15
原 著 論 文
徳之島と奄美大島に生息するコウモリ類の生態学的研究,
特にリュウキュウテングコウモリ
Murina ryukyuana について
船越 公威
1,山下 啓
2,北之口卓志
1,田中 広音
1,大坪 将平
1,大平 理紗
1,
内原 愛美
1,大澤 達也
1,渡辺 弘太
1,永山 翼
1,亘 悠哉
3,南 尚志
4 1鹿児島国際大学国際文化学部生物学研究室 2カエルPROJECT 3国立研究開発法人森林研究・整備機構森林総合研究所野生動物研究領域 4鹿児島県森林組合連合会 摘 要 徳之島と奄美大島のコウモリ類について,カスミ網, ハープトラップおよびアカメガシワトラップによる捕獲, 洞窟探査,音声録音や発信機装着個体の追跡によって調 査し,以下の 7 種と未同定の種が確認された.(1)リュウ キュウテングコウモリMurina ryukyuana は両島の常緑広 葉樹林に生息しているが,体のサイズは徳之島の方が大 きかった.本種は日毎にねぐらを変え,発信機装着個体 の追跡からねぐら間の移動距離は 25 ~ 178 m であった. ねぐら場所として,比較的乾燥した枯葉,群葉および樹 洞が利用されていた.出産・哺育期は 5 月~ 7 月,母子 集団は 11 月まで持続し母子最大 16 頭であった.独立後 は幼獣雌雄ともに単独生活に入った.成獣雄は秋季にな わばりを持つことが示唆された.(2)ヤンバルホオヒゲ コウモリMyotis yanbarensisは両島の限られた常緑広葉樹 林に生息しているが,徳之島における捕獲率は非常に低 く個体数が非常に少ないことが示唆された.(3)オリイ コキクガシラコウモリRhinolophus cornutus orii は両島に 点在する自然洞や廃坑をねぐらとして利用していた.こ の種の音声について徳之島のピーク周波数(111.3 kHz) は奄美大島(107.7 kHz)より高かった.(4)リュウキュ ウユビナガコウモリMiniopterus fuscus の飛翔域は各島 内の広範囲に及んでいた.(5)モモジロコウモリMyotis macrodactylus は河川域で飛翔していたが,両島ではきわ めて少なかった.(6)アブラコウモリPipistrellus abramus は両島の市街地に生息することが再確認された.(7)ク ビワオオコウモリPteropus dasymallus は徳之島で発見さ れた.(8)Tadarida sp. の生息が両島で確認され,徳之島 では初めての確認であった.この種の採餌空間は各島内 の広域に及んでおり,奄美大島の海岸 2 ヵ所でねぐら場 所が確認された.音声解析から,徳之島では既知種とは 異なる種が少なくとも 2 種,奄美大島では既知種と異な る種が少なくとも 2 種生息している可能性が示唆された. は じ め に 徳之島と奄美大島は南西諸島の亜熱帯地域に位置して いる(Fig. 1).中琉球に属する両島は,後期中新世から 更新世初期(約 1,000 万年前~約 200 万年前)までには 大陸から分離し,島嶼に取り残された遺存固有種を含む 特有の動物相を形成している(山田 2017).徳之島に は, こ れ ま で の 記 録 で ク ビ ワ オ オ コ ウ モ リPteropus dasymallus(船越 2017a),洞窟性コウモリとしてオリイ コキクガシラコウモリRhinolophus cornutus orii(以下, コ キ ク ガ シ ラ コ ウ モ リ ), モ モ ジ ロ コ ウ モ リMyotis macrodactylus, リ ュ ウ キ ュ ウ ユ ビ ナ ガ コ ウ モ リ Miniopterus fuscus,森林性コウモリとしてヤンバルホオ ヒゲコウモリMyotis yanbarensis とリュウキュウテング コウモリMurina ryukyuana,そして住家性のコウモリと してアブラコウモリPipistrellus abramus の計 7 種のコウ モリが生息していることが報告されている(Yoshiyuki 1989;前田ほか 2001).一方,奄美大島でも,上記の ク ビ ワ オ オ コ ウ モ リ を 除 く 6 種 が 確 認 さ れ て お り (Yoshiyuki 1989;澤田 1994;前田ほか 2002),スミイロ オヒキコウモリTadarida latouchei(Yoshiyuki et al. 1989) を加えて計 7 種のコウモリが生息している.特に森林性コウモリのヤンバルホオヒゲコウモリと
哺乳類科学 59(1):15-36,2019 ©日本哺乳類学会
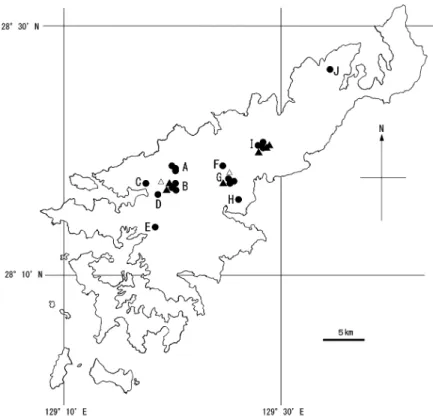
リュウキュウテングコウモリは,沖縄島北部のヤンバル の森林で 1996 年に同時に発見,新種記録され(Maeda and Matsumura 1998),その後,徳之島(前田 2000;前 田ほか 2001)と奄美大島(前田ほか 2002)で生息が確 認されている.両種は日本の固有種で絶滅危惧種にラン クされており(Kawai 2015b,c)リュウキュウテングコ ウモリについては徳之島における一部の生態や分布につ いて報告(船越ほか 2013a;亘・船越 2013)されている が,ヤンバルホオヒゲコウモリについては不明な点が多 く両島で本格的な調査がなされていない.本研究では, 各種の分布や生息状況を詳細に把握するとともに,両島 の種構成や前腕長,体重,音声を比較し,両島の気候や 自然環境の違いがコウモリの生息状況にどのように反映 されているのか検討した.特に,これまで不明な点が多 かったリュウキュウテングコウモリの分布や生態につい て詳細に論じた.また,音声解析を行い,既知の種以外 の種が生息している可能性を論じた. 調査地および方法 1.徳之島 徳之島は奄美大島から南西方向の北緯 28° 以南に位置 し,面積は 248 km2である(Fig. 1).森林性コウモリ類 の主な調査地点は,北部の天城岳の周辺地域(以下, A),中部の三京(以下,B),井ノ川岳(以下,C),犬 田布岳(以下,F),剥岳(以下,D),神嶺ダム(以下, E)地域および南部の東部ダム周辺地域(以下,G)で ある(Fig. 2).これらの地域には,ブナ科のスダジイ Castanopsis sieboldii とアラカシ Quercus glauca が優占す る二次林がみられるほか,谷や斜面の下の方のやや湿潤 な所にはオキナワウラジロガシCyclobalanopsis miyagii が 優 占 す る 一 次 林( 老 齢 林 ) が み ら れ( 大 野・ 寺 田 1996),巨木も混在している.こうした自然植生を有す る地域は,徳之島において上記の地域に限られている. 予備調査で,上記のブナ科の常緑広葉樹林(スダジイ, アラカシ,オキナワウラジロガシなど)が優占する一次 林と二次林ではリュウキュウテングコウモリの生息が確 認されている.一方,ブナ科以外の樹種(ホルトノキ Elaeocarpus sylvestris,モクタチバナ Ardisia sieboldii お よびリュウキュウマツPinus luchuensis など)が優占す る若齢林では,リュウキュウテングコウモリは捕獲でき ず,音声も録音できなかった.そこで,本調査では,ブ ナ科の常緑広葉樹が優占する二次林(高木層の樹齢 20 ~ 50 年)を調査対象地域とした.奄美大島における 調査対象地域の林況も同様である.
Fig. 1. Map of Tokunoshima Island and Amami-Oshima Island.
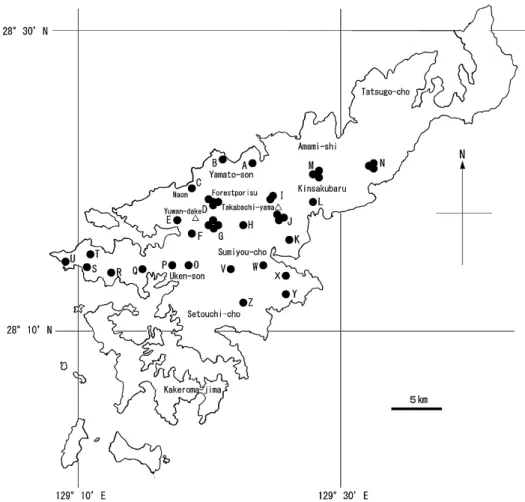
Fig. 2. Locations of the study areas (A–G) on Tokunoshima Island
and the distributions of Murina ryukyuana (●) and Myotis
徳之島と奄美大島のコウモリ類の生態 17
パ ル ス 終 わ り の 周 波 数( 終 部 周 波 数:EF, end fre-quency),最大振幅時の周波数(ピーク周波数:PF, peak frequency or frequency of maximum energy of the pulse)及 びパルスの長さ(持続時間:D, call duration)である. 飛翔時の音声パルスの測定値や形状から船越(2010)の それらと照合し,また飛翔個体のサイズや飛翔パターン から種を判定した.また,日没後数時間の音声記録から 各種の飛翔個体数をカウントした. リュウキュウテングコウモリの捕獲調査では,カスミ 網による捕獲に加えて,船越ほか(2009)に従い,アカ メガシワMallotus japonicus の枝を数本束ねたアカメガ シワトラップを現地で作成して,それらを約 20 ~ 50 m 間隔で林道から約 5 ~ 10 m 入った森林低層部の地上高 約 2 m と森林中層部の地上高約 5 m の横枝に吊るした. 設置されたトラップを昼間にチェックし,ねぐらとして 利用している個体を捕虫網で捕獲して本種の生息域や行 動域を調べた.各地域の調査日とトラップ設置数を Table 1 に記した.延べトラップ数に対する捕獲成功ト 調査は 2001 年,2006 年,2012 年~ 2014 年の 3 月~ 11 月に行った.調査はカスミ網(サイズ:6×4 m と 12 ×4 m を各 1 機併用)による捕獲と合わせてバットディ テクター(コウモリ超音波探知機:Pettersson D1000 bat detector, Pettersson Elektronik AB, Uppsala, Sweden)によ る音声の録音を行った.カスミ網による捕獲では,人工 的に作成された数種コウモリ類の音声をスピーカーで発 し, コ ウ モ リ を 誘 引 す る 装 置Acoustic Lure(Sussex AutoBat, Hill and Greenaway, Brighton, UK)を併用した. 音 声 の 録 音 で は, タ イ ム エ キ ス パ ン ジ ョ ン 式( time-expansion mode:オリジナルの 1/10 の速度に変換)にセッ トし,デジタルメモリーに記録した.また,夜間の音声 を録音するために,超音波無人測定機Pettersson D500X (Pettersson Elektronik AB, Uppsala, Sweden)を設置した.
記録した音声の解析は,Bat Sound 4.14 software(Pettrsson Elektronik AB, Uppsala, Sweden)を使用した.
録音された音声パルスの測定項目のパラメータは,パ ルス始まりの周波数(始部周波数:SF, start frequency),
Table 1. Records of the trapping efficiency and information on Murina ryukyuana captured in the traps made of dead-leaves of Mallotus japonicus
suspended from branches at about 2 m above the ground, on Tokunoshima Island
Date trap-placed (area trapped)*No. of traps capturedDate No. of solitary bats No. of colony and the size (A♀, S♀+S♂) Total no. of traps efficiency (%)Trapping Male Female 2012 8 Aug. 23 (F) 9 Aug. 3 1 0 23 17.4 9 Aug. 56 (F), 28 (D) 10 Aug. 4 1 0 84 6.0 56 (F), 28 (D) 11 Aug. 0 2 0 84 2.4 10 Sept. 18 (F), 15 (D) 11 Sept. 2 0 0 33 6.1 11 Sept. 26 (F), 32 (D)** 12 Sept. 2 1 0 58 5.2 26 (F) 13 Sept. 2 0 0 26 7.7 14 Nov. 25 (D) 17 Nov. 1 0 0 25 4.0 15 Nov. 19 (B) 18 Nov. 1 0 0 19 5.3 2013 7 Nov. 33 (F)** 9 Nov. 1 0 1 (10, 6) 33 6.1 8 Nov. 20 (B), 10 (D) 10 Nov. 3 1 0 30 13.3 33 (F) 11 Nov. 0 2 0 33 6.1 33 (F), 20 (B) 12 Nov. 4 0 0 53 7.5 33 (F), 20 (B) 13 Nov. 2 1 1 (1, 1)*** 53 7.5 33 (F) 14 Nov. 3 0 0 33 9.1
9 Nov. 22 (A), 33 (F) 15 Nov. 4 0 0 55 7.3 2014
25 Aug. 30 (F) 27 Aug. 3 1 0 30 13.3
30 (F)** 29 Aug. 1 0 1 (4, 6) 30 6.7 26 Aug. 30 (F), 10 (B) 31 Aug. 3 1 0 40 10.0 27 Aug. 30 (F), 10 (E), 10 (G) 1 Sept. 4 6 1 (3, 3) 50 22.0
30 (F) 2 Sept. 0 1 0 30 3.3
Total 43 18 4 (18, 16) 822 7.9 (65/822) *, Trap-setting points are as in Fig. 2; A♀, adult female; S♂, subadult male; S♀, subadult female.
**, Traps were set, in addition to the traps (total 10) suspended from a branch about 5 m above the ground. ***, pair (adult male and female).
翔段階(5 月~ 7 月)にあり,指骨関節の未骨化,腹部 皮膚の赤味や背面体毛が黒味を帯びていること,雄では ペニスが細く小さい状態の個体として定義し,亜成獣は, 8 月以降から成獣に達するまでの個体とした.特に,リュ ウキュウテングコウモリについては,夏季の亜成獣雄で は精巣や精巣尾部が体表面に突出せず,成獣雄では萎縮 した精巣や精巣尾部がみられる(船越ほか 2013a)こと で両者を区別した.一方,亜成獣雌では乳頭の突出がみ られず,成獣雌では粒状の乳頭や乳輪がみられることで 両者を区別した.ヤンバルホオヒゲコウモリの雌雄につ いても上記同様に区別した. リュウキュウテングコウモリの個体追跡のため,2013 年 11 月に 6 個体(雄 2,雌 4),2014 年 8 月~ 9 月には 3 個体(雄)に電波発信機(0.25 g,Blackburn Transmitters, Texas, USA)を装着した.発信機装着個体の昼間のねぐ らの特定は,折りたたみ式 3 素子の八木アンテナ(CM- 2H 型,ハムセンター札幌,札幌)と受信機(Yaesu FT- 290mkII, Yaesu,ハムセンター札幌,札幌)を使用した. 受信の際に,受信可能範囲(200 m)や発信機装着地点 から 500 m 前後に移動して異なる 3 方向以上から方探し て入信の方角を決定(三点測位法:Amelon et al. 2009) した.特定された地点はGPS(GPSMAP62, Garmin Ltd. UK)を使用して緯度経度を記録した. 調査期間中の昼間,道路や林道沿いにクワ科(ガジュ マルFicus microcarpa,アコウ F. superba およびイヌビ F. erecta)の結実した樹木があれば,樹下とそれらの周辺 にクビワオオコウモリの食痕があるかどうかを調査し た.また,聞き込み調査も実施した. 2.奄美大島 奄美大島は北緯 28° 以北に位置し,面積は 712 km2で ある(Fig. 1).森林性コウモリ類は 26 地点を調査した (Fig. 4).主要な調査地点は,金作原とその周辺域(M), タカバチ山麓とその周辺域(I,J),湯湾岳麓とその周 辺域(D ~ G),フォレストポリス(D)およびハイビ ス カ ス ロ ー ド(O) で あ っ た(Fig. 4). 調 査 地 域 は, 徳之島の調査地と同じ植生構成の常緑広葉樹の二次林 である.調査は,2001 年~ 2002 年,2006 年,2016 年~ 2017 年の 3 月~ 11 月に行った.調査のたびに,徳之島 の調査と同様に音声を記録し,カスミ網とアカメガシワ トラップに新たに加えたハープトラップ 1 ~ 2 機(サイ ズ:1.5×2 m)による捕獲を行った.各地域のアカメガ シ ワ ト ラ ッ プ 設 置 日 と ア カ メ ガ シ ワ ト ラ ッ プ 数 は Table 2 に記した.その他の地域(Fig. 4E,D,N,O) にも同調査期間に各 20 前後のアカメガシワトラップを ラップ数の割合を「捕獲効率(%)」とした.その際,1 トラップあたりの捕獲頭数は考慮しなかった.一方,カ スミ網による捕獲効率(%)は,延カスミ網設置日に対 する捕獲個体数の割合として算出した. 洞窟性コウモリ類の調査では,北部の松原銅山廃坑 (a),小島洞(f),義名山鍾乳洞(g),面縄鍾乳洞(i), 目手久鍾乳洞(j)および銀竜洞(h)を調査した(Fig. 3). 主 な 調 査 は 2013 年 11 月 7 日 ~ 15 日,2014 年 8 月 25 日~ 9 月 2 日に行った.日中の洞内探査・捕獲に加えて, 夕方洞口で音声録音による生息種の判定と個体数のカウ ントを行い,その翌日の夜間に洞口でカスミ網による捕 獲を試みた. コウモリを捕獲した場合は,各個体の性や繁殖状態を 調べ,電子体重計(TANITA,ハンディミニ 1476,東京: 最小目盛 0.1 g)で体重を測定し,ノギス(中村製作所, KSM-15,東京:最小目盛 0.05 mm)で前腕長を測定し た後,前腕部に標識用リングを着けて捕獲場所で放獣し た.年齢については,指骨関節の化骨の程度により幼獣 か否かを判定し,雄では精巣肥大の有無や長径のサイズ, 精巣尾部の発達の有無,雌では乳頭や乳腺の発達程度や 膣開口の有無をチェックして,幼獣,亜成獣および成獣 の区分を行った.幼獣は,出生から哺育終了後の独立飛
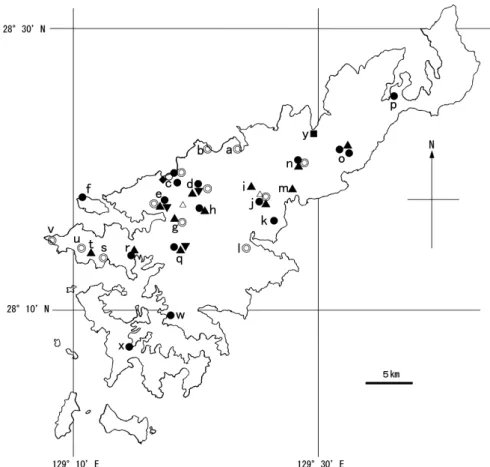
Fig. 3. Locations of the study areas (a–j) on Tokunoshima Island
and the distributions of Rhinolophus cornutus orii (●), Miniopterus
fuscus (▲), Myotis macrodactylus (▼), Pipistrellus abramus (■), Tadarida sp. (◎) , and V1s (○). △, mountain.
徳之島と奄美大島のコウモリ類の生態 19
Fig. 4. Locations of the study areas (A–Z) on Amami-Oshima Island. △, mountain.
Table 2. Records of the trapping efficiency and information on Murina ryukyuana captured in the dead-leaf traps suspended from
branches at about 2 m above the ground, on Amami-Oshima Island
Date trap-placed (area trapped)*No. of traps Date captured No. of solitary bats No. of colony and the size Total no. of traps efficiency (%)Trapping Male Female 2006 25 (G) 16 July 1 0 0 25 4.0 25 (K) 16 July 2 0 0 25 8.0 2016 19 Nov. 20 (J) 21 Nov. 1 0 0 20 5.0 20 (J) 22 Nov. 1 0 0 20 5.0 20 (J) 23 Nov. 1 0 0 20 5.0 20 (J) 24 Nov. 0 1 0 20 5.0 2017 30 Apr. 32 (J)** 4 May 2 0 0 32 6.3 2 May 15 (K) 5 May 1 0 0 15 6.7 18 Aug. 35 (J, I) 22 Aug. 4 1 0 35 14.3 20 Aug. 10 (M) 24 Aug. 1 0 0 10 10.0 35 (J, I) 24 Aug. 5 0 0 35 14.3 Total 19 2 0 257 8.2 (21/257)
*, Trap-setting points are as in Fig. 4.
152 kHz, Philip Blackburn)の電波がおよぶ範囲はきわめ て短距離であることや両島の森林内で使用するため,電 波法に抵触することはない.なお,本調査(徳之島と奄 美大島)は環境省九州地方環境事務所から鳥獣捕獲許可 証(1986 年 度 11-01 号,2001 年度 11-01 号,2002 年度 11-02 号,2006 年度 11-01 号,2008 年度 11-01 号,2010 年度 11-2 号,2012 年度第 11-01 号,2013 年度第 11-67 号, 2014 年度第 11-10 号,2016 年度第 11-9 号,2017 年度第 11-26 号),鹿児島県森林管理署から国有林(保護林) の入林許可証(2001 年度鹿管第 85 号,2002 年度鹿管第 135 号,2006 年度鹿管第 206 号,2012 年度鹿管第 120 号, 2013 度 鹿 管 第 898,899 号,2014 年 度 鹿 管 第 306 号, 2016 年度鹿管第 214 号,2017 年度鹿管第 579 号)を得 て行われた. 結 果 徳之島と奄美大島で得られた各種コウモリの生息地点 と個体数はAppendix 1,2 に記し,得られた音声のソナ グラムとパルスの測定値はFig. 6 と Table 3(アブラコウ 設置した. 洞窟性コウモリ類の調査はFig. 5 に記したように,龍 郷町屋入の銅山廃坑(p),大和村名音川中流域の用水路 内(c),名音の海蝕洞(c),フォレストポリスの隧道 2 ヵ 所(d),湯湾岳中腹の横穴(h),宇検村久慈大浜の洞窟 (r),宇検村西端の枝手久島の海蝕洞(f),瀬戸内町手 安の弾薬庫跡(w)および加計呂麻島三浦の防空壕(x) を調査した. リュウキュウテングコウモリとヤンバルホオヒゲコウ モリについて,2017 年 8 月に金作原(Fig. 4M)で捕獲 した前者 2 個体(雄 2),後者 3 個体(雄 2,雌 1)に発 信機を装着して個体追跡を行った.奄美大島における音 声録音・解析,捕獲後の個体の計測および発信機装着個 体の追跡は前述の徳之島の調査方法と同様である. また,徳之島と同様に,調査期間中の昼間,道路や林 道沿いにクワ科(ガジュマル,アコウおよびイヌビ)の 結実した樹木があれば,それらの樹下と周辺にクビワオ オコウモリの食痕があるかどうかを調査した.さらに, 聞き込み調査も実施した. 本研究で使用した小型発信機(Frequency range:150–
Fig. 5. Locations of the study areas (a–y) on Amami-Oshima Island and the distributions of Rhinolophus cornutus orii (●), Miniopterus fuscus
徳之島と奄美大島のコウモリ類の生態 21
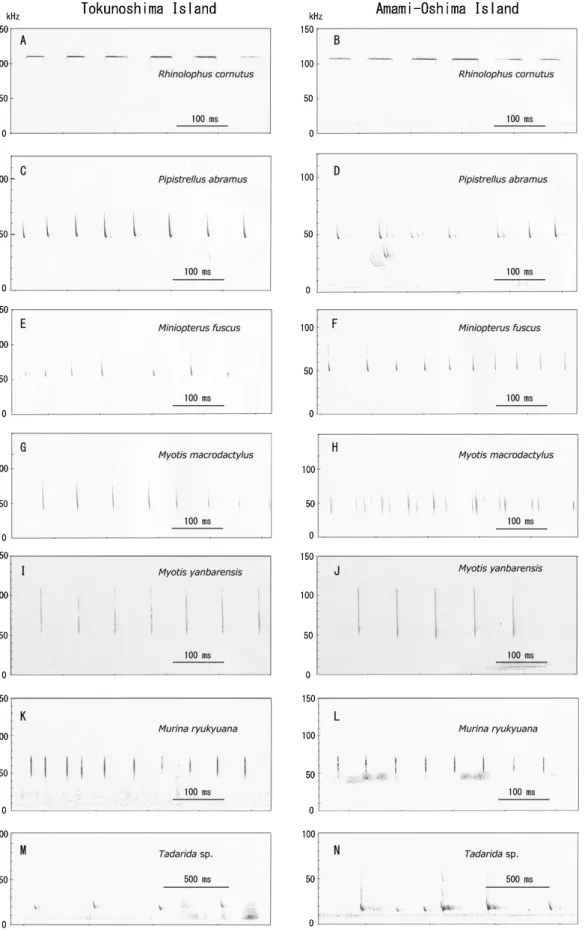
Fig. 6. Sonograms of the scanning echolocation calls emitted by Rhinolophus cornutus orii, Pipistrellus abramus, Miniopterus fuscus, Myotis macrodactylus, Myotis yanbarensis, Murina ryukyuana, and Tadarida sp. on Tokunoshima Island and Amami-Oshima Island.
音声(PF 値平均 24.9 kHz[21.5–27.4 kHz])は,これまで 徳之島から記録された 6 種のいずれとも一致しなかった. この音声をV1s として扱った(Tables 3 and 4,Fig. 7B1). 2)アカメガシワトラップで捕獲されたリュウキュウテ ングコウモリ 本調査期間の 2012 年~ 2014 年における捕獲記録を Table 1 にまとめた.ただし C 地域(Fig. 2)では,2013 年 11 月にアカメガシワ 20 束,2014 年 8 月に 15 束を設 置したが捕獲されなかった. 総捕獲数は 95 頭(成獣雄 39 頭,亜成獣雄 13 頭,成 獣雌 36 頭,亜成獣雌 7 頭)で,その内単独個体が 61 頭 (雄 43 頭,雌 18 頭)と 4 コロニーであった.アカメガ シ ワ ト ラ ッ プ に よ る 総 捕 獲 効 率 は 7.9 % で あ っ た (Table 1).捕獲された個体は,すべて森林低層部に設置 されたアカメガシワトラップを利用しており,中層部の アカメガシワトラップは全く利用されなかった.単独個 体以外に,成獣雌雄のペアがF 地域で 2013 年 11 月 13 日に 1 例あった.集団形成の例として,同地域で 2013 年 11 月 9 日に 16 頭(成獣雌 10 頭,亜成獣雌 3 頭,亜 成獣雄 3 頭),2014 年 8 月 29 日に 10 頭(成獣雌 4 頭, 亜成獣雌 3 頭,亜成獣雄 3 頭)および 9 月 1 日に 6 頭(成 獣雌 3 頭,亜成獣雌 2 頭,亜成獣雄 1 頭)がみられた. 8 月~ 9 月に捕獲された成獣雄について,隣接個体間の 距離は 69.1±11.23 m(Mean±SD,n=15)であった. 捕獲された個体のサイズについて,成獣雄の前腕長は 35.1±0.62 mm(n=39), 成 獣 雌 は 37.3±0.79 mm(n= 21)で,雄の最大値が雌の最小値に重ならず明らかに雌 雄差が認められ,雌が大きかった.非繁殖期の 8 月にお モリについては,サンプル数が少なかったのでパルスの 測定値を省いた)にまとめた. 1.徳之島 1)カスミ網による捕獲と音声録音によるコウモリ類 カスミ網による捕獲と調査地点における音声記録の結 果から,コウモリの確認地点をFig. 2 と Fig. 3 に示した. リュウキュウテングコウモリは,C 地域で生息が確認で きなかったがその他の地域(Fig. 2A,B,D,E,F およ びG)で確認された.また,2012 年 11 月 18 日のリュ ウキュウテングコウモリの捕獲時に,カスミ網設置周辺 (Fig. 2B)で本種の威嚇音(PF 値 28 kHz)が録音された (Fig. 7Ab).同様の音声は 2013 年 11 月の発信機装着個 体(成獣雄T201)の追跡の際にも録音された. ヤンバルホオヒゲコウモリの生息域はA ~ C に限ら れていた(Fig. 2).2013 年と 2014 年に行った A ~ C 地 域におけるカスミ網による本種の調査で,捕獲効率は 10.5%(2/19)で低かった. コキクガシラコウモリは,夜間にc 地域(Fig. 3)で 飛来していた.リュウキュウユビナガコウモリは,a ~ j 地域(b を除く)で広く音声が録音された(Fig. 3). ア ブ ラ コ ウ モ リ は,2012 年 11 月 18 日 に 花 徳 の 民 家 (Fig. 3b)で,出巣時に 15 頭がカウントされ,そのうち 3 頭(成獣雌 2 頭,成獣雄 1 頭)がカスミ網で捕獲された. 徳之島のa,c,d 地域(Fig. 3)で録音された FM-QCF 型の音声(PF 値平均 17 kHz)は奄美大島産の Tadarida sp. (船越 2010)と一致したので,この音声を Tadarida sp. の音声とした.c 地域で録音された異なる FM-QCF 型の
Table 3. Frequency of the maximum energy (PF) in the echolocation calls of six species of bats
Species Island Call structure PF (kHz)
n* Mean ± SD
Rhinolophus cornutus Tokunoshima CF 35 111.3 ± 1.27 a
Rhinolophus cornutus Amami-Oshima CF 25 107.7 ± 0.90 b
Myotis macrodactylus Tokunoshima FM 21 48.5 ± 2.37 a
Myotis macrodactylus Amami-Oshima FM 15 49.8 ± 2.61 a
Myotis yanbarensis Tokunoshima FM 16 66.1 ± 4.47 a
Myotis yanbarensis Amami-Oshima FM 22 62.9 ± 6.18 a
Miniopterus fuscus Tokunoshima FM 32 50.5 ± 1.55 a
Miniopterus fuscus Amami-Oshima FM 36 51.1 ± 1.23 a
Murina ryukyuana Tokunoshima FM 23 61.5 ± 4.67 a
Murina ryukyuana Amami-Oshima FM 36 59.9 ± 6.61 a
Tadarida sp. Tokunoshima FM-QCF 10 16.7 ± 1.56 a
Tadarida sp. Amami-Oshima FM-QCF 20 16.6 ± 1.11 a
*, number of measured pulses.
CF, Constant-frequency elements in the pulse; FM, Steep frequency-modulated components in the pulse; QCF, Quasi-constant-frequency.
徳之島と奄美大島のコウモリ類の生態 23 11 月には成獣雄 8.5±0.41 g(n=20),成獣雌 9.0±0.55 g (n=16) で 成 獣 雌 の 方 が 重 か っ た(Mann-Whitney’s U-test,U=74.0,P<0.005)が,雄は 9 月に比べて増加し, 一方雌は低下していた. 繁殖に関して,成獣雄の精巣のサイズ(長径)の季節 的変化をみると,8 月は精巣が委縮して小さかったが, 9 月 中 旬 に は 精 巣 が 肥 大 し 始 め て 長 径 4.3±0.06 mm (n=4),11 月中旬には 7.2±0.53 mm(n=18)と肥大し ていた.成獣雌では 8 月~ 11 月を通じて乳腺や乳頭が 委縮し,膣は開口していて,8 月上旬には出産・哺育を 終えていた.また,6 月 16 日に幼獣の雌雄(それぞれ 1 頭)が捕獲された. 再捕獲された個体のねぐら移動について,Table 5 に まとめた.成獣雄の移動距離は 17 ~ 390 m で,再捕獲 までの期間の長さに関係なく,個体によって付近に留ま るものや遠方 390 m に移動するものなど多様であった. 母子集団の母子ペアの移動が 2 例みられ,両ペアは 11 月に捕獲された後,翌年の 8 月~ 9 月に再捕獲された. その移動距離は 146 m であった. 3)発信機装着個体のねぐら利用と移動 リュウキュウテングコウモリの秋季 11 月における発 信機装着個体のねぐら利用をみると,日毎にねぐらを 変えており(Fig. 8),トラップ以外にも利用されるね ぐらとして,樹洞,群葉(広葉樹や蔓植物など),枯葉 (クワズイモAlocasia odora,オオタニワタリ Asplenium
antiquum など),シダ類の葉裏など森林内の多様な植物 を 利 用 し て い た. 最 大 移 動 距 離 は,A 個体(成獣雌 T204)178 m,B 個体(亜成獣雄 T216)163 m,C 個体(成 獣雌T205)25 m,D 個体(成獣雄 T201)43 m であった (Fig. 8).利用されたねぐらは地上高 1 ~ 3 m に位置し ていた. 8 月における発信機装着個体のねぐら利用をみると, 秋季と同様に日毎にねぐらを変えていた(Fig. 9).利用 されるねぐらの種類として,樹洞,枯葉,樹冠の群葉, シダの葉裏と多様であった.最大移動距離は,E 個体(成 獣雌S387)45 m,F 個体(成獣雄 S386)47 m,G 個体(成 ける成獣雄の体重は 8.3±0.31 g(n=13)で成獣雌は 9.5 ±0.47 g(n=12)で,成獣雌の方が重かった(Mann-Whitney’s U-test,U=0.00,P<0.001).9 月には成獣雄 8.0 ±0.39 g(n=11),成獣雌 9.6±0.36 g(n=11)で,雄の 最大値が雌の最小値と重ならず明瞭な雌雄差があった.
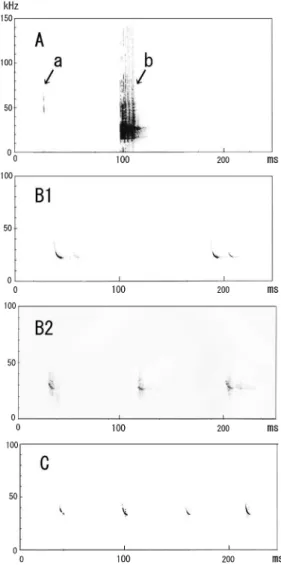
Fig. 7. Sonograms of the scanning echolocation calls (Aa) and
threat calls (Ab) emitted by Murina ryukyuana, and of the search phase echolocation calls (B1) emitted by V1s, search phase echolocation calls (B2) emitted by V2s, and those (C) emitted by Hs.
Table 4. Descriptive statistics for the echolocation calls of V1s, V2s, and Hs
Echolocation
calls Island Call structure n*
SF (kHz) PF (kHz) EF (kHz) D (ms) Mean ± SD Mean ± SD Mean ± SD Mean ± SD V1s** Tokunoshima FM-QCF 9 29.4 ± 1.88 24.9 ± 1.84 23.1 ± 1.66 11.7 ± 3.20 V2s** Amami-Oshima FM-QCF 15 29.2 ± 0.54 26.7 ± 0.48 25.3 ± 0.99 11.8 ± 1.27 Hs** Amami-Oshima FM-QCF 16 39.9 ± 1.11 35.5 ± 0.39 34.7 ± 0.41 8.3 ± 0.73 *, number of measured pulses.
**, V1s, V2s and Hs were emitted by unknown species, respectively.
同年 8 月 28 日における小島洞(Fig. 3f)の調査で,コ キクガシラコウモリが 18 時 45 分(日没前 5 分)に出洞 を開始し,その後断続的に出洞して合計 4 頭をカウント した.一方,リュウキュウユビナガコウモリが 19 時(日 没後 10 分)に出洞開始して合計 870 頭を記録した.同 月 29 日における面縄鍾乳洞(Fig. 3i)の同様の調査で, コキクガシラコウモリは 18 時 37 分(日没前 12 分)に 出洞を開始し,飛翔個体合計 41 頭をカウントした.リュ ウキュウユビナガコウモリは 18 時 55 分(日没後 6 分) に出洞を開始し,合計 410 頭を記録した.また,モモジ ロコウモリ 2 頭をカウントした.義名山鍾乳洞(Fig. 3g) では,同月 30 日の昼間に洞内調査を行い,コキクガシ ラコウモリ 1 頭の生息を確認した.同日夜間に目手久鍾 乳洞(Fig. 3j)の洞口で調査し,コキクガシラコウモリ 獣雄S385)78 m であった(Fig. 9).また,各雄のねぐら 移動域において隣接する雄間の重複がみられなかった. 4)洞窟調査におけるコウモリ類の分布と生息状況 洞窟性コウモリの確認地点について,Fig. 3 にまとめ た.2014 年 8 月 26 日の松原銅山廃坑(Fig. 3a)の入口 での調査で,コキクガシラコウモリが 18 時 43 分(日没 前 9 分)に出洞を開始し,その後連続的に出洞して合計 223 頭をカウントした.また,リュウキュウユビナガコ ウモリは遅れて 19 時 20 分(日没後 28 分)に出洞を開 始し,その後断続的に出動して合計 36 頭を記録した.
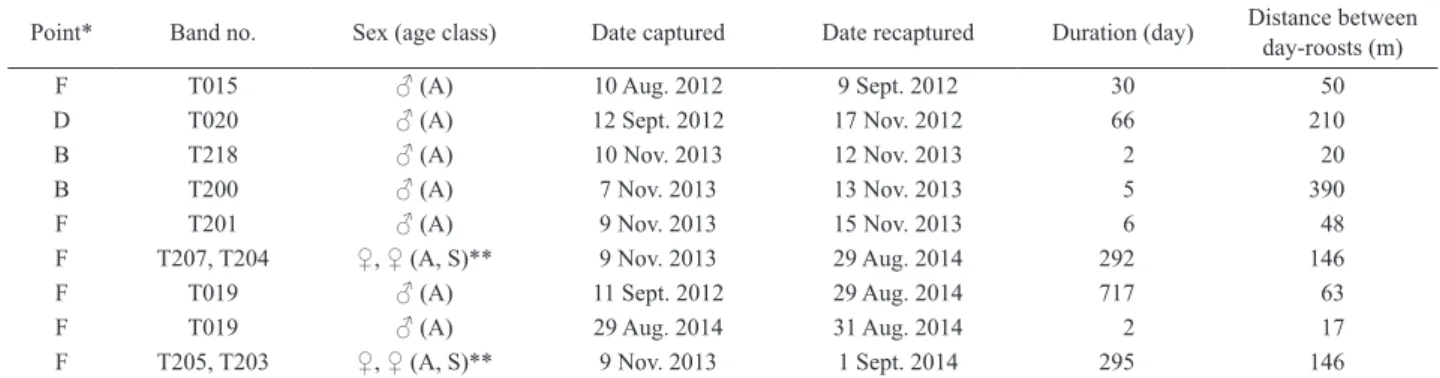
Table 5. Movement of day-roosts (traps) in Murina ryukyuana based on the recapture method
Point* Band no. Sex (age class) Date captured Date recaptured Duration (day) Distance between day-roosts (m) F T015 ♂ (A) 10 Aug. 2012 9 Sept. 2012 30 50 D T020 ♂ (A) 12 Sept. 2012 17 Nov. 2012 66 210 B T218 ♂ (A) 10 Nov. 2013 12 Nov. 2013 2 20 B T200 ♂ (A) 7 Nov. 2013 13 Nov. 2013 5 390 F T201 ♂ (A) 9 Nov. 2013 15 Nov. 2013 6 48 F T207, T204 ♀, ♀ (A, S)** 9 Nov. 2013 29 Aug. 2014 292 146 F T019 ♂ (A) 11 Sept. 2012 29 Aug. 2014 717 63 F T019 ♂ (A) 29 Aug. 2014 31 Aug. 2014 2 17 F T205, T203 ♀, ♀ (A, S)** 9 Nov. 2013 1 Sept. 2014 295 146 *, points are as in Fig. 2.
**, mother and child. A, adult; S, subadult.
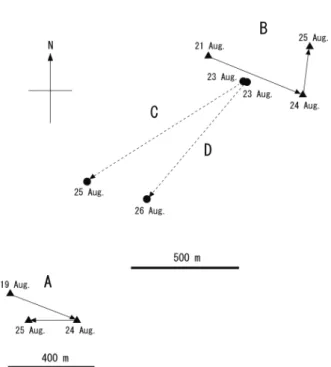
Fig. 8. Locations of the day-roost sites and movements of the
radio-tagged Murina ryukyuana on Tokunoshima Island from November 9 to 15, 2013. A, adult female (T204); B, subadult male (T216: male offspring of T204); C, adult female (T205); and D, adult male (T201).
Fig. 9. Locations of the day-roost sites and movements of the
radio-tagged Murina ryukyuana on Tokunoshima Island from August 27 to September 2, 2014. E, adult female (S387); F, adult male (S386); and G, adult male (S385).
徳之島と奄美大島のコウモリ類の生態 25 ウキュウテングコウモリは,10 地域(Fig. 10A ~ J)で 生息していたが,他の 16 地域(Fig. 4)では生息を確認 することができなかった.一方,ヤンバルホオヒゲコウ モリの生息域は 3 地域(Fig. 10B,G,I)に限られていた. I 地域では,本種が 2001 年 9 月 14 日に 5 頭(成獣雄 4 頭, 亜成獣雌 1 頭)捕獲され,2002 年 9 月 24 日に同捕獲地 点でその内の成獣雄 1 頭(No. H0331)が再捕獲された. 2016 年と 2017 年に行った上記 3 地域におけるカスミ網 による調査で,ヤンバルホオヒゲコウモリの捕獲効率は 34.8%(8/23)であった. コキクガシラコウモリは,9 地域における森林内を採 餌場所として利用していた(Fig. 5).本種の PF 値につ い て 奄 美 大 島 と 徳 之 島 の 間 で 有 意 差 が 認 め ら れ た (Table 3:Mann-Whitney’s U-test,U=2.5,P<0.001).
リュウキュウユビナガコウモリは,捕獲による生息 の 確 認 に 加 え て,12 地 域 の 林 道 で 音 声(PF 値 平 均 51.1 kHz:Table 3)と瞬時のライト照射による高速飛翔 個体の観察で確認された(Fig. 5).モモジロコウモリは, 捕獲と音声(PF 値平均 49.8 kHz:Table 3)から,3 地域 (Fig. 5d,e,g)で生息が確認された.2016 年~ 2017 年 の夏季の調査で,夜間に奄美市街で家屋からの出巣と飛 翔観察および音声(PF 値 45 kHz 前後)から,アブラコ ウモリの生息を確認した(Fig. 5y). の出洞開始は 18 時 35 分(日没前 13 分)で,合計 9 頭 を記録した.他方,リュウキュウユビナガコウモリの出 洞開始は 18 時 45 分(日没前 3 分)で合計 140 頭を記録 した.いずれも出洞開始は,コキクガシラコウモリがユ ビナガコウモリよりも早く,また洞口の比較的暗い面縄 鍾乳洞や目手久鍾乳洞では両種とも他洞よりも早く出洞 していた. コキクガシラコウモリの確認総数は,最大個体数を記 録した松原銅山廃坑(223 頭)と他 4 つの自然洞窟(計 55 頭)で合計 278 頭であった.リュウキュウユビナガ コウモリの確認総数は,最大個体数を記録した小島洞 (870 頭)と他 3 洞(586 頭)で合計 1,456 頭であった. 5)オオコウモリに関する食痕の有無と情報 調査期間中,林道沿いに点在するクワ科植物の結実し た樹木の周辺を調べたが,オオコウモリの食痕は見つか らなかった.聞き込み調査でも生息の情報は得られな かった. 2.奄美大島 1)カスミ網・ハープトラップによる捕獲と音声録音に よるコウモリ類 調査地域における捕獲と音声記録の結果から各種のコ ウモリの確認地点をFig. 5 および Fig. 10 に示した.リュ
日にタカバチ山周辺域(Fig. 10G)で捕獲した個体(成 獣雄:No. 468)が同年 8 月 22 日に同地域で再捕獲された. その移動距離は約 50 m であった. 3)発信機装着個体のねぐら移動 2017 年夏季の調査期間の後半は悪天候により発信機 装着個体の昼間のねぐら場所における追跡回数が少な かったが,発信機を装着してから数日を経過して受信し たので,ある程度の移動域を反映していると考えられる (Fig. 11).追跡できたリュウキュウテングコウモリ 2 個 体(成獣雄:S509,S510)について,最大移動距離は それぞれ 94 m と 83 m であった.一方,追跡できたヤ ンバルホオヒゲコウモリ 2 個体(成獣雄S516,亜成獣 雄S517)の最大移動距離は,それぞれ 140 m と 175 m であった. 4)洞窟調査におけるコウモリ類の分布と生息状況 洞窟性コウモリの分布と生息状況について,Fig. 5 に まとめた.コキクガシラコウモリについて,龍郷町屋入 の 銅 山 廃 坑(Fig. 5p)で 1986 年 7 月 28 日に母子総計 250 頭の集団,2002 年 9 月 23 日に約 60 頭の母子集団お よび 2008 年 6 月 12 日に母子総計 450 頭の集団,大和村 名音川中流域の用水路(Fig. 5c)で 1986 年 7 月 29 日に 母子総計 400 頭の集団,宇検村久慈大浜の洞窟(Fig. 5r) で 1986 年 7 月 29 日に 3 頭,瀬戸内町手安の弾薬庫跡 (Fig. 5w)で同年 7 月 30 日に約 30 頭,加計呂麻島三浦 オヒキコウモリ属の音声が奄美大島で広く記録された (Table 3,Figs. 5,6).この音声(PF 値平均 17 kHz)は Tadarida sp.(船越 2010)と一致したので,この音声を Tadarida sp. の音声とした.特に,c 地域や v 地域(Fig. 5) の断崖では日没 40 分後頃に高速飛翔するオヒキコウモ リ大の個体を観察することができ,ねぐら場所が確認さ れた.しかし,カスミ網による捕獲を 5 回試みたがカス ミ網の上空を通過するため捕獲できなかった. 2006 年 7 月 15 日にc 地域で録音された FM-QCF 型の 音声(PF 値平均 26.7 kHz[25.9–27.6 kHz])および 2017 年 8 月 25 日 に 録 音 さ れ た 音 声(PF 値 平 均 35.5 kHz [34.9–36.3 kHz])(Table 4,Fig. 7B2,C)は,奄美大島 でこれまで記録された 6 種の音声特徴と一致しなかっ た.前者の特徴を持つ音声をV2s,後者の特徴を持つ音 声をHs として扱った. 2)アカメガシワトラップで捕獲されたリュウキュウテ ングコウモリ 2006 年,2016 年 ~ 2017 年 に お け る 捕 獲 記 録 を Table 2 にまとめた.総捕獲数は 21 頭(成獣雄 17 頭, 亜成獣雄 2 頭,成獣雌 2 頭)で,すべて単独個体であっ た.総捕獲効率は 8.2%であった.捕獲された個体は, すべて森林低層部に設置されたアカメガシワトラップを 利用しており,中層部のアカメガシワトラップは全く利 用されなかった.その他の地域(Fig. 4E,D,O)では, 捕獲されなかった.それらを加えれば,総捕獲効率はそ れ(8.2%)よりさらに低下する.奄美大島ではアカメ ガシワトラップでの集団形成の例は得られなかった. 春~夏季に捕獲された成獣雄について,隣接個体間の距 離は 77.3±15.7 m(n=7)であった.前述の徳之島(平 均 69.1 m)の場合と比べて長いが,有意差は認められな かった(Mann-Whitney’s U-test,U=35.50,P=0.237). 捕獲された個体のサイズについて,成獣雄の前腕長は 34.4±0.62 mm(n=17),成獣雌は平均 35.8 mm(n=2) で あ っ た. 特 に, 成 獣 雄 に つ い て, 徳 之 島( 平 均 35.1 mm)のそれと比較すると短く,有意差が認められ た(Mann-Whitney’s U-test,Z=-3.563,P<0.001).8 月における成獣雄の体重は 7.5±0.54 g(n=14)で成獣 雌は 8.8 g(n=1)であった.11 月には成獣雄 7.8±0.81 g (n=3)で夏季より少し重くなっており,成獣雌は 9.1 g (n=1)であった. 繁殖に関して,成獣雄の精巣のサイズ(長径)の季節 的変化をみると,8 月は精巣が委縮していたが,11 月下 旬には精巣長径 8.3±0.64 mm(n=3)と肥大していた. 成獣雌では 8 月下旬の個体について乳腺や乳頭が委縮し ており,出産・哺育を終えていた.また,2017 年 5 月 5
Fig. 11. Locations of the day-roost sites and movements of the
radio-tagged adult males of Murina ryukyuana (A: S510 and B: S509) and Myotis yanbarensis (C: S517 and D: S516) on Amami-Oshima Island from August 19 to 26, 2017.
徳之島と奄美大島のコウモリ類の生態 27
ズイモやオオタニワタリの枯葉が追加された.同属近縁 のコテングコウモリも樹洞,枯葉,群葉などを昼間のね ぐら場所として利用している(船越 1998;本多 2002; 吉行・苅部 2002;Hirakawa and Kawai 2006;平川 2007; 船越ほか 2009).両島には本州や九州本土に比べて樹木 や草本類の葉のサイズが大きいため,量的にも多くの自 然物がねぐら場所として供給されていると思われる. リュウキュウテングコウモリの単独個体によって利用 されるねぐらの位置について,発信機装着個体の追跡で は,地上高 1 ~ 3 m であった.夜間の観察では低層部を 飛翔していることが多く,ねぐらの発見率が中層部より も低層部の方が高いためと推測される.森林中層部を利 用していた稀な例として,地上高 5 m のフカノキの群葉 枯葉にいた個体の観察がある(亘・船越 2013). テングコウモリ属のねぐらは,湿った樹皮下や枯葉よ りも比較的乾燥したものが選択されていると思われる. 徳之島(伊仙町)の年間降水量は,1,900 mm(統計期 間 1981 年~ 2010 年;気象庁URL:https://weather.time-j. net/Stations/JP/isen;2019 年 4 月 3 日確認)であり,奄 美大島(名瀬市)の 2,800 mm(統計期間 1981 年~ 2010 年;気象庁URL:https://weather.time-j.net/Stations/JP/naze; 2019 年 4 月 3 日確認)に比べて少ない.こうした降水 量に伴う湿度条件も本種の生息条件に密接に関係してい ると思われる.また,アカメガシワトラップでの総捕獲 効 率 は, 徳 之 島 で 7.9%,奄美大島で 8.2%であった (Tables 1,2).鹿児島県霧島市と宮崎県都城市で行った コテングコウモリのアカメガシワトラップ調査では 10.5%(船越ほか 2016)であったことから,リュウキュ ウテングコウモリの捕獲効率が低い原因の一つとして, 森林内のねぐら場所が樹洞や枯葉以外にも多様な生葉 (群葉)が選択されるためと考えられる. リュウキュウテングコウモリの個体追跡の結果から, ねぐらを日毎に変えていることが判明した.こうした特 性は,多くの森林性コウモリ類でみられる(O’Donnell 2000;Willis and Brigham 2004;Barclay and Kurta 2007; 松岡 2008;船越ほか 2009,2016;谷岡 2016).発信機 装着個体におけるリュウキュウテングコウモリのねぐら の最大移動距離は 25 ~ 178 m で比較的に短かった.発 信機装着地点から 500 m 前後の範囲で方探したことか ら,受信が途切れた場合は,500 m 以上の遠方に去って しまったことも考えられる.しかし,再捕獲個体の比較 的短い移動距離(Table 5)を考慮すれば,むしろ樹洞の 奥に入っていたことや樹洞内でアンテナが曲がってし まって受信できなかった可能性が高い. 本種の繁殖について,徳之島では 6 月に幼獣が捕獲さ の防空壕(Fig. 5x)で 1986 年 7 月 30 日に 6 頭,宇検村 西端の枝手久島の海蝕洞(Fig. 5f)で 2010 年 9 月 11 日 に約 500 頭,フォレストポリスの隧道 2 ヵ所(Fig. 5d) で同年 9 月 12 日に各 3 頭と 22 頭および湯湾岳中腹の横 穴(Fig. 5h)で同日に 2 頭の生息が確認された.コキク ガシラコウモリの確認個体数は,以上の廃坑や自然洞窟 計 9 地点で合計 1,411 頭であった. リュウキュウユビナガコウモリは,宇検村役場付近の マンガン廃坑(Fig. 5r)で 1986 年 7 月 29 日に 13 頭の 集団,瀬戸内町手安の弾薬庫跡(Fig. 5w)で約 50 頭の 集団が観察されたに過ぎない.また,これまで調査され た洞内でモモジロコウモリは観察されなかった. 5)オオコウモリに関する食痕の有無と情報 調査期間中,林道沿いに点在するクワ科植物の結実した 樹木の周辺を調べたが,オオコウモリの食痕は見つから なかった.聞き込調査でも生息の情報は得られなかった. 考 察 1.両島における各種の生態・分布と差異 1)森林性コウモリ類 リュウキュウテングコウモリについて,徳之島では 1999 年に天城町の丹発山や井之川岳周辺域で捕獲記録 されている(前田ほか 2001).今回の調査では,1 地域 を除く,6 地域で生息が確認された(Fig. 2).奄美大島 では 2000 年に湯湾岳登山口付近で本種の雄 1 頭が捕獲 されている(前田ほか 2002).今回の奄美大島の調査で 新たに生息地点が追加されたが,調査した 26 地点中, 10 地点に限られていた.両島の常緑広葉樹林内でも捕 獲されない地域があり,特に奄美大島では上記のよう に生息地域が限られていた.徳之島の森林面積(一次 林,二次林,植林地)107 km2は,奄美大島の森林面積 (685 km2)の 16%に過ぎない(九州森林管理局 2016). これを考慮すれば,森林面積に占める本種の生息域の割 合は高く,徳之島の方が奄美大島よりも高密度で生息し ていると考えられた. 予 備 調 査 に よ れ ば, 近 縁 種 の コ テ ン グ コ ウ モ リ Murina ussuriensis も九州においては同様に常緑広葉樹 林内に限られ,高木樹齢 30 年以下の二次林や雑木林で は生息していない.リュウキュウテングコウモリが生 息する常緑広葉樹林では,被食昆虫やねぐら場所が豊 富であると予想される.本種のねぐら場所として,樹 洞, イ イ ギ リIdesia polycarpa,アオバノキ Symplocos cochinchinesis,フカノキ Schefflera heptaphylla の枯葉, 群葉,シダ類が利用されている.今回の個体追跡でクワ
積脂肪への依存度はきわめて低いと考えられる.今後, 冬季の活動について調査する必要がある. ヤンバルホオヒゲコウモリは,徳之島において 1999 年に三京(Fig. 2B)で最初に 2 頭が捕獲記録されている (前田ほか 2001).今回の調査で,新たに 2 地域(Fig. 2A, C)が追加された.一方,奄美大島では 2000 年に金作 原国有林で 3 頭捕獲されている(前田ほか 2002).今回 の調査でこの地域に加えて新たに 2 地域(Fig. 10B,G) で生息が確認された.本種におけるカスミ網による捕獲 効率が徳之島で 10.5%であるのに対して奄美大島では 34.8%であったことから,徳之島の生息密度が非常に低 いことを示唆している.今後の調査で両島間の生息密度 の違いの要因を検討する必要がある. ヤンバルホオヒゲコウモリのねぐら場所に関して,不 明な点が多い.九州本土に生息する近縁のクロホオヒゲ コウモリMyotis pruinosus の生息場所は,比較的樹高の 高い照葉樹林に限られ,ねぐらとして隧道内天井の窪み が利用されている(船越ほか 2013b).また,同属の森 林 性 コ ウ モ リ 3 種( ヒ メ ホ オ ヒ ゲ コ ウ モ リMyotis ikonnikovi,ドーベントンコウモリ M. daubentonii,ウス リホオヒゲコウモリM. gracilis)の昼間のねぐら場所と して,樹洞のほかに,剥離した樹皮内,束になった枯葉 内,橋梁下が利用されている(Yasui et al. 2004;河合・ 赤坂 2008).ヤンバルホオヒゲホウモリもこうしたねぐ ら場所を含む類似の森林に生息していると考えられる. 奄美大島の金作原で,最初に捕獲放獣された成獣雄がそ の 1 年後に同地点の林道で再捕獲された.この事例や発 信機装着個体の追跡で得られたねぐら間の移動距離 (200 m 以内)の結果から,長距離移動することなく特 定の場所に定着していることが示唆された.また,夏季 における発信機装着個体の追跡で,雄 2 個体が別々のね ぐらを選択していたことから,リュウキュウテングコウ モリと同様に単独生活に入っていることが示唆された. 2)洞窟性コウモリ類 コキクガシラコウモリは,先行調査では,徳之島にお ける 1999 年秋季の洞窟調査では,廃坑や自然洞窟計 4 地点で合計 1,166 頭が記録されている(前田ほか 2001). 現状では,それらの生息地の多くは土地造成で崩壊して いるか,自然崩壊が進んでいた.今回の調査では,自然 崩壊が進んでいる松原銅山廃坑と 4 つの自然洞窟で合計 278 頭しか確認できなかった(Fig. 3).徳之島における 本種の個体数は減少傾向の可能性がある.一方,今回の 調査では奄美大島において,廃坑や自然洞窟計 9 地点の 調査で合計 1,411 頭を記録した(Fig. 5).夜間の飛翔個 体の調査で両島において広く生息していた(Figs. 3,5). れている(船越ほか 2013a).一方,7 月中旬には乳腺が 発達した成獣雌が捕獲されている(前田ほか 2001).8 月には成獣雌の乳頭・乳腺ともに委縮しており,成獣雄 の精巣も委縮していた.9 月には精巣の肥大が始まり, 11 月には精巣の長径平均 7.2 mm に達していた. これらの繁殖に関わる事実から,出産・哺育時期は 5 月 ~ 7 月と推定された.この期間はコテングコウモリの 6 月初旬~ 7 月下旬(船越ほか 2009)に比べて長く,個 体によって出産時期が大幅に遅延していると考えられ る.今回の調査で出産集団を観察することができなかっ たが,コテングコウモリ(船越ほか 2009,2016;谷岡 2016)やテングコウモリMurima hilgendorfi(佐藤・勝田 2007;渡邊・船越 2017)と同様に,出産直前頃から集 団を形成すると考えられる.出産や哺育時期の母子集団 の観察は今後の課題であるが,最近,龍郷町の「奄美自 然観察の森」で 2018 年 5 月 8 日に地上高 7 m の群葉内 に母子集団(約 15 頭)が発見され撮影された(Vladimir Dinets 私信).コテングコウモリ(Fukui et al. 2012;船 越ほか 2016;谷岡 2016)と同様に,中層部における母 子集団の形成は出産・授乳期の 5 月~ 7 月と予想される. リュウキュウテングコウモリの母子集団が 8 月~ 11 月 までみられた.その集団の規模は最大 16 頭であった. また,集団形成は,森林の低層部(地上高 1 ~ 2 m)に 形成され,中層部には形成されなかった.その要因とし て,夏~秋季の台風による悪天候下でその影響の少ない 低層部を選択していたと考えられる. 成獣雌は 11 月中旬頃から単独生活に入っていた.こ の時期に雌雄のペアが捕獲されたが,交尾行動について は観察されなかった.11 月中旬にリュウキュウテング コウモリの威嚇音を録音した.コテングコウモリでも同 様の威嚇音が記録されている(船越ほか 2016).加えて, 近隣雄個体間でねぐらの重複がみられないことから,こ の時期に雄間の縄張り争いまたは闘争が起きていること が示唆される. リュウキュウテングコウモリのサイズに関して,成獣 雄の前腕長では徳之島産(平均 35.1 mm)が奄美大島産 (平均 34.4 mm)に比べて長く,8 月の体重では徳之島 産(8.3 g)が奄美大島産(7.5 g)に比べて重かった.成 獣雌でも同様の傾向があり,徳之島産が奄美大島産に比 べて大型であった.成獣雌においても同様での傾向に あった沖縄島産の成獣雄の前腕長は 37.0 mm(Maeda and Matsumura 1998)で徳之島産よりさらに大きい.秋 季の体重変化について,越冬前の 11 月にも夏季と変わ らなかった.温帯産コウモリ類では冬眠前の顕著な脂肪 蓄積がみられるが(船越 2000),本種の冬季における蓄
徳之島と奄美大島のコウモリ類の生態 29 での記録は少なかったが今回の調査で,徳之島町花徳の 民家で捕獲され生息が再確認された.奄美大島では北部 の笠利町で取得された標本がある(Yoshiyuki 1989).ま た奄美大島でも今回の調査で,奄美市街において本種の 飛来の目撃と音声を記録し,生息が確認された.両島に おいて他の市街地や民家を精査すれば,生息地域がもっ と広がると思われる. クビワオオコウモリは両島で生息を確認することがで きなかった.徳之島の天城町当部では 2013 年 1 月 18 日 にクビワオオコウモリPteropus dasymallus の死体(亜成 獣雌)が発見された(船越 2017a).この個体は体サイ ズが小さいことから,亜種オリイオオコウモリP. d. inopinatus と思われる.生息が確認されている他島から 飛来したとすれば,徳之島に一番近い沖永良部島(島間 の距離約 40 km)か,さらに南方の島嶼から飛来したこ とが考えられる.一方,頭骨頭頂部が未完成であったこ とから前年の春~夏季に出生した個体と判明した(船越 2017a).若年個体は一般的に飛翔能力が劣っていること を考慮すれば,徳之島で生まれ育った個体の可能性も捨 てきれない.しかし,本調査以前の聞き込み調査でオオ コウモリの目撃情報は得られず,当地での繁殖の可能性 はきわめて低いと思われる.奄美大島では 1943 年に住 用村の山間で捕獲されている(森田忠義 私信).また, 1983 年 10 月に住用村で目撃されている(安間 1987). しかし,その後の奄美大島の調査で本種を目撃できず, 最後に目撃された後 30 年を経過した中で目撃情報が得 られないことから,その後消滅したと思われた.一方, 動物写真家の情報で瀬戸内町の嘉徳で十数年前に目撃し た(常田 守 私信)との情報を得たことから,定着の 可能性は低いが,稀に他島からの飛来があると予想され る.本種は液果への嗜好性が高く,被食樹種は自然林内 よりも住民の生活域に点在するクワ科のガジュマルやア コウなどである(船越・國崎 2003).こうした樹種への 飛来が目撃されて生息が確認されれば,これらの樹種の 植栽など保全への取り組みが求められる. 4)音声の記録により推測される生息種 今回の研究で奄美大島および徳之島において録音さ れ,Tadarida sp. とされた音声は,船越(2010)が指摘 し て い る よ う に ス ミ イ ロ オ ヒ キ コ ウ モ リ(Yoshiyuki 1989;Funakoshi and Kunisaki 2000)の可能性が考えられ る.徳之島においてTadarida sp. の生息はこれまで確認 されていないが,音声が記録されたことより同島におい てもTadarida sp. の生息している可能性が高まった.本 調査よって,本種のねぐら場所が奄美大島の大和村の名 音(Fig. 6c)と瀬戸内町半島部西端の曽津高崎(Fig. 6v) しかし,両島において,未発見・未調査の洞窟(海食洞 など)があると思われるので,今後の調査地点を増やし て生息状況を確認することが必要である. 両島間のコキクガシラコウモリの音声を比較する と,PF 値に有意差が認められ興味深い.一方,近縁の 沖縄本島のオキナワコキクガシラコウモリRhinolophus pumilus において,北部と南部の個体群間で CF 周波数 に大きな差異がみられたこと(松村 2005)から,島間 ばかりでなく島内の地域個体群間でも差異が生じている のか今後検討する必要がある. リュウキュウユビナガコウモリについて,夏季におけ る徳之島の各洞口での飛翔カウント総数は約 1,500 頭で あった(Fig. 3).最大個体数(870 頭)を記録した小島 洞は,かつて観光洞であったが,現在は放置されて人為 的影響が軽減され生息環境が回復したと思われる.先行 調査による徳之島の主要 4 洞窟における 1999 年おける 秋季の総個体数は 727 頭であった(前田ほか 2001).一方, 奄美大島では宇検村のマンガン廃坑で 10 頭の集団が観 察され,名音の海食洞で本種の音声が記録されただけで ある(Fig. 5w,c). モモジロコウモリについて,徳之島では 1984 年にお ける島内 10 洞窟の調査で本種の生息が確認されなかっ た(沢田 1996).その後,1999 年の調査で天城町の三京 林道や南部ダム付近で捕獲され,徳之島が本種の南限と して記録された(前田ほか 2001).今回の調査では徳之 島南部の洞窟で生息を確認したが,三京林道や南部ダム 付近では生息が確認できなかった.他方,奄美大島では モモジロコウモリの標本が,捕獲場所は不明であるが, 国立科学博物館に保管されている(前田 2001).今回の 調査で,モモジロコウモリはフォレストポリスで捕獲さ れ,他 2 地域の渓流で生息が確認された(Fig. 5d,e,g). モモジロコウモリの音声識別については,PF 値(平均 49.8 kHz:Table 3)およびパルスの終部がアブラコウモ リやユビナガコウモリのようなカーブがみられないこと (船越 2010)から判定された.両島ともモモジロコウモ リの生息域はきわめて限られている.前田(2001)は, 徳之島では比較的最近生息するようになったと推測して いるが,その逆に本種の生息環境が狭められたとも考え られる. 3)その他の種 アブラコウモリについて,徳之島では伊仙町内で捕獲 され液浸された標本 3 頭が伊仙町立歴史民俗資料館に保 管されている(前田ほか 2001).また,2015 年 11 月 30 日に天城町平土野の民家で本種 1 頭が捕獲された(岡崎 幹人 私信).このように本種は徳之島においては今ま
を持たないV1s が 1 種類録音された.このことから, 徳之島には既知 7 種に新たに 2 種加えた 9 種の小コウモ リ類が生息している可能性が高い.一方,奄美大島では, 既知 7 種の音声特徴とは異なる音声Hs と V2s が録音さ れた.このことから,奄美大島には 9 種の小コウモリ類 が生息している可能性が高い.また,徳之島の森林面積 は奄美大島の森林面積の 16%であることを考慮すれば, 面積あたりの種数は奄美大島よりも豊富である.さらに 長崎県対馬の面積(708 km2:森林率 89%)は徳之島 (248 km2)の約 3 倍であるが,コウモリの種数は 6 種で ある(河合ほか 2007).このことから,徳之島のコウモ リ相は豊富である. 奄美・徳之島両島のコウモリ類は,生態系の中で飛翔 昆虫の捕食者として,他方では被食者として位置づけら れる.しかし,今回の調査で,コウモリの捕食者は確認 できなかった.潜在的な捕食者としてリュウキュウコノ ハズクOtus elegans やヘビ類が予想される.一方,興味 深い事例として,1995 年の夏季にオオジョロウグモ Nephila pilipes の網にリュウキュウテングコウモリが掛 かっていた(恵沢岩生 私信).また,2007 年 7 月にオ リイコキクガシラコウモリがオオジョロウグモの網に捕 らえられた(朝日新聞:2007 年 9 月 19 日付).同様の 例は 2015 年 10 月 4 日にも撮影され,オオジョロウグモ がオリイコキクガシラコウモリの首筋を咬んでいた(石 田 健 私信).こうした事例は今回の調査では得られ ていないので,その頻度は極めて少ないと考えられる. 徳之島では,希少種のリュウキュウテングコウモリや ヤンバルホオヒゲコウモリの生息地は,北部の天城岳周 辺地域,中部の三京・犬田布岳・剥岳・井之川岳および 神嶺ダム地域,南部の東部ダム地域の二次林であった. これらの地域には,イタジイやアラカシを主体とする常 緑広葉樹がみられるほか,谷や斜面の下の方のやや湿潤 な所にはオキナワウラジロガシの一次林がみられる.奄 美大島では,希少種を含むコウモリ種数が多い地域は金 作原,タカバチ山周辺域,フォレストポリスおよび湯湾 岳麓であった.これらの地域は,徳之島と同様の樹木構 成の常緑広葉樹を有している.これらの地域の多くは奄 美群島国立公園の指定地域であるので森林保護の結果, 一部でオキナワウラジロガシの一次林がみられるが,今 後,森林の保全が進めば二次林から原生的自然林に変遷 することが予想される.今回の研究から,希少種のヤン バルホオヒゲコウモリの生息域は巨木が混在する常緑広 葉樹林に限られていたことが分かった.森林の保全とと もに,今回の研究成果がコウモリ類の具体的な保護など に関する資料として活用されることを期待する. の海岸の断崖の岩盤亀裂(割れ目)であることが判明し た.今後,本種の比較的低い飛行ルートを見つけて捕獲 し,種を確定したい. 徳之島で録音された音声V1s は,船越(2010)にお いて報告されたヒナコウモリの音声の特徴に一致してい た.奄美大島で録音された音声V2s は V1s と類似して いたが,PF 値が有意に異なっており(Mann-Whitney’s U-test,U=95.0,P<0.006),国内で録音されたコウモ リ類の音声のうちヒメヒナコウモリVespertilio murinus の音声の特徴(Fukui 2015a)に類似していた.徳之島と 奄美大島にはこれまでヒナコウモリやヒメヒナコウモリ の分布を示す記録はないが,音声分析によってこれらの 種が分布している可能性が示された.これらの音声がヒ ナコウモリまたはヒメヒナコウモリによって発せられた ものなのかに加え,両種ともに翼狭長型で長距離飛翔が 可能と考えられていること(Fukui 2015a,b;佐藤ほか 2017)から徳之島と奄美大島本島間(約 50 km)で行き 来があるのかどうかなど,今後の調査結果がまたれる. 奄美大島で録音された音声Hs は,船越(2017b)も すでに公表して指摘しているように,クロオオアブラコ ウモリ Hypsugo alashanicus の精査音(PF 値 34–37 kHz: Fukui et al. 2013)がこの変異内にあって一致しているこ とやその他のHypsugo 属の音声特徴(Russo and Jones 2002)と類似していた.また,浅利・木元(2018)によっ て捕獲された種不明コウモリの音声特徴(PF 値 35 kHz) とも類似していた. クロオオアブラコウモリは,近隣のアジア大陸にも生 息しているが,日本では北海道,青森県および長崎県対 馬からの記録しかない(Kawai 2015a).この音声がクロ オオアブラコウモリとすれば,日本での分布域はさらに 広がることになる.記録された生息域と類似の環境で生 息している可能性が高く,今後この種不明のHypsugo sp. を捕獲して種を同定する必要がある. 2.今後の課題 前述したように,徳之島と奄美大島における各種コウ モリについて,未同定の種の精査を含めて,今後の検討 課題を提案した.両島内において 2017 年に奄美群島国 立公園が設立され,世界自然遺産の推薦地として,今後 の生物多様性の維持と希少種の保全の取り組みが進めら れている.このタイミングで希少種を多く含むコウモリ 類の現状を本研究で示した. 徳之島において,音声解析からこれまで記録のない Tadarida sp. が生息している可能性が強く示唆された. また,これまで同島から知られている 7 種の音声の特徴
徳之島と奄美大島のコウモリ類の生態 31 Y. Ishibashi, M. A. Iwasa, D. Fukui and T. Saitoh, eds.) The Wild Mammals of Japan, Second edition, pp. 96–98. Shoukadoh Book Sellers and the Mammal Society of Japan, Kyoto.
Fukui, D., Hill, D. A. and Matsumura, S. 2012. Maternity roosts and behavior of the Ussurian tube-nosed bat Murina ussuriensis. Acta Chiropterologica 14: 93–104.
Fukui, D., Mochida, M., Yamamoto, A. and Kawai, K. 2013. Roost and echolocation call structure of the Alashanian pipistrelle
Hypsugo alashanicus: first confirmation as a resident species in
Japan. Mammal Study 38: 61–66.
船越公威.1998.鹿児島県口永良部島,屋久島および種子島産 の翼手類と食虫類.哺乳類科学 38: 293–298. 船越公威.2000.コウモリ.冬眠する哺乳類 3 コウモリ.冬 眠する哺乳類(川道武男・近藤宣昭・森田哲夫,編),pp. 103–142.東京大学出版会,東京. 船越公威.2010.九州産コウモリ類の超音波音声による種判別 の試み.哺乳類科学 50: 165–175. 船越公威.2017a.徳之島で発見されたクビワオオコウモリ
Pteropus dasymallus について.Nature of Kagoshima 43: 9–12.
船越公威.2017b.奄美大島におけるコウモリ類,特に絶滅危 惧種コウモリ類の現状と保全について.Pro Natura ニュー ス 27: 13.
船越公威・北之口卓志・田中広音・大坪将平・大平理紗・内原 愛美.2013a.徳之島における希少種リュウキュウテング
コウモリMurina ryukyuana の生態.Nature of Kagoshima 39:
1–6.
Funakoshi, K. and Kunisaki, T. 2000. On the validity of Tadarida
latouchei, with reference to morphological divergence among T. latouchei, T. insignis and T. teniotis (Chiroptera, Molossidae).
Mammal Study 25: 115–123. 船越公威・國崎敏廣.2003.エラブオオコウモリの食性,被食 植物の分布及び生息域の植生.エラブオオコウモリ天然記 念物緊急調査報告書(鹿児島県上屋久町教育委員会,編), pp. 44–54.鹿児島県上屋久町教育委員会,上屋久. 船越公威・長岡研太・竹山光平・犬童まどか.2009.コテング コウモリMurina ussuriensis におけるアカメガシワトラッ プのねぐら利用と繁殖生態.哺乳類科学 49: 245–256. 船越公威・坂田拓司・河合久仁子・荒井秋晴.2013b.熊本県 におけるクロホオヒゲコウモリMyotis pruinosus の生息初 記録と外部形態・頭骨およびミトコンドリアCyt b 遺伝子 の変異について.哺乳類科学 53: 351–357. 船越公威・玉利高志・市耒原優樹・北之口卓志・田中広音. 2016.コテングコウモリMurina ussuriensis の生態,特に ねぐら利用の雌雄差と繁殖関与年齢について.哺乳類科学 56: 117–128. 平川浩文.2007.コテングコウモリ(Murina ussuriensis)の夏 季におけるねぐら利用.東洋蝙蝠研究所紀要 6: 1–7. Hirakawa, H. and Kawai, K. 2006. Hiding low in the thicket: roost
use by Ussurian tube-nosed bats (Murina ussuriensis). Acta Chiropterologica 8: 263–269.
本多宣仁.2002.コテングコウモリの休息場所.コウモリ通信 10: 5.
Kawai, K. 2015a. Hypsugo alaschanicus (Bobrinskii, 1926). In (S. D. Ohachi, Y. Ishibashi, M. A. Iwasa, D. Fukui and T. Saitoh, eds.)
謝 辞 本調査において,コウモリの調査に協力していただい たコウモリの会の大沢夕志氏,大沢啓子氏,徳之島天城 町役場企画課自然保護専門員の岡崎幹人氏,徳之島虹の 会の美延睦美氏,美延治郷氏,関 政樹氏,宮前延代氏, 大坪博文氏,鹿児島県環境技術協会の岡田 滋氏,東京 大学医科学研究所・奄美病害動物研究施設の服部正策博 士,奄美大島在住の恵沢岩生氏,元大島北高校の永田武 志氏,鹿児島県立博物館の金井賢一氏,東京大学大学院 農学生命科学研究科附属演習林の福井 大博士,大阪大 学工学研究科環境・エネルギー工学専攻大学院生の増田 圭祐氏,東京大学大学院農学生命科学研究科の石田 健 博士,四国自然史科学研究センターの谷地森秀二氏,谷 岡 仁氏,奄美大島のオオコウモリの情報をいただいた 森田忠義氏,常田 守氏,奄美大島の道路事情等の情報 をいただいた奄美自然保護官事務所の方々,国有林野入 林許可をいただいた鹿児島県森林管理署の徳之島森林事 務所と奄美大島名瀬森林事務所の方々に厚くお礼申し上 げる.特に,調査に同行し,クロオオアブラコウモリと 類似する音声とのご指摘をいただいた福井 大博士,奄 美大島のリュウキュウテングコウモリの母子集団の情報 をいただいた沖縄科学技術大学院大学のVladimir Dinets 博士に感謝申し上げる.なお,本研究の一部は,鹿児島 国際大学 2012 年度フィールドワークアクションプログ ラムの助成,公益財団法人自然保護助成基金の第 24・ 27 期プロ・ナトゥーラ・ファンド助成により行われた ものである. 引 用 文 献 Amelon, S. K., Dalton, D. C., Millspaugh, J. J. and Wolf, S. A. 2009. Radiotelemetry: techniques and analysis. In (T. H. Kunz and S. Parsons, eds.) Ecological and Behavioral Methods for the Study of Bats, 2nd edition, pp. 57–77. The Johns Hopkins University Press, Baltimore.
浅利裕伸・木元侑菜.2018.奄美大島で捕獲された種不明のコ ウモリ類.哺乳類科学 58: 67–71.
Barclay, R. M. R. and Kurta, A. 2007. Ecology and behavior of bats roosting in tree cavities and under bark. In (M. J. Lacki, J. P. Hayes and A. Kurta, eds.) Bats in Forests, pp. 17–59. Johns Hopkins University Press, Baltimore.
Fukui, D. 2015a. Vesperrilio murinus Linnaeus, 1758. In (S. D. Ohachi, Y. Ishibashi, M. A. Iwasa, D. Fukui and T. Saitoh, eds.) The Wild Mammals of Japan, Second edition, pp. 94–95. Shoukadoh Book Sellers and the Mammal Society of Japan, Kyoto.