近畿大学学術情報リポジトリ
66
0
0
全文
(2) (英文題目). Functional analysis of Arabidopsis Nudix hydrolases having CoA- and guanosine-3,5-tetraphosphate (ppGpp)-specific pyrophosphohydrolase activities Daisuke Ito September, 2012. Graduate School, Kinki University Division of Agricultural Science Major: Applied Bioscience (Advisor: Prof: Shigeru Shigeoka) (和文題目). シロイヌナズナ CoA および. Guanosine-3,5-tetraphosphate. (ppGpp) 特異的 Nudix hydrolase の機能解析. 近畿大学大学院 農学研究科. バイオサイエンス専攻 伊藤. 大輔. (指導:重岡 成 教授) Submitted to the Graduate School, Kinki University, to fulfill the requirement for the Doctorate Degree..
(3) Acknowledgements. I wish to express my science graduate to Dr. Shigeru Shigeoka, Professor of Faculty of Agriculture, Kinki University, for his kind guidance, valuable advice, stimulating discussion and critical review throughout the work including the manuscript of this thesis. I am grateful to Dr. Tamo Fukamizo and Dr. Tsutomu Kawasaki, Professors of Faculty of Agriculture, Kinki University, for reading the entire text in its original form. I am thankful to Dr. Kazuya Yoshimura, College of Bioscience and Biotechnology, Chubu University, for his valuable help, kind suggestion, and stimulating discussion throughout the work. I wish to thank Dr. Masahiro Tamoi, Faculty of Agriculture, Kinki University, for their valuable discussions and suggestions throughout the work. Finally, special thanks are due to Takahiro Kato, for his many helpful collaborations. Thanks are also due to all the past and present members of our laboratory of Plant Molecular Physiology in the Faculty of Agriculture, Kinki University, for their kind cooperations. This research was supported by a Research Fellowships of the Japan Society for the Promotion of Science for Young Scientists..
(4) Abbreviations. ACP. acyl carrier protein. ACPS. ACP synthase. ADP-glucose. Adenosine 5’-diphosphoglucose. ADP-glucose. Adenosine 5’-diphosphoribose. ApnA. diadenosine polyphosphate. AtNUDX. Arabidopsis Nudix hydrolase. CoA. Coenzyme A. CRSH. Ca2+-activated RelA-SpoT homolog. DHNTP. dihydroneopterin triphosphate. DIP. diphosphoinositol polyphosphate. DPCK. dephospho-CoA kinase. EST. expressed sequence tag. FAD. Flavin adenine dinucleotide. GFP. green fluorescent protein. mTP. mitochondrial targeting peptide. NADPH-GAPDH. NADPH-dependent glyceraldehyde-3-phosphate dehydrogenase. NDP. nucleoside diphosphate. Nudix. nucleoside diphosphates linked to some moiety X. PANK. pantothenate kinase. pGp. ganosine 3’,5’-bisphosphate. P-pant. 4’-phosphopantetheinyl. PPAT. 4’-phosphopantetheine adenylyltransferase. PPC. 4’-phospho-N-panthothenoyl-cysteine. PPCD. PPC decarboxylase.
(5) PPCS. PPC synthase. ppGpp. guanosine 3’,5’-bispyrophosphate. PQ. paraquat. PTS1. type-1 peroxisomal targeting signal. RSH. RelA-SpoT homolog. SDH. succinate dehydrogenase. SPT. serine:pyruvate aminotransferase. TCA. tricarboxylic acid.
(6) CONTENTS. CHAPTER. I. Introduction. CHAPTER. II. Molecular. 1. characterization. of. Arabidopsis. pyrophosphohydrolases, AtNUDX11, 15, and 15a. CHAPTER. CHAPTER. III. IV. CoA 6. Identification and molecular characterization of Arabidopsis ppGpp pyrophosphohydrolases. 31. Conclusion. 45. REFERENCES. 47. PUBLICATIONS. 61.
(7) CHAPTER I. Introduction. Nudix hydrolases with pyrophosphohydrolase activity toward a wide variety of substrates containing a nucleoside diphosphate linked to some other moiety, X, are characterized by a conserved motif (Nudix box, PROSITE PS00893) consisting of 23 amino acids, GX5EX7REUXEEXGU, where U is an aliphatic, hydrophobic residue, although several interesting examples exist with altered consensus sequences (Bessman et al., 1996; McLennan, 2006; Xu et al., 2006).. The Glu residues in the core of the. motif, REUXEE, serve to modulate the reaction as the effector of catalysis and the ligand to the enzyme bound metal ion cofactor.. In most cases, Mg2+ is likely to be the. most physiologically relevant. More than 1800 open reading frames coding for Nudix hydrolases in over 360 different species have been identified by bioinformatics searches (Kraszewska, 2008; Gunawardana et al., 2009).. The number of Nudix in each species varies from one in. Mycoplasma sp. to over 50 in eukaryotes (Kraszewska et al., 2008).. For example,. yeast, human, and rice possess 6, 24, and 33, respectively, of genes encoding putative Nudix hydrolases. Enzymatic and molecular analysis revealed that Nudix hydrolases differ in substrate specificity and subcellular localization (Ogawa et al., 2005; 2008; McLennan, 2006; Yoshimura et al., 2007).. Accordingly, the enzymes have activity. hydrolyzing nucleoside di- and tri-phosphates and their oxidized forms, dinucleoside polyphosphates, nucleotide sugars, NADH, and also CoA (Ogawa et al., 2005; 2008; McLennan, 2006; 2009; Kraszewska et al., 2008; Gunawardana et al., 2009; Bessman et al., 1996) (Figure I-1).. Nucleoside diphosphates (NDPs), the mRNA cap,. 5’-triphosphorylated RNA, and guanosine 3’,5’-bispyrophosphate (ppGpp), and non-nucleoside 5-phosphoribosyl. substrates. such. 1-diphosphate. as. diphosphoinositol. (PRPP), 1. thiamine. polyphosphates. pyrophosphate. (DIPs),. (TPP),. and.
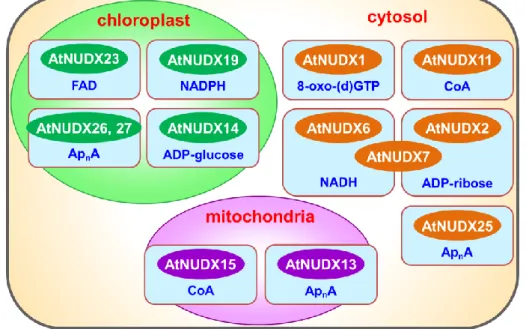
(8) dihydroneopterin triphosphate (DHNTP) are also recognized as substrates for Nudix hydrolases (Fisher et al., 2004; Klaus et al., 2005; Gunawardana et al., 2008; Ooga et al., 2008).. All these substrates are either potentially toxic compounds, cell-signaling. molecules, or metabolic intermediates, the concentrations of which must be closely regulated during the cell cycle and the growth period, and/or in response to environmental conditions.. Hence it has been postulated that the role of Nudix. hydrolases is to suppress or maintain the levels of such molecules (Bessman et al., 1996).. Figure I-1. Nudix hydrolase family in various organisms. Arabidopsis thaliana has 28 genes (AtNUDX1-27, and AtDCP2) encoding Nudix hydrolase homologues located in the cytosol, mitochondria, and chloroplasts (Ito et al., 2012; Ogawa et al., 2005; 2008).. There are various AtNUDXs having. pyrophosphohydrolase activity with a wide range of substrate specificities; 8-oxo-7,8-dihydro-2’-(deoxy) guanosine 5’-triphosphate (AtNUDX1), ADP-ribose 2.
(9) (AtNUDX2 and 7), ADP-glucose (AtNUDX14), long-chain diadenosine polyphosphates (ApnA) (AtNUDX13, 25, 26, and 27), NAD(P)H (AtNUDX6, 7, and 19), coenzyme A (AtNUDX11 and 15), FAD (AtNUDX23), and mRNA caps (AtDCP2) (Ito et al., 2012; Ogawa et al., 2005; 2008; Yoshimura et al., 2007) (Figure I-2).. Figure I-2. Subcellular localization and substrate specificity of AtNUDXs. Notably, there is increasing evidence of regulatory roles for the respective Nudix hydrolases in various physiological processes in plants such as biotic and abiotic stress responses and the metabolism of various molecules (Ishikawa et al., 2009; 2010a; 2010b; Ito et al., 2012; Muñoz et al., 2006; Maruta et al., 2012; Ogawa et al., 2009). It has been reported that AtNUDX1 plays an important role in protection against oxidative DNA and RNA damage through sanitation of oxidized nucleotides in plant cells (Yoshimura et al., 2007).. AtDCP2 acted as the mRNA de-capping enzyme which. is essential for mRNA turnover in postembryonic development (Gunawardana et al., 2009).. It is been reported that AtNUDX2 functions in the maintenance of NAD+ and. ATP levels by nucleotide recycling from ADP-ribose molecules under oxidative stress (Ogawa et al., 2009).. Similarly, AtNUDX7 served to maintain NAD+ levels by. supplying ATP via nucleotide recycling from free ADP-ribose molecules under 3.
(10) oxidative stress (Ishikawa et al., 2009). mechanisms. against. oxidative. In addition, AtNUDX7 regulated the defense. DNA damage. ADP-ribosylation reaction under the stress.. via. modulation. of. the. poly. On the other hand, AtNUDX6 modulated. the plant immune response as a positive regulator of NPR1-dependent SA signaling pathways, through regulation of NADH levels (Ishikawa et al., 2010).. AtNUDX14. regulated intracellular ADP-Glucose levels linked to starch biosynthesis (Muñoz et al., 2006).. More recently, it was reported that AtNUDX23 is involved in the flavin. homeostasis in plant cells via negative feedback regulation of the metabolism of flavins through the hydrolysis of FAD in plastids (Maruta et al., 2012). However, the substrates and physiological functions of a lot of AtNUDXs are still unclear.. Especially, Nudix hydrolases with pyrophosphohydrolase activity toward. CoA and its derivatives, such as acetyl-CoA and succinyl-CoA, have been identified in various organisms, including yeasts (Saccharomyces cerevisiae) (Cartwright et al., 2000), nematodes (Caenorhabditis elegans) (AbdelRaheim et al., 2002), mice (Gasmi et al., 2001; Reilly et al., 2008; Ofman et al., 2006), and plants (Arabidopsis thaliana) (Ogawa et al., 2005; 2008), suggesting that these enzymes are well conserved in various organisms across the evolutionary process.. This strongly suggests the importance of. CoA pyrophosphohydrolase in the metabolism of CoA and its derivatives.. However,. knowledge of the physiological functions of CoA pyrophosphohydrolase is still limited, since there is no direct evidence linking the actions of the enzyme with cellular processes. Recently, it was reported that guanosine 3’,5’-bispyrophosphate (ppGpp) is recognized as a substrate for a Nudix hydrolase, Ndx8, in Thermus thermophilus (Ooga et al., 2009). ppGpp is a low-molecular-weight effector that helps bacteria survive in limited environments, and therefore is called the bacterial alarmone (Xu et al., 2009; Stephens et al., 1975; Magnusson et al., 2005).. The level of ppGpp in Escherichia. coli (E. coli) increases 10-100-fold during amino acid starvation compared to the level present in exponentially growing cells (Rhaese et al., 1975; Lemons et al., 2008; 4.
(11) Nascimento et al., 2008). ppGpp is also accumulated in cyanobacteria under nitrogen starvation (Friga et al., 1981).. ppGpp binds to the β-subunit of RNA polymerase and. induces a structural change (Sato et al., 2009; Chatterji et al., 1998; Tedin et al., 1992; Glass et al., 1986; Wagner et al., 2002).. The, ppGpp accumulated in the cells leads to. an increase in the synthesis of many enzymes required for producing amino acids, and to change in the pattern of expression of various other products (Cashel et al., 1969; Magnusson et al., 2005; Potrykus and Cashel, 2008; Srivatsan and Wang, 2008; Dalebroux et al., 2010).. Notably, it has been reported that the level of ppGpp in plants. is increased by environmental stress, such as drought, salinity, and UV irradiation (Takahashi et al., 2004).. In addition, ppGpp has been found to act as a regulator of. chloroplast RNA polymerase (Sato et al., 2009).. The chloroplast is a semiautonomous. organelle that is originated from not only an ancestral host cell but also a cyanobacterium-like photosynthetic prokaryote.. Therefore, it seems feasible that a. stringent control system by ppGpp similar to that of bacteria exists in plant chloroplasts (Kasai et al., 2002; Givens et al., 2004; Braeken et al., 2006).. Notably, intracellular. ppGpp levels in the Thermus thermophilus ndx8 mutant briefly increased at the end of the logarithmic growth phase and then dropped during the stationary phase, indicating that Ndx8 is involved in the regulation of ppGpp levels during growth phase transition by acting on the degradation of ppGpp (Ooga et al., 2009).. These facts suggest that, in. plant chloroplasts, Nudix hydrolases act as a transcriptional regulator through hydrolysis of ppGpp.. However, it has been unclear about the existence of Nudix. hydrolase(s) having ppGpp pyrophosphohydrolase activity and the contribution of the enzyme(s) to the regulatory mechanism for ppGpp in plants. In this work, to clarify the functional diversity of Nudix hydrolases in higher plants, I studied the following: (1) molecular characterization of Arabidopsis CoA pyrophosphohydrolases, AtNUDX11, 15, and 15a, and (2) identification and molecular characterization of Arabidopsis ppGpp pyrophosphohydrolases.. 5.
(12) CHAPTER II. Molecular characterization of Arabidopsis CoA pyrophosphohydrolases, AtNUDX11, 15, and 15a. Introduction. CoA is required for the production and degradation of both primary and secondary metabolites in bacteria, plants, and animals, where it is involved in more than 100 different reactions (Tilton et al., 2006). five. enzymatic. reactions,. catalyzed. It is synthesized from pantothenate via by. pantothenate. kinase. (PANK),. 4’-phospho-N-pantothenoyl-cysteine (PPC) synthase (PPCS), PPC decarboxylase (PPCD), 4’-phosphopantetheine adenylyltransferase (PPAT), and dephospho-CoA kinase (DPCK) (Tilton et al., 2006; Daugherty et al., 2002; Kupke et al., 2003; Leonardi et al., 2005; Rudio et al., 2006; Spry et al., 2008; Olzhausen et al., 2009) (Figure II-1).. The activities of PANK and PPAT are known to be controlled by the. levels of CoA and CoA thioesters, the end-products of CoA biosynthesis (Leonardi et al., 2005; 2007; Jackowski et al., 1981; Vallari et al., 1987; Rock et al., 2003; Zhang et al., 2005; Miller et al., 2007).. The 4’-phosphopantetheinyl (P-pant) moiety of CoA. can be transferred to a serine residue of acyl carrier proteins (ACPs) by holo-ACP synthase (ACPS) (Lambalot and Walsh, 1995; Flugel et al., 2000; McAllister et al., 2006).. The resulting holo-ACP is active as a central coenzyme of fatty acid. biosynthesis.. CoA is degraded by CoA pyrophosphohydrolase belonging to the family. of Nudix hydrolase, by which 4’-phosphopantetheine and 3’,5’-ADP are produced (Cartwright et al., 2000; Xu et al., 2001; Gasmi and McLennan, 2001; AbdelRaheim and McLennan, 2002; Kang et al., 2003; Ogawa et al., 2005; 2008; Reilly et al., 2008) (Figure II-2).. 6.
(13) Figure II-1.. Biosynthetic pathway of CoA from pantothenate. Figure II-2.. Hydrolysis reaction catalyzed by CoA pyrophosphohydrolase.. CoA pyrophosphohydrolase hydrolyse CoA produce 4’-phosphopantetheine and 3’,5’-ADP. The arrows indicate the site of hydrolysis by CoA pyrophosphohydrolase.. 7.
(14) CoA pyrophosphohydrolases have been identified in various organisms, suggesting the importance of the enzymes in the metabolism of CoA and its derivatives. In fact, it has been reported that the expression of mouse Nudt7α, a peroxisomal Nudix hydrolase having pyrophosphohydrolase activity not only toward CoA but also toward acyl-CoAs, is negatively correlated with the levels of CoA as well as the expression of the enzymes involved in peroxisomal lipid metabolism (Reilly et al., 2008). findings suggest that Nudt7 α CoA/acyl-CoA in the peroxisomes.. These. is tightly associated with the homeostasis of The Arabidopsis Nudix hydrolases AtNUDX11. and AtNUDX15, which were predicted to occur in the cytosol and the mitochondria respectively, had pyrophosphohydrolase activity toward CoA and its derivatives, including not only malonyl-CoA and succinyl-CoA, metabolites of fatty acid biosynthesis in chloroplasts and the TCA cycle in mitochondria, but also oxidized CoA, a potentially toxic or nonfunctional form of CoA (Ogawa et al., 2005; 2008).. Notably,. AtNUDX11-overexpressing Arabidopsis plants showed enhanced growth and an extended life cycle compared to wild-type plants (Kupke et al., 2009).. Hence it has. been suggested that the action of AtNUDX11 is implicated in embryogenesis through the elimination of toxic or useless CoA derivatives.. However, it remains unclear. whether CoA pyrophosphohydrolases impact on the other cell physiology, although they have the potential to hydrolyze various CoA derivatives as the metabolites in diverse cellular processes. In this chapter, I found that the gene encoding AtNUDX15 produced alternatively spliced variants (designated AtNUDX15 and AtNUDX15a).. Hence, to. gain further insight into the physiological roles of plant CoA pyrophosphohydrolases, the regulatory system for gene expression, the kinetic parameters of enzymatic ability, and the intracellular localization of AtNUDX11, 15, and 15a were investigated.. 8.
(15) Materials and Methods. Materials and plant growth conditions Arabidopsis plants (ecotype Columbia) were grown on basic Murashige and Skoog (MS) medium in petri dishes containing 3% (w/v) sucrose for 2 weeks, and then grown in soil at 25°C under long-day conditions (16 h light at 100 μE/m2/s / 8 h dark). The vectors for the Gateway cloning system, pDONR201, and pGWB2, 5, and 6, were obtained from Dr. T. Nakagawa (Shimane University) (2007).. Restriction. enzymes and modifying enzymes were purchased from Takara (Kyoto, Japan).. All. other materials and enzymes were of analytical grade and were obtained from commercial sources.. Analysis of AtNUDX expression in various organs Total RNA was isolated from various organs (0.5 g of fresh weight), rosette leaves, stems, cauline leaves, inflorescences, siliques, and roots of 6-week-old Arabidopsis plants, as described previously (Ogawa et al., 2008). First-strand cDNA was synthesized using ReverTra Ace (Toyobo, Osaka, Japan) with an oligo (dT) primer as described above.. cDNAs encoding the various AtNUDXs and Actin2 were. semi-quantitatively amplified by PCR.. The specific primer sets used in the analysis. were as follows (Table II-1): AtNUDX11, AtNUDX11-F and AtNUDX11-R; AtNUDX15(a), AtNUDX15-15a-F and AtNUDX15-15a-R; Actin2, Actin2-F and Actin2-R. Equal loading of the various amplified gene sequences was determined with the control Actin2 PCR product.. 9.
(16) Table II-1. Primer sequences used for this study Primer names. Sequences (5' to 3'). Notes. AtNUDX11-F. CGCTTTATTCAGATCTGAAG. AtNUDX11-R. CTCAATCAGATCAAGTCAAG. AtNUDX15-15a-F. GTCTACTCACTCTGGAGAAG. AtNUDX15-15a-R. GGAGCTTCAGCTACTGCGTC. Actin2-F. GAGATCCACATCTGCTGG. Actin2-R. GCTGAGAGATTCAGGTGCCC. AtNUDX11-XhoI-F. CTCGAGATGTCTTCAACAAC. XhoI site is in bold. AtNUDX11-XhoI-R. CTCGAGCTCAATCAGATCAA. XhoI site is in bold. AtNUDX15-NcoI-F. CATATGACAACAACAACCCT. NcoI site is in bold. AtNUDX15-XbaI-R. TCTAGAAGCTACTGCGTCTG. XbaI site is in bold. AtNUDX15a-XbaI-R. TCTAGATTCAGCCATTCACA. XbaI site is in bold. attB1-AtNUDX11-cGFP. AAAAAGCAGGCTTGTTAATATGTCTTCAAC. attB site is in bold. attB2-AtNUDX11-cGFP. AGAAAGCTGGGTAAGTTAGGCTGATTCCAG. attB site is in bold. attB1-AtNUDX15-15a-nGFP. AAAAAGCAGGCTTCATGTTTTTGCTTTAC. attB site is in bold. attB2-AtNUDX15. AGAAAGCTGGGTATCATTAAGGCATACAAG. attB site is in bold. attB2-AtNUDX15a. AGAAAGCTGGGTATGCATTCAGCCATTCACA. attB site is in bold. attB1-AtNUDX15-15a. AAAAAGCAGGCTCGCAAATCGGATTCATGT. attB site is in bold. attB2-AtNUDX15-cGFP. AGAAAGCTGGGTAAAGGCATACAAGTATGTT attB site is in bold. attB2-AtNUDX15a-cGFP. AGAAAGCTGGGTAACATTTTAGGGTACTTAA. GFP-F. GTGAGCAAGGGCGAGGAGCT. GFP-R. CTTGTACAGCTCGTCCATGC. attB site is in bold. Expression and purification of the recombinant AtNUDX15a protein cDNA fragments encoding the mature AtNUDX15a protein, except for the predicted mitochondrial transit peptides (mTP) at the N-terminus, were amplified by PCR from first-strand cDNA synthesized from the total RNA of Arabidopsis using specific primers AtNUDX15a-NcoI-F and AtNUDX15a-XbaI-R (Table II-1).. The. amplified DNA fragment was ligated into pT7 Blue T-vector, digested with the desired restriction enzymes, and re-cloned into vector pCold II (Takara).. A recombinant form. of AtNUDX15a was produced using E. coli strain BL21 (DE3) pLysS cells and was purified from the extract using a HiTrap chelating HP column (GE Healthcare, Little 10.
(17) Chalfont, UK) following the manufacturer’s instructions.. The production and. purification of the recombinant AtNUDX11 and 15 were carried out following previous reports (Ogawa et al., 2005; 2008).. The protein content was determined by the. Bradford method (Bradford, 1976), using bovine serum albumin (BSA) as standard. The molecular masses of the recombinant AtNUDX11, 15, and 15a proteins agreed with the predicted values, calculated from the amino acid sequence of the mature protein plus the hexahistidine-tag (AtNUDX11; 27.9 kDa, AtNUDX15; 32.6 kDa, AtNUDX15a; 31.7 kDa).. Enzyme assay and HPLC The hydrolytic activities of recombinant forms of AtNUDX11, 15, and 15a toward CoA derivatives were assayed by a method described previously (Ogawa et al., 2008).. Sixty μL of the reaction mixture, containing 50 mм Tris-HCl (pH 8.0), 5 mм. MgCl2, 5-3,000 μм substrate, and 0.2-1.0 μg of the purified recombinant protein, was incubated at 37°C for 10 min. mм EDTA.. The reaction was terminated by adding 10 μL of 100. The mixture was then analyzed by HPLC using a COSMOSIL C18. column (4.6 x 250 mm, Nacalai tesque, Kyoto, Japan) at a flow rate of 0.6 mL/min for the mobile phase buffer.. It contained 73 mм KH2PO4, 5 mм tetrabutylammonium. dihydrogenphosphate, and 20% methanol.. The substrates (CoA, succinyl-CoA,. malonyl-CoA, lauroyl-CoA, myristoyl-CoA, and oxidized-CoA) and the reaction product (3’, 5’-ADP) were detected by their UV absorbance at 260 nm.. Leaves (0.3 g). of the Arabidopsis plants were homogenized with 1 mL of 100 mм Tris-HCl (pH 8.0) containing 20% glycerol.. After centrifugation (20,000 x g) for 20 min at 4°C, the. supernatant was used for analysis of enzymatic activity.. Crude extracts (approximately. 10.0 μg of protein) were used in this assay.. Subcellular localization of the green fluorescent protein (GFP) fusion protein The vectors for the generation of GFP-fused proteins were constructed using 11.
(18) GATEWAY cloning technology (Invitrogen, Carlsbad, CA, USA).. The cDNAs. encoding the open reading frames of the AtNUDXs were cloned into the donor vector, pDONR201, and then re-cloned into the destination vector, pGWB6 or pGWB5, in which the AtNUDX proteins, fused with GFP at N- or C-terminus, were expressed under the control of the cauliflower mosaic virus 35S promoter.. PCR and in vitro BP. and LR recombination reactions were carried out following the manufacturer’s instructions (Invitrogen).. The specific primer sets were as follows (Table II-1):. AtNUDX11-cGFP,. attB1-AtNUDX11-cGFP. and. attB2-AtNUDX11-cGFP;. AtNUDX15-cGFP,. attB1-AtNUDX15-15a. and. attB2-AtNUDX15-cGFP;. AtNUDX15a-cGFP,. attB1-AtNUDX15-15a. and. attB2-AtNUDX15a-cGFP;. AtNUDX15-nGFP,. attB1-AtNUDX15-15a-nGFP. and. attB2-AtNUDX15;. AtNUDX15a-nGFP, attB1-AtNUDX15-15a-nGFP and attB2-AtNUDX15a.. and. To fuse. the GFP cDNA in-frame with the cDNA encoding AtNUDX15 or 15a in the internal regions, pDONR/AtNUDX15 or 15a was digested with DraI, by which AtNUDX15 (882 bp) and 15a (858 bp) cDNAs are separated into 2 fragments (291 and 591 bp, and 291 and 567 bp, respectively), and dephosphorylated by Alkaline phosphatase, SAP (Takara, Kyoto, Japan) (Figure II-3). primers. (GFP-F. and. GFP-R). pDONR/AtNUDX15 or 15a. sequencing.. Amplified GFP cDNA using phosphorylated. (Table. II-1). were. ligated. into. linearized. The nucleotide sequence was verified by DNA. The fragment obtained was re-cloned into the destination vector, pGWB2.. Agrobacterium tumefaciens (strain C58), which was transformed with constructs obtained by electroporation, was used in the transformation of Nicotiana tabacum BY-2 (Nicotiana tabacum L. cv bright yellow 2) cells by a modification of the procedure reported by Gu and Verma (1997).. The fluorescence of the GFP fusion proteins in. protoplasts prepared from transgenic BY-2 cells was monitored using a Radiance 2100 confocal fluorescence microscope (Bio-Rad, Hercules, CA) (Ogawa et al., 2008). The mitochondria were stained with a mitochondria-selective dye, MitoTracker Orange (Invitrogen). Photographs were processed using LaserSharp2000 software (Carl Zeiss, 12.
(19) Oberkochen, Germany). Dra I attL1 AttL1. P. AtNUDX15(a) AtNUDX15. GFP. attL2 AttL2. P. pDONR201/AtNUDX15(a). pGWB5/GFP Amplification using phosphorylated primers. Dra I digestion P. GFP. P. Dephosphorylation. Ligation attL1 AttL1. AtN. UDX15(a) 15. GFP. attL2 AttL2. pDONR201/AtNUDX15(a)i. attR1 ccdB AttL2 attR2 CaMV35S-P AttL2. LR recombination attB1 CaMV35S-P AttL1. AtN. UDX15(a) 15. GFP. pGWB2. attB2 AttL2. pGWB2/AtNUDX15(a)i. Figure II-3. Plasmid construction for expressing AtNUDX15 or 15a fused to GFP at the internal regions. To fuse in-frame the GFP cDNA with the cDNA encoding AtNUDX15 or 15a at the internal regions, pDONR/AtNUDX15 or 15a was digested by DraI, by which AtNUDX15 (882 bp) and 15a (858 bp) cDNAs are separated into 2 fragments (291 and 591 bp, and 291 and 567 bp, respectively). The linearized plasmid was then dephosphorylated by Alkaline phosphatase, SAP.. The GFP. cDNAwas amplified by PCR using phosphorylated primers (GFP-F and GFP-R) (Table II-1). The amplified fragment was ligated into the linearized pDONR/AtNUDX15 or 15a. The nucleotide sequence was verified by DNA sequencing. The AtNUDX15 and 15a cDNAs containing GFP obtained were re-cloned into the destination vector, pGWB2, by GATEWAY LR recombination reactions.. 13.
(20) Protein analysis A polyclonal mouse antibody raised against the AtNUDX15 protein was prepared using His-tagged recombinant AtNUDX15, as described previously (Ogawa et al., 2005).. Immunoblot analysis was performed as reported previously (Yoshimura et. al., 2004).. AtNUDX11, 15, 15a, and 22 proteins were detected using the AtNUDX15. antibody as primary antibody and anti-mouse IgG-horseradish peroxidase conjugate (Bio-Rad) as secondary antibody.. Protein bands were detected using the enhanced. chemiluminescence detection system (GE Healthcare).. The protein concentration was. determined by the method of Bradford, using BSA as standard (Bradford, 1976).. Generation of transgenic plants Total RNA was isolated from the leaves of 2-week-old Arabidopsis plants (0.5 g fresh weight), as described previously (Yoshimura et al., 1999).. First-strand cDNA. was synthesized using ReverTra Ace reverse transcriptase (Toyobo) with an oligo (dT) primer.. The vector for generating the AtNUDX15- and 15a- overexpressing plants was. constructed using Gateway cloning technology (Invitrogen).. The cDNAs encoding the. open reading frame of AtNUDX15 and 15a were cloned into the donor vector, pDONR201, and then recloned into the destination vector, pGWB2.. The specific. primer sets were as follows (Table II-1): AtNUDX15, attB1-AtNUDX15-15a and attB2-AtNUDX15 and AtNUDX15a, attB1-AtNUDX15-15a and attB2-AtNUDX15a. PCR and in vitro BP and LR recombination reactions were performed following the manufacturer’s instructions (Invitrogen).. Agrobacterium tumefaciens, which was. transformed with the constructs obtained by electroporation, was used to infect Arabidopsis by the vacuum infiltration method.. T1 seedlings were selected on basic. MS medium in petri dishes containing 3% sucrose, 20 mg/L of hygromycin, and 20 mg/L of kanamycin over 2 weeks, and then transferred to soil. were harvested and used in the experiments.. 14. Homozygous T3 seeds.
(21) Isolation of organelles from Arabidopsis leaves Six-week-old Arabidopsis plants were transferred to dark conditions for induction of senescence, and then grown for 4 d by the method described by Reumann et al. (2007). Organelles were isolated from Arabidopsis (20 g FW) using Percoll (15-38%) and sucrose (0-36%) density gradients (10 mL). fractions were isolated. dehydrogenase. After centrifugation, 1-mL. The activities of catalase (Lück et al., 1963), succinate. (SDH). (Denton. et. al.,. 1996),. and. NADPH-dependent. glyceraldehyde-3-phosphate dehydrogenase (NADPH-GAPDH) (Ma et al., 2006), were measured as markers of peroxisomes, mitochondria, and chloroplasts respectively.. Results and Discussion. Alternative splicing of AtNUDX15 It has been reported that among Arabidopsis Nudix hydrolases, AtNUDX11 and 15 have CoA pyrophosphohydrolase activity (Ogawa et al., 2005; 2008). AtNUDX11 was predicted to be localized to the cytosol, since it had no transit peptide as judged by TargetP (www.cbs.dtu.dk/services/TargetP/). On the other hand, AtNUDX15 had the mTP at the N-terminus.. The mitochondrial localization was verified using a. GFP-fused protein (Ogawa et al., 2008). A search of the expressed sequence tag (EST) database identified a new alternatively spliced variant (designated AtNUDX15a) produced from the AtNUDX15 gene (Figure II-4A).. Due to intron retention-type. alternative splicing, the AtNUDX15a mRNA, which is shorter than the AtNUDX15 mRNA, was produced.. Translation of the AtNUDX15 mRNA was predicted to stop at. the UAA codon, resulting in the production of an AtNUDX15 protein composed of 294 amino acids (Figure II-4B). On the other hand, the AtNUDX15a mRNA translated a protein composed of 286 amino acids due to usage of the alternative stop (UGA) codon. Semiquantitative RT-PCR indicated that AtNUDX15 and 15a mRNAs were both 15.
(22) expressed in the leaves of 2-week-old Arabidopsis plants (Figure II-5).. The. nucleotide sequences of the various mRNAs were verified by DNA sequencing. The deduced AtNUDX15 and 15a proteins have the Nudix motif and the UPF0035 motif conserved in CoA pyrophosphohydrolases (Gasmi et al., 2001; AbdelRaheim et al., 2002; Ogawa et al., 2008).. Importantly, the predicted amino acid sequence of. AtNUDX15a has the type-1 peroxisomal targeting signal (PTS1) at the C-terminus, in addition to the mTP at the N-terminus, like AtNUDX15.. In other words, AtNUDX15a. has mTP and PTS1 at the N- and the C- terminus respectively, suggesting a dual subcellular distribution in mitochondria and peroxisomes.. 16.
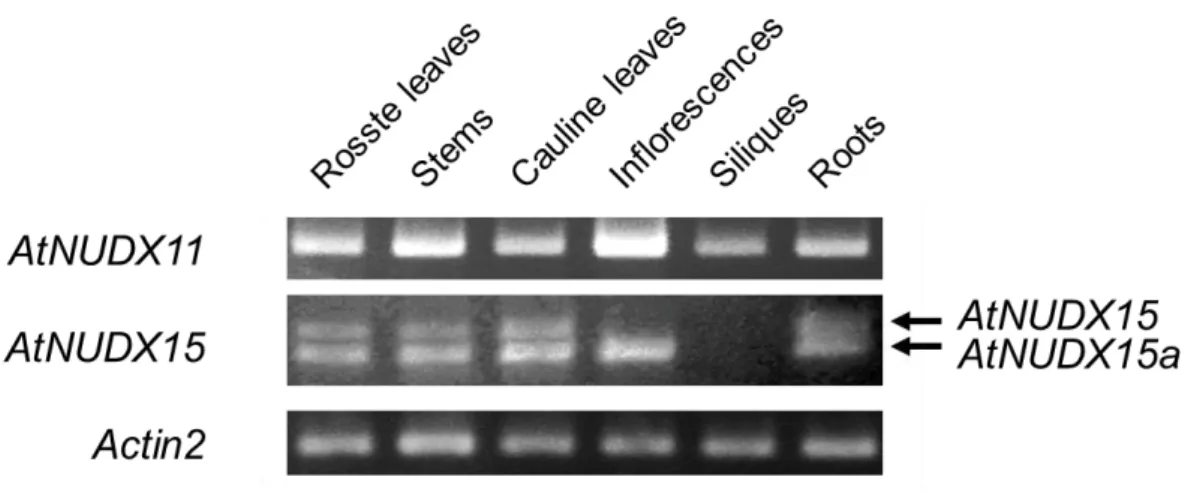
(23) Figure II-4. Alternative splicing patterns of AtNUDX15 mRNAs and putative structures of AtNUDX15 proteins. A, Schematic diagrams of AtNUDX15 pre-mRNA and alternatively spliced mRNA variants of it (AtNUDX15 and 15a). The constitutive exons are shown as black boxes, the alternative exons as gray boxes, and the introns as lines. The 5’- and 3’-untranslated regions in each mRNA are shown as white boxes. Start (ATG) and stop (UAA and UGA) codons are indicated. and P-2) indicate the locations of the primers in Figure II-5.. Arrows (P-1. B, Alignment of the amino acid. sequences of AtNUDX15, 15a, and 22, and mouse Nudt7α and C. elegans Y87G2A.14. Amino acids that are fully and highly conserved among CoA pyrophosphohydrolases are shown by black boxes. The bold-underlined, underlined, dotted-underlined, and double-underlined sequences are the Nudix motif, the UPF0035 motif, the mTP, and the PTS1 respectively.. Figure II-5. Changes in the expression levels of AtNUDX11, 15, and 15a in various plant organs. Semi-quantitative RT-PCR analysis of the AtNUDX11, 15, and 15a mRNAs in the roots, rosette leaves, stems, cauline leaves, inflorescences, silique, and roots of Arabidopsis plants grown in soil for 6 weeks under long-day conditions. PCR amplification was performed with 27-28 cycles of 95°C for 60 s, 55°C for 60 s, and 72°C for 60 s, followed by 72°C for 10 min. Aliquots of the products were analyzed on 1% agarose gel. Actin2 expression (18 cycles) was used as control in all experiments. Arrows indicate the bands of the AtNUDX15 and 15a mRNAs. The procedures are described in “Materials and Methods.”. 17.
(24) The expression of AtNUDX11, 15, and 15a in various plant organs Semiquantitative RT-PCR was used to examine the expression of the AtNUDX11, 15, and 15a mRNAs in rosette leaves, stems, cauline leaves, inflorescences, siliques, and roots.. AtNUDX11 mRNA was detected in all the organs tested, but the levels were. highest in the inflorescences (Figure II-5).. Consistent with this result, Kupke et al.. (2009) reported that AtNUDX11 expression is ubiquitous, but strong in flowers, vascular elements, and developing embryos.. More detailed information on the expression of. AtNUDX11 in the floral organs was obtained from the microarray data bank Genevestigator, with higher levels of AtNUDX11 observed in petals, stigmas, and abscission zones (Zimmermann et al., 2004) (Figure II-6).. Like AtNUDX11,. AtPANK1 and 2, enzymes for CoA biosynthesis, were highly expressed in these organs. Hence, it is likely that the function of AtNUDX11 is associated with CoA biosynthesis in floral development.. Figure II-6.. Expression profiles of AtNUDX11 and AtPANK in floral organs.. The expression data was obtained from the microarray data bank Genevestigator (Zimmermann et al., 2004).. 18.
(25) AtNUDX15 mRNA was expressed in the stems, roots, and rosette and cauline leaves, but not in the inflorescences or siliques.. The levels of AtNUDX15a mRNA. were similar to those of AtNUDX15 mRNA in the stems, roots, and rosette and cauline leaves, but AtNUDX15a mRNA was detected in the inflorescences (Figure II-5). These results indicate that the alternative splicing of AtNUDX15 is regulated in an organ-specific manner.. Although microarray analysis cannot independently detect. alternatively spliced variants, the data from Genevestigator indicates that the AtNUDX15 or 15a mRNA is highly expressed in the pollen, suggesting that the proteins are involved in the male reproductive organs.. Enzymatic characterization of AtNUDX11, 15, and 15a To characterize the recombinant forms of AtNUDX15a, the His-tagged AtNUDX15a gene product was expressed in E. coli transformed with the pColdII/AtNUDX15a plasmid.. Affinity chromatography on a HiTrap chelating. column yielded a highly purified recombinant protein, as found on the SDS/PAGE analysis (Figure II-7).. The hydrolytic activities of this enzyme toward various. nucleoside diphosphate derivatives were determined by HPLC.. AtNUDX15a. hydrolyzed CoA and its derivatives, including malonyl-CoA, succinyl-CoA, oxidized CoA, lauroyl-CoA (C12:0-CoA), and myristoyl-CoA (C14:0-CoA) (Table II-2).. A. product, 3’, 5’-ADP, of the reaction with CoA was detected (data not shown), indicating that CoA is hydrolyzed by the enzyme to form 3’,5’-ADP and 4’-phosphopantetheine. The Km value for the CoA (124.0 ± 5.4 μм) of AtNUDX15a was similar to or lower than those of other CoA pyrophosphohydrolases, such as AtNUDX15 and yeast Pcd1, mouse NUDT7, and C. elegans Y87G2A.15, and higher than that of AtNUDX11 (Ogawa et al., 2005; 2008; Cartwright et al., 2000; Gasmi and McLennan, 2001; AbdelRaheim et al., 2002) (Table II-2).. Previous studies have found that AtNUDX11 and AtNUDX15 are. highly active toward various CoA derivatives (Ogawa et al., 2008).. To compare the. enzymatic ability of AtNUDX11, 15, and 15a, kinetic parameters for various CoA 19.
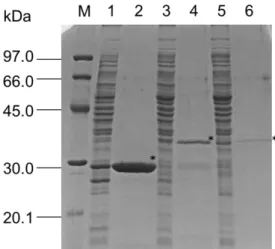
(26) derivatives were measured.. The production and purification of the recombinant. AtNUDX11 and 15 were carried out following previous reports (Ogawa et al., 2005; 2008). AtNUDX11 showed higher affinity for malonyl-CoA (52.3 ± 8.9 µм) than for CoA and the other CoA derivatives (Table II-2).. The Km value for the malonyl-CoA. of AtNUDX11 was lower than that of AtNUDX15 or AtNUDX15a. AtNUDX15 and 15a showed high affinity for oxidized-CoA (48.0 ± 3.4 and 53.0 ± 6.9 µм respectively) and succinyl-CoA (29.3 ± 9.5 and 35.2 ± 10.5 µм) compared with that for CoA. The Vmax values for the oxidized-CoA of AtNUDX15 and 15a (3.2 ± 0.52 and 3.4 ± 0.32 μmol/min/mg, respectively) were the highest among the CoA derivatives, and were similar to the S. cerevisiae PCD1p (Cartwright et al., 2000). The kcat/Km values for the lauroyl-CoA and myristoyl-CoA of these enzymes were higher than that of CoA or any other CoA derivatives.. All of the enzymes were highly active toward medium- to. long-chain acyl-CoAs, like mouse RP2p and NUDT7α (Ofman et al., 2006; Reilly et al., 2008).. Figure II-7.. Purification of recombinant AtNUDX11, 15, and 15a.. Recombinant AtNUDX11, 15, and 15a protein was overexpressed in E. coli, purified by Ni2+ affinity chromatography, and verified by SDS-PAGE (12.5%) with Coomassie Blue staining. The experimental conditions are described in “Materials and Methods.” 20 μg of crude extract prepared from E.coli expressing the recombinant AtNUDX11 (lane 1), AtNUDX15 (lane 3), and AtNUDX15a (lane 5), and 2 μg of purified recombinant AtNUDX11 (lane 2), AtNUDX15 (lane 4), and AtNUDX15a (lane 6) proteins were loaded. Bioscience). 20. M, molecular mass standards (Amersham.
(27) Table II-2. Comparison of Kinetic Parameters of the CoA Pyrophosphohydrolases in Arabidopsis, C.elegans, Mice, and S.cerevisiae. The standard assay was used with concentrations of 5 to 3,000 μм for CoA, succinyl-CoA, malonyl-CoA, lauroyl-CoA, myristoyl-CoA, and oxidized-CoA at 37°C with 5 mм Mg2+, as described in “Materials and Methods.” Data are the means of three independent determinations ± SD. -, Not determined. Protein. Km. Vmax. kcat. kcat/Km. Reference. μм. μmol/min/mg. 1/s. 1/s/м. 27.8 ± 2.3 118.7 ± 3.4 124.0 ± 5.4 220 240 280 600. 0.4 ± 0.01 1.4 ± 0.03 1.5 ± 0.04 5.3 0.56. 0.18 0.76 0.79 13.8 3.8 4.6 -. 6.5 x 103 6.4 x 103 6.4 x 103 6.3 x 104 1.5 x 104 1.6 x 104 -. Ogawa et al. (2008) Ogawa et al. (2008) This study AbdelRaheim and McLennan (2001) Gasmi and McLennan (2001) Cartwright et al. (2000) Ofman et al.(2006). Succinyl-CoA AtNUDX11 AtNUDX15 AtNUDX15a. 116.6 ± 12.8 29.3 ± 9.5 35.2 ± 10.5. 10.3 ± 1.81 1.6 ± 0.12 1.7 ± 0.18. 4.79 0.84 0.89. 4.1 x 104 2.9 x 104 2.5 x 104. This study This study This study. Malonyl-CoA AtNUDX11 AtNUDX15 AtNUDX15a Mouse Nudt7α. 52.3 ± 8.9 98.5 ± 12.5 102.5 ± 20.3 330. 6.3 ± 1.22 2.5 ± 0.37 2.6 ± 0.65 4.1. 2.90 1.35 1.38 3.0. 5.6 x 104 1.4 x 104 1.3 x 104 9.0 x 104. This study This study This study Gasmi and McLennan (2001). Lauroyl-CoA (C12-CoA) AtNUDX11 AtNUDX15 AtNUDX15a Mouse Nudt7α Mouse RP2p. 29.4 ± 4.6 22.3 ± 6.5 17.8 ± 4.6 22 80. 11.4 ± 1.31 2.8 ± 0.51 2.4 ± 0.68 1.06 0.2. 5.28 1.52 1.28 0.50 -. 1.8 x 105 6.8 x 104 7.2 x 104 2.2 x 104 -. This study This study This study Gasmi and McLennan (2001) Ofman et al.(2006). Myristoyl-CoA (C14-CoA) AtNUDX11 AtNUDX15 AtNUDX15a Mouse Nudt7α. 23.7 ± 5.8 17.5 ± 2.8 22.4 ± 4.9 34. 9.3 ± 1.78 1.7 ± 0.33 2.0 ± 0.48 0.41. 4.30 0.93 1.06 0.19. 1.8 x 105 5.3 x 104 4.7 x 104 8.1 x 103. This study This study This study Gasmi and McLennan (2001). 180.9 ± 21.4 48.0 ± 3.4 53.0 ± 6.9 580. 5.5 ± 0.63 3.2 ± 0.52 3.4 ± 0.32 0.012. 2.56 1.74 1.80 -. 1.4 x 104 3.6 x 104 3.3 x 104 -. This study This study This study Ofman et al.(2006). CoA AtNUDX11 AtNUDX15 AtNUDX15a C. elegans Y87G2A Mouse Nudt7α S. cerevisiae PCD1 Mouse RP2p. Oxidized-CoA (CoASSCoA) AtNUDX11 AtNUDX15 AtNUDX15a Mouse RP2p. 21.
(28) These results indicate that AtNUDX11, 15, and 15a prefer CoA derivatives to free CoA, suggesting that the actions of such proteins are associated with CoA derivative-related metabolic factors, such as the TCA cycle and fatty acid metabolism.. Subcellular localization of AtNUDX15 and 15a It has been reported that AtNUDX15 has N-terminal mTP and is localized to the mitochondrion; but the subcellular distribution of AtNUDX11 and AtNUDX15a remains unclear (Ogawa et al., 2008).. As described above, AtNUDX15a has not only. mTP at the N-terminus but also PTS1 at the C-terminal end.. To confirm the. subcellular distribution of these enzymes, full-length AtNUDX11, 15, and 15a cDNAs were fused in-frame with GFP at the N- or the C-terminus and then introduced into tobacco (Nicotiana tabacum) BY-2 cells.. The fluorescence of the GFP-fused proteins. in the transgenic cells was monitored by confocal microscopy.. Fluorescence of. AtNUDX11 fused to GFP at its C-terminus (AtNUDX11c-GFP) was detected in the cytosol (Figure II-8).. The fluorescence of AtNUDX15a fused to GFP at the. C-terminus (AtNUDX15ac-GFP) was co-localized with mitochondria stained with MitoTraker Orange, similarly to that of AtNUDX15 fused to GFP at C-terminus (AtNUDX15c-GFP), as reported previously (Ogawa et al., 2008). The. fluorescence. of. AtNUDX15a. fused. to. GFP. at. N-terminus. (AtNUDX15an-GFP) showed dot-like patterns in the cytoplasmic area distinct from mitochondria, probably due to their peroxisomal localization, whereas that of AtNUDX15 fused to GFP at N-terminus (AtNUDX15n-GFP) was detected only in the cytosol (Reumann et al., 2007; Carrie et al., 2006; Simkin et al., 2011) (Figure II-8). These results suggest that the actions of the N-terminal mTP and C-terminal PST1 of AtNUDX15a are disturbed by the fusion of GFP, and that both signals are potentially functional for organelle targeting.. Hence, to determine which targeting signal of. AtNUDX15a. subcellular. predominates,. the. distribution. of. AtNUDX15. and. AtNUDX15a fused to GFP via its internal regions (AtNUDX15i-GFP and 22.
(29) AtNUDX15ai-GFP respectively) was analyzed.. Fluorescence of AtNUDX15ai-GFP. was detected in mitochondria as well as that of AtNUDX15i-GFP (Figure II-8).. This. indicates that the N-terminal mTP of AtNUDX15a predominates and hence that the protein is localized to the mitochondria.. This result is similar to the case of rat. serine:pyruvate aminotransferase (SPT), in that the C-terminal PTS1 of SPT has been reported to be inhibited by the N-terminal mTP (Mizuno et al., 2002).. 23.
(30) Figure II-8.. Subcellular localization of AtNUDX11, 15, and 15a.. Confocal images of Tobacco BY-2 cells expressing the AtNUDX11, 15, and 15a proteins fused with GFP. Plasmids expressing AtNUDXs fused with GFP were introduced into Tobacco BY-2. Green fluorescence signals (a-g) of GFP and red fluorescence signals of mitochondria stained with MitoTracker (h-n) were detected with a laser confocal microscope (Bio-Rad, Hercules, CA). Merged images are shown in o-u. Scale bars are 10 μm. The procedures are described in the “Materials and Methods.”. Next, I confirmed the mitochondrial localization of AtNUDX15 and 15a in Arabidopsis plants.. First, an anti-AtNUDX15 polyclonal antibody was generated. using recombinant AtNUDX15.. In immunoblot analyses, the polyclonal antibody. recognized the recombinant protein in a range from 5 ng to 1 μg, and cross-reacted with AtNUDX11 and 15a (Figure II-9A).. In addition, it cross-reacted with the. recombinant AtNUDX22 protein, which is highly homologous to AtNUDX15 (Figure II-9B), although the protein had no activity toward any substrates (Ogawa et al., 2008). Using the antibody obtained, the AtNUDX15 and 15a proteins (deduced molecular weights 32.8 and 31.9 kDa respectively) were not be detected in the crude extract prepared from 2-week-old Arabidopsis leaves grown under normal conditions (data not shown).. Hence I isolated organelles from leaves of 6-week-old plants (15 g. fresh weight) using the method described by Reumann et al. (2007). Fractions isolated on a Percoll density gradient were examined for catalase, SDH, and NADPH-GAPDH activities, markers for peroxisomes, mitochondria, and chloroplasts respectively.. The. highest levels of catalase, SDH, and NADPH-GAPDH activities were detected in fractions 2, 8, and 11 respectively (Figure II-10A).. As in the case of SDH, the highest. level of the CoA pyrophosphohydrolase activity was detected in fraction 8, although activities were also detected in the low-density fractions, including the chloroplast fractions, probably due to contamination of the cytosolic AtNUDX11 protein in those factions.. However, on immunoblot analysis, AtNUDX15 and 15a were not detected in. any fraction.. These results suggest that the levels of AtNUDX15 and 15a in the leaves 24.
(31) of Arabidopsis plants are extremely low or else undetectable based on post-translational protein modification.. Figure II-9.. Production of anti-AtNUDX15 polyclonal antibody.. A, Antibody titers to the AtNUDX15 protein using the anti-AtNUDX15 polyclonal antibody. The recombinant AtNUDX15 protein (5, 50, 100, 250, 500, and 1,000 ng, lanes 1 to 6), was recognized by the anti-AtNUDX15 polyclonal antibody in the immunoblot analysis.. B,. Cross-reactivity of the anti-AtNUDX15 antibody to the recombinant AtNUDX11, 15a, and 22 proteins (2 µg each). Asterisks indicate the bands of the AtNUDX11 (27.9 kDa), 15 (32.6 kDa), 15a (31.7 kDa), and 22 (33.0 kDa) proteins.. 25.
(32) 26.
(33) Figure II-10. plants. Extracts. Subcellular distribution of AtNUDX15 and 15a in Arabidopsis prepared. from. wild-type. (A),. Pro35S:AtNUDX15-1-2. (B),. and. Pro35S:AtNUDX15a-2-1 (C) plants were fractionated by a discontinuous Percoll and sucrose gradient (15-38%, 0-36%). The distribution of CoA pyrophosphohydrolase activity and marker activities on the Percoll density gradient is shown. Catalase, SDH, and NADPH-GAPDH were markers of peroxisomes, mitochondria, and chloroplasts respectively.. Fraction 1 represents the. bottom of the gradient. Aliquots of the fractions (15 μL of each) were subjected to immnoblot analysis using the anti-AtNUDX15 polyclonal antibody.. 27.
(34) Figure II-11. plants.. Characteristics of AtNUDX15 and 15a-overexpressing Arabidopsis. A, Semi-quantitative RT-PCR analysis of the AtNUDX15 and 15a mRNAs in wild-type (WT), Pro35S:AtNUDX15, and Pro35S:AtNUDX15a (T3 generations of independent transformed lines) plants. The plants were grown on MS medium for 2 weeks under long-day conditions, and then the leaves were used for analysis. Equal loading of amplified cDNAs was determined with the control Actin2 PCR product. B, Immunoblot analysis of the AtNUDX15 and AtNUDX15a proteins in the wild-type (WT), Pro35S:AtNUDX15, and Pro35S:AtNUDX15a plants. The AtNUDX15 and 15a. proteins. were. detected. by. anti-AtNUDX15. pyrophosphohydrolase. activities. in. the. leaves. of. polyclonal the. WT,. antibody.. C,. Pro35S:AtNUDX15,. CoA and. Pro35S:AtNUDX15a plants. Data are means ± SD for three individual experiments (n = 3) using plants grown independently.. The details of the procedures are described in “Materials and. Methods.” Asterisks indicate values significantly different from the corresponding value of the wild-type plants (p < 0.05).. Hence I generated transgenic Arabidopsis plants overexpressing AtNUDX15 and 15a under the control of the cauliflower mosaic virus 35S promoter (Pro35S:AtNUDX15 and Pro35S:AtNUDX15a respectively).. There was no difference. in growth or morphology between the control and transgenic plants under normal conditions.. The levels of the AtNUDX15 and 15a mRNAs were higher in the T3. generation of the Pro35S:AtNUDX15 and 15a plants as compared with the control plants (Figure II-11A).. Immunoblotting detected the AtNUDX15 protein in extracts. prepared from the leaves of the Pro35S:AtNUDX15-1-1, -1-2, -10-1, and -10-2 plants (Figure II-11B).. Similarly, the Pro35S:AtNUDX15a-2-1, -2-2, -3-1, and -3-2 plants. accumulated the AtNUDX15a protein.. The pyrophosphohydrolase activities toward. CoA in the Pro35S:AtNUDX15 and Pro35S:AtNUDX15a plants were approximately 1.4- to 2.4-fold and 1.1- to 1.3-fold respectively higher than the levels in the wild-type plants (Figure II-11C).. To confirm subcellular distribution of these enzymes, I. isolated organelles from the Pro35S:AtNUDX15 and Pro35S:AtNUDX15a plants. Judging from the activities of catalase and SDH, fractions 4 and 9 isolated from the Pro35S:AtNUDX15a plants contained peroxisomes and mitochondria respectively 28.
(35) (Figure II-10C).. The highest level of CoA pyrophosphohydrolase activity was. detected in the mitochondrial fraction.. On immunoblot analysis, the AtNUDX15a. protein was detected in the mitochondrial fraction.. Similarly, AtNUDX15 was. detected only in the mitochondrial fraction isolated from the Pro35S:AtNUDX15 plants (Figure II-10B).. These results clearly indicate that AtNUDX15a as well as. AtNUDX15 is localized exclusively to the mitochondria.. Conclusion. CoA and. its. derivatives depend on. the. metabolic state,. and are. compartmentalized in the cytosol, mitochondria, and peroxisomes, suggesting the importance of their own pool and of mechanism to tightly controling their levels in each organelle.. In this study I characterized the molecular and enzymatic properties of. Arabidopsis CoA pyrophosphohydrolases AtNUDX11, 15, and 15a.. The cytosolic. pool of malonyl-CoA contributes to various pathways, including fatty acid elongation and flavonoid biosynthesis, whereas the plastidic pool contributes to de novo fatty acid synthesis (Baud et al., 2003). Hence it appears likely that the actions of cytosolic AtNUDX11 impact many cellular processes in various organs, especially in the inflorescences.. In addition, AtNUDX11 might contribute to the regulation of CoA. biosynthesis by acting on the degradation of malonyl-CoA’s, since the activity of a key enzyme in CoA biosynthesis, PANK, is inhibited by malonyl-CoA, but not by CoA or acetyl-CoA (Falk et al., 1993) (Figure II-12).. Analysis of the expression and. subcellular distribution of AtNUDX15 and 15a suggested that the two enzymes act similarly in the mitochondria of various plant organs, while the latter has a specific function in inflorescences.. In addition, it is possible that AtNUDX15 and 15a impact. the regulation of the TCA cycle by the hydrolysis of succinyl-CoA, a key molecule in the cycle (Araújo et al., 2008) (Figure II-12). 29.
(36) The actions of CoA pyrophosphohydrolases toward free CoA or CoA derivatives produce of 3’,5’-ADP and (acyl-) 4’-phosphopantetheine. The fate of the products is unknown, but 4’-phosphopantetheine might be exported from the mitochondria to the cytosol and might be reused in the synthesis of CoA by PPAT. Although more studies are required, my results provide a starting-point in uncovering the importance of the degradation of CoA and its derivatives by CoA pyrophosphohydrolases in plant cellular processes.. Figure II-12.. Proposed model for the physiological roles of AtNUDX11, 15 and 15a on. the CoA metabolisms. 30.
(37) CHAPTER III. Identification and molecular characterization of Arabidopsis ppGpp pyrophosphohydrolases. Introduction. The biosynthesis of ppGpp is catalyzed by two homologous enzymes, RelA and SpoT (Cashel et al., 1969; Magnusson et al., 2005).. In E. coli cells grown under. condition of amino acid depletion, pppGpp is synthesized from ATP and GTP by ribosome-associated RelA and subsequently converted to ppGpp by 5’-nucleotidase (Figure III-1). Thereafter, ppGpp is degraded to GDP and pyrophosphate by SpoT, which has reciprocal activities for ppGpp degradation and synthesis.. Van der Biezen. et al. (2000) isolated three RelA/SpoT homologues, designated RSH genes (AtRSH1, AtRSH2, and AtRSH3) from Arabidopsis thaliana.. In addition, Arabidopsis had a. single CRSH gene, a Ca2+-activated RelA-SpoT homolog, for an enzyme having ppGpp synthase activity (Tozawa et al., 2007; Masuda et al., 2008a; 2008b).. It has been. demonstrated that ppGpp and the ppGpp synthetase occur in chloroplasts along with all RSH and CRSH proteins in Arabidopsis plants (Givens et al., 2004; Kasai et al., 2004; Mizusawa et al., 2008; Takahashi et al., 2004). The chloroplast is a semiautonomous organelle that originates from not only an ancestral host cell but also a cyanobacterium-like photosynthetic prokaryote.. Therefore, it seems feasible that a. stringent control system by ppGpp similar to that of bacteria exists in plant chloroplasts (Kasai et al., 2002; Givens et al., 2004; Braeken et al., 2006). However, it is unclear how ppGpp levels are regulated in plant chloroplasts. In addition, there is no information on the contribution of ppGpp pyrophosphohydrolase such as Ndx8 to ppGpp metabolism in plants.. In this chapter, I identified the ppGpp. pyrophosphohydrolases (AtNUDX11, 15, 25, and, 26) in Arabidopsis. 31. Among them,.
(38) judging from its subcellular localization, enzymatic properties (enzymatic ability and kinetic parameters) toward ppGpp, and gene expression in response to environmental stress, AtNUDX26 seems to be involved in metabolism of ppGpp in chloroplasts.. Figure III-1.. Metabolic pathway for ppGpp. Materials and Methods. Expression and purification of the recombinant AtNUDX proteins Recombinant forms of AtNUDXs (AtNUDX1-27) were produced using E. coli strain BL21 (DE3) pLysS cells and purified from the extract using a HiTrap chelating HP column (GE Healthcare, Little Chalfont, UK) following previous reports (Ogawa et al., 2005; 2008; Yoshimura et al., 2007). The protein content was determined by the Bradford method, using bovine serum albumin as a standard (Bradford, 1976).. The. molecular masses of the recombinant AtNUDX proteins agreed with the predicted 32.
(39) values, calculated from the amino acid sequence of the mature protein plus the hexahistidine-tag (data not shown).. Enzyme assay and HPLC The ppGpp and ganosine 3’,5’-bisphosphate (pGp) was purchased from TriLink Biotechnologies (San Diego, CA, USA).. The hydrolytic activities of. recombinant forms of AtNUDXs toward ppGpp were assayed by a method described previously (Ogawa et al., 2005; 2008). The reaction mixture (60 μL) containing 50 mм Tris-HCl (pH 8.0), 5 mм MgCl2, 25 μм ppGpp, and 0.2-1.0 μg of the purified recombinant protein, was incubated at 37°C for 10 min.. For the assay of Km, ppGpp at. 25 µм to 800 µм was added to the reaction.. For the assay of divalent. cation-dependency, Mg2+ was substituted with Ni2+, Zn2+, Ca2+, or Mn2+ (5 mм each). For the assay of pH-dependency, the Tris-HCl buffer was replaced with Glycine-NaOH buffer (100 mм each) at basic pH (pH8.5-10.0). assayed at 20-60°C.. Temperature-dependency was. The reaction was terminated by adding 10 μL of 100 mм EDTA.. In addition, the reaction was carried out under reducing conditions with 1 mм DTT according to Olejnik et al. (2007). The mixture was then analyzed by HPLC using a COSMOSIL C18 column (4.6 x 250 mm, Nacalai Tesque, Kyoto, Japan) at a flow rate of 0.6 mL/min for the mobile phase buffer, which contained 73 mм KH2PO4, 5 mм tetrabutylammonium dihydrogenphosphate, and 21% methanol.. The substrates. (ppGpp) and the reaction product (pGp) were detected by their UV absorbance at 252 nm.. Plant materials and stress treatments Arabidopsis thaliana wild-type (Col-0) was grown on Murashige and Skong’s medium under a light intensity of 100 μmol photons/m2/s.. Two-week-old Arabidopsis. plants were subjected to various types of stress with salinity, paraquat, drought, and heat. Paraquat at 50 μм was sprayed on the plants. 33. Salinity stress was imposed by.
(40) transferring the plants to MS medium containing 250 mм NaCl for 0 to 9 h.. Drought. stress was imposed by subjecting the plants to dehydration on paper towels for 0 to 6 h. Heat stress was imposed by incubating cultures for 0 to 3h at 37°C.. Quantitative Real-Time PCR experiments Quantitative Real-Time PCR (RT–PCR) experiments were performed according to Nishizawa et al. (2006). Primer sequences were as follows; NUDX26-primerF (5’-ATCCGATTGGAAGGGTCAAGCAC-3’), NUDX26-primerR (5’-CTCAGGTTTCTCAGTACCATCGC-3’), RSH1-primerF (5’-TTCTTGCGTGGCCGAGATTGACA-3’), RSH1-primerR (5’-CACCTTAGCGCAAACACACTAACCAAACT-3’), RSH2-primerF (5’-CTGAAGATGGGTGATGTGGTGG-3’), RSH2-primerR (5’-CGCGAGTCCTCGATCATACATG-3’), RSH3-primerF (5’-ACACCTGTAAGTGATCTGAAATGCAAG-3’), RSH3-primerR (5’-CGCAAGCCCTCGATCATACATC-3’), CRSH-primerF (5’-CAGACGAATTCGATACGTTTCAGAAAC-3’), CRSH-primerR (5’-TCATGAAGCTTCTCGTCTAAAAGACTC-3’), Actin2-QF (5’-GGTGGTTCCATTCTTGCTTCCC-3’), Actin2-QR (5’-TCATACTCGGCCTTGGAGATCC-3’).. Data analysis The significance of differences between data sets was evaluated with a t-test. Calculations were carried out with Microsoft Excel software.. Accession numbers Arabidopsis Genome Initiative locus identifiers for major genes mentioned in this article are as follows: AtNUDX1 (At1g68760), AtNUDX2 (At5g47650), AtNUDX3 (At1g79690), AtNUDX4 (At1g18300), AtNUDX5 (At2g04430), AtNUDX6 (At2g04450), 34.
(41) AtNUDX7 (At4g12720), AtNUDX8 (At5g47240), AtNUDX9 (At3g46200), AtNUDX10 (At4g25434),. AtNUDX11. (At5g45940),. AtNUDX12. (At1g12880),. AtNUDX13. (At3g26690),. AtNUDX14. (At4g11980),. AtNUDX15. (At1g28960),. AtNUDX16. (At3g12600),. AtNUDX17. (At2g01670),. AtNUDX18. (At1g14860),. AtNUDX19. (At5g20070),. AtNUDX20. (At5g19460),. AtNUDX21. (At1g73540),. AtNUDX22. (At2g33980),. AtNUDX23. (At2g42070),. AtNUDX24. (At5g19470),. AtNUDX25. (At1g30110), AtNUDX26 (At3g10620), AtNUDX27 (At5g06340), RSH1 (At4g02260), RSH2 (At3g14050), RSH3 (At1g54130), CRSH (At3g17470), Actin2 (At3g18780).. Results and Discussion. Identification of AtNUDXs having pyrophosphohydrolase activity toward ppGpp To identify AtNUDXs having pyrophosphohydrolase activity toward ppGpp, I prepared His-tagged recombinant AtNUDX1-27 proteins in the absence of the predicted transit peptide expressed in E. coli cells.. The production and purification of the. recombinant AtNUDXs were carried out according to a method reported previously (Ogawa et al., 2005; 2008; Yoshimura et al., 2007) (Figure III-2). The hydrolase activity of AtNUDXs toward ppGpp in the presence of 5 mм Mg2+ as a cofactor was examined by an HPLC analysis. The recombinant AtNUDX26 hydrolyzed ppGpp with relatively high activity (0.19 ± 0.05 μmol/min/mg protein). AtNUDX11 (0.06 ± 0.01 μmol/min/mg protein), 15 (0.02 ± 0.01 μmol/min/mg protein), and 25 (0.06 ± 0.01 μmol/min/mg protein) exhibited barely detectable activity.. No ppGpp hydrolase. activity was detected in the other AtNUDXs (AtNUDX1-10, 12-14, 16-24, and 27).. 35.
(42) Figure III-2.. Purification of recombinant AtNUDX proteins.. Recombinant AtNUDX proteins were overexpressed in E. Coli, purified with Ni2+ affinity chromatography, and verified using SDS-PAGE with Coomassie blue staining. The experimental conditions are described in “Materials and Methods”. Left lanes and right lanes of all AtNUDXs contain 15 μg extract and 2 μg of purified recombinant proteins, respectively. M: molecular mass standards (Amersham Bioscience) as indicated on the left.. It has been reported that the activities of some Nudix hydrolases, such as AtNUDX13 and 26 having ApnA pyrophosphohydrolase Activity, increased under reducing conditions (Olejnik et al., 2007; Ogawa et al., 2008).. However, in the. presence of the reducing agent, dithiothreitol (DTT: 1 mм), in the reaction solution, the activities of AtNUDX11 (0.05 ± 0.02 μmol/min/mg protein), AtNUDX25 (0.05 ± 0.03 μmol/min/mg protein), and AtNUDX26 (0.12 ± 0.04 μmol/min/mg protein) were 36.
(43) similar to those in the absence of the agent.. No ppGpp hydrolase activity was detected. for the other AtNUDXs under reducing conditions (data not shown). The reaction products generated by the activity of AtNUDX26 toward ppGpp were determined by an HPLC analysis (Figure III-3).. In addition to the peak of. ppGpp as the substrate, three peaks having absorbance at 252 nm were detected in the reaction solution.. The peak that eluted first (approximately 9.50 min) corresponded to. that of standard pGp, while the others were estimated to be either pGpp or ppGp. These findings indicated that AtNUDX26 has pyrophosphohydrolase activity toward both the diphosphate linkages in ppGpp and generates pGp as a final product. At present, the fate of pGp as well as pGpp/ppGp is unknown even in bacterial cells (Ooga et al., 2009).. Figure III-3. AtNUDX26.. Identification of the products of ppGpp’s degradation by. Reaction mixtures containing 25 μм ppGpp were incubated with 5 mм Mg2+ in the absence or presence of the purified recombinant AtNUDX26 protein at 37˚C for 10 min and then subjected to HPLC with the COSMOSIL C18 column as described in “Materials and Methods.” The elution profile of the reaction mixture without the enzyme (gray line) or with the enzyme (black line) is shown. Positions of standards (15 µmol of GDP, 25 µmol of GTP, 25 µmol of ppGpp, and 25 µmol of pGp) are shown in the upper panel. Asterisks are estimated to be either pGpp or ppGp.. 37.
(44) As. described. above,. none. of. the. AtNUDXs. having. ppGpp. pyrophosphohydrolase activity showed significant homology to T. thermophilus Ndx8. Conversely, previous studies have shown that AtNUDX11 and 15 have the UPF0035 motif conserved in CoA pyrophosphohydrolases (Ito et al., 2012). AtNUDX25 and 26 were identified as ApnA pyrophosphohydrolases having an unique conserved amino acid sequence (GGGX5EX7REUXEEXGUX2GX6G) (Kraszewska et al., 2008).. These. results suggest that the mechanism for recognizing ppGpp is similar to that for recognizing CoA and ApnA and that the plant ppGpp pyrophosphohydrolases have evolved in their own way different from their bacterial counterparts.. Enzymatic properties and kinetic parameters of AtNUDX26 as a ppGpp pyrophosphohydrolase Most previously characterized Nudix hydrolases strictly require bivalent metal ions for their activity.. Therefore we studied the effect of a series of bivalent metal ions. on the activity of AtNUDX26 (Figure III-4A).. The activity of AtNUDX26 was. detected only in the presence of Mg2+ and Mn2+.. No activity was detected in the. absence of metal ions or in the presence of the other metal ions tested. The optimum pH for the activity was determined using Tris-HCl (pH 6.5-9.0) and glycine-NaOH (pH 8.5-10.0) buffers. As shown in Figure III-4B, AtNUDX26 was most active at pH 9.0, with 50% activity at pH 7.0 and 9.5.. The optimum. temperature for the enzyme was 50°C (Figure III-4). Kinetic parameters for ppGpp were measured (Table III-1).. The Km values of. AtNUDX11, 15, 25, and 26 were 29.1 ± 2.5, 145.3 ± 9.8, 63.7 ± 3.6, and 135.1 ± 9.6 µм, respectively. The Vmax of AtNUDX26 was 1.64 ± 0.30 μmol/min/mg, the highest value obtained. Therefore, AtNUDX26 had the highest catalytic efficiency (kcat/Km) for ppGpp among these AtNUDXs.. It has been reported that AtNUDX26 has the. predicted chloroplastic transit peptide and the protein fused with GFP fluorescent protein is distributed in chloroplasts (Ogawa et al., 2008). These findings and the 38.
(45) results obtained here suggest that AtNUDX26 contributes to the regulation of ppGpp levels via hydrolysis in the chloroplasts.. Figure III-4. Enzymatic characterization of AtNUDX26. (A) The requirement of divalent cations for ppGpp pyrophosphohydrolase activity was determined in the presence or absence of divalent cations (5 mм) at 37˚C for 10 min as described in “Materials and Methods.” (B) The pH-dependency of ppGpp pyrophosphohydrolase activity was determined under the assay conditions described in “Materials and Methods”, except that Tris-HCl buffer was replaced with Glycine-NaOH buffer (100 mм each) at the indicated pH.. (C) The. temperature-dependency of ppGpp pyrophosphohydrolase activity was determined as described in “Materials and Methods”, at 20–60°C. 39.
(46) Table III-1.. Analysis of the enzymatic properties of AtNUDX11, 15, 25, and 26.. The standard assay was used with concentrations of 5 to 1,500 µM for ppGpp at 37°C with 2+. 5 mм Mg as described in “Materials and Methods.” Data are means of three independent determinations ± SD.. Protein. AtNUDX11 AtNUDX15 AtNUDX25 AtNUDX26. Km μм. Vmax μmol/min/mg. kcat 1/s. kcat/Km 1/s/м. 29.1 ± 2.5 145.3 ± 9.8 63.7 ± 3.6 135.1 ± 9.6. 0.15 ± 0.03 0.05 ± 0.01 0.35 ± 0.07 1.64 ± 0.30. 0.07 0.03 0.12 0.49. 2.3 x 103 2.0 x 102 1.9 x 103 3.6 x 103. Comparison of amino acid sequences between AtNUDXs and T. thermophilus Ndx8 Nudix. hydrolases. contain. the. highly. conserved. Nudix. box. GX5EX7REXEEUXGU, where U is a bulky hydrophobic amino acid such as Ile, Leu, or Val (Bessman et al., 1996; Xu et al., 2006; McLennan, 2006).. In addition, some. Nudix hydrolases, having hydrolytic activity toward ADP-ribose (Proline 15 or 16 aa downstream of the Nudix box), ApnA (GGGX5EX7REUXEEXGUX2GX6G), coenzyme A (UPF0035;. LLTXR[SA]X3RX3GX3FPGG),. NADH. (SQPWPFPQS),. or. UTP. (L[VL]VRK and AANE), contain additional regions conserved across several species (PROSITE; http://prosite.expasy.org/) (Frick et al., 1995; Dunn et al., 1999; Gasmi et al., 2001; Xu et al., 2003; McLennan, 2006; Kraszewska, 2008).. Therefore, I. compared the amino acid sequences of AtNUDXs with T. thermophilus Ndx8 known as a ppGpp pyrophosphohydrolase (Ooga et al., 2009).. However, Ndx8 did not share. significant sequence homology to any of the AtNUDXs (3-24%), including AtNUDX11 (19%), 15 (17%), 25 (24%), and 26 (18%), and no region other than the Nudix motif was conserved between them (data not shown). These findings suggest that plant Nudix hydrolases having ppGpp pyrophosphohydrolase activity undergo their own evolutionary process.. Alternatively, there might be unidentified amino acid residue(s). essential for the ppGpp pyrophosphohydrolase activity, since a couple of residues are reported to be essential for activity or substrate recognition in some NUDXs such as 40.
(47) AtNUDX1 and E. coli NudG (Kraszewska, 2008).. Expression of AtNUDX26 in response to various types of stress It has been reported that ppGpp level in plant cells increased in response to various forms of environmental stress (Takahashi et al., 2004).. I analyzed the changes. in the expression of AtNUDX26 as well as the genes encoding enzymes involved in ppGpp metabolism, AtRSH1, AtRSH2, AtRSH3, and CRSH, under various stressful conditions.. RT–PCR analysis revealed that transcript levels of AtNUDX26 increased. approximately 4-fold at 6 h after drought stress (Figure III-5). A significant increase in expression under drought stress was observed for all genes tested here.. In particular,. the levels of AtRSH2 and AtRSH3 increased approximately 19- and 24-fold, respectively. Conversely, the expression of AtNUDX26 decreased under oxidative stress caused by treatment with paraquat (PQ: a generator of reactive oxygen species) and under salinity caused by treatment with NaCl (Figure III-5). Similarly, levels of CRSH decreased in response to both PQ and NaCl treatments. response to these treatments.. In contrast, levels of AtRSH2 increased in. The expression of AtNUDX26 was not significantly. changed by treatment with heat, although expression levels of some AtRSHs increased or decreased (Figure III-5). These results suggest that the metabolism of ppGpp is regulated depending on the type of stress and AtNUDX26 might, partially contribute to the regulation.. 41.
(48) Figure III-5. Expression profiles of AtNUDX26, AtRSH1, AtRSH2, AtRSH3, and CRSH under various stressful conditions. Two-week-old Arabidopsis plants were subjected to various forms of stress, treatment with salinity (250 mм NaCl), PQ (50 µм), drought (dehydration on paper towel) and heat (37°C under dark conditions). Total RNA extracted from Arabidopsis leaves was converted into first strand cDNA using the oligo (dt)20 primer.. A quantitative PCR analysis was carried out to. determine the expression levels of AtNUDX26 (A) and AtRSH1, AtRSH2, AtRSH3, and CRSH (B). Detailed procedures are described in “Materials and Methods.”. The relative amounts were. normalized to Actin2 mRNA. Data are the mean values ± SD for three individual experiments (n = 3). Asterisks indicate values significantly different from the control treatment (p <0.05). 42.
(49) Conclusion. In plant cells, ppGpp is an important signaling molecule, which affects various cellular processes including transcription, translation, DNA replication, amino acid and nucleotide metabolism, secondary metabolism, and infectivity (Cashel et al., 1996; Dalebroux et al., 2010; Magnusson et al., 2005; Potrykus and Cashel, 2008; Srivatsan and Wang, 2008; Nomura et al., 2012).. For the first time, I characterized the. molecular and enzymatic properties of ppGpp pyrophosphohydrolases in plants, AtNUDX11, 15, 25, and 26.. It has been demonstrated that, in plant chloroplasts,. ppGpp is synthesized and degraded by RSHs and directly affects the translation system (Givens et al., 2004; Kasai et al., 2004; Mizusawa et al., 2008; Takahashi et al., 2004; Nomura et al., 2012; Tozawa et al., 2011).. In addition to the importance of ppGpp. metabolism in plant chloroplasts, the findings that, among AtNUDXs having ppGpp pyrophosphohydrolase activity, chloroplastic AtNUDX26 has the highest catalytic efficiency and its expression is regulated in response to various types of environmental stress suggest that AtNUDX26 impacts on the metabolism of ppGpp in chloroplasts through the hydrolysis of ppGpp under stress (Figure III-6).. Since ppGpp is a. substrate for the hydrolysis reaction by AtNUDX26, this enzyme might play a role in the fine-tuning of ppGpp signaling in combination with AtRSHs in the chloroplasts. In addition, it is possible that not only ppGpp but also pGp, the final product by the reaction of AtNUDX26, have signaling roles in cellular responses, since its analog, 3’-phosphoadenosine 5’-phosphate, has an inhibitory effect on some enzymes such as 3’-phosphoadenosine 5'-phosphosulfate, nucleoside diphosphate kinase, and poly (ADP-ribose) polymerase, involved in various biological processes (Toledano et al., 2012; Todeschini et al., 2006; Dichtl et al., 1997; Schneider et al., 1998; Klaassen et al., 1997).. To uncover the importance of the degradation of ppGpp by AtNUDX26, we are. progressing toward analyzing the effect of the knockout and overexpression of. 43.
(50) AtNUDX26 on levels of ppGpp and the response to various types of stress.. Figure III-6. Proposed model for the physiological roles of AtNUDX26 on the ppGpp metabolisms under environmental stress condition. 44.
(51) CHAPTER IV. Conclusion. Nudix hydrolases act on the hydrolysis of various nucleoside diphosphate derivatives, suggesting their roles in diverse cellular metabolisms.. However, the. information on the physiological functions of Nudix hydrolases was still limited.. In. this thesis, I showed that Nudix hydrolases have a potential to regulate the energy metabolisms in the cytosol and mitochondria and the stress signaling in the chloroplasts through the hydrolysis of CoA-derivatives and ppGpp, respectively, in Arabidopsis plants. Arabidopsis Nudix hydrolases, AtNUDX11 and 15, had pyrophosphohydrolase activity toward CoA and its derivatives.. I identified two alternatively spliced variants,. AtNUDX15 and 15a, produced from the AtNUDX15 gene, and carried out comparative studies of the gene regulation, the kinetic parameters, and the intracellular localization of AtNUDX11, 15, and 15a.. The present findings indicated that AtNUDX11 and. AtNUDX15(a) function in the hydrolysis of malonyl-CoA in the cytosol and succinyl-CoA in the mitochondria, respectively, suggesting their impact not only on CoA biosynthesis but also on various CoA-related pathways such as the TCA cycle. Next, I identified Arabidopsis Nudix hydrolases, AtNUDX11, 15, 25, and 26, having. the. ppGpp. pyrophosphohydrolase. activity. from. Arabidopsis. plants.. AtNUDX26 was localized in chloroplasts and showed highest Vmax and kcat values, and catalytic efficiency, kcat/Km. Expression of not only genes encoding the Arabidopsis RelA and SpoT involved in the ppGpp metabolism but also AtNUDX26 was regulated by various types of stress. These results suggest that AtNUDX26 regulates the cellular ppGpp levels in response to stresses and impacts on the gene expression in chloroplasts. The results obtained here provide the valuable information to elucidate diverse roles of Nudix hydrolases in cellular metabolisms of not only plants but also the other 45.
(52) organisms.. 46.
(53) REFERENCES. AbdelRaheim SR and McLennan AG (2002) The Caenorhabditis elegans Y87G2A.14 Nudix hydrolase is a peroxisomal coenzyme A diphosphatase. BMC Biochem., 27: 3-5.. Araújo WL, Nunes-Nesi A, Trenkamp S, Bunik VI, and Fernie AR (2008) Inhibition of 2-oxoglutarate dehydrogenase in potato tuber suggests the enzyme is limiting for respiration and confirms its importance in nitrogen assimilation,. Plant Physiol., 148: 1782-1796.. Baud S, Guyon V, Kronenberger J, Wuillème S, Miquel M, Caboche M, Lepiniec L, and Rochat C (2003) Multifunctional acetyl-CoA carboxylase 1 is essential for very long chain fatty acid elongation and embryo development in Arabidopsis. Plant J., 33: 75-86.. Bessman MJ, Frick DN, and O'handley SF (1996) The MutT proteins or "Nudix" hydrolases, a family of versatile, widely distributed, "housecleaning" enzymes. J. Biol. Chem., 271: 25059-25062.. Bradford M (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Biochem., 72: 248-254.. Braeken K, Moris M, Daniels R, Vanderleyden J, and Michiels J (2006) New horizons for (p)ppGpp in bacterial and plant physiology. Trends Microbiol., 14: 45-54.. Carrie C, Murcha MW, Millar AH, Smith SM, and Whelan J (2006) Nine 3-ketoacyl-CoA thiolases (KATs) and acetoacetyl-CoA thiolases (ACATs) encoded by. 47.
(54) five genes in Arabidopsis thaliana are targeted either to peroxisomes or cytosol but not to mitochondria. Plant Mol. Biol., 63: 97-108.. Cartwright JL, Gasmi L, Spiller DG, and McLennan AG (2000) The Saccharomyces cerevisiae PCD1 gene encodes a peroxisomal nudix hydrolase active toward coenzyme A and its derivatives. J. Biol. Chem., 275: 32925-32930.. Cashel M (1969) The control of ribonucleic acid synthesis in Escherichia coli. IV. Relevance of unusual phosphorylated compounds from amino acid-starved stringent strains. J. Biol. Chem., 244: 3133-3141.. Chatterji D, Fujita N, and Ishihama A (1998) The mediator for stringent control, ppGpp, binds to the beta-subunit of Escherichia coli RNA polymerase. Genes Cells, 3: 279-287.. Dalebroux ZD, Svensson SL, Gaynor EC, and Swanson MS (2010) ppGpp conjures bacterial virulence. Microbiol. Mol. Biol. Rev., 74: 171-199.. Daugherty M, Polanuyer B, Farrell M, Scholle M, Lykidis A, de Crécy-Lagard V, and Osterman A (2002) Complete reconstitution of the human coenzyme A biosynthetic pathway via comparative genomics. J. Biol. Chem., 277: 21431-21439.. Denton H, Roberts CW, Alexander J, Thong KW, and Coombs GH (1996) Enzymes of energy metabolism in the bradyzoites and tachyzoites of Toxoplasma gondii. FEMS Microbiol. Lett., 137: 103-108.. Dichtl B, Stevens A, and Tollervey D (1997) Lithium toxicity in yeast is due to the inhibition of RNA processing enzymes. EMBO. J., 16: 7184-7195.. 48.
(55) Dunn CA, O'Handley SF, Frick DN, and Bessman MJ (1999) Studies on the ADP-ribose pyrophosphatase subfamily of the nudix hydrolases and tentative identification of trgB, a gene associated with tellurite resistance. J. Biol. Chem., 274: 32318-32324.. Falk KL and Guerra DJ (1993) Coenzyme A biosynthesis in plants: partial purification and characterization of pantothenate kinase from spinach. Arch. Biochem. Biophys., 301: 424-430.. Fisher DI, Cartwright JL, Harashima H, Kamiya H, and McLennan AG (2004) Characterization of a nudix hydrolase from Deinococcus radiodurans with a marked specificity for (deoxy)ribonucleoside 5’-diphosphates. BMC Biochem., 5: 7.. Fisher DI, Safrany ST, Strike P, McLennan AG, and Cartwright JL (2002) Nudix hydrolases that degrade dinucleoside and diphosphoinositol polyphosphates also have 5-phosphoribosyl 1-pyrophosphate (PRPP) pyrophosphatase activity that generates the glycolytic activator ribose 1,5-bisphosphate. J. Biol. Chem., 277: 47313-47317.. Flugel RS, Hwangbo Y, Lambalot RH, Cronan JE Jr, and Walsh CT (2000) Holo-(acyl carrier protein) synthase and phosphopantetheinyl transfer in Escherichia coli. J. Biol. Chem., 275: 959-968.. Frick DN, and Bessman MJ (1995) Cloning, purification, and properties of a novel NADH pyrophosphatase. Evidence for a nucleotide pyrophosphatase catalytic domain in MutT-like enzymes. J. Biol. Chem., 270: 1529-1534.. Friga GM, Borbély G, and Farkas GL (1981) Accumulation of guanosine tetraphosphate (ppGpp) under nitrogen starvation in Anacystis nidulans, a cyanobacterium. Arch. Microbiol., 129: 341-343.. 49.
図


+7




関連したドキュメント
Standard domino tableaux have already been considered by many authors [33], [6], [34], [8], [1], but, to the best of our knowledge, the expression of the
As in the previous case, their definition was couched in terms of Gelfand patterns, and in the equivalent language of tableaux it reads as follows... Chen and Louck remark ([CL], p.
The edges terminating in a correspond to the generators, i.e., the south-west cor- ners of the respective Ferrers diagram, whereas the edges originating in a correspond to the
To obtain the optimal time decay rates of the higher-order derivatives of the solution, we can represent the spatial derivatives of the solutions to the equation U t = BU + G with
For the multiparameter regular variation associated with the convergence of the Gaussian high risk scenarios we need the full symmetry group G , which includes the rotations around
理工学部・情報理工学部・生命科学部・薬学部 AO 英語基準入学試験【4 月入学】 国際関係学部・グローバル教養学部・情報理工学部 AO
[Mag3] , Painlev´ e-type differential equations for the recurrence coefficients of semi- classical orthogonal polynomials, J. Zaslavsky , Asymptotic expansions of ratios of
This makes a somewhat more sophisticated analysis of the behaviour of that vertex necessary, which represents the curvature minimum (Lemma 3). [Gr1, § 2, Main Theorem]), we are able
