Journal of Biomechanical Science and Engineering
Progress and challenges in vascular tissue engineering
using self-organization/pre-designed approaches
1. Introduction
Currently, organ transplantation is the most effective therapy for end-stage organ failure. However, the demand for life-saving organ transplants far exceeds the supply of available organs, including the kidney, liver, heart, and lung owing to organ shortage (Lauerer et al., 2016). To address this problem, tissue engineering has offered potential strategies for the construction of solid tissues and organs using cells, signaling factors, and scaffolds (Langer and Vacanti, 1993). Over the last several decades, various approaches have been developed for the construction of organ-specific tissues by utilizing cell culture platforms, such as porous scaffolds and decellularized matrices, and scaffold-free culture methods, including cell sheet engineering and spheroid culture (Uygun et al., 2010; Bao et al., 2011; No et al., 2012; Damania et al., 2014). Despite great progress, limitations remain for medical and clinical applications of tissue-engineered constructs owing to the lack of functional vascular networks, which may result in tissue dysfunction, such as hemorrhage, clot formation, and tissue hypoxia.
Functional vascular networks should be constructed in tissue-engineered constructs. Establishing a hierarchical tubular network is required to supply oxygen and nutrients to the tissue constructs. In particular, following morphological
Masafumi WATANABE*, *** and Ryo SUDO*, **
*School of Integrated Design Engineering, Keio University, 3-14-1 Hiyoshi, Kohoku-ku, Yokohama 223-8522, Japan **Department of System Design Engineering, Keio University, 3-14-1 Hiyoshi, Kohoku-ku, Yokohama 223-8522, Japan
***Current affiliation: Graduate School of Medicine, Ehime University, Shitsukawa, Toon, Ehime 791-0295, Japan E-mail: [email protected]
Abstract
Organ transplantation is the most effective therapy for end-stage organ failure. However, the demand for life-saving organ transplants far exceeds the supply of available organs owing to organ shortage. To address this problem, tissue engineering has offered potential strategies for in vitro construction of organs as medical and clinical applications. However, tissue-engineered organs are difficult to construct owing to the lack of functional vascular networks because avascular organs lead to tissue dysfunctions, such as hypoxia and clot formation. Therefore, establishing functional vascular networks is required for the construction and maintenance of organs in terms of morphology and function. Recent advances in tissue engineering have allowed the in vitro construction of a wide range of functional vascular networks, ranging from microvessels to organ-scale vascular networks, using self-organization and pre-designed approaches. In particular, various new models have been developed utilizing microfluidics, 3D bioprinting, and organ decellularization. These models have enabled the in vitro recapitulation of key features of physiological vascular networks, such as morphology (e.g., network formation, luminal structure, and perivascular cell coverage) and function (e.g., barrier and antithrombogenic functions). In this review, we summarize the progress and challenges in vascular tissue engineering based on two distinct categories: self-organization and pre-designed approaches. In addition, the advantages and limitations of these models are highlighted, and future perspectives are discussed. These models will provide useful insights for the construction of vascularized functional tissues and organs and can contribute to development in tissue engineering and regenerative medicine.
Keywords : Vascular networks, Tissue engineering, Self-organization approach, Pre-designed approach,
Microfluidics, 3D bioprinting, Organ decellularization
【
Review Paper】
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
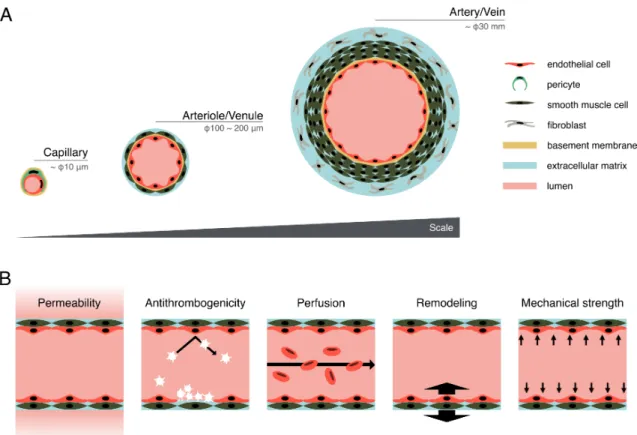
features are required to develop an ideal vascular model: i) continuous hierarchical network ranging from the micrometer-scale capillaries to millimeter-micrometer-scale arteries/veins, ii) lumen structure, iii) multilayer structure including endothelial cells (ECs) and perivascular cells such as, pericytes, smooth muscle cells (SMCs), and fibroblasts, iv) formation of basement membrane, and v) junction formation between ECs (Fig. 1A). Moreover, the functional features of vascular networks, such as i) permeability of biochemical factors, ii) antithrombogenic function, iii) blood perfusion, iv) vascular remodeling, and v) mechanical strength to blood pressure, are required for tissue growth and homeostatic maintenance (Michiels, 2003; Pashneh-tala et al., 2016) (Fig. 1B). The formation of vascular networks in tissue-engineered constructs improves their performance after transplantation (Jain et al., 2005; Cheng et al., 2011; Takebe et al., 2013). Therefore, functional vascular networks should be constructed to recapitulate physiological features of vasculatures in terms of morphology and function.
Recent advances in tissue engineering have allowed the construction of a wide range of vascular networks ranging from capillaries/microvessels to organ-scale vascular networks by combining biomaterials with various technologies, such as microfluidics, 3D bioprinting, and organ decellularization (Bogorad et al., 2015; Hussein et al., 2020). In this review, the progress and challenges of various techniques for in vitro vascular formation are summarized according to two distinct categories: self-organization (cell-based) approaches and pre-designed (scaffold-based) approaches, focusing on morphology and function of the vascular networks. Self-organization approaches have been utilized mainly for the construction of smaller vessels, such as capillaries/microvessels, while pre-designed approaches are suitable for the construction of larger vessels, such as arteries/veins. Finally, we discuss future perspectives for in vitro construction of solid tissues and organs in terms of vascular formation.
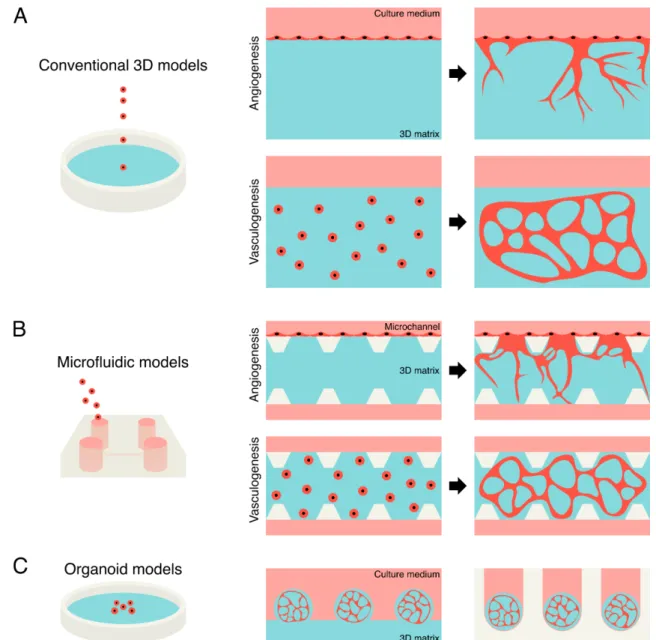
Fig. 1 Morphological and functional features of an ideal vascular model. (A) Morphological features of the ideal vascular model, such as hierarchical networks including capillary, arteriole/venule, and artery/vein, lumen structure, and multilayer structure including ECs, pericytes, SMCs, and fibroblasts. (B) Functional features of the ideal vascular model, such as permeability of biochemical factors, antithrombogenic function, blood perfusion, vascular remodeling, and mechanical strength to blood pressure.
2. Self-organization approaches
Since Folkman et al. (1980) reported in vitro angiogenesis, there has been growing evidence that vascular cells, such as ECs, have the capacity to self-organize into vascular networks in vitro under appropriate culture conditions (Czirok
and Little, 2012). For example, rat aortic explants cultured in clotted chick plasma provided vascular outgrowth and developed a complex vascular network mimicking in vivo angiogenesis (Nicosia et al., 1982). In addition to the aorta model, bovine ECs were capable of migrating into a three-dimensional (3D) matrix and forming interconnected networks when plated on a collagen matrix, which is a major component of the extracellular matrix (ECM) (Schor et al., 1983). Moreover, several studies have demonstrated that ECs can self-organize into tube-like structures when cultured on a two-dimensional (2D) matrix of basement membrane extract (Kubota et al., 1988; DeCicco-Skinner et al., 2014) or when suspended within a 3D collagen matrix via vasculogenesis (Davis and Camarillo, 1996). To construct vascular networks in vitro, biomaterials (e.g., collagen and fibrin) have been widely used as 3D matrices. Collagen is the main component of connective tissues and is abundant in the physiological mammalian body, while fibrin is the key factor of the thrombosis development under pathological conditions, such as wound healing. Over the last several decades, various self-organization approaches, such as conventional 3D matrix models, microfluidic models, and vascularized tissue models, have been developed to construct vascular networks in vitro via angiogenesis and vasculogenesis.
2.1 Conventional 3D models
In vitro angiogenesis and vasculogenesis models using 3D matrices are pioneering works in tissue engineering for the construction of vascular networks in culture. These models have been used to study in vitro morphogenesis of ECs, such as vascular sprouting, lumen formation, and vascular maturation owing to their reproducibility and simplicity (Vailhé et al., 2001). Several approaches have been developed to construct vascular networks in vitro based on the conventional angiogenesis and vasculogenesis models (Table 1).
2.1.1 In vitro angiogenesis models
Angiogenesis is defined as the formation of new blood vessels and capillaries from pre-existing blood vessels. An ex vivo aorta model is a conventional angiogenesis model. For example, this model demonstrated that fibronectin incorporated in the matrix promoted elongation of the newly formed microvessels when aortic explants were embedded in a collagen matrix (Nicosia et al., 1993). Moreover, several studies using an in vitro angiogenesis model reported that ECs invaded into 3D matrices and underwent lumen formation in response to exogenous factors such as sphingosine-1-phosphate (S1P), stromal cell-derived factor-1α, or basic fibroblast growth factor (bFGF) (Davis et al., 2000; Bayless and Davis, 2003; Saunders et al., 2006; Ueda et al., 2006) (Fig. 2A, upper panel). Hypoxia conditions can also promote the formation of 3D vascular networks within a collagen matrix by increased gene expression of angiogenic factors, such as vascular endothelial growth factor (VEGF) families and FGF (Ueda et al., 2008). Endothelial progenitor cells (EPCs) also contributed to 3D vascular network formation by secreting angiogenic factors, such as VEGF (Koga et al., 2009; Abe et al., 2013a). These findings suggest that in vitro vascular formation can be stimulated and regulated by various biochemical factors.
Vascular formation can also be regulated by mechanical factors, such as matrix stiffness and fluid shear stress (FSS). For example, matrix stiffness can regulate in vitro vascular morphogenesis when ECs are cultured in rigid or flexible gel (Yamamura et al., 2007). ECs formed thicker and deeper networks within rigid gel, whereas thin and dense networks were formed in flexible gel. In a parallel-plate flow chamber, ECs on a collagen matrix under flow conditions promoted the growth of vascular networks within the matrix compared to static conditions (Ueda et al., 2004). In addition, steady FSS promoted the formation of interconnected and stable vascular networks, while pulsatile FSS induced retraction of vascular networks, resulting in the formation of unstable networks (Abe et al., 2013b). Furthermore, a combination of FSS and angiogenic factors, including S1P, enhanced EC invasion into a collagen matrix via phosphorylation of Akt and matrix metalloproteinase (MMP)-2 activation (Kang et al., 2008). These findings suggest that mechanical factors can be used to control the morphology of vascular networks in vitro.
2.1.2 In vitro vasculogenesis models
Vasculogenesis is defined as the process of vascular formation in the absence of pre-existing blood vessels, which are conducted by EPCs during embryogenesis. In vitro vasculogenesis models have been widely used to study vascular morphogenesis (Fig. 2A, lower panel). For example, this model has been used to study multicellular interaction between ECs and perivascular cells (e.g., pericytes) during in vitro vascular formation because perivascular cells have pivotal roles in the formation and maintenance of vascular networks in vivo (Ribatti et al., 2011). In EC-pericyte coculture within a collagen matrix, bovine pericytes were shown to associate with developing EC tubes and stabilize their tube structures
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
by blocking activation of MMPs (Saunders et al., 2006). Moreover, a vasculogenesis model revealed that pericytes facilitated maturation events, including the deposition of a vascular basement membrane matrix of collagen IV and laminin during in vitro vascular formation (Stratman et al., 2009). Moreover, several studies also demonstrated that supporting cells, such as mesenchymal stem cells (MSCs) and human adipose-derived stromal cells (ASCs), could be used to construct stable vascular networks when the cells are embedded with ECs within 3D matrices, such as collagen, fibrin, glycosaminoglycan, and Matrigel (Rao et al., 2012; McFadden et al., 2013; Andrée et al., 2019). Glycosaminoglycan is a component of ECM present on the luminal surface of vascular endothelium, while Matrigel is a basement membrane ECM.
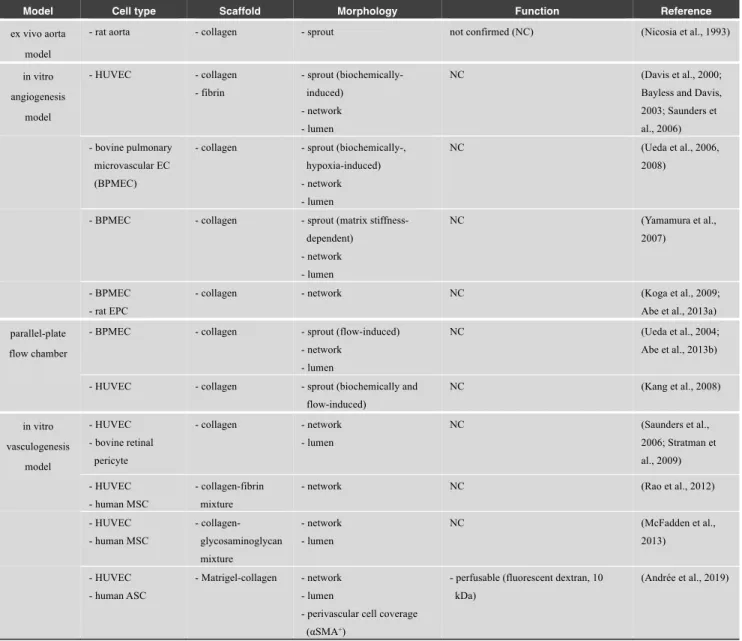
Table 1 List of conventional angiogenesis and vasculogenesis models
Model Cell type Scaffold Morphology Function Reference
ex vivo aorta model
- rat aorta - collagen - sprout not confirmed (NC) (Nicosia et al., 1993)
in vitro angiogenesis model - HUVEC - collagen - fibrin - sprout (biochemically-induced) - network - lumen NC (Davis et al., 2000; Bayless and Davis, 2003; Saunders et al., 2006) - bovine pulmonary
microvascular EC (BPMEC)
- collagen - sprout (biochemically-, hypoxia-induced) - network - lumen
NC (Ueda et al., 2006,
2008)
- BPMEC - collagen - sprout (matrix stiffness-dependent) - network - lumen NC (Yamamura et al., 2007) - BPMEC - rat EPC
- collagen - network NC (Koga et al., 2009; Abe et al., 2013a) parallel-plate
flow chamber
- BPMEC - collagen - sprout (flow-induced) - network
- lumen
NC (Ueda et al., 2004;
Abe et al., 2013b) - HUVEC - collagen - sprout (biochemically and
flow-induced) NC (Kang et al., 2008) in vitro vasculogenesis model - HUVEC - bovine retinal pericyte - collagen - network - lumen NC (Saunders et al., 2006; Stratman et al., 2009) - HUVEC - human MSC - collagen-fibrin mixture
- network NC (Rao et al., 2012)
- HUVEC - human MSC - collagen-glycosaminoglycan mixture - network - lumen NC (McFadden et al., 2013) - HUVEC - human ASC - Matrigel-collagen - network - lumen
- perivascular cell coverage (αSMA+)
- perfusable (fluorescent dextran, 10 kDa)
(Andrée et al., 2019)
2.2 Microfluidic models
Although conventional angiogenesis and vasculogenesis models have demonstrated that ECs can form vascular networks in vitro, there remain some technical limitations, such as spatial and temporal manipulation of cells, precise fluid control mimicking physiological environments, and live-cell analyses of vascular functions (e.g., barrier function and antithrombogenic function). To address these problems, microfabrication techniques were utilized for a cell culture platform. In 1979, a miniaturized gas chromatographic analyzer was developed using photolithography and chemical etching techniques, which enabled precise fluid control and high-throughput analysis (Terry et al., 1979). This device is now widely regarded as the first “Lab on a Chip” (Convery and Gadegaard, 2019). By applying these techniques, a microfluidic device was developed as a 3D cell culture platform (Smith and Gerecht, 2014). Microfluidic devices have
been widely used to understand how the complex cellular microenvironment directs vascular cell behavior and functionality (Table 2).
2.2.1 Microfluidic angiogenesis models
In 2008, a novel microfluidic platform was developed that could mimic the key features of in vivo microenvironments, such as FSS, interstitial flow through the matrix, gradients of biochemical factors, and the properties of gel scaffolds (Vickerman et al., 2008). This platform also enabled real-time monitoring of cells in response to changes in their microenvironment. In their study, human adult dermal microvascular ECs (HMVECs) invaded into a 3D collagen matrix and formed open luminal structures in a microfluidic device (Fig. 2B, upper panel). This was confirmed by 3D confocal images and perfusion with fluorescent microspheres. Using a similar platform, several types of ECs, including HMVECs and rat microvascular ECs (rMVECs), demonstrated the formation of vascular networks (Chung et al., 2009; Sudo et al., 2009; Shin et al., 2012). Because microfluidic devices allow the precise control of fluid flow, the mediation of vascular formation in vitro by biophysical factors owing to shear flow and interstitial flow has been investigated. For example, vascular sprouting was attenuated when ECs were exposed to FSS using a syringe pump, while interstitial flow triggered the formation of vascular sprouts and anastomoses between two pre-existing vessels (Song and Munn, 2011). Both intraluminal and transmural flow have triggered the formation of vascular sprouts into a collagen matrix (Galie et al., 2014). Moreover, a recent study demonstrated that the balance between the interstitial flow magnitude and VEGF concentration played an important role in the regulation of 3D microvascular network formation in vitro (Abe et al., 2019). A combination of VEGF, S1P, and phorbol 12-myristate 13-acetate provided the optimal cocktail for 3D angiogenic sprouting under hydrostatic pressure-driven flow conditions (van Duinen et al., 2019). These studies suggested that mechanical and biochemical factors can control the morphology of vascular networks, such as sprout formation in a microfluidic device.
Stable and mature vascular structures are required to construct functional vascular networks in vitro. However, vascular networks are difficult to maintain in an EC monoculture primarily owing to vascular regression. To address this problem, heterotypic cell culture models can be used for the construction of stable and mature vascular networks in a microfluidic device. For example, fibroblasts secrete various proangiogenic factors and induce vascular sprouting within a fibrin matrix (Newman et al., 2011). Similar results were obtained in a microfluidic device. HUVECs formed vascular networks with perfusable lumens when they were cultured with normal human lung fibroblasts (NHLFs) in a 3D fibrin-collagen matrix (Yeon et al., 2012). In this model, a tight junction protein, zonula occludens-1, was expressed between ECs. In addition, the deposition of basement membrane components, such as laminin and collagen IV, was observed around the perivascular ECM, which is a sign of vessel maturation (Kim et al., 2013). Permeability measurement using fluorescent molecules revealed that microvessels constructed in a microfluidic device exhibited reliable barrier functions similar to those measured in vivo (Lee et al., 2014).
To obtain stable and mature vascular networks, in vivo interaction between ECs and perivascular cells, such as pericytes, should be recapitulated. For example, several studies demonstrated that EC-pericyte coculture recapitulated the physiological capillary networks in terms of morphology, which corresponds to the smallest part of the circulatory system. To mimic capillary structures, human placental pericytes were cultured in HUVEC-NHLF coculture (Kim et al., 2013, 2015). This report showed pericyte-coverage and decreased permeability of vascular networks, which were similar to those in vivo. Furthermore, recent studies have demonstrated that MSCs can be a potential cell source for pericytes (Yamamoto et al., 2013; Watanabe and Sudo, 2019, 2021). In these studies, MSCs differentiated into pericyte-like perivascular cells in HUVEC-MSC coculture in vitro. In HUVEC-MSC coculture, HUVECs formed vascular networks and MSCs located around the microvascular networks. Moreover, the constructed capillary networks had continuous lumens with a < 10 µm diameter, and basement membranes, which contained collagen IV and laminin, were formed around the capillary structures (Yamamoto et al., 2019). These structures were maintained for at least 21 days. In this coculture model, MSC-derived perivascular cells were positive for α smooth muscle actin (αSMA), neural/glial antigen 2 (NG2), and platelet-derived growth factor receptor β (PDGFRβ) (Yamamoto et al., 2019; Uwamori et al., 2019). These markers are known as in vivo pericyte markers, while αSMA is expressed in the pathological state (Armulik et al., 2011).
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
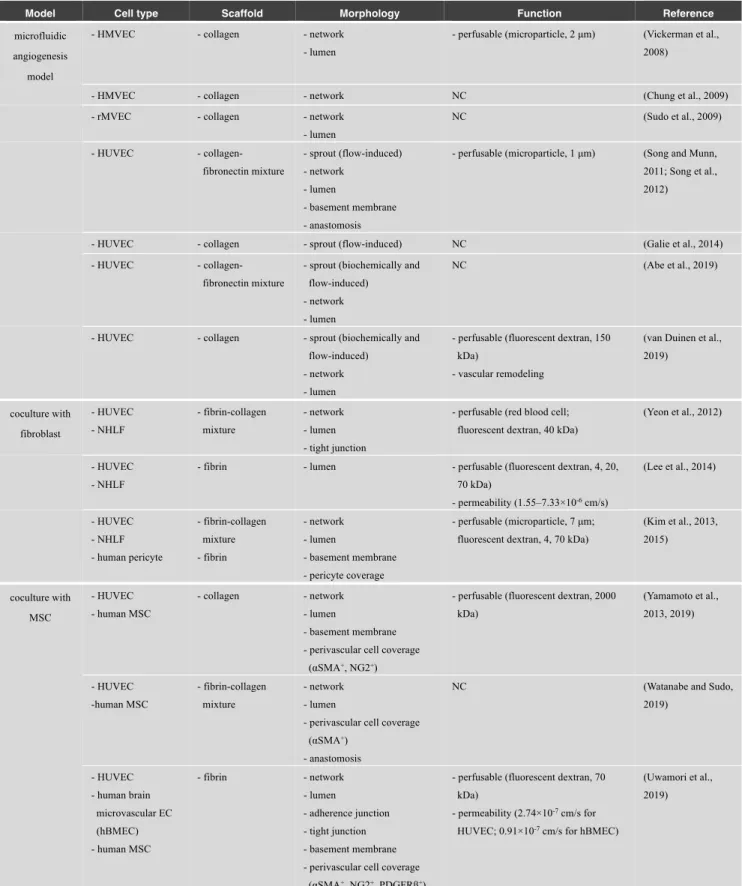
Table 2 List of microfluidic angiogenesis models
Model Cell type Scaffold Morphology Function Reference
microfluidic angiogenesis
model
- HMVEC - collagen - network - lumen
- perfusable (microparticle, 2 μm) (Vickerman et al., 2008)
- HMVEC - collagen - network NC (Chung et al., 2009) - rMVEC - collagen - network
- lumen NC (Sudo et al., 2009) - HUVEC - collagen-fibronectin mixture - sprout (flow-induced) - network - lumen - basement membrane - anastomosis
- perfusable (microparticle, 1 μm) (Song and Munn, 2011; Song et al., 2012)
- HUVEC - collagen - sprout (flow-induced) NC (Galie et al., 2014) - HUVEC -
collagen-fibronectin mixture
- sprout (biochemically and flow-induced)
- network - lumen
NC (Abe et al., 2019)
- HUVEC - collagen - sprout (biochemically and flow-induced)
- network - lumen
- perfusable (fluorescent dextran, 150 kDa)
- vascular remodeling
(van Duinen et al., 2019) coculture with fibroblast - HUVEC - NHLF - fibrin-collagen mixture - network - lumen - tight junction
- perfusable (red blood cell; fluorescent dextran, 40 kDa)
(Yeon et al., 2012)
- HUVEC - NHLF
- fibrin - lumen - perfusable (fluorescent dextran, 4, 20, 70 kDa) - permeability (1.55–7.33×10-6 cm/s) (Lee et al., 2014) - HUVEC - NHLF - human pericyte - fibrin-collagen mixture - fibrin - network - lumen - basement membrane - pericyte coverage - perfusable (microparticle, 7 μm; fluorescent dextran, 4, 70 kDa)
(Kim et al., 2013, 2015) coculture with MSC - HUVEC - human MSC - collagen - network - lumen - basement membrane - perivascular cell coverage
(αSMA+, NG2+)
- perfusable (fluorescent dextran, 2000 kDa) (Yamamoto et al., 2013, 2019) - HUVEC -human MSC - fibrin-collagen mixture - network - lumen
- perivascular cell coverage (αSMA+)
- anastomosis
NC (Watanabe and Sudo,
2019) - HUVEC - human brain microvascular EC (hBMEC) - human MSC - fibrin - network - lumen - adherence junction - tight junction - basement membrane - perivascular cell coverage
(αSMA+, NG2+, PDGFRβ+)
- perfusable (fluorescent dextran, 70 kDa)
- permeability (2.74×10-7 cm/s for
HUVEC; 0.91×10-7 cm/s for hBMEC)
(Uwamori et al., 2019)
2.2.2 Microfluidic vasculogenesis models
Vasculogenesis can also be recapitulated using a microfluidic device (Table 3). In a microfluidic vasculogenesis model, ECs were suspended in a 3D matrix within a microchannel, which induced the formation of vascular networks with lumens (Park et al., 2014) (Fig. 2B, lower panel). In their study, the fibrin matrix was preferable to collagen or collagen-fibrin matrices, by significantly reducing matrix retractions during maturation of microvascular networks for 7
days. A conventional vasculogenesis model also demonstrated that both pure fibrin and collagen-fibrin gel at a ratio of 2:3 were useful for forming vascular networks, in terms of total network length or number of segments (Rao et al., 2012). Moreover, several studies investigated the effects of biophysical factors owing to shear flow and interstitial flow on vasculogenesis. When the Péclet number (Pe), which is defined as the ratio of convective to diffusive transport, was controlled by hydrostatic pressure in a microfluidic device (Hsu et al., 2013), vasculogenesis was independently stimulated by interstitial flow (Pe > 10) or hypoxic conditions (Pe < 0.1) and not by the intermediate state in the HUVEC-fibroblast coculture. This platform is also useful as a perfusable vascular model because reliable barrier functions with minimal leakage of fluorescent molecules were observed (Moya et al., 2013; Wang et al., 2016).
To construct mature vascular networks, supporting cells, such as fibroblasts and MSCs, are important in vasculogenesis models (Carrion et al., 2010; Jeon et al., 2014). ECs suspended within a 3D fibrin matrix executed a morphogenetic process similar to vasculogenesis, forming a primitive vascular plexus and maturing into robust microvascular networks with well-defined lumens. MSCs differentiated into αSMA-positive perivascular cells and wrapped around the newly formed microvascular networks. Moreover, a recent study demonstrated that intraluminal perfusion induced dilation of vascular networks in a microfluidic vasculogenesis model, which was composed of HUVECs and MSC-derived perivascular cells (Sano et al., 2020). This model recapitulated the key features of outward vascular remodeling in vivo. These findings suggest that in vitro vascular morphogenesis can be controlled by mechanical fluid force, chemical gradients, and heterotypic cell–cell interactions.
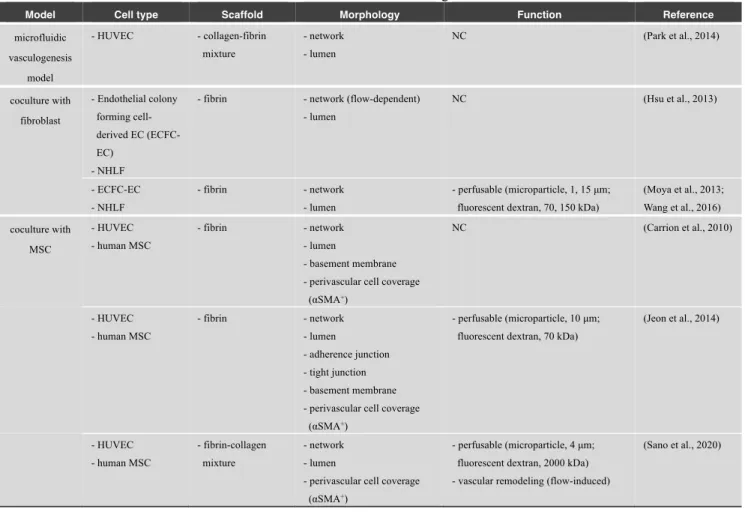
Table 3 List of microfluidic vasculogenesis models
Model Cell type Scaffold Morphology Function Reference
microfluidic vasculogenesis model - HUVEC - collagen-fibrin mixture - network - lumen NC (Park et al., 2014) coculture with fibroblast - Endothelial colony forming cell-derived EC (ECFC-EC) - NHLF
- fibrin - network (flow-dependent) - lumen NC (Hsu et al., 2013) - ECFC-EC - NHLF - fibrin - network - lumen - perfusable (microparticle, 1, 15 μm; fluorescent dextran, 70, 150 kDa)
(Moya et al., 2013; Wang et al., 2016) coculture with MSC - HUVEC - human MSC - fibrin - network - lumen - basement membrane - perivascular cell coverage
(αSMA+) NC (Carrion et al., 2010) - HUVEC - human MSC - fibrin - network - lumen - adherence junction - tight junction - basement membrane - perivascular cell coverage
(αSMA+)
- perfusable (microparticle, 10 μm; fluorescent dextran, 70 kDa)
(Jeon et al., 2014) - HUVEC - human MSC - fibrin-collagen mixture - network - lumen
- perivascular cell coverage (αSMA+)
- perfusable (microparticle, 4 μm; fluorescent dextran, 2000 kDa) - vascular remodeling (flow-induced)
(Sano et al., 2020)
2.3 Vascularized tissue models
Generation of vascularized tissues remains a significant challenge in tissue engineering and regenerative medicine. In recent years, self-organization approaches have been used to construct tissues with vascular networks (Table 4). For example, 3D vascularized cardiac tissues were obtained when cardiac cell sheets with ECs were cocultured on a vascular bed, which were perfused with a culture medium using a bioreactor (Sekine et al., 2013). The formation of in vitro organ buds is also a promising approach. In this model, induced pluripotent stem cell (iPSC)-derived hepatic cells self-organized
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
into 3D functional liver buds by recapitulating organogenetic interactions between ECs and mesenchymal cells (Takebe et al., 2013). Luminal structures of vascular networks within in vitro liver buds were confirmed by perfusion of fluorescent molecules in mice after transplantation. In addition to liver buds, this approach was also applied to other organ systems, such as the intestine, lung, kidney, heart, and brain (Takebe et al., 2015). Moreover, a recent study showed the development of a clinical-scale production platform for human liver bud organoids and the generation of liver buds entirely from iPSC-derived cells (Takebe et al., 2017). A recent work demonstrated the development of self-organizing 3D human blood vessel organoids from iPSCs (Fig. 2C). These human blood vessel organoids contained ECs and pericytes that self-organized into microvascular networks with basement membranes (Wimmer et al., 2019). These functional vascularized tissue concepts provide promising approaches toward regenerative medicine and clinical use.
Fig. 2 Representative illustrations of self-organization approaches. (A) Conventional 3D models of in vitro angiogenesis and vasculogenesis. In an angiogenesis model, ECs in a monolayer invade into an underlying 3D matrix and form vascular networks. In a vasculogenesis model, ECs embedded within a 3D matrix form connected vascular networks. (B) Microfluidic models of in vitro angiogenesis and vasculogenesis models. ECs form vascular networks within a 3D matrix formed in a microchannel. (C) Organoid models. Vascularized organoids are developed using 3D matrices and low-attachment substrates.
Table 4 List of vascularized tissue models
Model Cell type Scaffold Morphology Function Reference
cardiac cell sheet
- rat EC - rat cardiac cell
- none - network - lumen
- perfusable (microparticle, 4 μm; fluorescent dextran, 2000 kDa)
(Sekine et al., 2013)
organ buds - human EC - human MSC - organ-specific cell
- Matrigel - network - lumen
- perfusable (fluorescent dextran, 70 and 2000 kDa) after transplantation into mice
(Takebe et al., 2013, 2015, 2017) vascular
organoid
- human iPSC - Matrigel-collagen - network - lumen - tight junction - basement membrane - perivascular cell coverage
(PDGFRβ+)
- perfusable (fluorescent dextran, 70 kDa) after transplantation into mice
(Wimmer et al., 2019)
3. Pre-designed approaches
Cells are unable to self-organize into complex 3D structures without appropriate environments. Therefore, a scaffold can be used to provide a template of the required construct, as a popular approach in tissue engineering. A range of synthetic and natural materials has been evaluated as scaffolds in vascular tissue engineering (Pashneh-tala et al., 2016).
3.1 Tissue-engineered vascular graft (TEVG) models
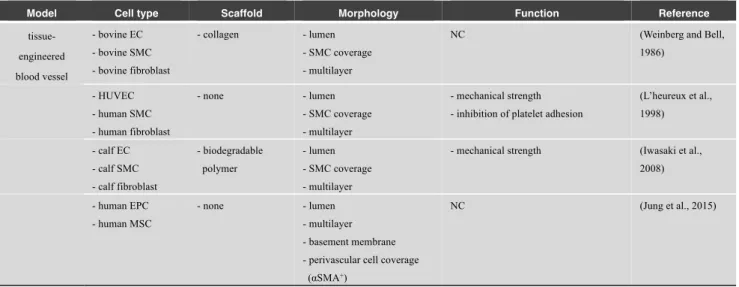
In 1986, the first TEVG model was developed by mimicking the multilayer structure of an in vivo artery (Weinberg and Bell, 1986). Collagen and bovine aortic SMCs were cast together to create an annular mold. The inner surface of the graft was seeded with bovine ECs, and the outer surface was seeded with bovine fibroblasts, resulting in the formation of a tubular structure in vitro (Fig. 3A). Moreover, multilayered TEVG can be fabricated without biological scaffolds by utilizing cell sheet engineering (L’heureux et al., 1998). Other reports also showed the construction of physiologically-relevant multilayer structures, including in vivo vascular intima, media, and adventitia, utilizing biodegradable polymers (e.g., polyglycolic acid and polycaprolactone) or the cell sheet technique to fabricate tubular structures (Iwasaki et al., 2008; Jung et al., 2015) (Table 5). These studies have primarily focused on the clinically relevant size of blood vessels and enabled the construction of larger blood vessels in vitro ranging from the submillimeter to centimeter size, which provides alternative clinical approaches to the transplantation of autologous vessels, such as the saphenous vein and internal thoracic artery. However, to achieve an effective delivery of oxygen and nutrients by convection and diffusion, large blood vessels and microvascular networks are required, especially in the peripheral tissues (Pries and Secomb, 2014).
Table 5 List of tissue-engineered vascular graft models
Model Cell type Scaffold Morphology Function Reference
tissue-engineered blood vessel - bovine EC - bovine SMC - bovine fibroblast - collagen - lumen - SMC coverage - multilayer
NC (Weinberg and Bell,
1986) - HUVEC - human SMC - human fibroblast - none - lumen - SMC coverage - multilayer - mechanical strength - inhibition of platelet adhesion
(L’heureux et al., 1998) - calf EC - calf SMC - calf fibroblast - biodegradable polymer - lumen - SMC coverage - multilayer
- mechanical strength (Iwasaki et al., 2008) - human EPC - human MSC - none - lumen - multilayer - basement membrane - perivascular cell coverage
(αSMA+)
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
3.2 Microfluidic pre-designed models
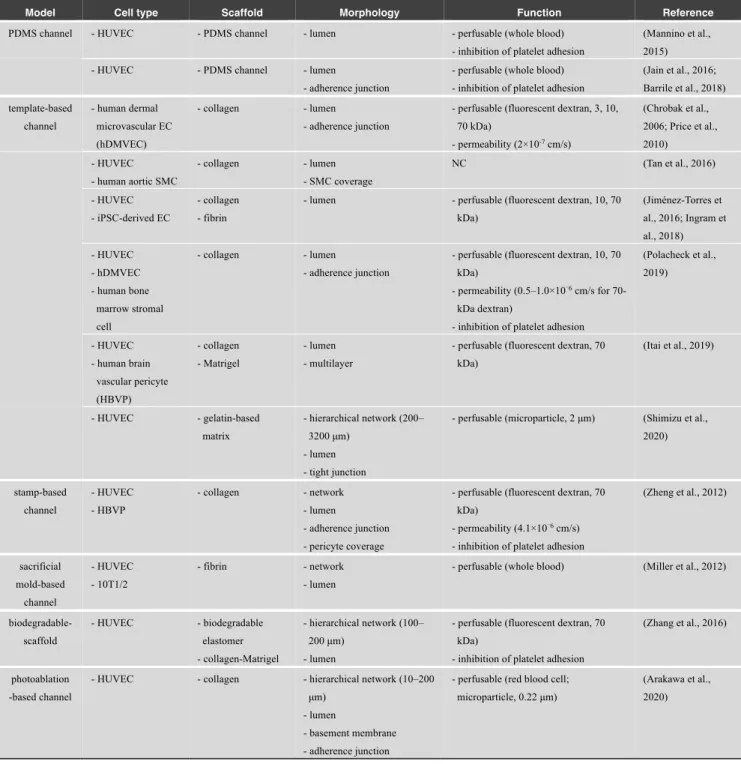
Because microfluidic techniques were introduced for the development of 3D culture devices, various models have been developed using pre-designed templates including microneedles and sacrificial molds (Table 6). Several studies have mimicked the luminal structures of blood vessels by lining ECs inside a polydimethylsiloxane (PDMS)-based microfluidic channel. This approach allowed the development of in vitro vascular models that recapitulated in vivo geometries, such as aneurysms, stenoses, and bifurcations. The endothelial monolayers formed on the luminal space of these vascular models reduced platelet adhesion inside the PDMS microchannels (Mannino et al., 2015). Conversely, other models showed the formation of platelet-rich thrombi under pathophysiological conditions (e.g., FSS and cytokine stimulation) inside the PDMS microchannels with endothelium (Jain et al., 2016; Barrile et al., 2018). Thus, these models can recapitulate both physiological and pathological conditions in vitro.
In addition to the PDMS microchannels, hydrogel-based microchannels have been developed in recent studies (Fig. 3B). For example, a cylindrical microchannel was formed in collagen gel using a stainless steel microneedle as a template (Chrobak et al., 2006). Confluent EC monolayers were formed in the microchannel, which displayed low permeability without chemical stimulation. Constructed microvessels exposed to intraluminal flow showed a barrier function similar to that observed in vivo, and these structures were maintained for at least 14 days (Price et al., 2010). This microneedle-based approach can also be used to form multilayered vascular structures. In an EC-SMC coculture model, an EC monolayer lining inside a collagen microchannel was covered by a lateral SMC layer, which mimicked an in vivo artery (Tan et al., 2016). In addition, flexible cylindrical PDMS rods were developed as a template for the formation of bifurcated microchannels in collagen and fibrin gels (Jiménez-Torres et al., 2016; Ingram et al., 2018). Moreover, a microneedle-based approach was utilized to perform vascular permeability tests in vitro (Polacheck et al., 2019). An alternative approach demonstrated the generation of vascular-like structures by detaching confluent EC monolayers from gold rods in a gelatin methacrylate (GelMA) matrix, which resulted in the formation of vascular structures with lumens (Sadr et al., 2011). Furthermore, a recent study fabricated a collagen-based perfusable microtube using a glass capillary and tungsten wire (Itai et al., 2019). This approach enabled the coculture of heterogeneous cell types in each layer. For example, ECs were seeded on the inner surface of the collagen tube, while pericytes were encapsulated within the collagen layer, mimicking the multilayer structure of in vivo microvessels. These pre-designed models were primarily utilized to construct vessels with a vascular diameter ranging from 50 μm to 1 mm in vitro.
As well as multilayer structures of blood vessels, several microfluidic models have mimicked hierarchical structures of vascular networks. For example, a 3D stamping technique allowed the formation of lattice vascular networks composed of 100 μm microvessels in a collagen matrix (Zheng et al., 2012). In addition, sacrificial lattice molds were used to fabricate microfluidic channels within a fibrin matrix (Miller et al., 2012). The result showed the generation of 3D lattice vascular networks with perfusable lumens. Biodegradable materials were also employed to construct hierarchical vascular networks, whose vascular widths ranged from 100 μm to 200 μm (Zhang et al., 2016). Furthermore, a recent study mimicked the complex geometries of vascular networks in vivo and fabricated complex microchannels with branches in a gelatin-based matrix using water-soluble sacrificial polyvinyl alcohol molds (Shimizu et al., 2020). Hierarchical vascular networks were generated whose vascular widths ranged from 200 μm to 3.2 mm. In contrast, smaller microchannels mimicking in vivo capillaries were fabricated by multiphoton ablation, resulting in the formation of vascular networks with vascular widths ranging from 10 μm to 200 μm (Arakawa et al., 2020).
Table 6 List of pre-designed microfluidic models
Model Cell type Scaffold Morphology Function Reference
PDMS channel - HUVEC - PDMS channel - lumen - perfusable (whole blood) - inhibition of platelet adhesion
(Mannino et al., 2015) - HUVEC - PDMS channel - lumen
- adherence junction
- perfusable (whole blood) - inhibition of platelet adhesion
(Jain et al., 2016; Barrile et al., 2018) template-based channel - human dermal microvascular EC (hDMVEC) - collagen - lumen - adherence junction
- perfusable (fluorescent dextran, 3, 10, 70 kDa) - permeability (2×10-7 cm/s) (Chrobak et al., 2006; Price et al., 2010) - HUVEC - human aortic SMC - collagen - lumen - SMC coverage NC (Tan et al., 2016) - HUVEC - iPSC-derived EC - collagen - fibrin
- lumen - perfusable (fluorescent dextran, 10, 70 kDa) (Jiménez-Torres et al., 2016; Ingram et al., 2018) - HUVEC - hDMVEC - human bone marrow stromal cell - collagen - lumen - adherence junction
- perfusable (fluorescent dextran, 10, 70 kDa)
- permeability (0.5–1.0×10−6 cm/s for
70-kDa dextran)
- inhibition of platelet adhesion
(Polacheck et al., 2019) - HUVEC - human brain vascular pericyte (HBVP) - collagen - Matrigel - lumen - multilayer
- perfusable (fluorescent dextran, 70 kDa) (Itai et al., 2019) - HUVEC - gelatin-based matrix - hierarchical network (200– 3200 μm) - lumen - tight junction
- perfusable (microparticle, 2 μm) (Shimizu et al., 2020) stamp-based channel - HUVEC - HBVP - collagen - network - lumen - adherence junction - pericyte coverage
- perfusable (fluorescent dextran, 70 kDa)
- permeability (4.1×10−6 cm/s)
- inhibition of platelet adhesion
(Zheng et al., 2012) sacrificial mold-based channel - HUVEC - 10T1/2 - fibrin - network - lumen
- perfusable (whole blood) (Miller et al., 2012)
biodegradable-scaffold - HUVEC - biodegradable elastomer - collagen-Matrigel - hierarchical network (100– 200 μm) - lumen
- perfusable (fluorescent dextran, 70 kDa)
- inhibition of platelet adhesion
(Zhang et al., 2016)
photoablation -based channel
- HUVEC - collagen - hierarchical network (10–200 μm)
- lumen
- basement membrane - adherence junction
- perfusable (red blood cell; microparticle, 0.22 μm)
(Arakawa et al., 2020)
3.3 3D bioprinting models
In the field of tissue engineering, 3D bioprinting has emerged as a promising technology for the construction of vascular networks (Table 7). This technology has allowed the precise control of spatial and temporal distribution of the cells and matrix in vitro (Fig. 3C). In pioneer studies, 3D printers have been utilized to assemble cells in vitro. For example, Kenzan bioprinting was developed for the generation of tubular tissues by assembling multicellular spheroids, including ECs, SMCs, and fibroblasts (Itoh et al., 2015). Moreover, 3D bioprinting has been applied to fabricate large-scale solid organs. To construct perfusable vascular networks within a dense cellular construct composed of cardiac spheroids, a hierarchical channel network was printed within the cellular construct using sacrificial ink (Skylar-Scott et al., 2019). After printing, the cellular construct was warmed to remove the sacrificial ink, which resulted in the formation of a perfusable cardiac cellular construct. Current bioprinting technologies have enabled the fabrication of 3D organ-scale tissues in vitro. For example, an alveolar model mimicking distal lung subunits was printed within a poly(ethylene glycol) diacrylate (PEGDA) hydrogel, which is a synthetic polymer, using stereolithography (Grigoryan et al., 2019).
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
Moreover, cellularized human hearts were fabricated within a patient-derived hydrogel (Noor et al., 2019). These technologies provide promising platforms for organ-scale tissue engineering.
Table 7 List of 3D bioprinting models
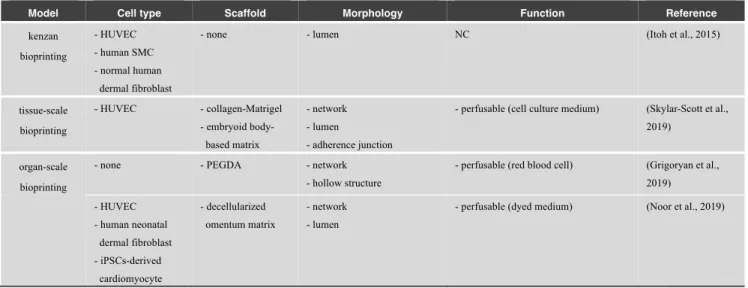
Model Cell type Scaffold Morphology Function Reference
kenzan bioprinting - HUVEC - human SMC - normal human dermal fibroblast
- none - lumen NC (Itoh et al., 2015)
tissue-scale bioprinting - HUVEC - collagen-Matrigel - embryoid body-based matrix - network - lumen - adherence junction
- perfusable (cell culture medium) (Skylar-Scott et al., 2019)
organ-scale bioprinting
- none - PEGDA - network - hollow structure
- perfusable (red blood cell) (Grigoryan et al., 2019) - HUVEC - human neonatal dermal fibroblast - iPSCs-derived cardiomyocyte - decellularized omentum matrix - network - lumen
- perfusable (dyed medium) (Noor et al., 2019)
3.4 Organ-decellularization models
Decellularization techniques have great potential for the generation of organ-scale hierarchical vascular networks, which is critical for the construction of large-scale solid organs. Decellularization completely removes the cellular and nuclear components from a tissue or organ while preserving its ECM around the parenchymal and vascular structures (Hussein et al., 2020). This approach has the distinct advantage of developing acellular matrices that preserve the mechanical, biological, and anatomical properties of the native tissue. In addition, decellularized matrices do not evoke a host immune response owing to the removal of cellular components. Currently, protocols for the preparation of acellular whole-organ matrices have been reported in terms of the liver, kidney, lung, and heart using detergent-based perfusion (Uygun et al., 2010; Guyette et al., 2014).
Several studies have constructed vascular networks in vitro using decellularized organs ranging from small animals to large animals (Table 8) (Fig. 3D). For example, ECs formed hierarchical vascular networks in a decellularized ferret liver with lumens, and antithrombogenic function was shown by reducing platelet deposition in the re-endothelialized liver tissue (Baptista et al., 2011). Hierarchical vascular networks were also formed in a rat decellularized kidney (Song et al., 2013). Furthermore, recent efforts allowed human-scale tissue construction using decellularized porcine organs. A previous study demonstrated reestablishment of hierarchical vascular networks with lumens in a decellularized porcine liver by conjugating anti-CD31 antibodies to maximize coverage of the vessel walls with ECs (Ko et al., 2015). The re-endothelialized liver was able to withstand physiological blood flow, prevent thrombosis, and deliver blood to the entire organ for up to 24 h when transplanted into a recipient pig. Heparin-gelatin coating was also useful to reestablish vascular networks in a decellularized porcine liver (Hussein et al., 2016). Ex vivo blood perfusion was maintained for 24 h, and thrombosis was not observed in the heparin-gelatin coated re-endothelialized livers. These results suggest that organ-decellularization models can recapitulate both morphological and functional features of vascular networks, including hierarchical networks and antithrombogenic function. However, maintaining tissue-engineered whole organs for long term is challenging owing to the lack of intact endothelium, including microvessels.
Table 8 List of organ-decellularization models
Model Cell type Scaffold Morphology Function Reference
small animal model - mouse EC (MS1) - decellularized (DC) ferret liver - hierarchical network - lumen - perfusable (microparticle, 5 μm; whole blood)
- inhibition of platelet adhesion
(Baptista et al., 2011)
- HUVEC - rat neonatal kidney
cell
- DC rat kidney - hierarchical network - lumen
- perfusable (whole blood) (Song et al., 2013)
large animal model
- MS1 - DC porcine liver - hierarchical network - lumen
- perfusable (whole blood) - inhibition of platelet adhesion
(Ko et al., 2015)
- human EA.hy926 EC
- HepG2
- DC porcine liver - hierarchical network - lumen
- perfusable (whole blood) - inhibition of platelet adhesion
(Hussein et al., 2016)
Fig. 3 Representative images of pre-designed approaches. (A) TEVG models. A multilayer structure can be formed using a tubular graft and vascular cells (e.g., ECs and SMCs). The inner surface of the graft is seeded with ECs, while SMCs are embedded within the graft. (B) Microfluidic pre-designed models, such as microneedle-based and stamp-based models. Confluent EC monolayers are formed in microchannels. Insets indicated by black frames show cross-section images of vascular structures. (C) 3D bioprinting models. This technology allows the precise spatiotemporal control of cells and matrices. (D) Organ-decellularization models. Organ-scale hierarchical vascular networks can be formed in decellularized organs.
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
4. Integration of self-organization and pre-designed approaches
Based on the previous studies described above, self-organization approaches can effectively construct smaller vascular networks, including microscale capillaries/microvessels (Yamamoto et al., 2019; Uwamori et al., 2019), while pre-designed approaches can construct larger vascular networks ranging from the submillimeter to centimeter size (Pashneh-tala et al., 2016). Therefore, integration of these two approaches may provide potential strategies for constructing complex vascular networks ranging from microvessels to large vessels.
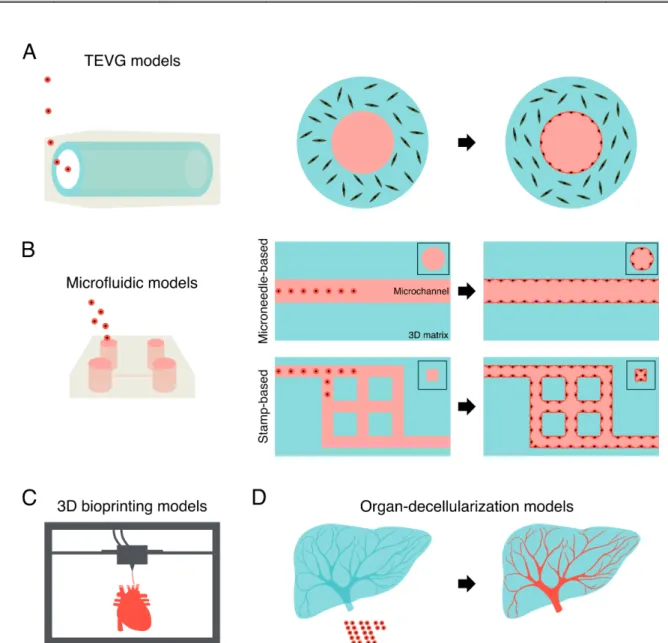
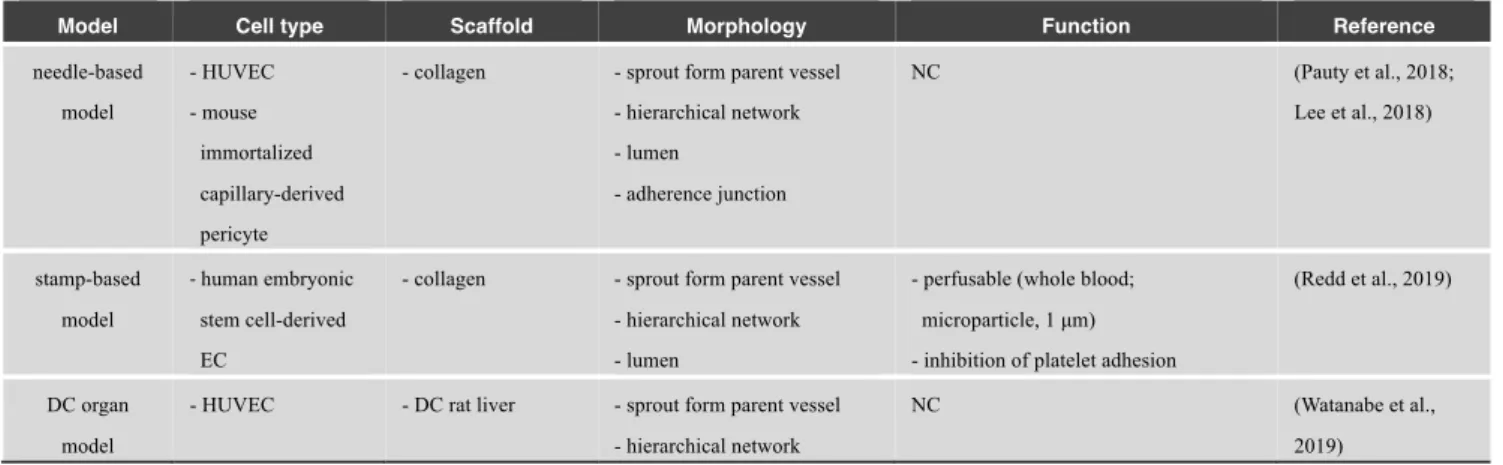
Recent studies have formed continuous vascular networks between self-organized small vessels and large vessels in pre-designed molds (Table 9). For example, several studies formed continuous vascular networks consisting of chemically-stimulated vascular sprouts via self-organization and pre-existing parent vessels formed in pre-designed microchannels created by microneedles (Pauty et al., 2018; Lee et al., 2018) (Fig. 4A). Another model seeded embryonic stem cell-derived ECs into patterned microchannels and the surrounding collagen matrix. The ECs lining the inner surface of microchannels were sprouted and anastomosed with EC tubes in the matrix via self-organization (Redd et al., 2019). Moreover, this approach constructed organ-scale hierarchical vascular networks ranging from microvessels to large vasculatures in a decellularized organ (Watanabe et al., 2019) (Fig. 4B). In their study, ECs self-organized into microvessels in response to flow-induced mechanical stress in a decellularized liver, while the endothelium of large vascular networks was formed in preserved natural ECM structures of the liver. These approaches may provide potential strategies and future directions toward the construction of hierarchical vascular networks in vitro.
Table 9 Integration of self-organization and pre-designed approaches
Model Cell type Scaffold Morphology Function Reference
needle-based model - HUVEC - mouse immortalized capillary-derived pericyte
- collagen - sprout form parent vessel - hierarchical network - lumen - adherence junction NC (Pauty et al., 2018; Lee et al., 2018) stamp-based model - human embryonic stem cell-derived EC
- collagen - sprout form parent vessel - hierarchical network - lumen
- perfusable (whole blood; microparticle, 1 μm) - inhibition of platelet adhesion
(Redd et al., 2019)
DC organ model
- HUVEC - DC rat liver - sprout form parent vessel - hierarchical network
NC (Watanabe et al.,
2019)
Fig. 4 Integration of self-organization and pre-designed approaches. (A, B) Formation of continuous vascular networks consisting of vascular sprouts via self-organization (arrowheads) and pre-existing parent vessels formed in pre-designed microchannels in microfluidic and organ-decellularization models.
5. Future perspectives
Various types of in vitro models have been developed to construct physiological vascular networks ranging from microscale capillaries/microvessels to organ-scale vascular networks. Although both self-organization and pre-designed approaches have provided promising platforms for constructing vascular networks, several crucial problems remain for medical and clinical applications, including drug development and organ transplantation, because each model cannot recapitulate all features of physiological vascular networks.
First, to obtain functional tissues and organs in vitro, hierarchical and continuous vascular networks of the artery, capillary, and vein, are required for mass transport to the entire tissue. In addition, a multilayer structure of blood vessels is inevitable for vascular strength (Pashneh-tala et al., 2016). The vascular functions, such as barrier and antithrombogenic function, should be recapitulated for maintenance of tissue homeostasis. Thus, an ideal model mimicking physiological vascular networks is required to construct functional tissues and organs. Second, while this review focuses on the construction of vascular networks, a multicellular system, including parenchymal cells (e.g., hepatocytes), should be developed to obtain organ-specific functions. The interaction of vascular cells and parenchymal cells should be considered. Finally, the cell source is also important for the development of personalized medical applications, which allow patients to receive drugs specific to their individual disease. Recent studies have focused on the use of patient-derived iPSCs and the matrix for the construction of tissues and organs (Ingram et al., 2018; Noor et al., 2019). Therefore, stem cells (e.g., embryonic stem cells and iPSCs) are potential cell sources for personalized tissue engineering. In summary, an ideal model of vascular networks is required to recapitulate the physiological features of individual patients in terms of morphology and function. As mentioned in the previous section, the integration of self-organization and pre-designed approaches with the use of patient-derived iPSCs is a potential platform to develop an ideal vascular model in vitro.
6. Summary
This review summarized various approaches for the construction of vascular networks in vitro utilizing a wide range of cell culture platforms, such as microfluidic devices, 3D printing structures, and decellularized organs. These technologies could be useful for the construction of vascularized functional tissues and organs and contribute to the development in tissue engineering and regenerative medicine.
Acknowledgement
This work was supported by Japan Society for the Promotion of Science (19H04452, 18K19937). We would like to thank Editage (www.editage.com) for English language editing.
References
Abe, Y., Ozaki, Y., Kasuya, J., Yamamoto, K., Ando, J., Sudo, R., Ikeda, M., and Tanishita, K., Endothelial progenitor cells promote directional three-dimensional endothelial network formation by secreting vascular endothelial growth factor. PLoS ONE. Vol. 8, No. 12 (2013a), e82085.
Abe, Y., Sudo, R., Ikeda, M., and Tanishita, K., Steady and pulsatile shear stress induce different three-dimensional endothelial networks through pseudopodium formation. Journal of Biorheology. Vol. 27, (2013b), pp. 38–48. Abe, Y., Watanabe, M., Chung, S., Kamm, R.D., Tanishita, K., and Sudo, R., Balance of interstitial flow magnitude
and vascular endothelial growth factor concentration modulates three-dimensional microvascular network formation. APL Bioengineering. Vol. 3, No. 3 (2019), 036102.
Andrée, B., Ichanti, H., Kalies, S., Heisterkamp, A., Strauß, S., Vogt, P., Haverich, A., and Hilfiker, A., Formation of three-dimensional tubular endothelial cell networks under defined serum-free cell culture conditions in human collagen hydrogels. Scientific Reports. Vol. 9, No. 1 (2019), 5437.
Arakawa, C., Gunnarsson, C., Howard, C., Bernabeu, M., Phong, K., Yang, E., DeForest, C.A., Smith, J.D., and Zheng, Y., Biophysical and biomolecular interactions of malaria-infected erythrocytes in engineered human capillaries. Science Advances. Vol. 6, No. 3 (2020), aay7243.
© 2021 The Japan Society of Mechanical Engineers
[DOI: 10.1299/jbse.20-00537]
problems, and promises. Developmental Cell. Vol. 21, No. 2 (2011), pp. 193–215.
Bao, J., Shi, Y., Sun, H., Yin, X., Yang, R., Li, L., Chen, X., and Bu, H., Construction of a portal implantable functional tissue-engineered liver using perfusion-decellularized matrix and hepatocytes in rats. Cell Transplantation. Vol. 20, No. 5 (2011), pp. 753–766.
Baptista, P.M., Siddiqui, M.M., Lozier, G., Rodriguez, S.R., Atala, A., and Soker, S., The use of whole organ decellularization for the generation of a vascularized liver organoid. Hepatology. Vol. 53, No. 2 (2011), pp. 604– 617.
Barrile, R., van der Meer, A.D., Park, H., Fraser, J.P., Simic, D., Teng, F., Conegliano, D., Nguyen, J., Jain, A., Zhou, M., Karalis, K., Ingber, D.E., Hamilton, G.A., and Otieno, M.A., Organ-on-chip recapitulates thrombosis induced by an anti-CD154 monoclonal antibody: Translational potential of advanced microengineered systems. Clinical Pharmacology and Therapeutics. Vol. 104, No. 6 (2018), pp. 1240–1248.
Bayless, K.J., and Davis, G.E., Sphingosine-1-phosphate markedly induces matrix metalloproteinase and integrin-dependent human endothelial cell invasion and lumen formation in three-dimensional collagen and fibrin matrices. Biochemical and Biophysical Research Communications. Vol. 312, No. 4 (2003), pp. 903–913.
Bogorad, M.I., DeStefano, J., Karlsson, J., Wong, A.D., Gerecht, S., and Searson, P.C., Review: In vitro microvessel models. Lab on a Chip. Vol. 15, No. 22 (2015), pp. 4242–4255.
Carrion, B., Huang, C.P., Ghajar, C.M., Kachgal, S., Kniazeva, E., Jeon, N.L., and Putnam, A.J., Recreating the perivascular niche ex vivo using a microfluidic approach. Biotechnol. Bioeng. Vol. 107, No. 6 (2010), pp. 1020– 1028.
Cheng, G., Liao, S., Wong, H.K., Lacorre, D.A., Di Tomaso, E., Au, P., Fukumura, D., Jain, R.K., and Munn, L.L., Engineered blood vessel networks connect to host vasculature via wrapping-and-tapping anastomosis. Blood. Vol. 118, No. 17 (2011), pp. 4740–4749.
Chrobak, K.M., Potter, D.R., and Tien, J., Formation of perfused, functional microvascular tubes in vitro. Microvascular Research. Vol. 71, No. 3 (2006), pp. 185–196.
Chung, S., Sudo, R., MacK, P.J., Wan, C.R., Vickerman, V., and Kamm, R.D., Cell migration into scaffolds under co-culture conditions in a microfluidic platform. Lab on a Chip. Vol. 9, No. 2 (2009), pp. 269–275.
Convery, N., and Gadegaard, N., 30 Years of microfluidics. Micro and Nano Engineering. Vol. 2, (2019), pp. 76–91. Czirok, A., and Little, C.D., Pattern formation during vasculogenesis. Birth Defects Research Part C - Embryo Today:
Reviews. Vol. 96, No. 2 (2012), pp. 153–162.
Damania, A., Jain, E., and Kumar, A., Advancements in in vitro hepatic models: Application for drug screening and therapeutics. Hepatology International. Vol. 8, No. 1 (2014), pp. 23–38.
Davis, G.E., Black, S.M., and Bayless, K.J., Capillary morphogenesis during human endothelial cell invasion of three-dimensional collagen matrices. In Vitro Cellular and Developmental Biology - Animal. Vol. 36, No. 8 (2000), pp. 513–519.
Davis, G.E., and Camarillo, C.W., An α2β1 integrin-dependent pinocytic mechanism involving intracellular vacuole formation and coalescence regulates capillary lumen and tube formation in three-dimensional collagen matrix. Experimental Cell Research. Vol. 224, No. 1 (1996), pp. 39–51.
DeCicco-Skinner, K.L., Henry, G.H., Cataisson, C., Tabib, T., Curtis Gwilliam, J., Watson, N.J., Bullwinkle, E.M., Falkenburg, L., O’Neill, R.C., Morin, A., and Wiest, J.S., Endothelial cell tube formation assay for the in vitro study of angiogenesis. Journal of Visualized Experiments. Vol. 10, No. 91 (2014), e51312.
van Duinen, V., Zhu, D., Ramakers, C., van Zonneveld, A.J., Vulto, P., and Hankemeier, T., Perfused 3D angiogenic sprouting in a high-throughput in vitro platform. Angiogenesis. Vol. 22, No. 1 (2019), pp. 157–165.
Folkman, J., and Haudenschild, C., Angiogenesis in vitro. Nature. Vol. 288, (1980), pp. 551–556.
Galie, P.A., Nguyen, D.-H.T., Choi, C.K., Cohen, D.M., Janmey, P.A., and Chen, C.S., Fluid shear stress threshold regulates angiogenic sprouting. Proceedings of the National Academy of Sciences of the United States of America. Vol. 111, No. 22 (2014), pp. 7968–7973.
Grigoryan, B., Paulsen, S.J., Corbett, D.C., Sazer, D.W., Fortin, C.L., Zaita, A.J., Greenfield, P.T., Calafat, N.J., Gounley, J.P., Ta, A.H., Johansson, F., Randles, A., Rosenkrantz, J.E., Louis-Rosenberg, J.D., Galie, P.A., Stevens, K.R., and Miller, J.S., Multivascular networks and functional intravascular topologies within biocompatible hydrogels. Science. Vol. 364, No. 6439 (2019), pp. 458–464.
Guyette, J.P., Gilpin, S.E., Charest, J.M., Tapias, L.F., Ren, X., and Ott, H.C., Perfusion decellularization of whole organs. Nature Protocols. Vol. 9, No. 6 (2014), pp. 1451–1468.
Hsu, Y.H., Moya, M.L., Abiri, P., Hughes, C.C.W., George, S.C., and Lee, A.P., Full range physiological mass transport control in 3D tissue cultures. Lab on a Chip. Vol. 13, No. 1 (2013), pp. 81–89.
Hussein, K.H., Park, K.-M., Kang, K.-S., and Woo, H.-M., Heparin-gelatin mixture improves vascular reconstruction efficiency and hepatic function in bioengineered livers. Acta Biomaterialia. Vol. 38, (2016), pp. 82–93.
Hussein, K.H., Park, K.M., Yu, L., Song, S.H., Woo, H.M., and Kwak, H.H., Vascular reconstruction: A major challenge in developing a functional whole solid organ graft from decellularized organs. Acta Biomaterialia. Vol.
103, (2020), pp. 68–80.
Ingram, P.N., Hind, L.E., Jiminez-Torres, J.A., Huttenlocher, A., and Beebe, D.J., An accessible organotypic microvessel model using iPSC-derived endothelium. Advanced Healthcare Materials. Vol. 7, No. 2 (2018), 1700497.
Itai, S., Tajima, H., and Onoe, H., Double-layer perfusable collagen microtube device for heterogeneous cell culture. Biofabrication. Vol. 11, No. 1 (2019), 015010.
Itoh, M., Nakayama, K., Noguchi, R., Kamohara, K., Furukawa, K., Uchihashi, K., Toda, S., Oyama, J.I., Node, K., and Morita, S., Scaffold-free tubular tissues created by a bio-3D printer undergo remodeling and endothelialization when implanted in rat aortae. PLoS ONE. Vol. 10, No. 9 (2015), e0136681.
Iwasaki, K., Kojima, K., Kodama, S., Paz, A.C., Chambers, M., Umezu, M., and Vacanti, C.A., Bioengineered three-layered robust and elastic artery using hemodynamically-equivalent pulsatile bioreactor. Circulation. Vol. 118, No. 14 Suppl (2008), pp. S52-57.
Jain, A., van der Meer, A.D., Papa, A.L., Barrile, R., Lai, A., Schlechter, B.L., Otieno, M.A., Louden, C.S., Hamilton, G.A., Michelson, A.D., Frelinger, A.L., and Ingber, D.E., Assessment of whole blood thrombosis in a microfluidic device lined by fixed human endothelium. Biomedical Microdevices. Vol. 18, No. 73 (2016), pp. 1–7.
Jain, R.K., Au, P., Tam, J., Duda, D.G., and Fukumura, D., Engineering vascularized tissue. Nature Biotechnology. Vol. 23, No. 7 (2005), pp. 821–823.
Jeon, J.S., Bersini, S., Whisler, J.A., Chen, M.B., Dubini, G., Charest, J.L., Moretti, M., and Kamm, R.D., Generation of 3D functional microvascular networks with human mesenchymal stem cells in microfluidic systems. Integrative Biology. Vol. 6, No. 6 (2014), pp. 555–563.
Jiménez-Torres, J.A., Peery, S.L., Sung, K.E., and Beebe, D.J., LumeNEXT: A practical method to pattern luminal structures in ECM gels. Advanced Healthcare Materials. Vol. 5, No. 2 (2016), pp. 198–204.
Jung, Y., Ji, H., Chen, Z., Fai Chan, H., Atchison, L., Klitzman, B., Truskey, G., and Leong, K.W., Scaffold-free, human mesenchymal stem cell-based tissue engineered blood vessels. Scientific Reports. Vol. 5, (2015), 15116. Kang, H., Bayless, K., and Kaunas, R., Fluid shear stress modulates endothelial cell invasion into three-dimensional
collagen matrices. American Journal of Physiology - Heart and Circulatory Physiology. Vol. 295, No. 5 (2008), pp. 2087–2097.
Kim, J., Chung, M., Kim, S., Jo, D.H., Kim, J.H., and Jeon, N.L., Engineering of a biomimetic pericyte-covered 3D microvascular network. PLoS ONE. Vol. 10, No. 7 (2015), e0133880.
Kim, S., Lee, H., Chung, M., and Jeon, N.L., Engineering of functional, perfusable 3D microvascular networks on a chip. Lab on a Chip. Vol. 13, (2013), pp. 1489–1500.
Ko, I.K., Peng, L., Peloso, A., Smith, C.J., Dhal, A., Deegan, D.B., Zimmerman, C., Clouse, C., Zhao, W., Shupe, T.D., Soker, S., Yoo, J.J., and Atala, A., Bioengineered transplantable porcine livers with re-endothelialized vasculature. Biomaterials. Vol. 40, (2015), pp. 72–79.
Koga, M., Sudo, R., Abe, Y., Yamamoto, K., Ando, J., Ikeda, M., and Tanishita, K., Contribution of rat endothelial progenitor cells on three-dimensional network formation in vitro. Tissue Engineering: Part A. Vol. 15, No. 9 (2009), pp. 2727–2739.
Kubota, Y., Kleinman, H.K., Martin, G.R., and Lawley, T.J., Role of laminin and basement membrane in the morphological differentiation of human endothelial cells into capillary-like structures. The Journal of Cell Biology. Vol. 107, No. 4 (1988), pp. 1589–1598.
L’heureux, N., Pâquet, S., Labbé, R., Germain, L., and Auger, F.A., A completely biological tissue-engineered human blood vessel. FASEB Journal. Vol. 12, No. 1 (1998), pp. 47–56.
Langer, R., and Vacanti, J.P., Tissue engineering. Science. Vol. 260, No. 5110 (1993), pp. 920–926.
Lauerer, M., Kaiser, K., and Nagel, E., Organ Transplantation in the Face of Donor Shortage - Ethical Implications with a Focus on Liver Allocation. Visceral Medicine. Vol. 32, No. 4 (2016), pp. 278–285.
Lee, E., Takahashi, H., Pauty, J., Kobayashi, M., Kato, K., Kabara, M., Kawabe, J.I., and Matsunaga, Y.T., A 3D in vitro pericyte-supported microvessel model: Visualisation and quantitative characterisation of multistep angiogenesis. Journal of Materials Chemistry B. Vol. 6, No. 7 (2018), pp. 1085–1094.
Lee, H., Kim, S., Chung, M., Kim, J.H., and Jeon, N.L., A bioengineered array of 3D microvessels for vascular permeability assay. Microvascular Research. Vol. 91, (2014), pp. 90–98.
Mannino, R.G., Myers, D.R., Ahn, B., Wang, Y., Margo Rollins, Gole, H., Lin, A.S., Guldberg, R.E., Giddens, D.P., Timmins, L.H., and Lam, W.A., Do-it-yourself in vitro vasculature that recapitulates in vivo geometries for investigating endothelial-blood cell interactions. Scientific Reports. Vol. 5, (2015), 12401.
McFadden, T.M., Duffy, G.P., Allen, A.B., Stevens, H.Y., Schwarzmaier, S.M., Plesnila, N., Murphy, J.M., Barry, F.P., Guldberg, R.E., and O’Brien, F.J., The delayed addition of human mesenchymal stem cells to pre-formed endothelial cell networks results in functional vascularization of a collagen-glycosaminoglycan scaffold in vivo. Acta Biomaterialia. Vol. 9, No. 12 (2013), pp. 9303–9316.
© 2021 The Japan Society of Mechanical Engineers [DOI: 10.1299/jbse.20-00537]
Miller, J.S., Stevens, K.R., Yang, M.T., Baker, B.M., Nguyen, D.H.T., Cohen, D.M., Toro, E., Chen, A.A., Galie, P.A., Yu, X., Chaturvedi, R., Bhatia, S.N., and Chen, C.S., Rapid casting of patterned vascular networks for perfusable engineered three-dimensional tissues. Nature Materials. Vol. 11, No. 9 (2012), pp. 768–774.
Moya, M.L., Hsu, Y.H., Lee, A.P., Christopher, C.W.H., and George, S.C., In vitro perfused human capillary networks. Tissue Engineering: Part C. Vol. 19, No. 9 (2013), pp. 730–737.
Newman, A.C., Nakatsu, M.N., Chou, W., Gershon, P.D., and Hughes, C.C.W., The requirement for fibroblasts in angiogenesis: Fibroblast-derived matrix proteins are essential for endothelial cell lumen formation. Molecular Biology of the Cell. Vol. 22, No. 20 (2011), pp. 3791–3800.
Nicosia, R.F., Bonanno, E., and Smith, M., Fibronectin promotes the elongation of microvessels during angiogenesis in vitro. Journal of Cellular Physiology. Vol. 154, No. 3 (1993), pp. 654–661.
Nicosia, R.F., Tchao, R., and Leighton, J., Histotypic angiogenesis in vitro: Light microscopic, ultrastructural, and radioautographic studies. In Vitro. Vol. 18, No. 6 (1982), pp. 538–549.
No, D.Y., Lee, S.A., Choi, Y.Y., Park, D.Y., Jang, J.Y., Kim, D.S., Lee, S.H., and Johnson, R., Functional 3D human primary hepatocyte spheroids made by co-culturing hepatocytes from partial hepatectomy specimens and human adipose-derived stem cells. PLoS ONE. Vol. 7, No. 12 (2012), e50723.
Noor, N., Shapira, A., Edri, R., Gal, I., Wertheim, L., and Dvir, T., 3D Printing of Personalized Thick and Perfusable Cardiac Patches and Hearts. Advanced Science. Vol. 6, No. 11 (2019), 1900344.
Park, Y.K., Tu, T.-Y., Lim, S.H., Clement, I.J.M., Yang, S.Y., and Kamm, R.D., In vitro microvessel growth and remodeling within a three-dimensional microfluidic environment. Cellular and Molecular Bioengineering. Vol. 7, No. 1 (2014), pp. 15–25.
Pashneh-tala, S., Macneil, S., and Claeyssens, F., The tissue-engineered vascular graft-past, present, and future. Tissue Engineering: Part B. Vol. 22, No. 1 (2016), pp. 68–100.
Pauty, J., Usuba, R., Cheng, I.G., Hespel, L., Takahashi, H., Kato, K., Kobayashi, M., Nakajima, H., Lee, E., Yger, F., Soncin, F., and Matsunaga, Y.T., A vascular endothelial growth factor-dependent sprouting angiogenesis assay based on an in vitro human blood vessel model for the study of anti-angiogenic drugs. EBioMedicine. Vol. 27, (2018), pp. 225–236.
Polacheck, W.J., Kutys, M.L., Tefft, J.B., and Chen, C.S., Microfabricated blood vessels for modeling the vascular transport barrier. Nature Protocols. Vol. 14, No. 5 (2019), pp. 1425–1454.
Price, G.M., Wong, K.H.K., Truslow, J.G., Leung, A.D., Acharya, C., and Tien, J., Effect of mechanical factors on the function of engineered human blood microvessels in microfluidic collagen gels. Biomaterials. Vol. 31, No. 24 (2010), pp. 6182–6189.
Pries, A.R., and Secomb, T.W., Making microvascular networks work: Angiogenesis, remodeling, and pruning. Physiology. Vol. 29, No. 6 (2014), pp. 446–455.
Rao, R.R., Peterson, A.W., Ceccarelli, J., Putnam, A.J., and Stegemann, J.P., Matrix composition regulates three-dimensional network formation by endothelial cells and mesenchymal stem cells in collagen/fibrin materials. Angiogenesis. Vol. 15, (2012), pp. 253–264.
Redd, M.A., Zeinstra, N., Qin, W., Wei, W., Martinson, A., Wang, Y., Wang, R.K., Murry, C.E., and Zheng, Y., Patterned human microvascular grafts enable rapid vascularization and increase perfusion in infarcted rat hearts. Nature Communications. Vol. 10, No. 1 (2019), 584.
Ribatti, D., Nico, B., and Crivellato, E., The role of pericytes in angiogenesis. International Journal of Developmental Biology. Vol. 55, No. 3 (2011), pp. 261–268.
Sadr, N., Zhu, M., Osaki, T., Kakegawa, T., Yang, Y., Moretti, M., Fukuda, J., and Khademhosseini, A., SAM-based cell transfer to photopatterned hydrogels for microengineering vascular-like structures. Biomaterials. Vol. 32, No. 30 (2011), pp. 7479–7490.
Sano, H., Watanabe, M., Yamashita, T., Tanishita, K., and Sudo, R., Control of vessel diameters mediated by flow-induced outward vascular remodeling in vitro. Biofabrication. Vol. 12, No. 4 (2020), 045008.
Saunders, W.B., Bohnsack, B.L., Faske, J.B., Anthis, N.J., Bayless, K.J., Hirschi, K.K., and Davis, G.E., Coregulation of vascular tube stabilization by endothelial cell TIMP-2 and pericyte TIMP-3. Journal of Cell Biology. Vol. 175, No. 1 (2006), pp. 179–191.
Schor, A.M., Schor, S.L., and Allen, T.D., Effects of culture conditions on the proliferation, morphology and migration of bovine aortic endothelial cells. Journal of Cell Science. Vol. 62, (1983), pp. 267–285.
Sekine, H., Shimizu, T., Sakaguchi, K., Dobashi, I., Wada, M., Yamato, M., Kobayashi, E., Umezu, M., and Okano, T., In vitro fabrication of functional three-dimensional tissues with perfusable blood vessels. Nature Communications. Vol. 4, (2013), 1399.
Shimizu, A., Goh, W.H., Itai, S., Hashimoto, M., Miura, S., and Onoe, H., ECM-based microchannel for culturing in vitro vascular tissues with simultaneous perfusion and stretch. Lab on a Chip. Vol. 20, No. 11 (2020), pp. 1917– 1927.