Introduction
There has been a recent focus on plant biomass as a source of commercial biomaterials as both energy resources and valuable chemical products. However, because we also consume plant biomass as food, it is necessary to avoid competition between their industrial and agricultural utilization. One of the strategies to solve this problem is the utilization of lands that are currently suboptimal for agriculture, including regions of alkaline calcareous soil, which occupy up to 30% of the cultivated soil worldwide (Kobayashi and Nishizawa, 2012).
Although iron is the fourth most abundant element in the geosphere, it forms ferric hydroxide in alkaline soil, and plants cannot absorb this insoluble form of iron (Robin et al., 2008; Skinner, 1979). The plant strategies invoked for utilizing this insoluble form of iron have been extensively studied (Curie et
al., 2009; Kobayashi and Nishizawa, 2012), and transgenic rice
plants that are resistant to alkaline soil have been successfully generated on the basis of clarified molecular mechanisms against iron deficiency (Kobayashi and Nishizawa, 2012; Takahashi et
al., 2001). Another strategy against iron deficiency employed by
plants is the more efficient utilization of limited iron. For example, Cu/Zn superoxide dismutase (SOD), Mn SOD, and Fe SOD are modified in response to the availability of the corresponding metal in chloroplasts (Page et al., 2012; Yamasaki
et al., 2007). However, it remains unclear how limited iron is
utilized in chloroplasts under iron-limited conditions. Because iron is essential for photosynthetic electron transport and ROS scavenging, it is important for plants to optimize their chloroplast function to the iron conditions. Plants are likely to regulate their chloroplast function prior to the occurrence of the visible symptom of chlorosis, yet our knowledge is still limited with regard to the regulation of photosynthesis, especially in higher plants.
Chlorophyll fluorescent analysis is widely used for monitoring photosynthetic electron transport in intact leaves (Baker, 2008). The electron transport rate (ETR) is a chlorophyll fluorescence parameter that represents the rate of electron transport through photosystem II (PSII), and the non-photochemical quenching (NPQ) of chlorophyll fluorescence largely depends on the safe dissipation of absorbed light energy as heat from PSII, a process that is induced by acidification of the thylakoid lumen (Krause and Weis, 1991). The 1-qL parameter (qL means the fraction of open PSII centers) represents the redox state of the plastoquinone pool (Miyake et al., 2009). From these parameters, the efficiency of light energy utilization can be estimated in leaves, and we can monitor early responses and/or damage to chloroplast function using chlorophyll fluorescence analysis. As a first step, we evaluated the conditions of iron deficiency and found that a disruption of photosynthetic electron transport was earlier than
Analysis of photosynthetic electron transport in Arabidopsis thaliana
under iron deficiency
Ryoichi Araki and Toshiharu Shikanai
Graduate School of Science, Kyoto University(Kitashirakawa, Sakyo, Kyoto 606−8502, Japan)
Summary: Iron is an essential nutrient for plants, and intensive studies have focused on plant responses to iron deficiency. In the field, iron availability often limits the productivity of crops, particularly in alkaline calcareous soil with insoluble iron. Iron is essential for chloroplasts, as a cofactor of many proteins involved in electron transport and reactive oxygen species (ROS) scavenging. Because chloroplasts are the major site of ROS generation, it is important for plants to maintain proper chloroplast function under iron deficiency to avoid photodamage. To study the primary damage to the photosynthetic apparatus by iron deficiency in Arabidopsis
thaliana, we took advantage of chlorophyll fluorescence analysis, a nondestructive method of monitoring
photosynthetic electron transport in leaves. Chlorosis was observed, and photosystem II (PSII) was severely photodamaged in seedlings grown under severe iron-deficient conditions. In contrast, mild iron deficiency decreased the non-photochemical quenching of chlorophyll fluorescence, which represents the dissipation of absorbed light energy from PSII prior to PSII photodamage. Although transferring seedlings from iron-supplemented to iron-deficient conditions is often used to study plant responses to iron deficiency, photosynthetic electron transport was disturbed before visible symptoms of iron deficiency were observed.
Materials and methods
Growth conditions
Sterilized seeds of Arabidopsis thaliana (Col-0) were stored in water at 4℃ for 2 or 3 days and then sown on MS medium and incubated in a growth chamber (LH-350S, NK system, Japan) with white fluorescent lights (70 µmol photons m−2 s−1 at
23℃ ) under a photoperiod of 16-h light / 8-h dark. After 1 or 2 week(s), the seedlings were transferred to MS medium plus iron as an iron-sufficient condition (+Fe) or iron-free MS medium as an iron-deficient condition (−Fe). We exposed seedlings to two types of iron-deficient conditions. In the severe −Fe conditions, seedlings cultured on the +Fe medium for 7 days were transferred to the −Fe medium and then incubated for 7 days. In the mild −Fe conditions, seedlings grown under the +Fe for 14 days were cultured on the −Fe medium for 6 days. The control seedlings were continuously cultured on the +Fe medium for the corresponding periods. These seedlings were subjected to the measurement of chlorophyll fluorescence and SPAD values. The MS medium was supplemented with 5 µM of CuSO4 to avoid a
possible cross-talk with Cu deficiency (Yamasaki et al., 2007). Iron was supplied in the form of FeSO4・7H2O chelated by Na2
-EDTA (Murashige and Skoog, 1962).
Chlorophyll fluorescence analysis
Seedlings transferred to the +Fe or −Fe medium were dark-adapted at least for 20 min prior to the chlorophyll fluorescence analysis using a MINI-PAM portable chlorophyll fluorometer (Walz, Germany). The ETR, NPQ, and 1-qL parameters were calculated as follows:
ETR = (Fm'−F)/Fm' × photon flux density
(µmol photons m−2 sec−1),
where Fm' is the maximal fluorescence from a light-adapted leaf
and F is the steady-state fluorescence level in the light; NPQ = Fm/Fm'−1,
where Fm is the maximum fluorescence (Bilger and Björkman,
1994); and
1−qL = 1−((Fm'−F)/Fm'/(1−(Fm'−F))/Fm') ×
(1−(Fv/Fm)/ (Fv/Fm)) × (NPQ + 1)),
where Fv = Fm−F (Miyake et al., 2009).
Estimation of chlorophyll contents
SPAD values were measured by SPAD 502 chlorophyll meter (Konica Minolta, Japan). The chlorophyll contents were calculated by following equation (Ling et al., 2011):
Chl content (nmol chl / mg fresh wt.) = 0.0007 × SPAD2 + 0.0230 × SPAD + 0.0544
Results and discussion
Optimization of iron-deficient conditions for chlorophyll fluorescence analysis
To evaluate the effects of iron deficiency on the photosynthetic electron transport, plants grown under severe or mild −Fe were used for a chlorophyll fluorescence analysis. Seedlings exposed to severe −Fe exhibited a visible symptom of the chlorophyll content. Consistently, chlorophyll contents were significantly lower in the severe −Fe plants than those in the +Fe plants (Table 1).The values of Fv/Fm, representing the maximum
quantum yield of PSII, were 0.59 ± 0.17, whereas the values were 0.71 ± 0.03 in the control plants grown under +Fe for 14 days. These results indicate that PSII was photodamaged by growth under the severe −Fe condition. The light-intensity dependence of the chlorophyll fluorescent parameters was also impacted by the severe −Fe (Fig. 1). ETR was saturated at a lower level, and 1-qL was higher in the severe −Fe plants than in the +Fe plants at a low light intensity. The magnitude of NPQ was also lower in the severe −Fe plants than the +Fe plants. Because chlorophyll fluorescence analyses depend on PSII activity, we cannot assess the state of electron transport in detail on the basis of chlorophyll fluorescence parameters in plants with considerably lower Fv/Fm values. Rather, the plants must be
analyzed at earlier stages to examine the primary response of or damage to photosynthesis by iron deficiency.
The plants exposed to the mild −Fe conditions for 6 days exhibited levels of Fv/Fm that were 0.77 ± 0.01, indicating that
PSII was not seriously damaged at this stage, although the chlorophyll contents was lower in mild −Fe plants than in +Fe plants (Table 1). We considered that this condition is optimal to evaluate the effect of iron deficiency on photosynthetic electron transport.
Early response or damage of photosynthetic electron transport to iron deficiency
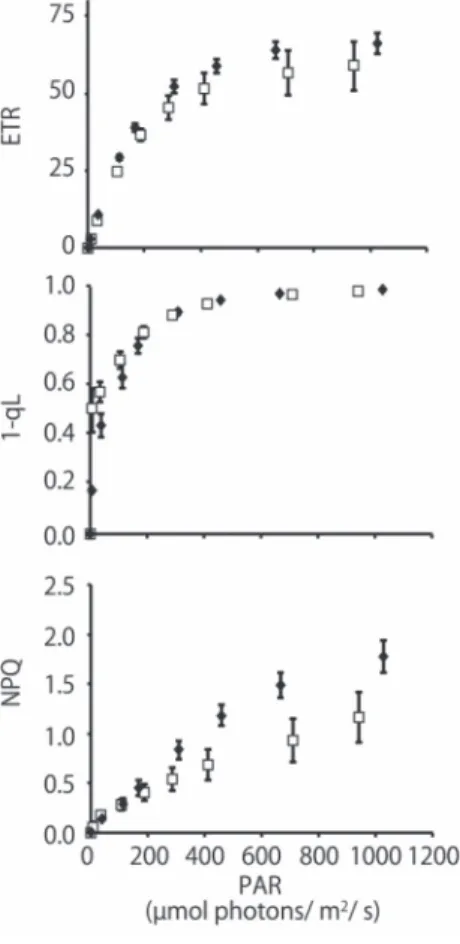
ETR was not significantly different between the mild −Fe and +Fe plants, whereas 1-qL was slightly higher in the mild −Fe plants at a low light intensity (Fig.2). These results suggested that the plastoquinone pool was slightly more reduced by electrons. This level of iron deficiency may somehow restrict electron transport downstream of QA of PSII at a low light
intensity. NPQ was also considerably lower in the mild −Fe
mainly consists of a qE (energy-dependent quenching) component, depending on ∆pH formation. Lumen acidification, which is regulated by electron transport and modulated by NPQ mechanisms (Foyer et al., 2012), may be restricted by iron deficiency at high light intensity in −Fe plants. It is also possible that the NPQ machinery is less sensitive to lumen acidification under iron deficiency. Although no symptom of iron deficiency was observed in the maximum activity of PSII in the plants exposed to mild −Fe, photosynthetic electron transport was already affected, particularly with regard to NPQ.
Photosynthetic electron transport is mediated by the protein complexes of photosystem I (PSI), PSII, and Cytochrome b6f,
which contain 12, 3, and 5 iron atoms, respectively (Behrenfeld and Milligan, 2013; Nouet et al., 2011). In addition to the main complexes, ferredoxin, which accepts electrons from PSI, is also an iron-containing protein. Cyclic electron transport (PSI) generates ∆pH, which is consequently is involved in ATP synthesis and NPQ induction, without the accumulation of NADPH (Shikanai, 2007). In flowering plants, PSI cyclic
Fig. 1. Effects of severe iron deficiency on the chlorophyll fluorescence parameters ETR, 1-qL, and NPQ. The chlorophyll fluorescence of seedlings grown on − Fe or +Fe (as a control) medium was measured using MINI-PAM portable chlorophyll fluorometer. The white boxes indicate severe −Fe conditions; the black diamonds indicate +Fe conditions. PAR = photosynthetically active radiation. Error bars represent SDs (n = 5−7).
Fig. 2. Effects of mild iron deficiency on the chlorophyll fluorescence parameters, ETR, 1-qL, and NPQ. The chlorophyll fluorescence of seedlings grown on −Fe or +Fe (as a control) medium was measured using a MINI-PAM. The white boxes indicate mild −Fe conditions; the black diamonds indicate +Fe conditions. PAR = photosynthetically
electron transport is mediated by both PGR5 (PROTON GRADIENT REGULATION 5)/PGRL1 (PGR5-LIKE PHOTOSYNTHETIC PHENOTYPE 1) (DalCorso et al., 2008) and chloroplast NADH dehydrogenase-like complexes (Munekage et al., 2004; Peng et al., 2009). Machinery of PSI cyclic electron transport also includes iron atoms, implying that iron deficiency may directly affect the levels of photosynthetic machinery. Interestingly, rice and barley grown under the growth chamber conditions show the opposite response to iron deficiency (Saito et al., 2010). In rice grown under iron deficiency, ETR and NPQ were decreased compared to growth in the presence of sufficient iron, consistent with our observations, whereas NPQ induction in barley was accelerated by iron deficiency, even though iron deficiency scarcely affected ETR. The authors suggested that several light-harvesting chlorophyll a/b-binding proteins of PSII (LHCII) (Lhcb1s) are specifically expressed under iron deficiency and may induce enhanced NPQ as a protective mechanism of the photosynthetic machinery. Although enhancing NPQ is the strategy employed by barley to prevent photodamage under iron deficiency, we observed the opposite in Arabidopsis (Fig. 2). It is most likely that an early defect in photosynthesis caused by iron deficiency in rice and Arabidopsis was monitored rather than a protective response. It is possible that our conditions of iron deficiency were too severe for monitoring a plant response for adaptation. In Chlamydomonas
reinhardtii, iron deficiency resulted in a reduction in the levels of
PSI, PSII, and Cytochrome b6f and remodeling of the antenna
complexes to establish a new steady state of the electron transfer chain (Moseley et al., 2002). Thus, it remains unclear how the balance of electron transport machinery is maintained by using a limited amount of iron in chloroplasts. To answer this question, our strategy is to clarify the primary defect in iron deficiency in chloroplasts, which most likely results in low NPQ induction and consequent severe photodamage of PSII, as observed in the plants exposed to the severe iron-deficient condition.
Acknowledgement
This work was supported partly by Grants-in-Aid from the NC-CARP project, the Ministry of Education, Culture, Sports, Science and Technology, to TS.
References
Baker, N.R. (2008) Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 59: 89−113. Behrenfeld, M.J. and A.J. Milligan (2013) Photophysiological
expressions of iron stress in phytoplankton. Annu. Rev. Mar. Sci. 5: 217−246.
Bilger, W. and O. Björkman (1994) Relationships among
violaxanthin deepoxidation, thylakoid membrane conformation, and nonphotochemical chlorophyll fluorescence quenching in leaves of cotton (Gossypium hirsutum L.). Planta 193: 238−246.
Curie, C., G. Cassin, D. Couch, F. Divol, K. Higuchi, M. Le Jean, J. Misson, A. Schikora, P. Czernic and S. Mari (2009) Metal movement within the plant: contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 103: 1−11.
DalCorso, G., P. Pesaresi, S. Masiero, E. Aseeva, D. Schünemann, G. Finazzi, P. Joliot, R. Barbato and D. Leister (2008) A complex containing PGRL1 and PGR5 is involved in the switch between linear and cyclic electron flow in
Arabidopsis. Cell 132: 273−285.
Foyer, C.H., J. Neukermans, G. Queval, G. Noctor and J. Harbinson (2012) Photosynthetic control of electron transport and the regulation of gene expression. J. Exp. Bot. 63: 1637− 1661.
Kobayashi, T. and N.K. Nishizawa (2012) Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 63: 131−152.
Krause, G.H. and E. Weis (1991) Chlorophyll fluorescence and photosynthesis: the basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 42: 313−349.
Ling, Q., W. Huang and P. Jarvis (2011) Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis
thaliana. Photosynth. Res. 107: 209−214.
Miyake, C., K. Amako, N. Shiraishi and T. Sugimoto (2009) Acclimation of tobacco leaves to high light intensity drives the plastoquinone oxidation system−relationship among the fraction of open PSII centers, non-photochemical quenching of Chl fluorescence and the maximum quantum yield of PSII in the dark. Plant Cell Physiol. 50: 730−743.
Moseley, J.L., T. Allinger, S. Herzog, P. Hoerth, E. Wehinger, S. Merchant and M. Hippler (2002) Adaptation to Fe-deficiency requires remodeling of the photosynthetic apparatus. EMBO J. 21: 6709−6720.
Munekage, Y., M. Hashimoto, C. Miyake, K.-I. Tomizawa, T. Endo, M. Tasaka and T. Shikanai (2004) Cyclic electron flow around photosystem I is essential for photosynthesis. Nature 429: 579−582.
Murashige, T. and F. Skoog (1962) A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15: 473−497.
Nouet, C., P. Motte and M. Hanikenne (2011) Chloroplastic and mitochondrial metal homeostasis. Trend. Plant Sci. 16: 395− 404.
Page, M.D., M.D. Allen, J.Kropat, E.I. Urzica, S.J. Karpowicz, S.I. Hsieh, J.A. Loo and S.S. Merchant (2012) Fe sparing and Fe recycling contribute to increased superoxide dismutase capacity in iron-starved Chlamydomonas reinhardtii. Plant
Cell 24: 2649−2665.
Peng, L., Y. Fukao, M. Fujiwara, T. Takami and T. Shikanai (2009) Efficient operation of NAD(P)H dehydrogenase requires supercomplex formation with photosystem I via minor LHCI in Arabidopsis. Plant Cell 21: 3623−3640. Robin, A., G. Vansuyt, P. Hinsinger, J.M. Meyer, J.F. Briat and P.
Lemanceau (2008) Chapter 4 Iron dynamics in the rhizosphere: consequences for plant health and nutrition. In "Advances in Agronomy" Donald, L.S. (eds), Academic Press. 99: 183−225.
Saito, A., T. Iino, K. Sonoike, E. Miwa and K. Higuchi (2010) Remodeling of the major light-harvesting antenna protein of PSII protects the young leaves of barley (Hordeum vulgare L.) from photoinhibition under prolonged iron deficiency. Plant
Cell Physiol. 51: 2013−2030.
Shikanai, T. (2007) Cyclic electron transport around photosystem I: genetic approaches. Annu. Rev. Plant Biol. 58: 199−217. Skinner, B.J. (1979) Earth resources. Proc. Natl. Acad. Sci. USA
76: 4212−4217.
Takahashi, M., H. Nakanishi, S. Kawasaki, N.K. Nishizawa and S. Mori (2001) Enhanced tolerance of rice to low iron availability in alkaline soils using barley nicotianamine aminotransferase genes. Nat. Biotech. 19: 466−469.
Yamasaki, H., S.E. Abdel-Ghany, C.M. Cohu, Y. Kobayashi, T. Shikanai and M. Pilon (2007) Regulation of copper homeostasis by micro-RNA in Arabidopsis. J. Biol. Chem. 282: 16369−16378.