol., 3, 906918 (2002).
19) Marks K. M., Nolan G. P.,Nat. Methods, 3, 591596 (2006).
20) Keppler A., Gendreizig S., Gronemeyer T., Pick H., Johnsson K., Nat. Biotechnol., 21, 8689 (2003).
21) Vivelo-Pol L., George N., Krumm H., Joh-nsson K., JohJoh-nsson N., J. Am. Chem. Soc., 127, 1277012771 (2005).
22) Miller L. W., Cai Y., Sheetz M. P., Cornish V. W.,Nat. Methods, 2, 255257 (2005). 23) Marks K. M., Braun P. D., Nolan G. P.,
Proc. Natl. Acad. Sic. U. S. A., 101, 9982 9987 (2004).
24) Gri‹n B. A., Adams S. R., Tsien R. Y., Science, 281, 269272 (1998).
25) Adams S. R., Campbell R. E., Gross L. A., Martin B. R., Walkup G. K., Yao Y., Llopis J., Tsien R. Y.,J. Am. Chem. Soc., 124, 6063 6076 (2002).
26) Rozinov M. N., Nolan G. P.,Chem. Biol., 5, 713728 (1998).
27) Ojida A., Honda K., Shinmi D., Kiyonaka S., Mori Y., Hamachi I., J. Am. Chem. Soc., 128, 1045210459 (2006).
28) Hochuli E., Dobeli H., Schacher A.,J. Chro-matogr., 411, 177184 (1987).
29) Ueda E. K. M., Gout P. W., Morganti L.,J. Chromatogr., A, 988, 123 (2003).
30) Guignet E. G., Hovius R., Vogel H., Nat. Biotechnol., 22, 440444 (2004).
31) Goldsmith C. R., Jaworski J., Sheng M., Lip-pard S. J.,J. Am. Chem. Soc., 128, 418419
(2006).
32) Kapanidis A. N., Ebright Y. W., Ebright R. H., J. Am. Chem. Soc., 123, 1212312125
(2001).
33) Soh N., Seto D., Nakano K., Imato T.,Mol. BioSyst., 2, 128131 (2006).