シアノバクテリアの概日システムにおける 入力・出力機構の解析
Analyses on input and output mechanisms
in the cyanobacterial circadian system
2013 年 2 月 細川 徳宗
Norimune HOSOKAWA
シアノバクテリアの概日システムにおける 入力・出力機構の解析
Analyses on input and output mechanisms
in the cyanobacterial circadian system
2013 年 2 月
早稲田大学大学院 先進理工学研究科 電気・情報生命専攻
細胞分子ネットワーク研究
細川 徳宗
Norimune HOSOKAWA
目次
用語解説 2
第一章 序論 3
第二章 シアノバクテリア概日時計の入力機構の解析 13
第三章 シアノバクテリア概日時計の出力機構の解析 51
第四章 総合議論 78
謝辞 86
研究業績 87
用語解説
CT circadian time の略。概日リズムの周期を 24時間として,それを 24等分し た時間。例えば,概日リズム周期が28時間の場合は概日時間の1時間は28/24時 間に相当する。
LL 連続明条件の略。
DD 連続暗条件の略。
LD 明暗条件の略。本研究では,12 時間明期-12 時間暗期のサイクルによって,
概日時計を同調させた。
主観的黄昏,主観的夜明け 明暗サイクル同調後の連続明条件下における主観的 時間のことで,CT12-18を主観的黄昏,CT24-CT6を主観的夜明けとした。
IPTG isopropyl-β-D-thiogalactopyranosideの略。
Prx 酸化還元酵素ペルオキシレドキシンの略。
第一章 序論
概日時計の概念
バクテリアから高等動植物に到るまで,地球上に生息するほとんどの生物は明期と暗 期 (昼と夜)の周期的な環境変化にさらされて生きている。この周期的な環境変化に対し て適応するために「約 24 時間周期の内因性の生体リズム」を持っている。それが「概日 (サーカディアン) リズム」である。そして,この約 24 時間周期の基本振動を発生させてい るのが「概日時計 (体内時計)」であり,「振動体」である。この概日時計を利用することに よって,生物は両立できない生体内反応の時間的制御や,明期と暗期の環境変化を予 測し,効率的に順応している。例えば,動物は安全で効率的な活動時間を選択すること ができ,植物では開花時間の決定や,光合成機能をより効率的に行うために,暗期から 明期に移り変わるタイミングを予測してその準備をする。さらに,人間においても睡眠・覚 醒,脳波,ホルモン分泌,体温など様々な生命活動において概日リズムが見られる。また,
概日時計は光周性などの高度な環境適応メカニズムにも関わっていることがわかっており,
概日時計を基盤として日長を測ることで適切な季節に植物の花成や動物の生殖,休眠を 実現させている。
この概日時計の特性としては以下の3つの要素が挙げられる。第一に,周期が連続明 期や連続暗期など外部環境を一定にした状態でも約24時間を維持することができる「自 由継続性」を持つこと。第二に,光や暗パルスなどの特定の外界刺激によって位相が変 化し同調することができる「位相応答性」を持つこと。第三に,生理的な温度の範囲内で 周期の長さを比較的一定に保つことができる「温度補償性」を持つこと。以上の特性を持 つことによって外界の環境変化に応じて時計の時刻を調節することができ,夏や冬など 様々な温度環境下でも一定のリズムを刻むことが可能である。
念的には,簡単に記述すると三つの要素で説明ができる。まず一つ目として,中心に約 24時間周期の概日リズムを発生させる振動体 (概日時計) がある。二つ目に,その概日 時計に外界の環境情報 (昼夜など) を伝え,外部環境との調和を取るために概日時計 のリセットをしたり,リズムを整えたりする入力系がある。そして三つ目に,概日時計から発 生した振動を様々な生命活動に利用する出力系がある。この振動体,入力系,出力系で 構成される概日時計システムによって様々な生命活動の制御が行なわれている。
概日時計の発振機構
概日時計の研究は,現在では分子遺伝学,生理学,神経科学,理論生物学など幅広 い分野で展開されている。その中でも,特に概日時計の分子メカニズムについての研究 が盛んに行われるようになったきっかけが,ショウジョウバエにおける突然変異体の単離で ある (Konopka et al., 1971)。1984年に,この原因遺伝子がクローニングされperiod (per)と命名された (Bargiello et al., 1984)。それ以降,アカパンカビfrequency (frq) やショウジョウバエのtimeless (tim),マウスのClock,Bmal1 など次々と時計遺伝子が 同定されていった (McClung et al., 1989; Myers et al., 1995; King et al., 1997;
Gekakis et al., 1998)。
このように同定されていった時計遺伝子を比較すると,モデル生物間において広く保 存されているわけではないことが明らかとなった。哺乳類,鳥類,ショウジョウバエの間に はそれぞれ共通の時計蛋白質のホモログが発見されたが,アカパンカビ,シロイヌナズナ,
シアノバクテリアの時計遺伝子との間には,相同性は全く見られなかった。以上の理由か ら,概日時計の発振機構は,遺伝子や蛋白質に還元されないシステムレベルで保存され ていることが予想された。全生物に共通の概日時計の発振機構として「転写・翻訳フィー ドバックループ」モデルが提唱されていた。このモデルは,概日時計を構成する時計遺伝 子の転写を,自身が生産する翻訳産物で抑制することで 24 時間周期の振動を発生させ
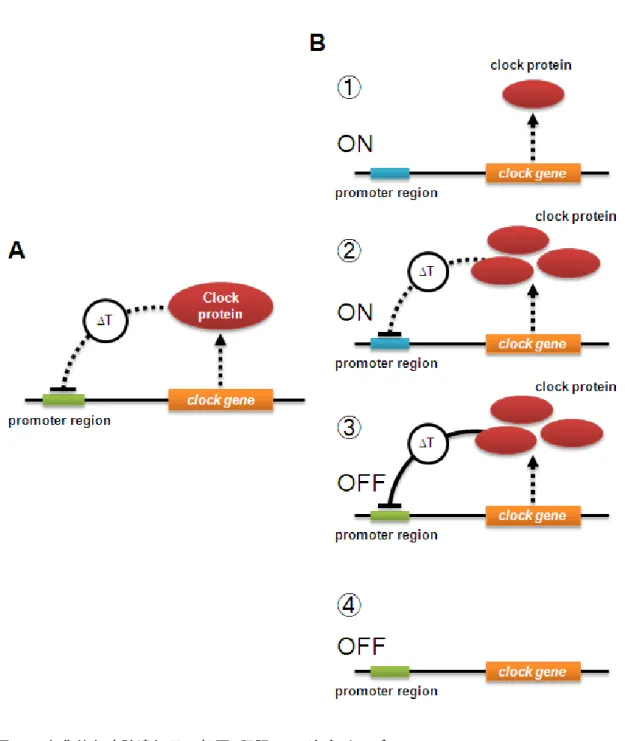
図1. 古典的な時計遺伝子の転写・翻訳フィードバックモデル
(A) 古典的な時計遺伝子の転写・翻訳フィードバックに必要な4つの要素。古典的な転写・翻訳 フィードバックモデルは,時計遺伝子,その翻訳産物である時計蛋白質,時計蛋白質による時計 遺伝子のプロモーター領域の抑制機構とその抑制までの時間遅れ (ΔT)によって説明される。
(B) 古典的な時計遺伝子の転写・翻訳フィードバックモデルの概要。古典的な転写・翻訳フィード バックモデルは,下記のように説明される。① 時計遺伝子のプロモーターは常に活性化状態に あり,時計蛋白質が生産される。② 時計蛋白質は時計遺伝子のプロモーター領域を抑制する働 きを持つため,その蓄積量に応じて,時間遅れ (ΔT)を伴ってプロモーターを抑制する。③ 時計 蛋白質の蓄積量が増加すると時計遺伝子の転写が停止する。④ 時計蛋白質が分解され,また
るというもので,全てのモデル生物において時計遺伝子の転写・翻訳リズムが観察される ため,当初どの生物にも適用しうるモデルであった (図1; Zhang et al., 2010)。しかし,
後にシアノバクテリアにおいて,このモデルは一部否定されることとなった。
シアノバクテリアを用いた概日時計研究
シアノバクテリアは,概日時計を示す最も単純なモデル生物である。酸素発生型の光 合成生物で,約35億年前に発生したとされている。機能・構造共に植物の葉緑体に近い ことから,葉緑体の系統的起源生物とされている。また,単細胞性,多細胞性,淡水性,
海洋性,土壌性,耐冷性,常温性,好熱性などの多様性を持つ。さらに原核生物が概日 リズムを持つことが初めて確認されたことで有名である (Kondo et al., 1993)。原核生物 であるシアノバクテリアを用いて概日リズムの解析をすることの最大の利点は,システムの 単純さと分子遺伝学的な解析の容易さにある。単細胞性シアノバクテリア S. elongatus はそのゲノムサイズがおよそ2.7Mbで大腸菌のそれよりも小さく,遺伝子数も3000以下と 少ないため,遺伝子の特定や生体内制御ネットワークの推定を容易に行える。さらには,
形質転換効率が非常に高いため,遺伝子の相同組換えを利用して遺伝子破壊や遺伝 子ターゲッティングがとても容易に行なえる。また,寒天培地上で数多くのコロニーを選択 することによって簡単に変異体が得られる。このため実験に非常に扱いやすい生物であ る。以上の理由から,この生物を用いて研究を行うことで,概日時計システムの基本原理 に鋭く迫ることが可能である。
シアノバクテリアにおける概日時計の研究は 1990 年代初頭に開始された。シアノバク テリアS. elongatus において,psbAI 遺伝子は光合成系 II 遺伝子の中核タンパク質を コードしている。この遺伝子は細菌内で非常に強く発現することが報告されていたが (Li et al., 1993),発光細菌のルシフェラーゼ遺伝子luxAB 遺伝子を発光レポーターに用
いて psbAI 遺伝子のプロモーター領域の下流に luxAB 遺伝子を融合させることで
psbAI 遺伝子の発現を発光で連続的に観察する方法が報告された (Kondo et al.,
1993)。この psbAI 遺伝子の転写パターンは,連続明期において顕著な概日リズムを示
していた。さらに,psbAI 遺伝子の概日リズム周期変異体,原因遺伝子のクローニングを 試みると,ほとんどがkaiA,kaiB, kaiC,という3つの遺伝子群のどこかに変異があること が明らかとなり,時計遺伝子kaiA,kaiB,kaiCが同定された (Ishiura et al., 1998)。k ai 遺伝子をそれぞれ欠失,ないしは不活化した場合,遺伝子発現に見られる概日リズム は消滅する。さらに,オペロンである時計遺伝子kaiBCの転写をkaiA 遺伝子が活性化 し,kaiC 遺伝子が抑制することが確認された。このため,概日時計の中核はkaiABC遺 伝子であると考えられ,Kai 蛋白質との転写・翻訳フィードバックが提唱された (Ishiura et al., 1998)。しかしその後,転写だけではなく直接的にタンパク質レベルでの相互作用 が確 認 され,KaiABC 蛋 白 質 の相 互 作 用 に関 する詳 細 な研 究 が展 開 されていった (Iwasaki et al., 1999; Nishiwaki et al., 2000; Hayashi et al., 2003)。
時計蛋白質 KaiABC の性質
シアノバクテリアで同定された時計蛋白質KaiA,KaiB,KaiCは,真核生物の蛋白質 と相同性を持たない新規蛋白質である。KaiA,KaiB ともに特徴的なモチーフは存在し ないが,KaiCに関してはATP/GTP結合モチーフ (P-loop) を持っており,in vitro で ATPと結合することが確認されている (Iwasaki et al., 1999; Nishiwaki et al., 2000;
Hayashi et al., 2003)。また,KaiCには自己リン酸化活性が存在し,細胞内でKaiCリ ン酸化の割合には概日リズムが観測される。このリン酸化反応は,KaiCの431番目のセ リン残基と432番目のスレオニン残基の2ヶ所で起こることが質量分析法やX線構造解 析の結果から明らかとなった (Nishiwaki et al., 2004; Xu et al., 2003)。この2つの KaiCのリン酸化サイトをアラニンに置換するとkaiBC遺伝子のプロモーター活性に概日 リズムが見られなくなることから,KaiC のリン酸化リズムは概日リズム発生に必須と考えら
れた (Nishiwaki et al., 2004)。
KaiCのリン酸化は,基本的にはKaiC 自体の自己リン酸化と自己脱リン酸化によって 調節されているが,そのリン酸化反応をKaiA蛋白質が促進し,KaiAとKaiCが複合体 を形成する。そして,KaiB は KaiA-KaiC の複合体に結合し, KaiC リン酸化促進効 果を抑制することで脱リン酸化が進み,複合体が解離する。このKaiAによるKaiCのリン 酸化の促進からKaiABC複合体の解離までのまでの一連の変化が 24 時間周期で起こ ることが知られており,上述の時計蛋白質間相互作用が 24 時間周期の振動を発生させ ていると考えられている (Iwasaki et al ., 2002; Kitayama et al., 2003; Kageyama et al., 2003)。
時計蛋白質 KaiB と KaiC は明期でそのタンパク質量も変動しているが,暗期では蓄 積量が一定となることが確認された。しかしながら,KaiA,KaiB,KaiCのタンパク質蓄積 量に全くリズムが見られなくなるにも関わらず,KaiC のリン酸化リズムは持続することが明 らかとなった。このことから,それまで様々な生物で概日リズムの基本振動は転写翻訳フィ ードバックループによって起こるとされてきたが,S. elongates では時計遺伝子の転写翻 訳フィードバックは概日振動発生に必須でないことが示された (Tomita et al., 2005)。 さらに,試験管内で時計遺伝子の翻訳産物である時計蛋白質 KaiA,KaiB,KaiC と ATPを特定の濃度で反応させると,4~5日もKaiCのリン酸化リズムが減衰せず,きれい な概日リズムを示した (Nakajima et al., 2005)。これらの研究から,KaiABCによる生 化学振動が概日時計の基本振動子であると考えられている。
このように,中心振動体 (KaiABC 振動子)については非常に研究が進んでいる。一 方で,概日システムにおける入力・出力機構の研究については振動体の研究と比較して まだまだ解明すべき点を多く残している。本論文では,概日システムの中の入力・出力機 構をターゲットに研究を展開した。
引用文献
Bargiello TA, Jackson FR, Young MW. (1984) Restoration of circadian behavioural rhythms by gene transfer in Drosophila. Nature 312(5996):752-4.
Gekakis N, Staknis D, Nguyen HB, Davis FC, Wilsbacher LD, King DP, Takahashi JS, Weitz CJ. (1998) Role of the CLOCK protein in the mammalian circadian mechanism. Science 280(5369):1564-9.
Hayashi F, Suzuki H, Iwase R, Uzumaki T, Miyake A, Shen JR, Imada K, Furukawa Y, Yonekura K, Namba K, Ishiura M. (2003) ATP-induced hexameric ring structure of the cyanobacterial circadian clock protein KaiC. Genes Cells 8(3):287-96.
Ishiura M, et al. (1998) Expression of a gene cluster kaiABC as a circadian feedback process in cyanobacteria. Science 281:1519–1523.
Iwasaki H, Nishiwaki T, Kitayama Y, Nakajima M, Kondo T. (2002) KaiA-stimulated KaiC phosphorylation in circadian timing loops in cyanobacteria. Proc Natl Acad Sci U S A 99(24):15788-93.
Iwasaki H, Taniguchi Y, Ishiura M, Kondo T. (1999) Physical interactions among circadian clock proteins KaiA, KaiB and KaiC in cyanobacteria. EMBO J 18(5):1137-45.
Kageyama H, Kondo T, Iwasaki H. (2003) Circadian formation of clock protein complexes by KaiA, KaiB, KaiC, and SasA in cyanobacteria. J Biol Chem 278(4):2388-95.
King DP, Zhao Y, Sangoram AM, Wilsbacher LD, Tanaka M, Antoch MP, Steeves TD, Vitaterna MH, Kornhauser JM, Lowrey PL, Turek FW, Takahashi JS. (1997) Positional cloning of the mouse circadian clock gene. Cell 89(4):641-53.
Kitayama Y, Iwasaki H, Nishiwaki T, Kondo T. (2003) KaiB functions as an attenuator of KaiC phosphorylation in the cyanobacterial circadian clock system. EMBO J 22(9):2127-34.
Kondo T, Strayer CA, Kulkarni RD, Taylor W, Ishiura M, Golden SS, Johnson CH. (1993) Circadian rhythms in prokaryotes: luciferase as a reporter of circadian gene expression in cyanobacteria. Proc Natl Acad Sci U S A 90(12):5672-6.
Li R, Golden SS. (1993) Enhancer activity of light-responsive regulatory elements in the untranslated leader regions of cyanobacterial psbA genes. Proc Natl Acad Sci U S A 90(24):11678-82.
McClung CR, Fox BA, Dunlap JC. (1989) The Neurospora clock gene frequency shares a sequence element with the Drosophila clock gene period. Nature
339(6225):558-62.
Myers MP, Wager-Smith K, Wesley CS, Young MW, Sehgal A. (1995) Positional cloning and sequence analysis of the Drosophila clock gene, timeless. Science 270(5237):805-8.
Nakajima M, Imai K, Ito H, Nishiwaki T, Murayama Y, Iwasaki H, Oyama T, Kondo T. (2005) Reconstitution of circadian oscillation of cyanobacterial KaiC phosphorylation in vitro. Science 308(5720):414-5.
Nishiwaki T, Iwasaki H, Ishiura M, Kondo T. (2000) Nucleotide binding and autophosphorylation of the clock protein KaiC as a circadian timing process of cyanobacteria. Proc Natl Acad Sci U S A 97(1):495-9.
Nishiwaki T, Satomi Y, Nakajima M, Lee C, Kiyohara R, Kageyama H, Kitayama Y, Temamoto M, Yamaguchi A, Hijikata A, Go M, Iwasaki H, Takao T, Kondo T. (2004) Role of KaiC phosphorylation in the circadian clock system of Synechococcus elongatus PCC 7942. Proc Natl Acad Sci U S A 101(38):13927-32.
Konopka RJ, Benzer S. (1971) Clock Mutants of Drosophila melanogaster. Proc Natl Acad Sci U S A 68(9): 2112–2116.
Tomita J, Nakajima M, Kondo T, Iwasaki H. (2005) No
transcription-translation feedback in circadian rhythm of KaiC phosphorylation. Science 307(5707):251-4.
Xu Y, Mori T, Johnson CH. (2003) Cyanobacterial circadian clockwork: roles of KaiA, KaiB and the kaiBC promoter in regulating KaiC. EMBO J 22(9):2117-26.
Zhang EE, Kay SA (2010) Clocks not winding down: unravelling circadian networks. Nat Rev Mol Cell Biol 11:764–776.
第二章 シアノバクテリア概日時計の入力機構
要約
多くのモデル生物において,概日振動発生に時計遺伝子の転写・翻訳フィード バックは必須であると考えられてきた。しかしながら,シアノバクテリアではそ の転写・翻訳フィードバックの役割がこれまで不透明であった。それは概日振動 発生にも転写出力機構にも時計遺伝子の転写・翻訳フィードバックは必須ではな いことが明らかとなっているからである。本研究では,シアノバクテリア概日時 計システムにおける転写・翻訳フィードバックの一つの役割を明示し,概日シス テムの入力機構との関与を示した。
シアノバクテリアの時計遺伝子 kaiABC は,連続明条件下においてゲノムワイ ドな概日転写リズムの発生に必須である (Ishiura et al., 1998; Ito et al., 2009)。 一方,連続明条件下から細胞を暗条件下に移すと kaiABC の転写は速やかに停止 し,転写蓄積量もゼロレベルにまで低下するが,時計蛋白質 KaiC のリン酸化リ ズムに非常にきれいな概日リズムが観察される。このことから,時計遺伝子の転 写・翻訳は概日振動発生に必須ではないことが明らかとなった (Tomita et al., 2005)。さらに,KaiABC蛋白質とATPを試験管内で混合するだけで24時間周期 の KaiC リン酸化リズムが再構成できることが明らかとなり,概日振動発生に転 写・翻訳フィードバックは必須ではないという結果が裏付けられた (Nakajima et al., 2005)。また,kaiABC 遺伝子の新規転写・翻訳の停止する暗条件下でも概日 リズムの特性を示す転写振動が観察されることが明らかとなった (第三章で記 述;Hosokawa et al., 2011)。このことから,概日システムの転写出力機構にも時 計遺伝子の転写・翻訳は必須でないことが示唆された。よって,シアノバクテリ
アでは時計遺伝子の転写・翻訳フィードバックは発振機構と転写出力機構に必須 ではないことがわかる。
一方,多くのモデル生物で時計遺伝子の転写・翻訳フィードバックの入力機構 への関与が多数報告されているが,シアノバクテリアにおいても転写・翻訳フィ ードバックと入力機構との関連が報告されている。例えば,時計蛋白質 KaiC を 過剰発現させると数時間の位相シフトが起こることが観察され,KaiCの細胞内蓄 積量を減少させると暗パルスに対する位相シフトが鋭敏になることも観察されて いる (Ishiura et al., 1998; Xu et al., 2000)。さらに最近の研究から KaiABC蛋白 質からなる翻訳後修飾レベルで振動するKaiABC振動子 と転写・翻訳フィードバ ックの相互作用の検討が行われ,シアノバクテリアにおいては KaiABC振動子が 概日システムの中核であり,転写・翻訳フィードバックはその補助機構であるこ とが示唆された (Qin et al., 2010)。同論文中で,KaiCの蓄積量の変化が時計の 同調に重要であるという議論がなされていた。しかしながら,転写・翻訳フィー ドバックの入力機構への関与について,システマティックな理解は行われておら ず,暗刺激に対する概日時計のリセット機構 (暗刺激により位相シフトが引き起こ されるメカニズムについて)も不明瞭な点を多く残している。
本論文では,時計遺伝子の転写・翻訳フィードバックが主観的夜明けに KaiA に対するKaiCの細胞内存在比 (KaiC/KaiA比)を小さくすることでKaiABC振動 子の性質を変化させ,暗刺激に対する位相シフトを促進させることを強く示唆し た。時計遺伝子の転写・翻訳フィードバックの効果により,連続明条件下ではKaiC の蓄積量の変化に 24時間周期のリズムが生まれ,KaiC の蓄積量の変化に伴って 細胞内の KaiCと KaiAの存在比 (KaiC/KaiA 比)にリズムが見られることを確認 した。一方,暗条件下では細胞内の代謝活性が著しく低下するため,KaiC,KaiA,
KaiC/KaiA 比が一定となる。主観的夜明けから細胞に暗刺激を与えると暗期では
KaiC/KaiA比が小さい状態となり,KaiABC振動子の振動状態 (KaiCリン酸化リ ズム)が短周期・低振幅化し,脆弱化することが明らかとなった。このとき,細胞 を再び明期に移行すると大きな位相シフトが観察された。さらにkaiA過剰発現株 (ox-kaiA株)を用いて細胞内のKaiC/KaiA比を人為的に変化させると,KaiC/KaiA 比が小さくなるにつれて概日リズムが短周期・低振幅化し,暗刺激に対する位相 シフトが大きくなっていくことが明らかとなった。以上の結果から,シアノバク テリアにおいて時計遺伝子の転写・翻訳フィードバックはKaiC/KaiA比を主観的 夜明けで小さくすることで KaiC リン酸化リズムを脆弱化させ,位相シフトを鋭 敏にしていることが強く示唆された。
序論
真核生物における時計遺伝子の転写・翻訳フィードバックの入力機構への寄与 真核モデル生物を用いた研究で,時計遺伝子の転写・翻訳フィードバックと概 日システムの入力機構 (位相シフト)との関連について多数の報告がされている。
例えばマウスを用いた研究では,転写・翻訳フィードバックの構成要素であるPer1, Per2 遺伝子と位相シフトの関係が報告されている。suprachiasmatic nucleus (SCN; 視交叉上核)においてPer1,Per2遺伝子には光応答性が見られ,特定の位 相で光刺激があたえられるとPer1,Per2遺伝子のmRNA および蛋白質蓄積量の 急激な上昇が見られ,概日時計の位相シフトが観察される (Shigeyoshi Y et al., 1997; Albrecht et al., 1997; Zylka et al., 1998; Okamura et al., 1999; Field et al., 2000; Yan et al., 2002; Yan et al., 2004) 。アカパンカビを用いた研究では,frq 遺 伝子およびFRQ蛋白質は転写・翻訳フィードバックを構成する主要な要素である。
アカパンカビは光や温度が概日時計の同調因子であることが知られているが,こ れらの刺激によって,frq 遺伝子およびFRQ蛋白質の蓄積レベルが変化し,位相 シフトが起こることが報告されている (Dunlap, 1999; Aronson et al., 1994)。こ のように,マウスとアカパンカビにおいて時計遺伝子の転写・翻訳フィードバッ クと位相シフトには密接な関係がある。
シアノバクテリア概日時計の入力機構に関わる因子 CikA, LdpA
シアノバクテリアの概日時計の同調因子として,トランスポゾンを用いた突然 変 体 の ス ク リ ー ニ ン グ に よ っ て 初 め て 同 定 さ れ た の が CikA (circadian input kinase)である (Schmitz et al., 2000)。CikAは,植物の光受容体であるフィトク ロムと相同性を持つ蛋白質であり,発見当初は光受容体であると予想された。し かし,その後の解析によってS. elongatusにおいては発色団と結合するGAFドメ
インが欠損していることが明らかとなり,今のところCikAが発色団を結合してい るとの知見は得られていない (Mutsuda et al., 2003)。cikA遺伝子破壊株では,
概日リズムの周期が 3 時間程度短縮されることに加え,5 時間の暗パルスによる 概日リズムの位相シフト量が著しく減少する (Schmitz et al., 2000)。よって,概 日時計の入力系に関わる因子として考えられた。CikA以外の入力系に関わる因子 として,LdpA (light dependent period A) が同定されている (Katayama et al.,
2003)。LdpA も CikA と同様にトランスポゾンを用いた変異体スクリーニングに
よって同定された。野生株において,概日リズムの周期は光強度に依存して変化 することが知られており,光強度の増加に伴って短周期化する。しかしながら,
ldpA遺伝子破壊株では野生株で観察されるような概日リズムの短周期化は観察さ れず,光強度を変化させても周期長の変化は殆ど観察されなかった (Katayama et al., 2003)。LdpAは電子の授受に関わる4Fe-3Sクラスターの結合領域を含むため,
細胞内の酸化還元状態のセンサーとして働き,細胞への光入力機構に関与するこ とで概日リズムの入力機構にも関与していることが示唆された (Katayama et al.,
2003)。このように,シアノバクテリア概日時計の入力機構の研究は,入力系に関
わる蛋白質の解析を主軸に開始された。
概日時計の入力機構と細胞内代謝
CikA はシアノバクテリア概日時計の入力系に関与していることが示唆された が,前述のように発色団を結合しないことから光を感受しているのではなく,昼 夜交替によって起こる細胞内変化を感受して機能していることが予測された。単 細胞性シアノバクテリアS. elongatus は光合成独立栄養生物であるため光依存性 が非常に高く,光条件によって細胞内環境が劇的に変化することが知られている。
例えば,暗期では細胞分裂が停止し細胞内のほとんどの遺伝子の転写蓄積量がゼ
ロレベルにまで低下する (Ito et al., 2009)。さらに,明期から暗期へ細胞を移行 後 ~5 時間で細胞内 ATP/ADP 比が明期の半分程度にまで減少する (Rust et al.,
2011)。また,光化学反応が明暗で切り替わることにより細胞内の酸化還元状態の
変化がもたらされる。CikA 蛋白質を標的とした詳細な解析により,CikA はキノ ンアナログである DBMIB (2,5-ジブロモ-3-メチル-6-イソプロピル-1,4-ベンゾキ ノン) によって分解が促進されることが明らかとなった (Ivleva et al., 2006)。
シアノバクテリア概日システムの入力機構と転写・翻訳フィードバックの関係に ついての先行研究
一方で,シアノバクテリアにおいても時計遺伝子の転写・翻訳フィードバック と位相シフトとの関係がいくつか報告されている。Ishiura ら (1998)による先行 研究では,KaiCの過剰発現によって数時間の位相シフトが起こることが報告され ている。またXu ら (2000)により,細胞内のKaiC蓄積量を減少させると位相応 答が鈍感になることが報告されている。
最近の研究 (Qin et al., 2010)ではKaiABC振動子と転写・翻訳フィードバック との関係について報告がされており,KaiCの 2つのリン酸化サイトをグルタミン 酸に置換した変異株kaiCEE株を用いた解析が行われた。kaiCEE株ではKaiCのリ ン酸化サイトがグルタミン酸に置換されていることでリン酸化状態が模倣されて おり,KaiABC 振動子の KaiC リン酸化状態はある特定の状態 (位相)で固定され ていると考えることができる。このときkaiBCの転写に~60時間のリズムが観察 されるがリズムが非常に不安定であり,概日リズムの特性である温度補償性がみ られないことから,KaiABC 振動子が概日発振機構の中核であり,転写・翻訳フ ィードバックはその補助機構であることが示唆された。さらに,転写・翻訳フィ ードバックの構成要素である KaiC 蛋白質の翻訳が KaiABC振動子の同調機構に
寄与しているという議論がされている。このように時計遺伝子の転写・翻訳フィ ー ド バ ッ ク が 位 相 シ フ ト (入 力 機 構)に 寄 与 し て い る と い う 報 告 は さ れ て き た (Ishiura et al., 1998; Qin et al., 2010)。しかしながら,「転写・翻訳フィードバッ
クはKaiABC振動子にどのように寄与し,そしてどのように位相シフトを引き起
こしているのか」,その詳細は全く不明であった。本研究では,時計遺伝子の転写・
翻訳フィードバックの入力機構への寄与について,その詳細を解析した。
実験方法
使用した野生株および培養条件
本研究ではシアノバクテリア S.elongatusの野生株として,S.elongatus のゲ ノム上の Neutral Site I (NSI)にバクテリア由来のルシフェラーゼ遺伝子 luxAB のプロモーターを除いた断片とkaiBC のプロモーター領域を連結させたDNA断
片 (PkaiBC ::luxAB)に,さらにクロラムフェニコール遺伝子をセレクションのマー
カーとして組み込んだ株 (NUC42)を使用した (Nakahira et al., 2002)。
培養条件は,温度条件30℃,白色光・光条件~30 μE· m-2· s-1とした。また培 地はBG -11 (Bustos and Golden,1992) に 0.03 mMのクエン酸と,培地のpH調 節のため1 mM HEPES-NaOH (pH7.5) を加えた液体培地もしくは,その溶液に 1.5% (wt/vol) Bacto Agarを加え,固めた寒天培地を使用した。
発光レポーター株を用いた概日時計の位相解析
バクテリア由来のルシフェラーゼ遺伝子luxABをターゲット遺伝子プロモータ ーの下流に連結したシアノバクテリア細胞を用いて,ターゲット遺伝子のプロモ ーター活性を経時的に測定し,細胞の概日時計の位相をモニタリングする生物発 光測定を行った。luxAB 遺伝子にコードされるルシフェラーゼは還元型フラビン モノヌクレオチド (FMNH2),長鎖のアルデヒド (デカナール)および酸素を基質 とする反応を触媒し,490 nmの生物発光を示す。反応式は以下の通りである。
FMNH2 + n-decanal + O2 → n-decanoic acid + H2O + light
この生物発光を,光電子増倍管を用いた連続生物発光測定装置によってリアルタ イムに近い状態で測定した。シアノバクテリア株は泳体培養後,OD730 を 0.2 に そろえ1/10000に希釈してBG-11寒天培地に 10 μl プレーティングした。5日ほ ど連続明条件下で培養を行った後,1% n-decanalを細胞に直接接触しないように
適当量加え,生物発光測定を行った。細胞を 2回の 12L12Dで明暗同調をかけた 後,発光測定を行った。
ノーザンブロット解析 (1)細胞のサンプリング
シアノバクテリア細胞を液体培地でOD730が 0.3以上になるまで培養をした後,
1.4 lのBG-11の入った大型培養槽で培養した。大型培養槽で光条件片面~30 μE•
m-2• s-1で,OD730= 0.3まで培養をした。それから,12L12Dの LDサイクルを2 回かけ,概日時計の同調を行った。その後,適切な時間帯にサンプリングを行っ た。サンプリング時には,50 mlのファルコンチューブに30 mlほど氷をと入れ たものを用意し,培養槽からサンプルを取り出し,冷やしておいたファルコンチ ューブに移した。1つのファルコンチューブに取るサンプルの量は細胞数を 6 ml
/OD730値として設定した。その後50 mlファルコンチューブを4℃,3500 rpm で8 分遠心し,上清を捨てさらに1 分遠心をした。遠心後すぐに 50 mlファルコ ンチューブを氷上に移し,下に溜まったサンプルを再懸濁し,氷上に用意した1.5 mlエッペンチューブに移した。エッペンチューブを4℃,15000 rpmで 1 分遠心 し,上清を完全に捨ててシアノバクテリア細胞のみを-80℃に保存した。
(2) RNA抽出
ノーザンブロッティングを行うため,集菌した細胞からRNAを抽出した。RNA 抽出には改変ホットフェノール法を用いた (Mohamed and Jansson, 1989)。まず Suspension buffer,Lysis buffer,250 mM EDTA (pH 7.5)の 3種類のバッファー を調合した。Suspension bufferは0.3 M sucroseと10 mM sodium acetate (pH 4.5)を等量混ぜ,120℃ 15分オートクレーブをして滅菌をした。10 mM sodium
acetate (pH 4.5)のpH調節には,塩酸を使用した。Lysis bufferは 2% (w / v) SDS と先ほど使用したものと同じ 10 mM sodium acetate (ph 4.5) を等量混ぜた。
EDTAは 500 mM を水 (DW)で 250 mMに希釈した。さらにこの 3 つの buffer をSuspension buffer 125 μl,Lysis buffer 188 μl,250 mM EDTA 38 μlの割合 で混ぜlysis-EDTA-suspension bufferを作製した。
集菌した細胞にフェノール350 μlと Lysis-EDTA-suspension buffer 350 μlを 加え,よく混和し,65℃で3 分,時々vortexしながらインキュベートした。その 後,4℃で15000 rpm,10 分遠心をして,ピンク色の上澄みを新しい1.5 mlエッ ペンチューブに入れた。これによって,細胞から内容物のみを取り出してきた。
さらに,この回収した溶液に350 μlのクロロホルムを加え,vortexして,20℃で 15000 rpm,10 分遠心をし,上澄みを1.5 mlエッペンチューブに入れた (クロロ ホルム抽出)。その後,8 M LiCl 75 μlとエタノール800 μlを回収した上澄みに加 え,混和して,4℃で15000 rpm,20 分遠心をした。エタノールをピペットで吸 出し,ペレットを確認後,ペレットに100 μlのエタノールをかけ,加えたエタノ ールを完全に吸い出して,エタノールが乾くまでエッペンチューブのふたを開け たまま 2,3 分放置した (エタノール沈殿およびエタノール洗浄)。エタノールが 完全に乾いたら,100 μlのDW を加え,ペレットを溶解させた。これにISOGEN (ニッポンジーン) を 500 μl 加え混和後,3 分室温で静置した。その後,100 μl クロロホルムを加え混和をして, 4℃で 15000 rpm,15 分遠心した。上澄みを 回収し,新しいエッペンチューブに入れたら,クロロホルム抽出を行い,8 M LiCl を85 μlとエタノール800 μlを加え,混和をして,エタノール沈殿→エタノール 洗浄→エタノールを乾かした後173 μlの DWを加える。このチューブを65℃で2 分インキュベートして,ペレットを溶解し,さらにチューブミキサーで溶液を混 ぜる。この後,DNaseI 4 μl,DNaseI buffer 20 μl,RNase inhibitor 3 μl,を加
え 37℃で1 時間インキュベートした。処理の後,フェノール 100 μlとクロロホ ルム100 μlを加え混和をして,20℃で15000 rpm,10 分遠心し,上澄みを新し いエッペンに移した。クロロホルム抽出をした後,回収した上澄みに8 M LiCl 50 μlと,エタノール 400 μlを加えエタノール沈殿→エタノール洗浄をした。エタノ ールが完全に乾いたことを確認したら,50 μl の DW に溶解し,濃度を測定して RNAが抽出できていることを確認した。
(3) ノーザンブロット解析
ノーザンブロット解析には,ターゲット遺伝子を検出するためジゴキシゲニン
(DIG)標識されたプローブを PCR により作製し,使用した。具体的なノーザンブ
ロッティング解析の操作は以下のように行った。まず抽出しておいたRNAの濃度 をそろえた。希釈する際にはNuclease Free Water (Ambion)を使用した。濃度調 節後,1.2%アガロースゲルに1ウェルにつきRNA量が2 μgとなるようにロード
し,100 Vで 4 分泳動した。RNAが泳動されていることを写真で確認後,タッパ
ーに20×SSC bufferをゲルが浸るくらいまで入れ,ゲルをタッパーに移し 15 分 振盪させた。Nuclease Free Water 50 mlと 20×SSCバッファー 50 mlを新 しいタッパーの中に入れた。タッパーの上にガラスを乗せ,その上にろ紙を乗せ,
20×SSCバッファーを50 ml程度かけてろ紙をしめらせておいた。アガロースゲ ルをしめらせておいたろ紙の上に裏返して乗せ,ゲル穴の部分を全てピンセット で貫通させた。それからゲルの上にメンブレン,ろ紙,紙タオルと乗せ,その上 に重しを乗せた。紙タオルは数回換え,タオルが水をあまり吸わなくなってきた ら,そのまま一晩ブロッティングをさせた。翌日,メンブレンをゲルからはがし,
UVクロスリンカーでメンブレンを処理した。ハイブリバックを用意し,その中に メンブレンと10~30 mlの DIG EasyHyb (Roche) を入れた。ハイブリバックの
口をしっかりとシーラーでとめ,ハイブリバックごと振盪させながら50℃で1 時 間保温した。この間に標的RNAを検出する2本鎖DIG-DNAプローブ (ラベルさ れた2本鎖DNA) を95℃で10 分保温した。インキュベート後,氷水につけて急 冷した。ファルコンチューブを用意し,先ほど処理したDIG-DNAプローブ 12 μl とDIG‐EasyHyb 30 ml を入れて混和した。ハイブリオーブンからハイブリバッ クを取り出し,中の液のみを全て捨てDIG-DNAプローブとDIG‐EasyHybの混 合液を新たにハイブリバックの中に入れ再び口を閉じた。その後,振盪させなが ら50℃で一晩反応させた。翌日,メンブレンを洗う作業とフィルムの現像を行っ た。実験で使うWashing buffer,Blocking buffer, Detection buffer, 抗体溶 液の各溶液を調合した。washing bufferは, 10×washing buffer (Roche) 2.5 ml に対して,DW 22.5 ml を混ぜた。Blocking buffer は,10×Blocking solution (Roche) 2.5 ml,マレイン酸 2.5 ml,DW 20 ml を混ぜたものを使用した。
Detection bufferは,10×Detection buffer (Roche) 2.5 mlに対し,DW 22.5 ml を混ぜた。抗体溶液はBlocking buffer 25 mlにさらに AP (アルカリフォスファタ ーゼ) 標識抗ジゴキシゲニン (DIG) 抗体 2.5 μlを混ぜ使用した。前日から一晩反 応させていたメンブレンをハイブリバックから取り出し,新しいタッパーの中に 入れた。その中にさらに0.1%SDS 溶液と 2×SSC (20×SSC bufferを DWで 10 倍希釈した) 溶液を入れ,室温で 5 分振盪させた。この反応後,タッパーの中の 液を完全に捨て,再度同じ操作を行った。次に 0.1%SDS溶液と0.1×SSC (20× SSC bufferを 200倍希釈した) 溶液を入れ,65℃で15 分振盪させながら保温し た。その後,タッパーの中の液を全て捨て再度同じ反応を行った。washing buffer
25 mlをタッパーの中に入れ,室温で2 分メンブレンを振盪させた。タッパーの
液を完全に捨てた後,25 mlのBlocking bufferを入れ,室温で30 分振盪させた。
次いで,タッパーの液を全て捨て,新たに抗体溶液を25 ml入れ,30 分振盪させ
た。その後,タッパーの中の溶液を全て捨て,washing buffer 25 mlをタッパー の中に入れて室温で 15 分振盪させた。タッパーの中の液をすべて捨て,再び同 じ操作を行った。発光基質溶液調合のために25 mlのDetection bufferから400 μl
を1.5 mlチューブに移し,残りの液を全てタッパーの中に入れ3 分室温で振盪さ
せた。この間に,1.5 mlチューブのDetection buffer 400 μl に対して100×CSPD 4 μl を加えて発光基質溶液をつくった。厚紙の上に白い紙,さらに全体をサラン ラップで包んだメンブレンを乗せる台紙を用意した。この台紙に振盪の終わった メンブレンを,水気を切って乗せた。メンブレンに発光基質溶液をまんべんなく かけた。その上に透明なフィルムを乗せ, 30℃で10分保温した。ノーザンブロッ ティングの結果は,Image-J (NIH) を用いて解析した。
ウェスタンブロット解析
前 述 の ノ ー ザ ン ブ ロ ッ ト の と き と 同 様 に 集 菌 し た 細 胞 に 1×SDS (Sodium Dodecyl Sulfate) バッファーを300 μl加え,多検体細胞破壊装置 (YASUI KIKAI) を用いて[2500 rpm on 30秒/off 30秒] × 6回の条件で細胞を破壊した。破壊した 細胞を90℃で 1分インキュベートしてから 4℃,15000 rpm,3 分遠心して可溶 画分を新たなエッペンチューブに取り出した。BCA法によって細胞抽出液のタン パク質濃度を測り,それぞれが同じ濃度になるように1×SDS (with 0.2% BPB, 2-mercaptoethanol) バッファーを使ってサンプルの調製をした。10~15%ポリア クリルアミドゲルを用いてSDS-PAGE (Polyacrylamide gel electrophoresis)を行 った。この際,マーカーはRPN800E full-rangeマーカー (GE healthcare)を用い た 。 SDS-PAGE が 終 了 す る 前 に , あ ら か じ め PVDF (MILLIPORE;
PolyVinylidine DiFluoride)メンブレンをメタノールに5分振盪させた後,転写バ ッファーに 15 分振盪させた。転写用ケースのプラス側を下にして,スポンジ→
ろ紙→PVDF メンブレン→アクリルアミドゲル→ろ紙→スポンジの順に挟み,余 分な所を切り取った後200 mA,90分転写を行った。次に,blocking buffer (10%
skim milk,0.3% Tween-20)にメンブレンを浸し,90 分振盪した。PVDF膜を1
×PBSTで洗い,KaiCと KaiA抗体をそれぞれ1:3000,1:1000の割合で blocking
bufferを用いて希釈し,抗体反応を室温で1 時間おこなった。その後,1×PBST
でメンブレンを洗い,anti-rabbit IgG (GE healthcare) を 1:1000 で blocking bufferに混ぜ,抗体反応を1 時間おこなった。抗体反応後,PVDF膜を1×PBST で 洗 浄 し た 。 そ の 後 ,Western blotting detection reagents (Amersham Bioscience) にメンブレンを浸してフィルム (GE healthcare) に露光したものを 現像した。シグナルの定量方法は,ノーザンブロット解析のときと同様におこな った。
結果
主観的夜明けに与える暗刺激は概日時計の位相を明期開始時刻に固定する 初めに暗刺激による概日時計の位相応答を検討した。発光レポーター株を12時
間明期・12 時間暗期 (12L12D)の明暗サイクルに同調させた後,明期の様々な時
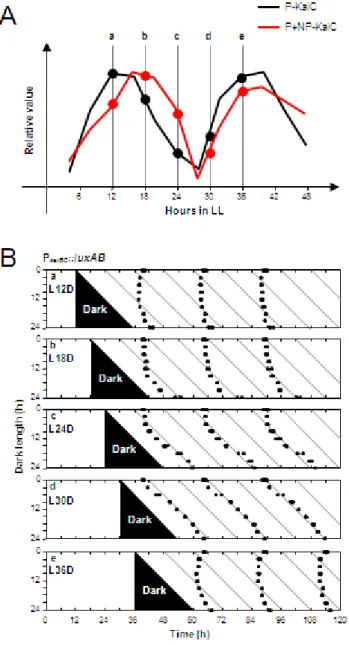
刻で 3~24 時間の長さの暗刺激を与え,刺激後の位相変化を観察した。実験スケ ジュールを図1A に示す。検討の結果,主観的黄昏 (明期12,18,36時間目) に 暗刺激を与えた場合は概日時計の位相は暗刺激前とほとんど変わらず,刺激前の 位相を刺激後も維持することがわかった。これは,先行研究の結果と一致した (図 1B; Xu et al., 2000)。しかしながら主観的夜明け (明期24,30時間目) に暗刺激 を与えた場合は,発光リズムの位相が暗刺激を与えていないコントロールと比べ て大きく変化し,刺激の長さによらず刺激後明期12時間目に発光リズムのピーク が観察された。そして,図 1Bに示すように明期開始の直線と平行に発光リズムの ピークタイムが並ぶlight onリセットパターンを示すことが明らかとなった。こ れらの結果から,主観的夜明けから細胞に暗刺激を与えた場合,暗期 (刺激後) に おける概日時計の状態に変化がみられることが予想された。さらに注意深く見て みると暗期でのリン酸化リズムの初めのピークタイムから,その前の 3 時間の間 (Light on の時間をCT0とすると,主観的昼から夜にかけて;おおよそ CT 9~12 までの間) に暗刺激が当たっていると,大きな位相応答が見られることも同時に観 察された。
主観的夜明けの暗刺激による連続暗条件下の KaiC リン酸化リズムの減衰
そこで主観的黄昏と主観的夜明けから細胞を暗期に移行した場合,連続暗期中 で概日時計の状態に変化が見られるかを検討した。概日時計の状態を観察するた めに,連続暗期中のKaiCリン酸化リズムを解析した。図2Aに実験スケジュール
図1. 主観的黄昏時の暗刺激は概日時計の位相を刺激後の明期開始時に固定する
細胞を2回の明暗サイクルに同調させた後,明期12~36時間目まで6時間毎に細胞を暗期に
移行させ (a-e),様々な長さの暗刺激を加えた。その後,再び細胞を明期に移行させ,発光レポ
ーター株を用いて概日時計の位相を検討した。
(A) 実験の概略図を示した。
(B) 実験結果を示した。プロットは生物発光リズムのピークタイムを示す。実験の結果,主観的黄 昏から細胞を暗期に移行した場合は,概日時計の位相は暗刺激を与えていないコントロールと 比較して大きな変化は観察されない。一方で,主観的夜明けから細胞を暗期に移行した場合は,
概日時計の位相は暗刺激後の明期でコントロールと比較して大きな位相変化が観察された。
を示す。検討の結果,主観的黄昏から細胞を暗期に移行したときには,KaiCリン 酸化リズムの状態は暗刺激を与えていないコントロールのものとほぼ一致した。
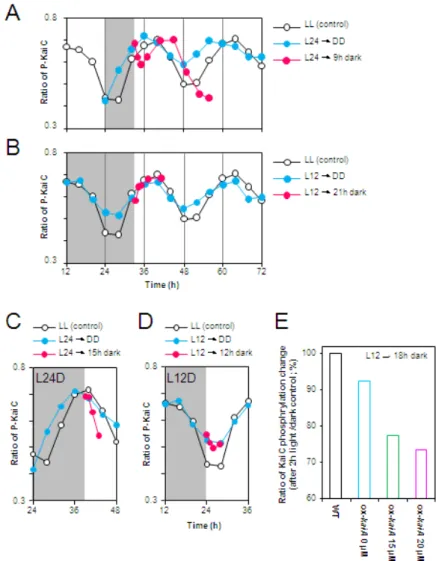
しかしながら,主観的夜明けから細胞を暗期に移行した場合は,暗期中でのKaiC リン酸化リズムがコントロールに比べて低振幅化・短周期化することが明らかと なった。特に,明期30時間目から細胞を暗期に移行した場合 (L30D),暗期中の KaiCリン酸化リズムの周期は ~21時間とコントロールに比べて 3~4時間程変化 することが明らかとなった (図 2B,2C)。暗期中にKaiCリン酸化リズムの低振幅 化・短周期化が観察されるとき位相シフトが大きいことから,暗期中に概日時計 の状態が変化することによって,位相シフトが大きくなることが示唆された。
明期中の時計遺伝子の転写・翻訳フィードバックは暗期中の KaiC リン酸化リズム の減衰を引き起こす
次に,暗期でKaiCリン酸化リズムが短周期化・低振幅化する要因を検討した。
Rust らは,in vitro において ATP/ADP 比の変化 (ATP/ (ADP+ATP)の変化) が KaiABC 生化学振動子 (KaiABC 振動子) の位相シフトの要因となることを示し た。またRustら (2011)はATP/ (ADP+ATP)比が低下すると概日時計の周期が長 周期化することを明らかにした (Rust et al., 2011)。S. elongatusは光合成独立栄 養生物であり,暗期ではATP/ (ADP+ATP) 比が明期の半分程度にまで低下するこ とが報告されている。よって, Rustらの研究からKaiCリン酸化リズムは暗期で 長周期化することが予想された。しかしながら,実際に解析をおこなったところ 図 2B に示したように本研究においては短周期化が観察された。よって,ATP/
(ADP+ATP)比の変化がKaiC リン酸化リズムの短周期化・低振幅化の要因ではな
いことが示唆された。Kim ら (2012)は,in vivo において酸化型キノンが KaiA の凝集を促進することで位相シフトを引き起こすことを示唆した。Kim らの研究
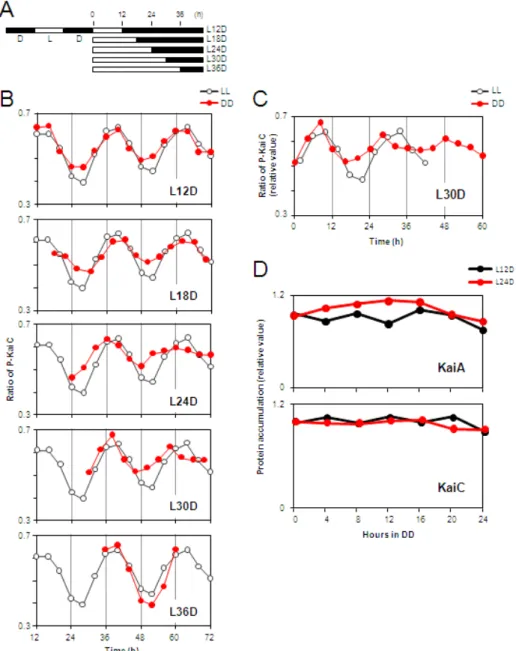
図2. 暗刺激中に見られるKaiCリン酸化リズムの減衰
細胞を2回の明暗サイクルに同調させた後,明期12~36時間目まで6時間毎に細胞を暗期に 移行させ,連続暗条件下でのKaiCリン酸化リズムを解析した。
(A) 実験の概略図を示した。
(B) 連続明期中でのKaiCリン酸化リズム (黒; コントロール)と連続暗期中でのKaiCリン酸化
リズム (赤)の解析結果を示した。L24DとL30Dでは,連続暗期中でKaiCリン酸化リズムの減
衰が観察された。
(C) L30Dにおける連続暗期中のKaiCリン酸化リズム。(B) の細胞の暗期移行時間をtime 0 h として暗期を延長させ,再表示した。
(D) 連続暗期中におけるKaiA,KaiC蛋白質の蓄積量の推移。L12D,L24Dの結果のいずれも 暗期では蛋白質量が一定となり,蓄積量の顕著な変化は観察されなかった。
によれば暗期では酸化型キノンの効果により KaiA の凝集が促進されることが予 測できる。KaiAはKaiCリン酸化を促進する役割を持つため,KaiAの凝集がKaiC リン酸化を促進することは考えにくい。しかし,図 2BではL24Dで暗期開始直後 KaiCリン酸化が促進されている。よって,リン酸化リズムの短周期化・低振幅化 の要因は酸化型キノン以外に存在することが示唆された。Nakajima ら (2010)に よるKaiABC振動子の詳細な解析から,in vitroではKaiABC振動子のKaiAと KaiCの量比を変動させることで KaiCリン酸化リズムの周期・振幅が変化するこ とが確認されており,KaiAに対して KaiCの量を減少させていくと時計の周期が 短周期化し,振幅が低振幅化する。In vivoにおいて,夜間では時計遺伝子の転写・
翻訳フィードバックは停止し KaiABC振動子のみが駆動している(Tomita et al., 2005)。よって,in vivo の KaiABC振動子は in vitroに近い状態であると考える こともできる。そこで,主観的夜明けから細胞を暗期に以降した際に,暗期で観 察されるKaiCリン酸化リズムの低振幅化・短周期化の要因は,暗期中のKaiAに 対する KaiC の量 (KaiC/KaiA 比) が主観的黄昏から細胞を暗期に移行したとき よりも少ない (比が小さい) ためであると考え,検証を行った。連続暗期中では,
時計蛋白質の合成も停止し,蓄積量は一定となる (Tomita et al., 2005)。よって KaiC/KaiA比は明期に変化し,暗期では明期中のKaiC/KaiA比が維持されると考 えた。そこで,連続明期中でKaiA/KaiC比が経時的に変化するかを確認した。そ の結果,KaiC蛋白質蓄積量は明期12~16時間目をピークにリズミックに変化し,
KaiA蛋白質はほぼ一定であること。KaiC/KaiA比を計算すると比が明期12~16 時間目に最も高く,明期28時間目付近で最も低くなるようにリズミックに変化す ることが明らかとなった (図 3A,3B)。暗期において時計蛋白質量の変化がない ことを確認するため,L12Dと L24DのKaiA蛋白質とKaiC蛋白質の蓄積量を解 析した。すると,ほぼ一定であることが確認できた (図2D)。よって,明期では
図3. KaiABC振動子の周期・振幅を調節する時計遺伝子の転写・翻訳フィードバック
(A) 連続明期におけるkaiA, kaiC 遺伝子の転写蓄積量と蛋白質蓄積量のプロファイル。データ はそれぞれ平均値を計算し,時系列平均が1となるように標準化した(n = 4)。
(B) 連続明条件下における時計蛋白質KaiCとKaiA (KaiC/KaiA)の量比の変化。計算には (A)
のKaiC,KaiA蛋白質の平均値を用いた。
(C) 野生株とkaiA過剰発現株 (ox-kaiA; 0, 10, 15, 20 μM)におけるPkaiBC::luxABのリズム変 化。
(D) 野生株とkaiA過剰発現株 (ox-kaiA ; 0, 10, 15, 20 μM)におけるKaiC/KaiA量比の定量 結果。明期4 ~28時間目まで4時間毎に細胞をサンプリングし,その平均値をグラフ化した(n = 4)。
(E and F) 野生株とkaiA過剰発現株 (ox-kaiA)におけるPkaiBC::luxABレポーターの周期測定 結果 (E),振幅測定結果 (F)。明期0 ~12時間目までの発光プロファイルから周期を算出し,
グラフ化した (n = 5)。
KaiC/KaiA 比が経時的に変化し,暗期ではその比が一定に保たれることが示唆さ れた。さらに,主観的夜明けから細胞を暗期に移行したときには,KaiC/KaiA 比 が主観的黄昏から暗期に移行したときと比べて小さいことが明らかとなった。
In vitroではKaiC/KaiA比を変化させるとKaiCリン酸化リズムの周期・振幅 に変化がみられるが,in vivo においても同様に変化するかは自明ではない。そこ で,isopropyl-β-D-thiogalactopyranoside (IPTG) によってkaiA遺伝子の誘導量 を調節することが可能なkaiA 過剰発現株 (ox-kaiA 株)を用いて KaiC/KaiA比を 人為的に小さくしたときに概日時計の周期と振幅がそれぞれ短周期化・低振幅化 するかを確認した。その結果,IPTG誘導量を 0,10,15,20,50 μMと増加さ せていくに従って KaiC/KaiA 比が小さくなり,連続明条件下での PkaiBC::luxAB の生物発光リズムがそれぞれ低振幅化・短周期化していくことが明らかとなった (図 3C-3F)。PkaiBC::luxAB の生物発光リズムの周期・振幅の変化と KaiC リン酸 化リズムの周期・振幅の変化には高い相関関係がみられ,PkaiBC::luxABの生物発 光リズムの周期・振幅の低振幅化・短周期化にともなって KaiC リン酸化リズム も低振幅・短周期化することが示されている (Qin et al., 2010)。よって,in vivo においてもKaiABC振動子の周期・振幅がin vitroと同様に,KaiC/KaiA比に伴 って変化することが示唆された。さらに,IPTG誘導性の過剰発現株 (ox-kaiC株)
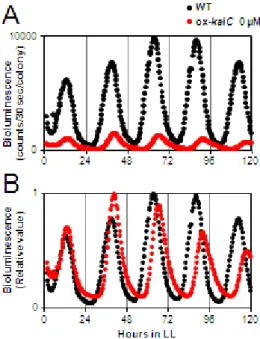
を用いてKaiC/KaiA比を大きくした場合の概日リズムの周期・振幅変化も検討し
た。ox-kaiC株はIPTG誘導性の kaiC遺伝子断片がゲノム上に元々の kaiC遺伝 子とは別に持っているため,IPTG誘導前の状態でも IPTG誘導性プロモーターか ら の リ ー ク に よ っ て KaiC 量 が 野 生 株 に 比 べ て 少 し 多 い 状 態 に な っ て い る 。 ox-kaiC株においてPkaiBC::luxABの生物発光リズムを観察した結果,リズムが長 周 期 化 す る こ と が 明 ら か と な り , こ れ も 先 行 研 究 の 結 果 と 一 致 し た (図 4;
Nakajima et al., 2010)。
以上の結果から,明期中での時計遺伝子の転写・翻訳フィードバック (KaiC蛋 白質のリズミックな蓄積量の変化) によりKaiC/KaiA比が周期的に変化すること で,夜間における KaiC リン酸化リズムの周期・振幅が調節されていること。そ して,夜間でのKaiCリン酸化リズムの短周期化・低振幅化はKaiC/KaiA比が小 さくなるために起こることが示唆された。
暗期中の KaiC リン酸化リズムの減衰 (短周期化・低振幅化) は位相応答を鋭敏に する
図1B では,主観的夜明けに細胞に対して暗刺激を与えると概日時計の位相シフ トが大きくなり,light onの時刻に位相がそろうようになること,図 2Bではlight onの時刻に位相がそろうような大きな位相シフトが観察されるときには,暗期中 でKaiCリン酸化リズムが低振幅化・短周期化することを示唆した。
次に,暗期中での KaiC リン酸化リズムの低振幅化・短周期化が光に対する位相 応答を鋭敏にしていることを確認した。野生株とox-kaiA株に0,15,20,50 μM の IPTGを加え段階的に KaiAの誘導をおこなった株を用意し, 2回の明暗サイ クルに同調後,明期12時間目から様々な長さの暗刺激を与えた。その結果,KaiA の誘導に従って暗刺激に対する位相応答が大きくなり,明期開始時刻に位相がそ ろう傾向が見られた (図 5A)。このとき,暗期開始時刻の KaiC/KaiA 比は KaiA の誘導に伴って小さくなることも確認した (図5B)。興味深いことに,暗刺激に対 する位相シフト量はKaiC/KaiA比の変化に応じて大きくなることが明らかとなっ た。図 5Cに 6時間暗刺激による位相シフトとKaiC/KaiA比の変化の関係を,図 5Dに18時間暗刺激による位相シフトとKaiC/KaiA比の変化の関係を示した。暗 期におけるリン酸化リズムの減衰が ox-kaiA 株で見られるかを確認するため,
ox-kaiA株に20 μM のIPTGを加え,2回の明暗サイクル後,明期12時間目か
図4. 時計蛋白質 KaiC量を増加させた際に見られる概日リズムの周期変化
細胞を2回の明暗サイクルに同調させ,連続明条件下でkaiC過剰発現株のPkaiBC::luxABリズ ムを測定した。
(A) kaiC過剰発現株 (ox-kaiC; IPTG 0 μM)と野生株の発光測定結果を示した。
(B) (A) の結果の最大値を1としてそれぞれ標準化した結果を示した。
ら連続暗条件下に細胞を以降させ,KaiCリン酸化リズムを解析した (図 5E)。そ の結果,20 μM でIPTG誘導をかけたox-kaiA株の暗期中でのリン酸化リズムは 野生株と比べ,低振幅化・短周期化することが明らかとなった。以上の結果より,
暗期中の KaiC リン酸化リズムの減衰が光に対する位相応答を鋭敏にすることが 強く示唆された。
明刺激は減衰した KaiC リン酸化リズムの急激な脱リン酸化を促進し位相シフト を引き起こす
暗刺激による位相シフト完了まで,細胞は「暗期開始」と「明期開始」の 2 つ の環境変化を経験する。暗期開始の刺激は概日時計の位相シフト (リセット) に非 常に重要ではあるが,図2B を見ると暗期でのリン酸化リズムの位相シフト量は細 胞の暗期移行時刻によらず明期のコントロールと比べてそれほど大きくない。つ まり暗期開始の刺激のみでは位相シフトは完了しない。よって,私は KaiC リン 酸化リズムが暗期で減衰しているときには,明期開始に伴うリセット刺激の効果 をKaiCリン酸化リズムがロバストな状態に比べて強く受けると考えた。図1Bを 見ると,主観的夜明け時に暗刺激を与えた場合,大きな位相シフトが観察される。
よって,主観的夜明け時に暗刺激を与え,その後細胞を再び明期へ移行した際に 概日時計に変化が現れると考え,明期開始後のKaiCリン酸化リズムを解析した。
細胞を2回の明暗サイクルに同調させ,明期24時間 (主観的夜明け) 経過後 9時 間の暗刺激を与えた後,再度明期に移行した際の KaiC リン酸化リズムを解析し た (図 6A)。解析の結果,明期開始後4時間以内に急激な KaiCの脱リン酸化が見 られ,KaiCリン酸化リズムは暗刺激をかけていないコントロールと比較して 3時 間の遅れが観察された。図 1Bの結果からkaiBCのプロモーター活性も同じ刺激
図5. 細胞内KaiC/KaiA量比の減少に伴う位相変位の増加
(A) 野生株とkaiA過剰発現株 (ox-kaiA; 0, 15, 20, 50 μM)における暗期延長実験結果。生物 発光レポーター株を用いて解析を行った。図中のプロットはピークタイムを示す (n = 4)。
(B) 野生株とkaiA過剰発現株 (ox-kaiA; 0, 15, 20 μM)における明期12時間目のKaiC/KaiA 蛋白質量比解析結果。値は野生株の量を1として標準化した。
(C, D) KaiC/KaiA蛋白質量比と位相シフト量の関係。横軸にKaiC/KaiA蛋白質量比,縦軸に 位相シフト量をプロットした (n = 4)。6時間暗刺激 (D)と18時間暗刺激 (F)の結果を示す。
(E) 野生株と20 μM IPTGを加えたkaiA過剰発現株の連続暗条件下でのKaiCリン酸化リズム
でコントロールと比べて3時間の遅れ (位相シフト) が起こり,リン酸化リズムの 位相シフトと一致した。よって,KaiCリン酸化リズムが暗期で減衰しているとき には,明期開始後に急激な脱リン酸化が観察され,その結果位相シフトが起こる ことが示唆された。
次に,KaiCリン酸化リズムが暗期中でロバストな場合の明刺激後のKaiCリン 酸化の状態変化を観察した。細胞を明暗サイクルに同調後,明期 12 時間目から 21時間の暗刺激を与えた後に KaiC リン酸化リズムを解析した。その結果,明期 24時間目から暗刺激を与えたときに,明刺激後に見られた KaiC の急激な脱リン 酸化は観測されなかった (図6B)。よって暗期中のKaiCリン酸化リズムがロバス トな状態のときには,明刺激を与えても KaiC の急激な脱リン酸化が観測されな いことが明らかとなった。
次に,暗刺激後の明期開始直後に見られる KaiC 脱リン酸化の位相依存性を検 討した。図 6Aでは,暗期中のKaiCリン酸化リズムがリン酸化フェーズの際に明 刺激を与えたが,脱リン酸化フェーズに明刺激を与えた場合も,明期開始後に KaiCリン酸化リズムの急激な脱リン酸化が観測されるかを検討した。細胞を明暗 サイクルに同調後,明期24時間目から15時間の暗刺激を与え,その後再び細胞 を明期に移行した。このとき,暗期における KaiC リン酸化リズムの位相は脱リ ン酸化フェーズである。検討の結果,明期開始 4 時間以内に急激な脱リン酸化が 観察された (図 6C)。よって,暗期中のKaiCリン酸化リズムに減衰が見られる際 には,明期開始後の急激なKaiCの脱リン酸化は,暗期のKaiCリン酸化リズムの 位相によらず見られることが示唆された。一方で,明期 12時間目から12時時間 の暗刺激を与えた場合は,明期開始後の KaiC の急激な脱リン酸化は観察されな かった (図6D)。よって暗期中のKaiCリン酸化リズムに減衰が見られない場合は,
暗期中の位相によらず,明期開始後に急激なKaiCの脱リン酸化が起こらないこ
図6. 明刺激が誘導するKaiCの脱リン酸化
細胞を2回の明暗サイクルに同調後,細胞に暗刺激を与え,暗刺激後のKaiCリン酸化リズムを 解析した。
(A) 明期24時間目に9時間の暗刺激をかけた後のKaiCリン酸化リズム解析結果 (白, LLコ ントロール; 青,DDコントロール; マゼンダ,暗刺激後のLL)。
(B) 明期12時間目から21時間の暗刺激を与えた後のKaiCリン酸化リズム解析結果。
(C) 明期24時間目から15時間暗刺激を与えた後のKaiCリン酸化解析結果。
(D) 明期12時間目から12時間の暗刺激を与えた後のKaiCリン酸化リズムの解析結果。
(E) 野生株とkaiA過剰発現株 (ox-kaiA; 0, 15, 20 μM)における,明期12時間目から18時間 暗刺激を与えた後のKaiCリン酸化状態の解析結果。それぞれの株で刺激後明期2時間目の KaiCリン酸化状態を,細胞を明期に移行していない同じ時間帯 (DD20 h)のサンプルで標準化
とが示唆された。
次に,刺激を与える時間と長さを固定して暗期中の KaiC のリン酸化リズムの み減衰させていった際に,明期開始後の急激なKaiCの脱リン酸化がKaiCリン酸 化リズムの減衰に伴って観察されるかを確認した。野生株とox-kaiA株に0,15,
20,50 μM の IPTGを加え段階的に誘導をおこなった株を用意し,明暗サイクル
同調後の明期12時間目から18時間の暗刺激を与え,その後再度明期に細胞を移 行した際の明期 2 時間目の KaiC 脱リン酸化率を測定した。測定結果は,明刺激 をかけていない暗期の KaiC リン酸化リズムをコントロールとして変化量を計算 し,野生株の解析結果が100%となるように標準化した。標準化した結果を図 6E に示した。この結果から,KaiCリン酸化リズムが暗期で減衰するに伴って,明期 での KaiC の脱リン酸化効果が大きくなることが明らかとなった。よって,暗期 中の KaiC リン酸化リズムの減衰は,暗刺激後の明刺激の効果を受けやすくする ことが強く示唆された。
考察
転写翻訳フィードバックを介した KaiC/KaiA 比の低下により KaiABC 振動子の 脆弱化が起こり,位相シフトが促進される
以 上 の 結 果 よ り , 明 期 に お け る 時 計 遺 伝 子 の 転 写 ・ 翻 訳 フ ィ ー ド バ ッ ク が
KaiC/KaiA比の低下を引き起こし,暗期中のKaiABC振動子を脆弱化させること
により概日時計の位相シフトを促進させるモデルを提案する。まず,連続明条件 下では時計遺伝子の転写・翻訳フィードバックの効果,特に KaiC 蛋白質の蓄積 リズムにより,時計蛋白質 KaiCと KaiAの細胞内量比 (KaiC/KaiA比)が変化す る 。 こ れ に よ り 主 観 的 黄 昏 で は KaiC/KaiA 比 が 大 き く , 主 観 的 夜 明 け で
KaiC/KaiA 比が小さくなる。連続明期の各時刻から細胞が暗刺激を受けると,暗
刺激中は時計蛋白質が安定化することによりKaiC/KaiA比が暗期開始時刻で固定 される。主観的夜明けから暗刺激が与えられた場合はKaiC/KaiA比が小さくなり,
それに伴って暗期中のKaiCリン酸化リズムが減衰する。減衰した KaiCリン酸化 リズムに明刺激が与えられると KaiC の急激な脱リン酸化が起こり,その結果 KaiCのリン酸化リズムの位相が大きくシフトする (図 7)。
真核生物における概日時計の脆弱化と位相シフトの関係
真核生物においても,概日時計の脆弱化と位相シフトの関係が報告されている。
例えばショウジョウバエでは,時計遺伝子per とtimが概日時計の発振機構を担っ ているが,ショウジョウバエのtimUL変異体は,野生株と比べて時計遺伝子per と timのmRNA蓄積リズムの振幅が小さくなることが観察されている。このとき,野 生株は10分の光パルスに対する位相シフト量が4時間以内であるのに対し,timUL 変異体は10時間の位相前進と6時間の位相後退が見られ,概日時計のペースメーカ ーの振幅が小さくなることによって光に対する位相応答が鋭敏になっていること
図7. 転写・翻訳フィードバックによるKaiABC振動子の光応答性調節機構
連続明期中では,KaiC蛋白質蓄積リズムによりKaiC/KaiA比がKaiCの蓄積リズムに従って調 節される。連続明期中に暗刺激が与えられると,KaiC/KaiA量比に従ってKaiABC振動子の周 期や振幅が変化する。主観的明期から暗刺激を与えられた場合は,KaiABC振動子のリン酸化 リズムが減衰する。その後,再び明期に細胞を移行すると,暗期中のKaiCリン酸化リズムに減 衰が見られた場合は位相が明期開始時に固定され,大きな位相シフトが観察される。一方で,暗 期中にKaiCのリン酸化リズムの減衰が見られない場合は,位相変化は小さく,暗刺激前の位相 が暗刺激後も維持される。