識別番号 P16
研究課題
金属酵素モデルとしての二価金属錯体の合成と構造及び酵素活性の測定
研究代表者 猪俣芳栄(理工学部・化学科)共同研究者 F.S.Howell(理工学部・化学科)
木山雅博(応用化学専攻・大学院生)・小林愛実(応用化学専攻・大学院生)
Summary A small amount of indispensable metallic elements exist in vivo, and some part is related to the reaction in vivo as the metal-containing enzyme. The copper ion is included in the ascorbic acid oxidaze and the zinc ion is included in the protease and phosphate hydrolase.
In this research, copper(Ⅱ) complexes with 4-amino-3-hydroxy butyric acid and L-citramalic acid (complexes 1 and 2) were synthesized as the ascorbic acid oxidase model, and zinc(Ⅱ) complexes with N-hydroxysuccinimide (complexes 3 and 4) were synthesized as a model of the phosphate hydrolase. The results of X-ray analysis show that the complex (1) has a trinuclear structure, the complex (2) has polynuclear structure, and the complexes (3, 4) have a mononuclear structure. In addition, the functions as the enzyme of these complexes were examined by a spectroscopy method, and the relations between the structures and the functions are considered.
Ⅰ 本研究の背景及び目的 背景 :生体内には微量で正常な機能を発揮する微量必須金属元素(V,Cr,Mn,Fe,Co,Ni,Zn,Mo) が存在し、それらは生命の基本的機能に直接関与する役割や、基本的機能を補助する役割を もつている。当研究室では、この微量元素の働きに関心をもち、アミノ酸錯体及び類似化合 物錯体の合成と性質および構造について研究を続けてきた。 酵素は生体内の穏やかな環境と中性に近い状態で、特定の化合物(基質)とだけ反応(基質特 異性)し、化学変化を素早く進行させる(活性発現)。これら酵素の約3分の1は、その活性発現 のために金属を必要とし、酵素の活性部位において金属イオンが働いている物質を一般的に金属 酵素とよび、Fe,Zn,Cu は生体内で様々な形で利用されている。 金属酵素の反応活性中心付近は、金属イオンとそれに結合している化合物(配位子:アミ ノ酸や水など)から成り、高い頻度でヒスチジンが存在しており、その活性に大きな役割をも っている。 図1 アスコルビン酸酸化酵素の活性中心の模式図 その中でもアスコルビン酸酸化酵素(A O)に は 、図 1 に 示 す 様 に 3 種 類 の 銅 イ オ ン周りの状態が存在する。 また、亜鉛含有酵素はリン酸エステルや ペプチドの加水分解に重要な働きをして いる。酵素活性を有する化合物は、2つ の金属原子を含む二核構造をとっている 場合が多いが、近年、単核構造で酵素活 性を有する化合物の合成例が報告されて いる(図2)。
目的:本研究ではAOと同様の作用をもつ銅錯体および、 リン酸エステル加水分解反応を促進する亜鉛錯体を合 成し、それら錯体の構造の解析を明らかにするととも に、錯体構造と酵素活性の有無、酵素反応の速度との関 係を明らかにすることを目的として研究を行った。 Zn N OH2 N N NH NH HN His His His Zn N O O HO Ser NH Asp O O Asp His Zn に 配 位 し た H2O や OH 種が反応機構に関与 Ⅱ 実験 1)錯体合成 ① 4-アミノ-3-ヒドロキシ酪酸(H2ahb)銅錯体(1): 水溶液中でH2ahbとCu(OH)2・2H2Oとを、物質量 比1:1で加熱混合し、不溶物をろ別後、濃縮して 図 2 リン酸エステルの加水分解 銅錯体(1)を得た。 ② L-(+)-シトラマル酸(H2cma)銅錯体(2):水溶液中で H2cmaとCu(CH3COO)2を用いて、錯体(1)と同じ方法で錯体濃縮溶液を作り、エタノール雰囲 気下に静置して銅錯体(2)を得た。 ③ N-ヒドロキシスクシンイミド(Hnhs)亜鉛錯体(3):水溶液中でHnhsとZn(CH3COO)2・2H2O を用いて、錯体(1)と同じ方法で合成して、亜鉛錯体(3)を得た。 ④ N-ヒドロキシスクシンイミド(Hnhs)-イミダゾール(im) 亜鉛錯体(4):水溶液中で錯体(3) とイミダゾールを物質量比1:1で加熱混合し、濃縮して亜鉛-イミダゾール錯体(4)を得た。
C O O H
C O O H
O H
C
H
3COOH
CH
2NH
2H
O
H
H2cma Hnhs H2ahb 図 3 錯体合成に用いた化合物 2)諸測定 これらの錯体について元素分析、IR を測定し、単結晶 X 線構造解析を行った。 3)酵素活性の測定 3-1. アスコルビン酸酸化酵素(AO):アスコルビン 酸(H2AsA)はpH 5.60 付近において 266 nmに吸収 が存在するが、酸化されると L-デヒドロアスコル ビン酸(AsA)が生成し、266 nm の吸収が消失する。(図 4) OH OH H2AsAと銅錯体との反応についてUVスペクトル を用いて、H2AsA溶液に銅錯体溶液を加え、吸光度の 図 4 アスコルビン酸酸化反応 時間変化を追い、H2AsA酸化の影響を調べた。(図 4) 3-2. リン酸エステル加水分解酵素: リン酸エステル(基質)として、トリス(4-ニトロ フェニル)リン酸エステル(TNPP)を用いた。 TNPPは加水分解すると、 p-ニトロフェノールを 生じ、404 nmに吸収が現れる。(図 5) TNPP溶液に亜鉛錯体溶液を加え、吸光度の時間 変化を溶液のpH、亜鉛錯体濃度、基質濃度な どの条件を変化させながら測定した。得られた O OH O H O O H O O O O O H -2H H2AsA O2 + 4H+ + 4e- → 2H2O AsA 分解物 p-nitrophenolate ion (λmax= 402 nm) O O2N 分解物 p-nitrophenolate ion (λmax= 402 nm) O O2N O O2N O O2N O O2N P O O O2N O O2N O O2N P O TNPP 図 5 リン酸エステル加水分解反応 加水分解結果から、速度定数を算出し、活性評価を行った。 Ⅲ 結果と考察 1)組成 元素分析の結果より錯体の組成は、表1の通りであった。 表1 元素分析及び熱重量分析結果 (計算値) 錯体 C / % H / % N / % H2O / % (1) Cu3(DL-Hahb)3(H2O)8 21.09 (21.01) 5.43 (5.44) 6.15 (6.12) 20.9 (21.00) (2) Cu(L-cma)(H2O)3 22.98 (22.70) 4.39 (4.59) - (0) 18.9 (20.48) (3) Zn(nhs)2(H2O)2 29.10 (29.15) 3.61 (3.67) 8.36 (8.50) 10.7 (10.92) (4) Zn(nhs)2(imi)2 38.61 (39.13) 3.56 (3.75) 19.39 (19.56) - (0) 2)単結晶 X 線構造解析 4種の錯体は図 6~9 に示す構造をとっていた。錯体(1)はカルボキシル基 O 原子、ヒドロキ シル基O原子、アミノ基N原子が隣接 Cu 原子に配位して三核錯体を構成していた。Hahb は 不斉炭素を有するため、三核錯体は S-S-R 体と R-R-S 体から成り立っていた。 錯体(2)はカルボキシル基 O 原子と水分子の O 原子が Cu 原子に配位した多核錯体であった。 錯体(3)はヒドロキシル基 O 原子とカルボニル基 O 原子及び水分子の O 原子が Zn 原子に配位 をした八面体錯体であった。錯体(4)はヒドロキシル基 O 原子とイミダゾール(im)の N 原子が Zn 原子に配位した四面体錯体であった。 図 7 錯体(2) Cu(L-cma)(H2O)3 の構造 図 6 錯体(1) Cu3(DL-Hahb)3の構造 錯体(2)Cu(cma)(H2O)2 3)酵素活性 3-1.アスコルビン酸(H2AsA)酸化 1) pH 変化よる吸収極大波長のシフト 基質としてH2AsAを、酵素として錯体(2)を用い て、溶液の pH を変化させてスペクトル測定を行 った。pH を 8.00 から酸性側へと変化させていく にともない、H2AsAに由来する吸収極大波長が 266 nm から 245 nm へとシフトした。(図 10) 2) 酸化反応促進効果の pH 依存性 錯体(1)と(2)ついて、上記 1)の結果から決定した 各pHのH2AsAによる吸収極大波長における時 R体 S体 図 8 錯体(3)Zn(nhs)2(H2O)2の構造 S体 図 9 錯体(4)Zn(nhs)2(imi)2の構造 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 200 220 240 260 280 300 320 340 波長 / nm 吸光度 pH3.50_acetate pH4.50_acetate pH5.60_MES pH6.00_MES pH6.50_MES pH7.00_HEPES pH7.50_HEPES pH8.00_HEPES 図3.1.1.1 スペクトル変化測定([Cu(L-cma)(H2O)]・2H2O) 図 10 錯体(2)のスペクトル変化
0.00 0.50 1.00 1.50 2.00 2.50 3.00 3.00 5.00 7.00 9.00 pH v0 / 10 -3 s -1 間経過にともなう吸光度変化を測定した。 錯体(1)■ 錯体(2)◆ その結果から求めた反応初速度を、pH に対し てプロットした(図 11)。両錯体について活性 が最大になる pH はおよそ 6.5 付近であること がわかった。酵素反応における至適 pH は、反応 の活性種の種類に依存するので、これら 2 つの錯 体を用いた場合の酸化反応における活性種は互い 図 11 pH と初速度の関係 に似ていると考えられる。 3) 酸化反応促進効果の温度依存性 錯体(1)と(2)を用いて、266 nm における 3600 秒間の吸光度変化を、測定温度を 25℃から 10℃ 毎に、65℃まで変化させて測定を行った。錯体(1)と(2) では、互いに異なる挙動を示した。ア スコルビン酸酸化酵素(AO)自身を用いての温度依存性の測定では、活性は 30℃付近で最大と なり、その後ほぼ一定の活性を保つことがわかっている。従って、温度依存性の挙動はこれ ら、モデル錯体を用いた場合と AO 自身とでは異なると考えられる。 4) 酸化反応促進効果の酸素濃度依存性 銅(Ⅱ)錯体が酸化反応の間にMichaelis-Menten挙動を示すと、反応式は下式であらわされ、ここで 生成したO2 -は、O 2とH2O2へと不均化される。 0.00 0.50 1.00 1.50 2.00 0.00 2.00 4.00 6.00 8.00 錯体濃度 / 10-6 M v 酵素濃度(錯体濃度)プロットの勾配から算出できる。 0 / 10 -3 s -1
H2AsA + Cu(Ⅱ) Cu(Ⅱ)‐H2AsA Cu(Ⅰ) + AsA
Cu(Ⅰ) + O2 Cu(Ⅱ) + O2 -k1 k-1 k2 k3 この機構に対する速度式は、Vmax / v = 1 + Km / [S] + (k2 / k3)×1 / [O2] であらわされる。 ここで、Vmaxは最大速度、vは基質濃度[S]における反応速度、KmはMichaelis定数である。 本測定では、反応溶液中の酸素濃度を変化させ、酸素濃度が初速度に影響を及ぼさないことを確 認し、酸素濃度の寄与がないとして取り扱った。 5) 酸化反応促進効果の錯体濃度依存性 酵素反応は次の経路で進行する。E(酵素) + S(基質) → ES(酵素-基質複合体) → P(生成物) 反応速度(v)は、酵素と基質の濃度に依存するため、 v = k[S][E] (1) であらわされる。 ここで、基質が酵素に対して大過剰に存在してい 錯体(1)■ 錯体(2)◆ るとする。すると、基質濃度は酵素濃度に対して その変化量が無視でき、式(1)は、v = kobs[E] (2) と表現でき、反応速度は酵素濃度のみに依存する。 kobs(擬一次速度定数)は、初速度法により初速度- 初速度と錯体濃度の関係は、ほぼ直線となった (図 12)。 このグラフの勾配より、擬一次速度定数(kobs)は、 Cu3(DL-Hahb)3(H2O)8について 1.85×10 2 M-1s-1と、 図 12 初速度と錯体濃度との関係 Cu(L-cma)(H2O)3について 2.44×10 2 M-1s-1と決定 された。
0.00 0.50 1.00 1.50 2.00 2.50 3.00 3.50 4.00 0.00 5.00 10.00 15.00 [S] / 10-5 M 6) 酸化反応促進効果の基質濃度依存性 酵素‐酵素基質複合体を形成しているとすれば、 錯体(1)■ 錯体(2)◆ Michaelis-Menten の式(3) に従い、 v0 / 10 -1 -3 s 反応速度‐基質濃度プロットは直角双曲線になる。
]
S
[
]
E
][
S
[
m 0 cat+
=
K
k
v
(3) 0 cat 0 cat m ] E [ 1 ] E ][ S [ 1 k k K v= + (4) KmはMichaelis定数であり、結合定数の逆数である。 従って、Kmの値が小さいほうが酵素‐基質複合体を 形成して、酵素反応の機構で進行しているといえる。 Kmは、Lineweaver-Burk両逆数プロット((初速度)-1‐ 図 13 初速度と基質濃度との関係 (基質濃度)-1) (4)式より求められる。 0.00 2.00 4.00 6.00 8.00 10.00 0.00 1.00 2.00 3.00 [S]-1 / 105 M-1 v0 / 10 3 s 図 13、14 に初速度‐基質濃度プロットと Lineweaver-Burk 両逆数を示す。 図 13 より、この酸化反応は、基質‐錯体複合体を 形成して進行する Michaelis-Menten 挙動を示すとい うことがわかった。 3-2 リン酸エステル加水分解反応 反応は基質の初濃度を大過剰にし、擬一次反応と して取り扱い、擬一次速度定数kobs(s-1)を次のように 定義した。ln[(A∞-At) / (A∞-A0)] = -kobs t A∞は反応終了時 図 14 Lineweaver- Burk 両逆数
プロット の吸光度、Atはt sec時の吸光度、A0は 0 sec時の吸
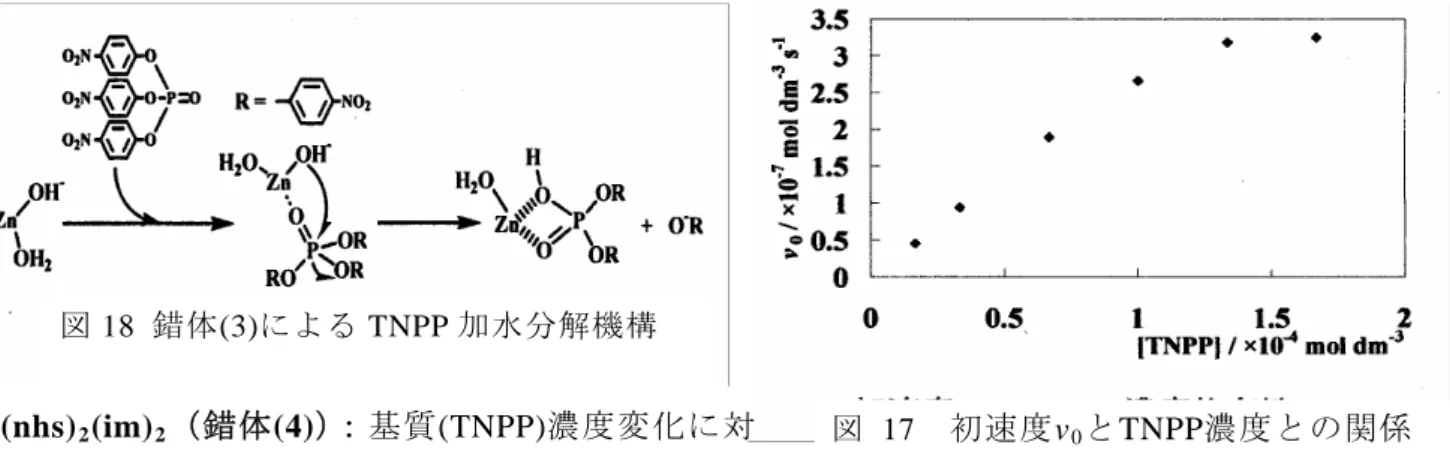
光度、kobsは擬一次速度定数、tは測定時間(sec)である。 Zn(nhs)2(H2O)2(錯体(3)):錯体溶液のpHを 6.50~ 11.00 に変化させて測定したところ、およそ pH 8 あた りでkobsが極大をもつことがわかった(図 15)。 また、図 16 に示すように pH 7.00、8.00、9.00 での錯体 濃度変化とkobsとの間には比例関係が成りたった。 直線の傾きを見かけ上の二次速度定数kapp とすると pH 7.00、8.00、9.00 でそれぞれ 1.388×10-2 M-1 s-1、 図1 kobs(錯体1)のpH依存性 pH 0.0 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 6.00 7.00 8.00 9.00 10.00 11.00 kob s / × 1 0 -4 s -1 6 7 8 9 10 11 pH 図 15 錯体(3)のkobs とpH依存性 1.946×10-2 M-1 s-1、9.803×10-3 M-1 s-1 になり、各pH の濃度によって反応性が違うことがわかった。 pH 8.00 でZn(nhs)2(H2O)2溶液に対しTNPPの濃 度を変化させて初速度を求めたところ、図 17 の ように Michaelis-Menten 挙動を示した。 Zn(nhs)2(H2O)2に配位するH2OのpKaを測定し、 化学種の分布曲線を作成した。 その結果、活性が最大なpH8 付近では[Zn(nhs)2 -(OH)(H2O)] -が主な化学種として存在し、これを 活性種と考え、反応機構を推定した(図 18)。 錯体(3)/×10-4 mol dm-3
図 18 錯体(3)による TNPP 加水分解機構 図 17 初速度v0とTNPP濃度との関係 Zn(nhs)2(im)2(錯体(4)):基質(TNPP)濃度変化に対 する擬一次速度定数(kobs)及び初速度(v0)を求めたと ころ、kobsに基質濃度依存性はなく、錯体(3)よりも値が小さいため、活性が低いことが示唆され た。また、v0はTNPPの濃度増加と共に増加しているが、Michaelis-Menten挙動は示さず、錯体 (3)とは異なる機構で反応が進んでいると考えられる。これは錯体(4)においては、Zn原子に水 分子が配位していないことによると考えられる。 Ⅳ 結論 金属酵素の機能と構造との関係に関する知見を得るために、アスコルビン酸酸化酵素モデ ルとして2種の銅(Ⅱ)錯体(錯体 1,2)を、およびリン酸エステル加水分解酵素モデルとして 2種の亜鉛(Ⅱ)錯体(錯体 3,4)を合成した。それら4種の錯体の構造を X 線構造解析により明 らかにした。アスコルビン酸酸化酵素モデル錯体の場合、温度依存性がアスコルビン酸酸化 酵素と異なり、反応機構が違うことが考えられる。リン酸エステル加水分解酵素モデル錯体 の場合、錯体(3)は、一般的なエステル加水分解反応機構と同じであった。しかし、錯体(4) は、活性が低く、錯体(3)とは異なる反応機構が考えられる。これは、錯体(4)では反応に関与 すると考えられる水分子が、亜鉛イオンに配位していないためと推測される。