イヌのホスホフルクトキナーゼ
-1
の活性調節日本大学大学院獣医学研究科 金井 修一郎
2019
目 次
第
1
章 序論1
第
2
章 イヌの各種臓器におけるPFK-1
の比活性およびアイソフォーム構成 72.1
緒言8
2.2
材料および方法10
2.2.1
材料10
2.2.2
供試臓器11
2.2.3
細胞質画分の分離および低分子物質の除去12
2.2.4 PFK-1
の活性測定12
2.2.5 FBP-1
の活性測定13
2.2.6
ウェスタンブロッティング13
2.2.7 real-time RT-PCR 14
2.3
結果15
2.3.1 PFK-1
比活性15
2.3.2 FBP-1
比活性15
2.3.3 PFK-1
アイソフォームおよびFBP-1
のタンパク質発現ウェスタンブロッティング
16
2.3.4 real-time RT-PCR 16
2.4
考察17
第
3
章 イヌ骨格筋PFK-1
の活性調節26
3.1
緒言27
3.2
材料および方法28
3.2.1
材料28
3.2.2
供試臓器29
3.2.3 PFK-1
部分精製29
3.2.4
タンパク質濃度測定30
3.2.5 PFK-1
の活性測定30
3.3
結果31
3.3.1
イヌ骨格筋PFK-1
の精製度31
3.3.2 pH
の効果32
3.3.3 ATP
の効果32
3.3.4 F-6-P
の効果32
3.3.5 F-2,6-P
2の効果33
3.3.6 AMP
の効果33
3.3.7 cAMP
の効果33
3.3.8
クエン酸の効果34
3.3.9 UTP
の効果34
3.4
考察35
第
4
章 イヌ肝臓PFK-1
の活性調節47
4.1
緒言48
4.2
材料および方法49
4.2.1
材料49
4.2.2
イヌ肝臓PFK-1
の部分精製49
4.2.3 PFK-1
の活性測定50
4.2.4
タンパク質濃度測定51
4.3
結果51
4.3.1 pH
の効果51
4.3.2 ATP
の効果52
4.3.3 F-6-P
の効果52
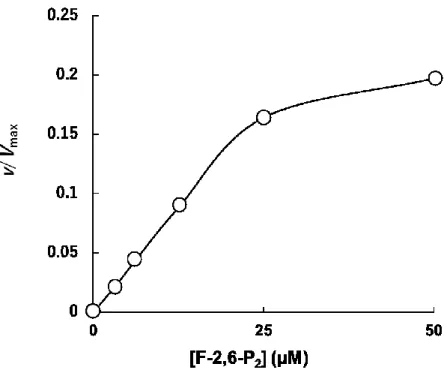
4.3.4 F-2,6-P
2の効果52
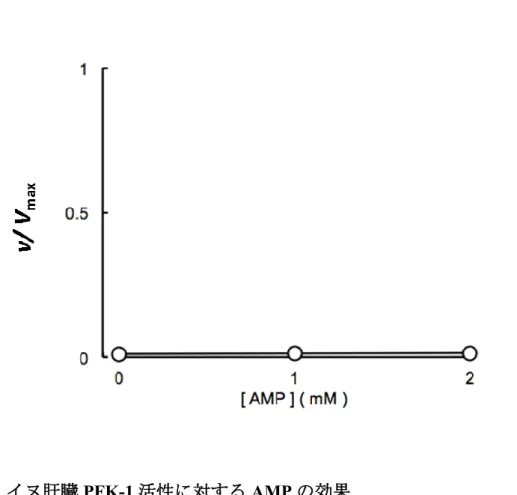
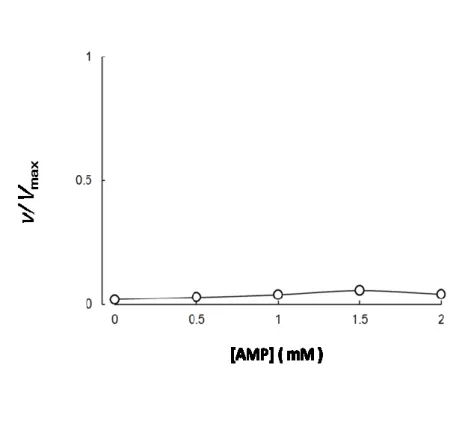
4.3.5 AMP
の効果53
4.3.6
クエン酸の効果53
4.3.7
コハク酸の効果53
4.4
考察54
第
5
章 イヌ脳PFK-1
の活性調節64
5.1
緒言65
5.2
材料および方法66
5.2.1
材料66
5.2.2
イヌ脳PFK-1
の部分精製66
5.2.3 PFK-1
の活性測定67
5.2.4
タンパク質の定量68
5.3
結果68
5.3.1 pH
の効果68
5.3.2 ATP
の効果69
5.3.3 F-6-P
の効果69
5.3.4 F-2,6-P
2の効果69
5.3.5 AMP
の効果70
5.3.6
クエン酸の効果70
5.4
考察70
第
6
章 総括79
謝辞
83
引用文献
84
1
第
1
章 序 論2
グルコースは,生体を構成する様々な細胞に取り込まれ,アデノシン三リン 酸(ATP)産生の基質となる重要な栄養素である。グルコース輸送体により細
胞内に取り込まれたグルコースがリン酸化され,グルコース
6-リン酸となって
ピルビン酸まで代謝される経路が解糖である。解糖は嫌気的条件下での反応で あり,ピルビン酸はさらに還元されて乳酸が生成する。この解糖の過程では,1
分子のグルコースから2
分子のATP
を産生する。一方,好気的な条件下では,ピルビン酸はミトコンドリア内に輸送され,クエン酸回路,電子伝達系を
経て完全に酸化されて二酸化炭素と水となるが,多くの
ATP
が産生される。そ のため,解糖はあらゆる細胞においてATP
産生の重要な初期代謝過程であり,解糖調節は細胞機能にとって重要となる(図
1-1)
。解糖系は,細胞質に存在する酵素により触媒される代謝系であり,代謝速度 は,ヘキソキナーゼ,ホスホフルクトキナーゼ-1(PFK-1),ピルビン酸キナー
ゼの
3
種の律速酵素により制御されている。PFK-1は,フルクトース6-リン酸
(F-6-P)を
ATP
依存的にリン酸化し,フルクトース1,6-ビスリン酸(F-1,6- P
2)とADP
生成を不可逆的に触媒する酵素であり,解糖調節において中心的な 役割を担っていると考えられている [Dunaway, 1983; Mor et al., 2011; Sola-Pennaet al., 2010]。哺乳類の PFK-1
には筋肉型PFK-M,肝臓型 PFK-L
および血小板型
PFK-P(脳型 PFK-C)の 3
種類のアイソフォームが存在し,それぞれの遺伝3
子によりアイソフォームがコードされる [Vora et al., 1981a; 1981b; 1982; 1983]。
3
種類のPFK
アイソフォームは4
量体を形成し,様々な組織において,活性制 御に関わり,組織特異的な解糖系の調節を行っている [Dunaway et al., 1988;Dunaway & Kasten, 1987]。
糖代謝は,各組織におけるエネルギー要求量やエネルギー源の違いにより,
特徴的な性質を有することが知られている。骨格筋は,好気的条件では解糖か
らクエン酸回路,電子伝達系を介して多量の
ATP
が産生されるのに対し,嫌気 的条件ではピルビン酸から乳酸が生成され,必要なATP
を解糖のみで産生する ことが可能である。さらに,細胞内のグルコースからグリコーゲンを合成し貯 蔵することで,必要に応じグルコースに分解し解糖を介してエネルギーの産生 に利用することも可能である [Evans et al., 2019; Murray, 2016]。肝臓は,血糖維 持において中心的な役割を果たす。摂食時には門脈を介して取り込んだグルコ ースからグリコーゲンを合成し貯蔵し,絶食時にはグリコーゲン分解だけでな く,糖新生により糖質以外からグルコースを産生し血糖を供給することが可能 である [Petersen et al., 2017]。脳は,飢餓状態を除きエネルギー源を血糖に依存 するが,常に十分な酸素供給があり好気的代謝のみを行うため,解糖からクエ ン酸回路,電子伝達系を介して多量のATP
を産生する [Mergenthaler et al.,2013]。また,脳の機能維持のためにグルコースを消費して乳酸産生も行ってい
4
る [Magistretti & Allaman,2018]。
近年,イヌはコンパニオンアニマルとしての地位が確立され,イヌに対して もヒト医学領域に遜色のない診療が提供されてきている。しかし,ヒトと他の 動物種では,解剖学的構造,食性,行動様式が異なることから,糖代謝調節に ついても異なる点があると考えられるが,イヌの糖代謝調節に関する報告は少 ない。そこで,本研究では,イヌのグルコース代謝を明らかにする目的の1つ として,特徴的な糖代謝を有する骨格筋,肝臓,脳に注目し,解糖系の律速酵
素である
PFK-1
について,比活性,アイソフォーム構成,細胞内因子による活性調節について検討した。
第
2
章においては,イヌの骨格筋,肝臓および脳におけるPFK-1
について,酵素活性を測定し比活性を求め,タンパク質発現と
mRNA
発現からアイソフォ ーム構成について比較検討した。第
3
章では,比活性が高く,M型のみのアイソフォームで構成されることが 明らかとなったイヌの骨格筋PFK-1
について,細胞内因子による活性調節につ いて検討した。第
4
章では,比活性が低く,主にL
型と少量のP
型のアイソフォームで構成 されるイヌの肝臓PFK-1
について,細胞内因子による活性調節について検討し た。5
第
5
章では,比活性が高く,3種のアイソフォームが混在するイヌの脳PFK-
1
について,細胞内因子による活性調節について検討した。6
図
1-1.
解糖系とクエン酸回路の概略図7
第
2
章イヌの各種臓器における
PFK-1
の比活性およびアイソフォーム構成8
2.1
緒言解糖は,グルコースの異化によりエネルギーを産生する嫌気的な反応経路で
ある。すべての細胞の細胞質内に存在し,ヘキソキナーゼ,
PFK-1
およびピルビ ン酸キナーゼの3
種の律速酵素が解糖制御に関わっている。PFK-1
は,F-6-P
をATP
依存的にリン酸化し,F-1,6-P
2とADP
生成を不可逆的に触媒する酵素であ り,解糖調節において中心的な役割を担っていると考えられている [Dunaway,1983; Mor et al., 2011; Sola-Penna et al., 2010]。
哺乳類の
PFK-1
には,筋肉型(PFK-M),肝臓型(PFK-L)および血小板型(PFK-P)あるいは脳型(PFK-C)といわれる 3
種類のアイソフォームが存在する。PFK-
1
は,その 3種のアイソフォームを基本単位として4
量体で構成されているが,動物種により,また,臓器により構成が異なっている。これまでに,ラット
[Dunaway & Kasten, 1986],ウサギ [Foe & Kemp, 1985; Kemp, 1971; Tsai & Kemp, 1974],ウシ[Fukushima & Sugiya, 1992]
およびヒト [Dunaway et al., 1988; Vora etal., 1981a; 1981b]
における各種臓器のアイソフォーム構成について,精製したPFK-1
を用いて,電気泳動法による解析がなされている。グルコースは,生体を構成する様々な細胞に取り込まれ,
ATP
産生のための基 質となる重要な栄養素であるが,グルコース代謝は,組織や臓器の独自な機能に 依存してその調節系が異なっている。骨格筋は,安静時には脂肪酸を主なエネル9
ギー源として利用するが,運動時にはグルコースの利用が亢進する。好気的条件
では,グルコースから代謝されたピルビン酸はアセチル
CoA
に代謝され,クエ ン酸回路,電子伝達系を介して多量のATP
が産生される。一方,嫌気的条件で はピルビン酸は乳酸となり,解糖によるATP
産生が優先される。また,細胞内 のグルコースからグリコーゲンを合成し,貯蔵するが,必要に応じてグリコーゲンを分解し,解糖を介して
ATP
産生に利用する [Murray, 2016]。肝臓は,様々な 代謝において重要な役割を担っているが,血糖調節においても中心的な役割を 果たす。グリコーゲン合成能と糖新生能を有し,摂食時はグルコースからグリコ ーゲンを合成し,貯蔵するが,低血糖時はグリコーゲン分解や糖新生によりグル コースを産生し,血糖として供給する [Petersen et al., 2017]。脳は,多量のエネ ルギーを必要とするが,飢餓状態を除きエネルギー源として血糖に依存する。常 に十分な酸素が供給されることから,解糖の代謝産物であるピルビン酸は,ミトコンドリアにてアセチル
CoA
となり,クエン酸回路,電子伝達系を介し多量のATP
が合成される[Mergenthaler et al., 2013]。また一方で,近年,脳ではその機能維持のためのグルコース取込みが亢進し,乳酸産生が亢進することも知られて いる[Cunnane et al., 2011; Magistretti & Allaman, 2018]。
このように臓器や組織による代謝の違いに依存して,各臓器における
PFK-1
のアイソフォーム構成や活性調節が異なるなど,特徴的であることが示唆され10
ている [Dunaway et al., 1988; Dunaway, 1983; Kemp, 1971; Sola-Penna et al., 2010;
Tsai & Kemp, 1974]。
しかし,イヌのPFK-1
に関する報告は少なく[Giger et al.,1988; Mhaskar et al., 1992; Smith et al., 1996a; 1996b; Washizu et al., 1999],各臓器に
おける
PFK-1
については比較検討はなされていない。本章では,イヌのグルコース代謝の一端を理解することを目的とし,グルコース代謝の異なる骨格筋,肝
臓,脳における
PFK-1
の比活性の違いを検討し,mRNAおよびタンパク質の発 現よりアイソフォーム構成について検討した。また,糖新生能を有する肝臓については,
PFK-1
の逆向きとなるF-1,6-P
2からF-6-P
への反応を触媒するフルクトースビスホスファターゼ-1(FBP-1)についても比活性の違いと,mRNAおよびタンパク質発現について検討した。
2.2
材料および方法2.2.1
材料酵素活性の測定に用いたアデノシン一リン酸(AMP),
aldolase
(ALD),glycerol 3-phosphate dehydrogenase
(G3PDH),glucose-6-phosphate dehydrogenase
(G6PDH)は和光純薬工業株式会社 (Osaka, Japan) から購入した。ATP,F-6-P,F-1,6-P2,
NAD, NADP, triosephosphate isomerase
(TPI),phosphoglucose isomerase (PGI)
は
Sigma-Aldrich (St. Luis, MO, USA)
から購入した。ウェスタンブロッティング11
に用いた
SDS
サンプルバッファー(Red Loading Buffer)は New England Biolabs(Ipswich, MA, USA),7.5%ポリアクリルアミドゲル(Mini-PROTEAN TGX ゲ
ル)は
Bio-Rad Laboratories
(Hercules, CA, USA),ポリフッ化ビニリデン(PVDF)膜(Immobilon-P)は
Merck Millipore(Burlington, MA, USA)
,ブロッキング試薬(ブロックエース)は
DS Pharma Biomedical(Osaka, Japan)
,抗PFK-P・PFK-M
抗体(ab204131),抗PFK-L
抗体(ab181064)および抗FBP-1
抗体(ab109732)は
Abcam(Cambridge, UK)
,HRP 結合抗ウサギIgG
抗体および化学発光試薬(ECL Western blotting Analysis System)は
GE Healthcare(Chicago, IL, USA)から
購入した。
real-time RT-PCR
に用いたTRIzol
はThermo Fisher Scientific
(Waltham,MA, USA)
,RNeasy Mini Kit
はQiagen
(Venlo, Netherlands),PrimeScript RT Master Mix, SYBR Premix Ex Taq™II
およびプライマーはTakara Bio Inc.
(Kusatsu,Japan)
から購入した。その他の一般試薬は,和光純薬工業株式会社および
Sigma- Aldrich (St. Louis, MO, USA)から購入した。
2.2.2
供試臓器本研究に使用した骨格筋,肝臓,脳は,日本大学動物実験委員会によって承認 された他の実験に使用された後,ペントバルビタールナトリウム(150 mg / kg体 重)の静脈内投与により人道的に安楽死されたビーグル犬(オス
2
頭,メス2
頭,12
1~2
歳)より,安楽死後,速やかに摘出した。臓器は,摘出後,実験に使用する まで-80°Cで保存した。本研究のプロトコルは,日本大学動物実験委員会(承認 番号:AP13B074-1)によって承認された。2.2.3
細胞質画分の分離および低分子物質の除去実験は
4°C
環境で実施した。骨格筋,肝臓,脳(各300 mg)を外科剪刀で細
切し,ペレットミキサーを用いて,10 mM ジチオトレイトール(DTT)を含む50 mM
トリスリン酸バッファー(pH 8.0)溶液にてホモジナイズした後,
100,000×g
で30
分間,超遠心し,細胞質画分を得た。PD-10
カラム(GE Healthcare)を用いたゲルろ過クロマトグラフィーにより
PFK-1
の活性調節因子となる低分 子を除去した細胞質画分を実験に供した。細胞質画分のタンパク質濃度 はBradford
法により測定した[Bradford, 1976]。2.2.4 PFK-1
の活性測定PFK-1
の活性測定は,活性調節因子の影響を受けない至適条件下で実施した[Fukushima & Sugiya, 1992]。50 mM HEPES
バッファー(pH 8.2),100 mM KCl,6.5 mM MgCl
2,1 mM NH4Cl,5 mM KH
2PO
4,0.3 mM NADH,0.5 units ALD,0.5units G3PDH,5 units TPI,1 mM F-6-P,5 mM ATP
および0.1 mM AMP
を含む混13
合溶液に細胞質画分を加えて
1 mL
とし,25°C にて反応を行い,U-2900 Spectrophotometer(Hitachi High-Technologies Corporation,Tokyo,Japan)を使用
して, NADH の酸化の変化を340 nm
の吸光度の減少で継時的に測定した。NADH
のモル吸光係数から,25°C で1
分あたり1 µmol
のF-6-P
をリン酸化する酵素活性を
PFK-1
活性の1 unit
として示した。2.2.5 FBP-1
の活性測定FBP-1
の活性測定は,50 mM HEPES
バッファー(pH 7.4),100 mM KCl, 5 mM EGTA, 1 mM MgSO
4,1 mM NH4Cl, 0.5 mM NADP, 5 units G6PDH,10 units PGI
および
0.2 mM F-1,6-P
2を含む溶液に,細胞質画分を加えて1 mL
とし,25°Cにて反応を行い,U-2900 分光光度計を使用して, NADP の還元の変化を
340 nm
の吸光度の増加で継時的に測定した[Sawada et al., 2000]。NADP のモル吸光係数から
25°C
で1
分あたり1 µmol
のF-1,6-P
2を加水分解する酵素活性をFBP-1
活 性の1 unit
として示した。2.2.6
ウェスタンブロッティング細胞質画分を
SDS
サンプルバッファーにて95°C 5
分間処理し,7.5%ポリア クリルアミドゲルの各レーンにPFK-1
活性0.3 munits
相当をロードし,電気泳14
動によりタンパク質を分離した。分離されたタンパク質は
PVDF
膜に転写した。PVDF
膜は,ブロッキング試薬で50
分間処理した。その後,PVDF膜は,一次抗体である抗
PFK-P
・PFK-M抗体(1:1,000),抗 PFK-L
抗体(1:500)
,抗FBP- 1
抗体(1:1,000)と120
分間反応させた。洗浄後,PVDF膜を二次抗体であるHRP
結合抗ウサギIgG
抗体(1:10,000)と90
分間反応させた。検出には化学発光を用い,化学発光シグナルを冷却
CCD
カメラ(ImageQuant LAS 4000 mini; GEHealthcare)で測定した。
2.2.7 real-time RT-PCR
骨格筋,肝臓および脳から
TRIzol(Thermo Fisher Scientific)もしくは RNeasy Mini Kit
(Qiagen)を用いてtotal RNA
を抽出し,RNA
チップ電気泳動(Experion™Automated Electrophoresis System, Experion™ RNA StdSens Analysis kit, Bio-Rad Laboratories)にて実験に適切な品質を確認した。 total RNA 500 ng
よりPrimeScript RT Master Mix(Takara Bio Inc.)を用いて逆転写し,cDNA
を合成した。cDNA 2µL,SYBR Premix Ex Taq™II(Takara Bio Inc.),特異的なプライマー(表 2-1)を
加えて総反応量
25 µL
でreal-time PCR
を実施した。サーマルサイクラー(ThermalCyclerDice®Real Time System II TP900,Takara Bio Inc.)を使用して熱変性 95°C 5
秒間,アニーリングおよび伸長反応
60°C 30
秒間のサイクルでPCR
反応を行っ15
た。増幅産物について,解離曲線で単一のピーク,アガロースゲル電気泳動で単
一のバンドを示すことを確認した。
PCR
増幅効率は,連続希釈したcDNA
を基 に作成した検量線の勾配から計算し,定量に適していることを確認した。 real-time PCR
の結果は,ソフトウェア(Thermal Cycler Dice®Real Time System Software Ver.5.11B, Takara Bio Inc.)で second derivative method
およびcomparative cycle
threshold (ΔΔCt) method
を使用して分析した。ハウスキーピング遺伝子は,BestKeeper
ソフトウェアを使用して[Okabayashi et al., 2019],Ct値の標準偏差に 基づき評価し,GAPDH,ACTB,RPS18,およびTBP
の値は1.15,1.43,0.69,
およびそれぞれ
1.34
であったため,RPS18
を内部標準遺伝子として相対的mRNA
発現量を算出した。2.3
結果2.3.1 PFK-1
比活性各臓器における
PFK-1
の比活性は,骨格筋0.309±0.044 units/mg protein(平均
値±標準誤差),肝臓0.038±0.0078 units/mg protein,脳 0.290±0.033 units/mg protein
であった(図2-1)
。2.3.2 FBP-1
比活性16
肝臓における
FBP-1
の比活性は,0.057±0.0132 units/mg protein
であった(図2-2)
。データとしては示さないが,骨格筋および脳にはFBP-1
の活性は認められ なかった。2.3.3 PFK-1
アイソフォームおよびFBP-1
のタンパク質発現ウェスタンブロッティング
ウェスタンブロッティングの結果を図
2-3
に示す。PFK-1
の各アイソフォーム について,骨格筋ではM
型,肝臓ではP,L
型,脳ではP,M,L
型が検出され た。また,肝臓ではFBP-1
が検出された。2.3.4 real-time RT-PCR
各臓器における内部標準遺伝子
RPS18
のmRNA
発現量には差はなかった(図2-4)
。また標的遺伝子の増幅効率および定量性について検討し,すべて適正な範囲であった(図
2-5)
。real-time RT-PCR
の結果を図2-6
に示す。PFK-1
の各アイソフォームについて,骨格筋ではほぼ
M
型のみ,脳ではM, P, L
型の順で,肝臓ではL
型が主でP
型 とわずかにM
型が認められた。また,肝臓においてFBP-1
のmRNA
発現が確認 された。17
2.4
考察至適条件下における
PFK-1
活性は,活性調節因子の影響を排除した最大活性 を示しており,骨格筋や脳で比活性が高く,FBP-1
活性を有する肝臓では比活性 が低いことが明らかとなった。本章で得られた結果から,イヌの骨格筋
PFK-1
はM
型のみで構成されている ことが明らかとなり,他の動物における過去の報告と一致した[Dunaway et al.,1988; Dunaway & Kasten, 1987; Fukushima & Sugiya, 1992; Kemp, 1971]。骨格筋は
安息時/運動時でエネルギー要求量が大きく変化し,PFK-1 のアイソフォームが
M
型のみであることが運動状況等に応じた活性調節に適していると示唆され た。雑食性のヒトおよびラットの肝臓
PFK-1
では,L,M,P 型の順で構成され[Dunaway et al., 1988; Dunaway & Kasten, 1987],草食性のウサギおよびウシでは
主に
L
型で構成されると報告されている[Fukushima & Sugiya, 1992; Kemp, 1971]。イヌの肝臓
PFK-1
は主にL
型および少量のP
型で構成されており,肝臓PFK-1
のアイソフォーム構成と食性には関連性があることが示唆された。また,糖新生 能を有することから,解糖と糖新生のバランスを調節する因子により活性調節 がなされることが示唆された。18
イヌの脳
PFK-1
は, P,M,L型の順で構成されており,他の動物と同様,3種のアイソフォームにより構成されていることが明らかとなった[Dunaway et al.,
1988; Dunaway & Kasten, 1987; Fukushima & Sugiya, 1992; Kemp, 1971]。これは,
常に嫌気的および好気的にそれを消費する臓器という特徴がアイソフォーム構 成にも反映していると考えられる。
19
表
2-1. Real-time PCR
に用いたプライマーの配列Symbol GenBank accession no. Primer sequences Length (bp) RPS18 NM_001048082.1 F: ATAGCCTTTGCCATCACAGCAATTA
R: TTGGTGAGATCGATGTCTGCTTTC 86
PFK-M NM_001003199.1 F: GCTGACACAGCCCTCAACACTATC R: CGCAGTAGCCACCCATAGTTTC 109
PFK-L XM_544922.6 F: AGACCTGAAAGCCAACGTGGAG R: GGCCCAGAACGTTGGTCCTA 173
PFK-P XM_005617183.3 F: GTCCAACGTGGAGCACCTGA
R: CCTCGGAGTACAGCTGGTAGATGAA 122
20
図
2-1.
イヌ骨格筋,肝臓および脳におけるPFK-1
の比活性イヌ骨格筋,肝臓および脳の細胞質画分の
PFK-1
活性をpH 8.2
の至適条件で測定した。
PFK-1
活性は,1
分間あたりの340 nm
の吸光度の減少を測定し,NADH
のモル吸光係数から
unit
を算出し,タンパク質濃度当りの比活性をとして示し た。値は4
例の平均値±
標準誤差を示す。21
図
2-2.
イヌ肝臓におけるFBP-1
の比活性イヌ肝臓の細胞質画分の
FBP-1
活性をpH 7.4
の至適条件で測定した。FBP-1
活 性は,1分間あたりの340 nm
の吸光度の増加を測定し,NADPのモル吸光係数 からunit
を算出し,タンパク質濃度当りの比活性をとして示した。値は4
例の 平均値±標準誤差を示す。22
(a)
(b)
図
2-3.
イヌ骨格筋,肝臓および脳PFK-1
アイソフォームおよびFBP-1
のタンパク質発現
(a)
イヌ骨格筋,肝臓および脳において,約0.3 munits
のPFK-1
活性を有する細 胞質画分を用い,PFK-P, -M およびPFK-L
に対する特異的抗体を用いたウェス タンブロッティングによりPFK-1
アイソフォームタンパク質発現の検討を行っ た。(b) FBP-1に対する特異的抗体を用いたウェスタンブロッティングにより,イヌ肝臓の細胞質画分の
FBP-1
タンパク質発現の検討を行った。独立に繰り返 し実施した3
例中の代表的な結果を示す。23
図
2-4.
イヌ骨格筋,肝臓および脳における内部標準遺伝子RPS18
のPCR
標準曲線
イヌ骨格筋
(
●)
,肝臓(
▲)
および脳(
■)
から抽出したtotal RNA
から合成したcDNA
をそれぞれ階段希釈したサンプルを用いてRPS18 mRNA
発現をreal-time
RT-PCR
にて検討した。横軸は希釈率,縦軸はCt
値を示す。PCR
効率は標準曲線の傾きから計算し,相関係数
(R
2)
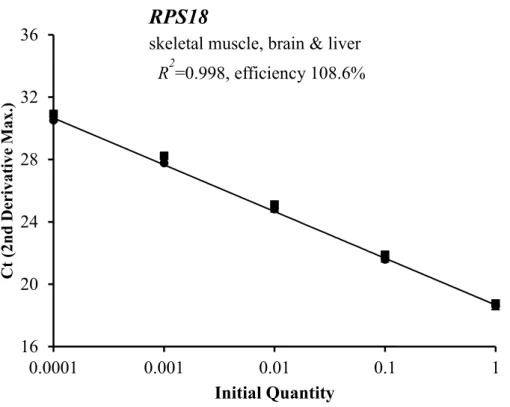
は近似線より求めた。各臓器間に差はなかっ た。16 20 24 28 32 36
0.0001 0.001 0.01 0.1 1
Ct (2nd Derivative Max.)
Initial Quantity
RPS18
skeletal muscle, brain & liver
R
2=0.998, efficiency 108.6%
24
図
2-5.
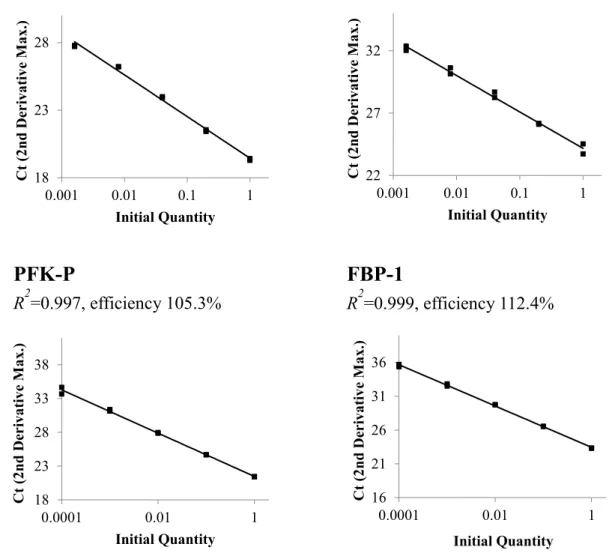
標的遺伝子の増幅効率および定量性PFK-1
アイソフォームmRNA
発現については脳より抽出したtotal RNA,FBP-1
については肝臓より抽出した
total RNA
から合成したcDNA
をそれぞれ階段希 釈したサンプルをreal-time RT-PCR
にて検討した。横軸は希釈率,縦軸はCt
値 を示す。PCR
効率は標準曲線の傾きから計算し,相関係数(R2)は近似線より求め
た。すべてのプライマーが定量性を示した。18 23 28
0.001 0.01 0.1 1
Ct (2nd Derivative Max.)
Initial Quantity
22 27 32
0.001 0.01 0.1 1
Ct (2nd Derivative Max.)
Initial Quantity
16 21 26 31 36
0.0001 0.01 1
Ct (2nd Derivative Max.)
Initial Quantity 18
23 28 33 38
0.0001 0.01 1
Ct (2nd Derivative Max.)
Initial Quantity
FBP-1
R
2=0.999, efficiency 112.4%
PFK-M
R
2=0.994, efficiency 111.1%
PFK-P
R
2=0.997, efficiency 105.3%
PFK-L
R
2=0.991, efficiency 120.1%
25
図
2-6.
イヌ骨格筋,肝臓および脳におけるPFK-1
アイソフォームおよび肝臓における
FBP-1
のmRNA
発現イヌ骨格筋,肝臓および脳における標的遺伝子の
PFK-1
アイソフォームmRNA
の相対発現量をreal-time RT-PCR
にて検討した。値は3
例の平均値±標準誤差を 示す。skeletal
muscle liver
liver
brain
26
第
3
章イヌ骨格筋
PFK-1
の活性調節27
3.1
緒言哺乳類の
PFK-1
には筋肉型(M),肝臓型(L)および血小板型(P)の3
種類のアイソフォームが存在し,様々な臓器でその臓器の特徴を反映した
4
量体で 構成されるが [Sola-Penna et al., 2010],骨格筋はM
型のアイソフォームのみで構 成される [Dunaway & Kasten, 1987; Dunaway et al., 1988]。PFK-1は様々な細胞内因子によりアロステリックな活性調節が行われるが,骨格筋の
PFK-1
において も活性調節についての検討がなされ [Kemp & Foe, 1983],さらにラットM
型PFK-1
アイソフォームのcDNA
クローニングがなされ,基質であるATP
やF-6-
P
の結合部位や活性促進因子や活性抑制因子の反応部位のアミノ酸配列が明ら かとなった [Ma et al., 1996]。近年,ウサギ [Banazak et al., 2011]およびヒト [Klooset al., 2014]の M
型PFK-1
アイソフォームの結晶構造が報告され,酵素触媒部位や活性促進および抑制部位の詳細な構造がさらに明らかにされつつある。
ヒト医学領域では,垂井病として知られるヒト
M
型PFK-1
活性を欠損した糖 原病があり,症状として運動不耐,運動時有痛性筋けいれん,横紋筋融解症,ミ オ グ ロ ビ ン 尿 症 お よ び 溶 血 を 特 徴 と す る 常 染 色 体 劣 性 遺 伝 性 疾 患 で あ る[Brüser et al., 2012b; Sherman et al., 1994 ; Tarui et al., 1965]。獣医学領域において
も,イングリッシュスプリンガースパニエルやウィペットにおいて,M型PFK-
1
のナンセンス変異による慢性溶血と労作性ミオパシーを伴う遺伝性疾患が報28
告されており [Gerber et al., 2009; Giger et al., 1985; Inal et al., 2012; Vora et al., 1985],
イヌにおいても
M
型PFK-1
アイソフォームの重要性が報告されている。本章では,イヌの骨格筋から部分精製された
PFK-1
を使用して,活性調節因 子による効果について検討した。3.2
材料および方法3.2.1
材料ATP,アデノシン二リン酸(ADP)
,AMP,ウリジン三リン酸(UTP)
,環状AMP
(cAMP),クエン酸,
ALD,および G3PDH
は,和光純薬工業株式会社(Osaka,Japan)から購入した。 F-6-P, F-1,6-P
2,D-フルクトース-1,2-サイクリック-6-二リ
ン酸(F-1,2c-6-P2),NAD, phenylmethylsulfonyl fluoride
(PMSF)およびTPI
は,Sigma-Aldrich(St. Louis, MO, USA)から購入した。Blue Sepharose 6 Fast Flow
(Cibacron Blue)は,
GE Healthcare
(Little Chalfont,UK)から購入した。 Bio-Rad Protein Assay Dye Reagent Concentrate
はBio-Rad(Hercules, CA, USA)から入手し
た。 ウシ血清アルブミン(BSA)ははRoche(Mannheim, Germany)から入手し
た。フルクトース
2,6-ビスリン酸(F-2,6-P
2)は,F-1,2c-6-P
2から合成した [Uyedaet al., 1981b]。F-2,6-P
2のモル濃度は,リン濃度を測定し [Itaya & Ui, 1966],それに基づいて算定した。その他の試薬は,和光純薬工業株式会社および
Sigma-
29
Aldrich
から入手した。3.2.2
供試臓器第
2
章と同様に,日本大学動物実験委員会によって承認された他の実験にお いて,ペントバルビタールナトリウム(150 mg / kg体重)の静脈内投与により人道的に安楽死されたビーグル犬(オス
2
頭,メス2
頭,1∼2
歳)より摘出した骨 格筋を-80°Cで保存した後に使用した。本研究のプロトコルは,日本大学動物実 験委員会(承認番号:AP13B074-1)によって承認された。3.2.3 PFK-1
部分精製イヌの骨格筋
PFK-1
の部分精製は,既に報告された方法 [Fukushima & Sugiya,1992]
を一部改変して行った。熱処理を除き,すべての精製ステップは4°C
環境で実施した。外科剪刀で
15 g
の骨格筋を細切し,テフロンホモジナイザーを用 いて,1 mM PMSFおよび10 mM DTT
を含む50 mM
トリスリン酸バッファー(pH 8.0)溶液にて
500 rpm
で5
分間ホモジナイズし,100,000×gで30
分間,超遠心し,細胞質画分を得た。得られた細胞質画分を
52°C
で3
分間加熱処理した後,
10,000×g
で10
分間遠心し,変性タンパク質を除去した。熱処理した上清を,0.1 mM EDTA,0.05 mM F-1,6-P
2および10 mM DTT
を含む50 mM
トリスリン酸30
バッファー(pH 8.0)溶液(A バッファー溶液)で平衡化した
Blue Sepharose 6 Fast Flow(Cibacron Blue)カラム(1×5 cm)にロードした。 A
バッファー溶液に
0.15 mM ADP
を加えた溶液(50 mL)でカラムを洗浄後,A
バッファー溶液に5 mM ATP
および2 mM F-6-P
を加えた溶液(Bバッファー溶液)を流速1 mL /
min
で流し,PFK-1を溶出した。PFK-1活性を有する画分を回収し,50%硫酸ア
ンモニウムでタンパク質を沈殿させ,
10,000×g
で30
分間遠心分離した。沈査画分を
10 mM DTT
を含む50 mM
トリスリン酸バッファー(pH 8)溶液に溶解した後,硫酸アンモニウムを除去するために透析を行い,透析された溶液を部分精
製
PFK-1
として使用した。3.2.4
タンパク質濃度測定PFK-1
部分精製における各ステップで得られたPFK-1
画分について,ウシ血清アルブミンを指標に
Bradford
法 [Bradford, 1976]でタンパク質濃度を測定した。3.2.5 PFK-1
の活性測定至適条件での
PFK-1
活性(Vmax)は,50 mM HEPESバッファー(pH 8.2),100 mM KCl,6.5 mM MgCl
2,1 mM NH4Cl,5 mM KH
2PO4,0.3 mM NADH,0.5
units ALD,0.5 units G3PDH,5 units TPI,1 mM F-6-P,5 mM ATP
および0.1 mM
31
AMP
を含む混合溶液に部分精製PFK-1
を加えて最終容量を1 mL
とし,25°Cで 反応を行った。U-2900 Spectrophotometer(Hitachi High-Technologies Corporation)を使用して,
NADH
の酸化を340 nm
の吸光度の減少で継時的に測定した。NADH
のモル吸光係数から,25°Cで1
分あたり1 μmol
のF-6-P
をリン酸化する酵素活性を
PFK-1
活性の1 unit
と定義した。PFK-1
の活性調節因子の効果の検討においては, 50 mM HEPES バッファー(pH 7.3),100 mM KCl,6.5 mM MgCl2,1 mM NH4
Cl,5 mM KH
2PO
4,0.3 mMNADH, 0.5 units ALD, 0.5 units GDH, 5 units TPI
および濃度の異なるF-6-P, ATP
および
AMP
を含む混合溶液に部分精製PFK-1
を加え,容量を1 mL
とし,25°C で反応を行い,340 nmの吸光度の減少を継時的に測定した。ここで得られた値 をv
とし,至適条件下で得られた最大反応速度(Vmax)に対する比率v/V
maxとし て表した [Uyeda et al., 1981a; Uyeda & Racker, 1965]。3.3
結果3.3.1
イヌ骨格筋PFK-1
の精製度イヌ骨格筋の細胞質画分を分離し,そこから
PFK-1
を部分精製した。表3-1
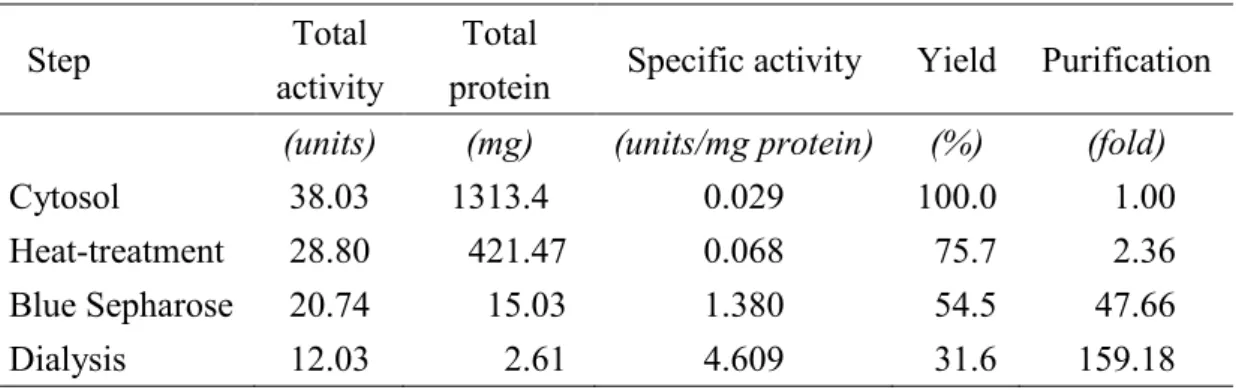
に 精製の各ステップのタンパク質濃度とPFK-1
活性を示す。イヌ骨格筋PFK-1
は,細胞質画分より
159
倍精製され,比活性は4.6 units /mg protein,回収率は 31.6%
32
であった。
3.3.2 pH
の効果部分精製されたイヌ骨格筋
PFK-1
の活性を1 mM F-6-P,5 mM ATP
および0.1
mM AMP
の条件下にてpH 7.0〜8.8
で測定し, pH依存性を検討した。 その結果,図
3-1
に示すように,pH 8.2
で最も高い活性が認められたことから,イヌの 骨格筋PFK-1
の至適pH
は8.2
であった。3.3.3 ATP
の効果PFK-1
の基質の1
つであるATP
の効果を検討した。0.1 mM F-6-P および0.1
mM AMP
の条件下で,ATP濃度を0∼10 mM
と変化させイヌ骨格筋PFK-1
活性を検討したところ,低濃度(<1 mM)の
ATP
は活性を亢進し,さらに濃度を上 げると活性は抑制され,5 mM ATPでほぼ完全に阻害した(図3-2)
。3.3.4 F-6-P
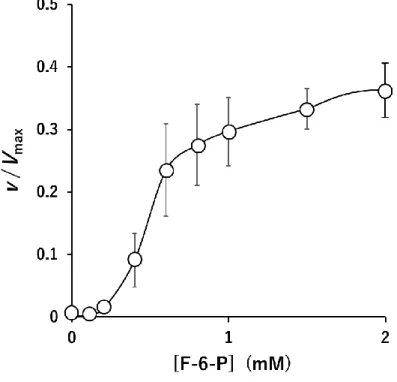
の効果PFK-1
のもう1
つの基質であるF-6-P
の効果を検討した。5 mM ATP
および0.1
mM AMP
の存在下の活性抑制がなされた条件下で,F-6-P
濃度を0∼2 mM
と変化させイヌ骨格筋
PFK-1
活性を検討したところ,図3-3
に示すようにF-6-P
の濃度33
に依存してシグモイド型に活性は上昇した。イヌ骨格筋
PFK-1
活性は1 mM F- 6-P
でほぼプラトーレベルに達した。3.3.5 F-2,6-P
2の効果PFK-1
の強力な活性化因子とされているF-2,6-P
2の効果について検討した。0.1
mM F-6-P,5 mM ATP
および0.1 mM AMP
の存在下の活性抑制がなされた条件下で,F-2,6-P2濃度を
0∼90 μM
と変化させイヌ骨格筋PFK-1
活性を検討したところ,図
3-4
に示すように,用量依存的に活性の亢進が認められた。3.3.6 AMP
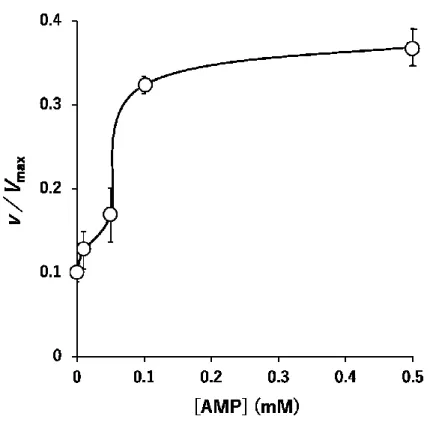
の効果PFK-1
の活性化因子の1
つとされているAMP
の効果について検討した。0.1mM F-6-P
および2.5 mM ATP
の存在下の活性抑制がなされた条件下で,AMP濃度を
0∼0.5 mM
と変化させイヌ骨格筋PFK-1
活性を検討したところ,図3-5
に示すように,用量依存的に活性の亢進が認められ,
0.1 mM AMP
でプラトーに達し た。3.3.7 cAMP
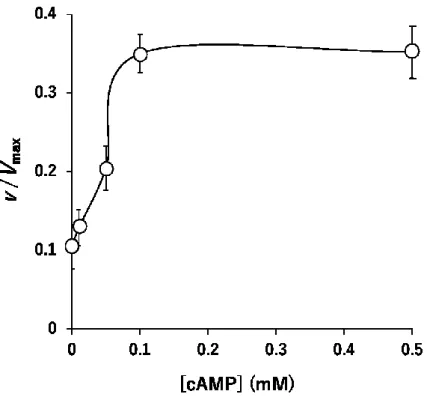
の効果細胞内情報伝達に関わる分子
cAMP
の効果について検討した。0.1 mM F-6-P34
および
2.5 mM ATP
の存在下の活性抑制がなされた条件下で,また,cAMP分解酵素ホスホジエステラーゼの阻害のために
1 mM 3-イソブチル-1-メチルキサン
チンを加えたcAMP
濃度を0∼0.5 mM
と変化させイヌ骨格筋PFK-1
活性を検討 したところ,図3-6
に示すように,AMP と類似した用量依存的な活性の亢進が 認められた。3.3.8
クエン酸の効果PFK-1
の抑制因子の1
つとされているクエン酸の効果について検討した。0.1mM F-6-P
および0.1 mM AMP
の存在下でATP
を0.3 mM
として活性が認められる条件下で,クエン酸濃度を
0∼5 mM
と変化させイヌ骨格筋PFK-1
活性を検討 したところ,図3-7
に示すように,用量依存的に活性が阻害された。3.3.9 UTP
の効果ヌクレオシド三リン酸の
1
つであるUTP
はATP
のように代謝反応の基質とし ての機能を有することから,UTPの効果について検討した。0.1 mM F-6-P および
0.1 mM AMP
の存在下で,ATPの代わりにUTP
濃度を0∼10 mM
と変化させ たところ,図3-8
に示すように,UTPは低濃度でイヌ骨格筋PFK-1
活性を亢進 し,高濃度では活性を抑制し,ATP
に類似した効果を示した。しかし,活性の亢35
進は
ATP
に比べ低濃度(<0.5 mM)で認められ,また,5 mMでは最大活性の約 半分の活性を示が認められ,抑制するのには10 mM
を要した。3.4
考察本章では,イヌ骨格筋から
PFK-1
を部分精製し,活性調節因子の効果につい て検討した。PFK-1のM
型アイソフォームには,酵素触媒部位,活性促進部位および活性抑制部位と
3
つのアデニンヌクレオチド結合部位が 存在する[Banaszak et al., 2011; Brüser et al., 2012a]。イヌ骨格筋の PFK-1
は,構成するアイソフォームが
M
型のみであることから,骨格筋PFK-1
の活性調節は,M
型アイ ソフォームの特徴を反映していると考えられる。ATP
は,高エネルギーリン酸化合物でありエネルギー代謝の目的生成物であ りながら,PFK-1 による反応においてリン酸供与体としてだけでなく活性抑制因子としての機能も有する。本章の結果から,ATP は低濃度では
PFK-1
の触媒部位に結合し,高濃度では活性抑制部位に結合することで,活性を抑制すること
が示唆された。ラットやウサギの
M
型PFK-1
アイソフォームにはATP
による 活性部位と抑制部位が存在し,活性調節に関わることから [Ma et al., 1996;Banaszak et al., 2011],イヌも同様な活性部位を有していると考えられる。
36
イヌ骨格筋
PFK-1
は,ATPで阻害された活性をF-6-P,AMP,F-2,6-P
2は解除 したことから,これらは活性調節因子として機能することが示唆された。また,cAMP
もAMP
と同様な効果を示した。M
型PFK-1
アイソフォームはリン酸化部位を有することから
cAMP
依存性リン酸化酵素による活性調節が考えられたが,リン酸化状態でも脱リン酸化状態でも調節因子の効果は明確でないことから,
リン酸化による調節は未だ不明である [Foe & Kemp, 1982; Kemp & Foe, 1983]。
本研究で使用した
cAMP
濃度も高く,AMP と同様な効果を示したことから,cAMP
の効果はリン酸化を介するものではなく,AMP の結合部位に作用したも のと考えられる。F-2,6-P
2 によるアロステリックな触媒部位は他の活性化因子と異なっており,この部位への
F-2,6-P
2の結合はM
型PFK-1
の活性化にとって重要である [Ma etal., 1996; Banaszak et al., 2011]。ラット M
型PFK-1
においてF-2,6-P
2は1∼2 μM
でV
maxの50%程度まで ATP
抑制を解除し,また抑制因子の効果も解除する [Uyedaet al., 1981a]が,イヌ骨格筋 PFK-1
はさらに高濃度のF-2,6-P
2を必要とし,ATP阻害の解除の割合も
V
maxの30%であることから,他種の M
型PFK-1
とは異なっ た性質を有するかもしれない。内在性のF-2,6-P
2 はホスホフルクトキナーゼ-2(PFK-2)/フルクトース
2,6-ビスホスファターゼ(FBP-2)により産生調節がな
されることから [Rider et al., 2004],PFK-2の発現の検討も必要と思われる。37
イヌ骨格筋
PFK-1
アイソフォームにおいて,クエン酸は抑制因子として機能 することが示された。ラットやウサギのM
型PFK-1
においてもクエン酸の阻害 部位が同定されており,イヌにおいても同様な構造の存在が考えられる。イヌ骨格筋
PFK-1
アイソフォームにおいて,UTP
は基質であるATP
と同様に 低濃度では活性化因子として,また,高濃度では抑制因子として活性調節に関わることが示唆された。
UTP
はATP
より低濃度でイヌ骨格筋PFK-1
を活性化した ことから,PFK-1のATP
による触媒部位に結合し,ATPよりも効率的なリン酸供与体として機能すると考えられる。しかし,抑制効果には
ATP
より高い濃度 が必要であったことから,ATP と比べて活性抑制部位には結合しにくいと考え られる。M
型PFK-1
アイソフォームは,アクチン [Liou & Anderson, 1980] ,カルモジ ュリン [Marinho-Carvalho et al., 2006],カベオリン [Vallejo & Hardin, 2005] など のタンパク質と結合して活性が修飾されることが知られており,それが骨格筋 の機能に関わるグルコース代謝の調節に関わっていると考えられていることか ら,イヌ骨格筋のPFK-1
の活性化における他の調節因子も明らかにすることが 必要と考えられる。38
表
3-1.
イヌ骨格筋PFK-1
の精製Step Total
activity
Total
protein Specific activity Yield Purification
Cytosol
(units) 38.03
(mg) 1313.4
(units/mg protein) 0.029
(%) 100.0
(fold)
1.00
Heat-treatment 28.80 421.47 0.068 75.7 2.36
Blue Sepharose 20.74 15.03 1.380 54.5 47.66
Dialysis 12.03 2.61 4.609 31.6 159.18
39
図
3-1.
イヌ骨格筋PFK-1
に対するpH
の効果イヌ骨格筋から部分精製した
PFK-1
活性についてpH
を変化させて測定した。PFK-1
活性は1
分間あたりの340 nm
の吸光度の減少で示した。pH 8.2
で最も高い活性が認められた。値は
3
例の平均値±
標準誤差を示す。7.0 7.6 8.2 8.8
40
図
3-2.
イヌ骨格筋PFK-1
活性に対するATP
の効果イヌ肝臓から部分精製した
PFK-1
の活性を,pH 7.3
および0.1 mM F-6-P
の条件 下においてATP
濃度を0
∼10 mM
まで変化させ,測定した。低濃度ではPFK-1
活 性の活性化されたが,高濃度では阻害がみられ,5 mM
でほぼ完全に阻害された。値は
3
例の平均値±
標準誤差を示す。41
図
3-3.
イヌ骨格筋PFK-1
に対するF-6-P
の効果イヌ骨格筋から部分精製した
PFK-1
の活性を,pH 7.3
および5 mM ATP
の条件 下でF-6-P
濃度を0
∼2 mM
まで変化させ,測定した。F-6-P
の濃度依存的にPFK-
1
に対するATP
阻害が解除された。値は3
例の平均値±
標準誤差を示す。42
図
3-4.
イヌ骨格筋PFK-1
に対するF-2,6-P
2の効果イヌ骨格筋から部分精製した
PFK-1
の活性を,pH 7.3
,0.1 mM F-6-P
,5 mM ATP
および0.1 mM AMP
の条件下でF-2,6-P
2濃度を0-90 μM
まで変化させ,測定した。
F-2,6-P
2の濃度依存的にPFK-1
に対するATP
阻害が解除された。値は3
例の平均値±標準誤差を示す。
43
図
3-5.
イヌ骨格筋PFK-1
活性に対するAMP
の効果イヌ骨格筋から部分精製した
PFK-1
活性を,pH 7.3
,0.1 mM F-6-P
および2.5 mM
ATP
の条件で測定した。PFK-1
に対するATP
阻害がAMP
により解除された。値 は3
例の平均値±標準誤差を示す。44
図
3-6.
イヌ骨格筋PFK-1
活性に対するcAMP
の効果イヌ骨格筋から部分精製した
PFK-1
活性を,pH 7.3
,0.1 mM F-6-P
および2.5 mM ATP
の条件で測定した。PFK-
1に対するATP
阻害がcAMP
により解除された。値は
3
例の平均値±標準誤差を示す。45
図
3-7.
イヌ骨格筋PFK-1
活性に対するクエン酸の効果イヌ骨格筋から部分精製した
PFK-1
の活性を,pH 7.3
,0.1 mM F-6-P
,0.1 mM
AMP
および0.3 mM ATP
の条件下で,クエン酸の濃度を0
∼5 mM
まで変化させ,測定した。縦軸はクエン酸が存在しないときの活性を