Effect of a Macrofilaricidal Agent on the Bioenergetics of Acanthocheilonema viteae as Studied by 31P-NMR and
Biochemical Analysis
Amita SHUKLA-DAVE1, Raja ROY2, V. M. L. SRIVASTAVA3, A. P. BHADURI2, and R. K. CHATTERJEE1+
^Divisions of Parasitologツ, ℃central Drug Research Institute, Lucknow, INDIA 2Dtvisions of Medicinal ℃hemistクツ, Central Drug Research Institute, Lucknow,
INDIA
Divisions of Biochemistry, Central Drug Research Institute, Lucknow, INDIA
Abstract: 31P-NMR has been applied to the study on the energy metabolism of intact ro- dent filariids Acanthocheilonema viteae. Based on chemical shifts and analysis of worm ex- tracts, the phosphorus components included sugar phosphates, inorganic phosphate, glycerophosphoryl choline (GPC) and -ethanolamine (GPE), phosphoenol pyruvate (PEP), nucleotide mono, -di and -tri phosphates, nicotinamide adenine dinucleotide and uridine diphosphate glucose. Effect of adulticidal candidate drug (C. D. R. I. Comp. 82/437, in its clinical phase I trial) on the bioenergetics of A. viteae adult filariids was assessed by 31P- NMRand revalidated by metabolic and enzymatic studies. Comp. 82/437 at the active dose of 50mg/kg, orally•~5 days, showed maximumeffect on day-16 post treatment. 31P-NMR data revealed significantly low amount of GPE (52.2%), GPC (43.5%), ATP (54.8%) and PEP (77.2%) in the treated worms. Biochemically also, ATP and PEP levels in these worms were found to be reduced by 30.9 and 44% respectively. Amongst enzymes the activity of hexokinase rose by 58%. By this enhancement, the enzyme may be able to effectively mediate the entry of extra glucose (48%), into glycolysis. On the other hand, a substantial (30%) decrease in activity seems to make phosphofructokinase a real rate limiting step in the glycolysis. This would ultimately lead to the lower production of ATP. In the energy deprived worm all the metabolic activities will gradually decline and may result in the penultimate death due to drug action. NMR observations and conventional biochemical methods substantiate the findings of one another and direct towards the hitting of bioener- getic machinery of A. viteae by macrofilaricidal agent (Comp. 82/437).
Key words: 31P‑NMR, Acanthocheilonema viteae, Compound 82/437, PEP, ATP, glycolysis.
Received for publi℃ati〇n, June 16, 1997
℃. D. R. I. ℃ommun. No. 5540.
‑Author for ℃orresp〇nden℃e. Telephone No. 91‑〇522‑212411‑18 PABX extn. 4315, 4259, FAX No. 91‑〇522‑223405
INTR〇DU℃TI〇N
Lymphati℃ filariasis is a major ve℃t〇r‑borne parasitic disease of the tr〇pi℃al and sub‑
tropical Countries, affe℃tinμ 12〇 million worldwide (Ottesen and Rama℃handran, 1995)・ The presently available drug, diethyl℃arbamazine (DE℃) is insuffi℃lent because of its inadequate
effect on adult parasites (Kumaraswami et al, 1988; Fan, 1992). The disease remains un‑
℃onquered largely due t〇 non‑availability 〇f a drug whi℃九 ℃an kill the adult filarnds・ There is thus need for generation 〇f better ℃hemi℃al lead for development 〇f an effective ma℃rofilari‑
℃ide・ It is now evident that Nuclear Maμnetti℃ Resonance (NMR) spe℃tr〇s℃opy is an exqui‑
sitely powerful t〇〇1 for design and development 〇f new drugs (℃raik, 1996). The technique is used in visualizing and assessinμ the drug‑parasite interactions, thereby identifymμ vulnerable targets of filarial parasites for chemotherapeutic attack. In our earlier study (Shukla et al}
1995) we explored the energy metabolism 〇f rodent filariids. Acanthocheilonema viteae by 31 p‑NMR spe℃tros℃opy. The bi〇enerμeti℃ status of these filariids identified high energy pnos‑
phate m〇1e℃山e ph〇sph〇en〇1 pyruvate (PEP) as key metab〇lite a℃tinμ as energy reservoir.
Aden〇sine triphosphate (APT) serves as primary ℃urren℃y of Cells, fuellinμ Cellular pr〇℃esses from ma℃r〇mole℃ular synthesis to signal transdu℃tion. It is well‑known that ATP pr〇du℃ti〇n is tightly ℃oupled t〇 its ℃onsumption・ The viability of these filariids depends on their energy utilization and it is probable that ℃ell death, which is related to a breakdown in membrane function andノor inhibit!〇n in ma℃rOm〇1e℃ular synthesis, is pr〇du℃ed by a l〇℃al decrease 〇f ATP below a ℃riti℃al level. This suμμests that a larμe and unfavourable pertubati〇n in the
energy pathways could be responsible for an impaired cellular function (Thompson et al,
1987). Very often, ℃hanμes in the normal fun℃ti〇ninμ of Cells have been shown to be affected due t〇 the alterations in the energy utilization pathways. A detailed understanding of Cellular
energy metabolism, is therefore vital. NMR spectroscopy is ideally suited for investigating
. ・
the Cellular energy metabolism (Gadian et al, 1979).
In the present study a benzimidazoyl derivative ℃〇mp・ 82/437 (2,2'‑di℃arb〇metnoxyl‑
amino 5.5'・dibenzimidaz〇lyl ket〇ne) which was earlier shown to be effective against adult
filarial parasites (Abuzar et al, 1986; Fatma et al, 1989) has been used to understand the
drug‑parasite intera℃ti〇ns and the vulnerable targets of parasites'enerμy metabolism. The results 〇f NMR study was also substantiated by parallel Conventional bio℃hemi℃al estimati〇nS・
MATERIALS AND METHODS
Host‑Parasite Model: The studies were Carried out in experimental filarial infe℃ti〇n A.
viteae in rodent host Mastomys coucha. Mastomys were infected by injecting subcutaneously
.50 infective larvae of A. viteae obtained from the infected vector Ornithodorus moubata (worms et al, 1961; Singh et al, 1989). On day 75, the animals showing progressive rise in
mi℃r〇filariae (mf) were sele℃ted・ These animals were sa℃rifi℃ed and intact, m〇tile adult A.
viieae parasites were recovered from the subcutaneous tissue for ex vivo studies.
candidate drug used: C. D. R. I. Comp. 82/437, a candidate macrofilaricidal agent, was
used in the present study <Abuzar et al, 1986; Fatma et al, 1989).
℃血emi℃als and enzymes: NADP, μ1y℃oμen, horse radish peroxidase, hex〇kmase, μ1u℃〇se‑
6‑phosphate dehydr〇μenase (G6PDH), aldolase, la℃tate dehydroμenase (LDH), μ1y℃er〇ph〇sphate dehyroμenase‑tri〇se phosphate is〇merase (GDH‑TPI) and sodium salts of fru℃t〇se‑6‑phosphate (F‑6‑P), ph〇sphoenol pyruvate (PEP), fumarate and aden〇sine triph〇sphate (ATP) were obtained from Siμma ℃hemi℃als ℃o., St・ L〇uis, Missouri (U. S. A・).
NAD, NADH, ADP, sodium pyruvate, aden〇sine mon〇ph〇sphate (AMP) and oxala℃etate were pr〇℃ured from SIS℃〇 Research Laboratories, Bombay, India.
All other ℃hemi℃als and reaμents used were of high purity grade available.
Perfusion method (Thompson et al, 1987; Shukla et al, 1995)
Adult and intact worms of A. viteae (55‑197 rag) were placed in a 10 mm NMR tube. A
spa℃er was pla℃ed at the bottom 〇f the NMR tube so that the parasites reside in the sensitive reμi〇n of the NMR tube. The apparatus used for maintaining worms during NMR analysis
℃〇nsisted of a fluid reservoir filled with Kreb's Ringer saline, pH 7.4, saturated with 95%
Ⅳ2/5% ℃〇2 and an indiμenously built assembly w〇rkinμ with the phenomenon of gravity and siphon system. The saturated solution was perfused and the per fusion rate was monit〇red with sterile i. v. set. For quantitative experiments measured amount of methylene
diphosph〇ni℃ acid was diss〇Ived in the buffer medium・ The ℃hemi℃al shift 〇f the external
reference does not interfere in the region of interest. The worms were per fused in the NMR
●tube at a rate of 1.5 ml miirl.
NMR analysis (Shukla et al, 1995)
The 31P‑NMR spectra were generated at 162 MHz at 22℃ in a Bruker WM 40〇 MHz spe℃tr〇meter with a narrow bore (54 mm) and a 9・398 Tesla super℃〇ndu℃tinμ magnet. The FID s were ℃olle℃ted using inverseμated de℃ouplinμ techniques. The lock was fixed at D2〇 field initially by adjusting with D2〇 sample and then replaced with the sample. The l〇℃k amplitude was raised to the maximum and thereafter the spectra were ℃〇11e℃ted 〇n l〇℃k m〇de. Shimmmμ of the sample was Carried out using the iH FID till the line width of the
・
water signal was below l〇 Hz in the non‑spinning mode. Two thousand forty eight (2〇48) transients were acquired to obtain spectra with satisfactory signal to noise ratio for mixed
●
population. A pulse 〇f 45‑(22 us), 〇・8 s delay and de℃〇uplinμ 〇f 3 Watt was found necessary t〇 maximise signal to noise ratio 〇f all the resonan℃es under non‑saturating Conditions. In
℃ase 〇f mixed population the a℃quisiti〇n time was 45 min・ Peak assignments were made on
.
the basis of chemical shifts as reported in the literature (Gadian et al, 1979). The total am〇unt 〇f ATP in the worms was ℃al℃ulated and the level expressed per mμ tissue fresh weiμht. The relative level 〇f adenosine diph〇sphate (ADP) was estimated by substra℃tion 〇f the peak area for the βATP (peak 12) resonance from the 7ATP + βADP integral (peak 8), and ATPノADP ratio ℃al℃ulated as βATPノβADP. Initially six Controls were used as standard referen℃e.
Worm extraction
Phosphorus metabolites of A. viteae were isolated by the method of Matthews et al,
1985. Extracts were prepared by freezing nearly 81喝〇f adult worms in liquid N」 and
.
p〇wderinμ them with a mortar and pestle・ The powder was extracted with 1.5 ml of 7%
per℃hlori℃ acid and after ℃entrifuμation, the supernate was neutralised to pH 6.5 with 3〇%
K〇H. The precipitate was discarded and the final supernate was lyophilised・ 31P‑NMR spec.
tra 〇f the extract was a℃℃umulated by diss〇Ivinμ the ly〇philised extract in a solution ℃〇n.
taininμ HEPES, 5・9喝and EDTA, 〇.9喝in 〇.4ml of triple distilled water and 〇.1 ml of D20. The analysis 〇f 31P‑NMR experiment was generated at 121 MHz in a DRX 30〇 MHz
・
FT‑NMR spe℃trometer equipped with a multinu℃lear 5皿broad band probe head・ Typical experimental ℃〇nditions were as f〇11〇ws: pulse width. 9〇° relaxati〇n delay, 2s; Composite pulse de℃ μ WALTZ・16 and a℃quisiti〇n 35,0〇O were acquired t〇 obtain a satisfa℃t〇ry spe℃tra. The whole experiment was repeated five times.
Drug treatment: A. vtteae infected animals (Mastomys) were treated in vivo with Comp.
82/437 at the dose level 〇f 5〇喝/kμ orally for 5 ℃onse℃utive days in two batches. The parasites were recovered on days 8 and 16 〇f start 〇f treatment for ℃arrymμ Out 31P‑NMR stuides. The whole set 〇f experiment was repeated four times.
Biochemical revalidation was, also, carried out at the dose of 50 mg/kg, p. o. x 5 days
and the parasites were recovered 〇n day 16 〇f start of treatment. The experiment was
repeated four times.
Metaboli℃ studies: 2〇%3O喝inta℃t, motile worms isolated from ℃〇ntr〇1 and drug treated mastomys were incubated in Hank's balanced salt s〇luti〇n (pH 7・4) ℃ontaininμ 〇.5% μ1u℃ose.
Exa℃tly after 4 hrs the worms were removed and the medium assayed for μ1u℃ose (Burleiμh
et at, 1968) and lactate (Barker et alf 1941) contents. Glucose disappeared from the medium
was used as a measure of μ1u℃〇se uptake by the parasites. Similarly la℃tate ℃〇ntent 〇f the medium was a measure 〇f la℃tate excreted by the parasites.
Fifty‑Seventy喝℃〇ntrol and drug treated parasites were horn〇μenized (5% wノv) in 〇・15 M K℃1. The horn〇μenate was ℃entrifuμed at 9〇〇μ for l〇 mm and the supernatant was som℃ated at 2〇 K ℃y℃les for 4×1 min with su℃℃essive ℃〇〇1inμ intervals 〇f 1 mm each. The som℃ate was ℃entrifuμed at l〇5,〇〇Og for 3〇 mm and the supernate was used for assay 〇f
glycolytic enzymes by standard methods as described elsewhere (Mercus et al, 1973). Protein content of the pellet fraction was measured according to Lowry et al (1951) using BSA as a
standard・ ATP and PEP ℃〇ntents 〇f the worm extract were determined by spe℃trophoto‑
metri℃ method ℃oupled to G6PDH and LDH respectively (Lampre℃ht and Trauts℃h〇Id, 1963;
Sh〇nk and Boxer, 1964; K〇rnberμ, 1955),
RESULTS
In vivo 31P‑NMR spectra of live normal A. viteae
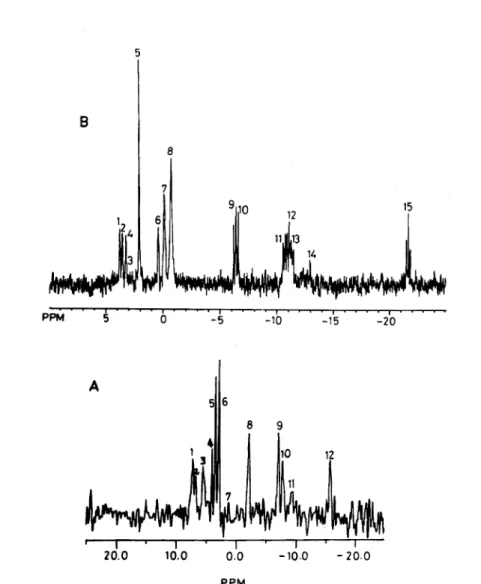
The in vivo 31P‑NMR spectrum of live and normal (untreated) A vtteae was found to be
℃omposed of 12 peaks a℃℃ordinμ to the ℃hemi℃al shift analysis (Fig. 1)・ The shifts for peaks 9 and 12 were assigned respectively to rATP + βADP, αATP and αADP, and βATP phosphorus res〇nan℃es bound t〇 Mμ. Peak l〇 ℃〇ntamed NAD + phosphorus resonance 〇f μ1u℃ose phosphate from uridine diphosphate glu℃〇se (UDPG) while that of ll belonged t〇
Fig. 1A: Typical 31P‑NMR spectra of intact live filarial parasites (A. viteae).
Fig. IB: Typical 31P‑NMR spectra of perchlorate extract of A. viteae filariids.
ph〇sphorus res〇nan℃e of uridine phosphate 〇f UDPG. Peak 7, remained unassiμned whereas
.
the signals 4, 5 and 6 were assigned to μ1y℃er〇phosphoryl ethanolamine, μ1y℃er〇ph〇sph〇ryl
.
℃holme and phosphoen〇1 pyruvate (PEP), respectively. Rest 〇f the other siμnals (1‑3) represented sugar phosphates, phosphoryl ℃h〇line and in〇rμani℃ phosphate.
31P‑NMR spectra of P℃A extract of A. viteae adults
Spe℃tra 〇f the P℃A extract (Fig. IB) based on direct ℃hemi℃al analysis indicated that resonan℃es in the d〇wnfield monophosphate region arise from primarily μ1u℃ose‑6‑P (Peak 1),
AMP (Peak 3), fructose‑6‑phosphate (peak 2), phosphocholine and fructose‑l‑6‑diphosphate
(Peak 4) and in〇rμani℃ phosphate (Peak 5)・ A res〇nan℃e of 0.55±O.〇1 ppm down field 〇f
μ1y℃er〇ph〇pSh〇ryl℃h〇line (GP℃ Peak 7) in the 31p‑NMR spectrum was assigned to
μ1y℃er〇phosph〇 ethan〇lamine (GPE, Peak 6) 〇n the basis of its ℃hemi℃al shift. The most
pr〇mment resonance in the spectrum arose from ph〇sph〇en〇1 pyruvate (Peak 8). Doublets of
^‑phosphate of ADP (peak 9) and /3‑phosphate of ADP (peak 10) as well as double doublets of
α.phosphates 〇f ATP and ADP (Peak ll and 12) were also observed m the spectra.
NAD+(H) and uridine phosphate of UDPG appeared as peak 13. Peak 14 of μ1u℃〇se ph〇s.
phate of UDPG and a triplet of /3‑phosphate of ATP (Peak 15) were also detected. The
Confirmation of the res〇nan℃e assignment 〇f PEP (Peak 8) was done by adding tris〇dium salt
〇f the authentic sample in the P℃A extract after the normal 31P‑NMR experiment・
The res〇Ived spectra 〇f the extract Confirmed the assignments of the 31P‑NMR
spectra of live, normal A. viteae filariids.
Evaluation of the effect of Candidate drug 82/437 on A. viteae/mastomys system
The 31P‑NMR spectra of the adult parasites re℃〇vered on day 8 post treatment with 82ノ437 at dose of 5〇喝ノμ (Fig 2A) exhibited no siμnifi℃ant ℃hanμe in ℃omparis〇n t〇 the spectra of normal adult parasites.
The spectra of parasites recovered 〇n day 16 of start 〇f treatment at the same dose, however, showed siμnifi℃ant effect (Fig. 2B) i. e・ 52.2% loss in GPE, 43.5% 1〇ss in GP℃ and 77.2% 1〇ss in PEP and 54・8% loss in the ATP level as well as differentiation of intra and extra℃ellular Pi was easily detectable. There was a total loss 〇f UDPG peak.
℃
,∩.n ・1O・〇 ‑2〇.O
PPH
Fig. 2A: 31P‑NMR spectra of drug (82/437) treated A. viteae 50 rag/kg, p. o. x5 days, D‑8.
Fig. 2B: 31P‑NMR spectra of drug (82/437) treated A. viteae 50 mg/kg, p. o. x5 days, D‑16.
Bio℃nemi℃al analysis revealed that ℃〇mp・ 82/437 markedly enhanced the uptake 〇f μ1u℃〇se but had no effect on la℃tate pr〇du℃ti〇n (Table 1). The treated worms were also found to possess lower amounts 〇f ATP and PEP (Table 1)・
℃〇mp. 82/437 altered activities of all the enzymes studied except that 〇f LDH (Table 2). Amongst these the activities of hex〇kinase (HK), aldolase. enolase, pyruvate kinase (PK) were enhanced whereas those 〇f ph〇sphoμ1u℃〇se is〇merase (PGI) and ph〇sphofru℃tokinase (PFK) were depressed・ Statist!℃ally the alterations induced by the ℃omp〇und in most 〇f the
・
℃ases were hiμhly siμnifi℃ant.
Table 1: Effect of Candidate drug 82/437 on μ1u℃〇se uptake and la℃tate produ℃ti〇n,
APT and PEP contents in vitro using A. viteae
℃〇‑p.para‑eterA‑℃Cone,of
。ntr。l℃〇m≡‑TreatedL℃重s重,oe二μ〇jew cttc
。l〇ith
...1...1....1...1...1.1
82/437 「3Ta℃ose uptake 0‑6〇±〇.〇8 〇・89±0.16 +48%*
t一oネZt事e pr〇du℃ 0. 97±〇・ 13 〇. 92±〇. 〇4 ‑5%
L/ 1.62 ・〇3
ATP ℃ontentb 71・ 69±〇・ 06 49, 16±O. 〇6 .3〇.9%"
PEP ℃ontentb 68. 5〇±〇. 71 38. 5〇±〇. 11 ‑44%*
[Values・expressed as aμm〇1eノmμ worm/4 hr, bpmoleノmμ protein are mean±SD of 4 experiments]
♯Pく〇・〇〇5
Table 2: Activities of carbohydrate metabolizing enzymes in A. viteae after treatment with Candidate drug 82/437
...11...1...1...1.
Enzy‑e℃S4effネit℃h℃:hang Lresp
。ntr。宮;e℃Bt
1...1.1...1...1...1
Hexokmase(HK)〇4.74±〇・43165.62±〇.51+58%a 宮3hospho:
s。merasμe17発ose
サGI)282.〇5±O・78195.53±2・12‑30%a
pho (pR溪ofru℃tokinase131・17±〇.4290・92±1.〇6.3〇%a
Aldolase::86.93±〇.28162.66±0.52+87%a En〇lase254.56±1・8〇379.〇2±1.27+48%a Lacta
hydro〇tμede‑342.23
renase(LDH)±1・45371.13±O.5〇+8%b
Pyruvatekinase(PK)97・77±〇.48229.86±1.71+135%a
[Values expressed in nmole/min/mμ protein, are mean±SE of 4 experiments]
P values: a<〇. 005, bNS (N〇t siμnifi℃ant)
DIS℃USSION
Parasiti℃ helminths Obtain their nutrients and several essential elements from their hosts, but generate enerμy 〇n their own・ Hence uptake me℃hanisms and bi〇enerμeti℃ path‑
ways have always been ℃〇nsidered loμi℃al targets for Chemotherapy (Sharma and Snvastava, 1988; Srivastava, 1995)・ Maki and Yanaμisawa (198〇a, b) 〇n the basis of dete℃tinμ high acid phosphatase activity in the cuticle of Setaria digitata have expressed the possibility of trans‑
・
℃uti℃ular absorption 〇f small molecules in the tissue dwellmμ parasites・ However uptake
. .
throuμh this route seems t〇 be limited t〇 the phosphorylated ℃〇mpounds required only m minute amounts. The supply for molecules like μ1u℃ose required in very high amounts Cannot
be effectively met by transcuticular absorption (Roy et al, 1988). Hence blocking of cuticular
abs〇rption dose not appear to yield fruitful results. From filarial Chemotherapy point 〇f view,
.
therefore, evasion of host attack and energy yielding processes seem to be of prime imp〇r.
tan℃e. Adult fiiariids utilize μ1u℃ose as the principal, if not exclusive s〇ur℃e of energy. The enerμy lS generated mostly by μ1y℃olysis which yields two m〇1e℃ules of ATP for each
・
m〇1e℃ule of μ1u℃〇se. degraded. Nonetheless minor pathways also Operate which may become effe℃tive under Certain Conditions (Barrett, 1983).
In Our earlier studies ℃omp・ 82ノ437 has been reported t〇 Cause 〇xidative damage t〇
filariids including A. viteae through selective inhibition of catalase and glutathione peroxidase
(Batra et al, 199〇, 1992a, b). In the present study, detrimental effects of this ℃〇mp〇und in vivo on the functionality of glycolysis has been shown to occur by metabolic, enzymatic and NMR studies. A℃℃〇rdinμ to NMR data, the parasites recovered on day 16 of the treatment
with 50 rag/kg dose of compound 82/437 had significantly lower amounts of GPE, GPC, ATP
and PEP (Fig. 2B). Bi〇℃hemi℃al estimation also indicated markedly reduced ATP and PEP
levels in these worms (Table 1). Surprisingly, treated A. viteae utilized about 1.5 times
μreater amounts 〇f μ1u℃ose than the untreated parasites. La℃tate production, however, remained un℃hanμed. Consequently la℃tateノμ1u℃ose (L/G) ratio dropped from 1.62 t〇 1.〇3・ At enzymati℃ level the activity of hex〇kinase (HK) rose by 58% (Table 2) and hence may ef‑
fectively mediate the entry of extra glucose (48%, Table 1) into glycolysis. Phos‑
ph〇μ1u℃〇seisomerase (PGI) is not rate limiting and therefore an observed decrease in the a℃.
tivity by 3〇 would not hamper μ1y℃〇lyti℃ rate. Nonetheless, subdued activity of ph〇s‑
℃t〇kinase (PFK) in the drug treated worms appeared t〇 be the real rate limitmμ step leading to further slowed down metabolism of μ1u℃ose and finally ℃ausmμ marked decrease in pr〇du℃ts like tri〇se phosphates, PEP and la℃tate. A℃℃ordinμly the LノG ratio and PEP level showed marked decrease in the dr噸exposed filariids (Fig. 2B, Table 1)・ Since ATP thr〇uμh μ1y℃olyti℃ pathway is formed at steps falling after PFK, the subdued fun℃ti〇ninμ of this en‑
zyme and therefore of μ1y℃〇lysis would ultimately result in lower generation 〇f ATP. This noti〇n is substantiated both bio℃hemi℃ally and by NMR analysis.
In the event of less effective μ1y℃olysis, the filariids appear t〇 derive energy by
divertinμ extra amounts of μ1u℃〇se‑6‑phosphate伽rmed from μ1u℃ose) into pentose phosphate
pathway. The NADPH generated by the action of G6PDH (Batra et al, 1990) may subse‑
quently be utilized in two ways. Firstly, for regeneration of reduced glutathione for providing protection against the drug induced oxidative damage (Batra et al, 1990; 1992a, b). Secondly, for μeneratmμ ATP through oxidation by site I 〇n.the respiratory ℃ham shown t〇 be opera‑
tive in A. viteae (Mendis and Townson, 1985). However, both G6PDH and the respiratory
℃hain operate in this filariid 〇r in general in most of the nematode parasite at extremely low rates. Consequently even this diversion of μ1u℃ose ℃ann〇t substantiate for the loss 〇f ATP due to the subdued fun℃tionmμ 〇f μ1y℃olysis・
. .
It may therefore be summarized that the adverse effect of ℃omp. 82/437 〇n PFK results in the lower production of ATP thr〇uμh μ1y℃olysis and oxidative damage (as men‑
ti〇ned earlier)・ The energy deprived worms die ultimately due to drug a℃ti〇n. Thus NMR is a valuable tool in Chemotherapy 〇f filariasis for assessmμ the effi℃a℃y of Candidate drugs in
・
pipeline as novel druμs・
A℃KNOWLED GMENTS
〇ne 〇f the authors (Amita Shukla‑Dave) is thankful to the ℃〇un℃il 〇f S℃lentm℃ and Industrial Research (℃SIR), New Delhi for financial support in the form of a fellowship.
REFEREN℃ES
1) Abuzar, S., Sharma. S.. Fatma, N・, Gupta, S., Murthy, P. K., Katiyar, J. ℃・, ℃hatterjee, R. K・ & Sen,
A. B. (1986): Studies in potential filari℃ides. 18. Synthesis of 2,2'‑disubstituted 5,5'‑dibenzimidazolyl
ketones and related compounds as potential anthelmintics. J. Med. Chem., 29, 1296‑1299.
2) Barker, B. S. & Summerson, W. H・ (1941): The ℃〇lorimetri℃ determination 〇f la℃ti℃ acid in bioloμi℃al
material. T・ Biol. ℃hem., 138, 535‑554・
3) Barrett, J. (1983);,!Biochemistry of filanal worms. Helminthol. Abstracts Series. An Animal and Hu‑
man Helmmtholoμy, 52, 1‑18.
4) Batra, S., ℃hatterjee, R. K. & Srivastava, V. M. L. (199〇): Anti〇xidant enzymes in Acanthochielonema vitea and effect of antifilarial agents. Biochem. Pharmacol., 40. 2363‑2369.
5) Batra, S., Chatterjee, R. K. & Srivastava, V. M. L. (1992a): Antioxidant system of Litomosoides cannii and Setaria cervi: Effect of a macrofilaricidal agent. Vet. Parasitol., 43, 93‑103.
6) Batra, S., Sinμh, S. P.; Fatma, N., ℃hatterjee, R. K. & Srivastava, V. M. L. (1992b): Effect of 2,2' dicarbomethoxyl amino‑5,5 ‑dibenzimidazolyl ketone on antioxidant defenses of Acanthocheilonema mteae and its laboratory host Mastomys natalensis. Biochem. PharmacoL 44. 727‑731.
7) Burleiμh, I. G. & S℃himke, R. T. (1968): On the activities 〇f some enzymes ℃〇n℃erned with μ1y℃olysis and μ1y℃oμenolysis in extracts 〇f rabbit skeletal mus℃1es・ Bio℃hem・ Biophys. Res. ℃〇mmun., 31.
831‑836.
8) Craik, D. J. (1996): The role of NMR in drug design and development, pp. 15‑29. In D. J. Craik (ed・). NMR in drug design ℃R℃ Press, Florida.
9) Fan, P. ℃. (1992): Diethyl℃arbamazine treatment of ban℃roftian and malayan filariasis with emphasis
on side effects. Ann. Trop. Med. ParasitoL, 86, 399‑4〇5.
10) Fatma, N., Sharma, S. & ℃hatterjee, R. K. (1989): 2,2'‑Di℃arbomethoxylamino, 5,5J‑dibenzimidazolyl ket〇ne: A new antifilanal agent. A℃ta Trop・, 46, 311‑321.
ll) Gadian, D. G., Radda, G. K., Ri℃hards, R・ E^ & Seeley, P. J・ (1979): 31P‑NMR in living tissue: the road from a promismμ to an important to〇1 in biology. pp. 463‑535. In R. G. Shulman (ed.). Bio‑
1oμi℃al apph℃ation 〇f maμneti℃ reS〇nan℃e. A℃ademi℃ Press, New York.
12) Kornberg, A. (1955): Lactic dehydrogenase in muscle, pp. 441. In S. P. Colowick & N. 0. Kaplan,
(ed・). Methods in Enzym〇1〇μy. A℃ademi℃ Press, New York.
13) Kumaraswami, V., Ottesen, E. A., Vijayasekaran, V., Devi, S. V., Swaminathan. M・. Aziz, M. A..
Sharma, G. R., Prabhakar, R. & Tripathi, S. P. (1988): Ivermectin for Wuchereria bancrofti filariasis:
effi℃a℃y and adverse reactions. J. Am. Med. Asso℃., 259, 315〇‑3153.
14) Lampre℃ht, W. & Trauts℃h〇Id, J. V. (1963): Determination 〇f ATP with hexokinase and μ1u℃〇se‑6・
phosphate dehydr〇μenase・ pp. 453‑551. In H. U・ Berμmeyer (ed.). Methods of Bio℃hemi℃al Analysis.
A℃ademi℃ Press. New York・
15) Lowry, 〇. H., R〇sebr〇uμh, N. JL Farr, A. L. & Randall, R. J. (1951): Protein measurement with the f〇lin phen〇1 reaμent. J. Biol. ℃hem., 193, 265‑275.
16) Maki, J・ & Yanaμisawa, T. (198〇a): Histo℃hemi℃al studies on acid phosphatase 〇f the body wall and intestine 〇f adult filarial worms in ℃ompanson with that of other parasitic nematodes. J. Helminth〇1.,
54,39‑41.
17) Maki, T. & Yanaμisawa, T・ (1980b): A Comparison of the sites 〇f acid pho叩hatase activity in an adult filaria Setaria spp・ and in some μastromtestmal nemat〇des. Parasitoloμy 81, 6〇3.6〇8.
18) Matthews, P. M., Shen, L., F〇xall, D. & Mansour, T. E. (1985): 31P‑NMR studies 〇f metab〇lite compartmentation in Fasciola hepatica. Biochim. Biophy. Acta. 845, 178‑ 188.
19) Mar℃us, R. E・ & Srivastava, V・ M. L. (1973): Effect of low oxygen tensions on μ1u℃ose‑metabolizinμ
enzymes in cultured articular chondrocytes. Proceed. Soc. Exptl. Biol. Med., 143, 488‑491.
20) Mendis. A. H. W. & Townson, S. (1985): Evidence for the occurrence of respiratory electron trans‑
.
port in adult Brugia pahangi and Dipetalonema viteae・ Mol. Bio℃h占m. parasitol・, 14, 337‑354.
21) 〇ttesen, E・ A. & Rama℃handran. ℃. P (1995): Lymphatic filariasis infection and disease: ℃〇ntrol strateμies. Parasitol. Today, ll, 129‑131.
22) Roy, T. K., Masood, K. & Srivastava, V. M. L. (1983): Transcuticular uptake of methyl glucose by Setoria cervi. Indian J. Exptl. BioL, 21, 280‑284.
23) Sharma, S. & Srivastava, V. M. L. (1988): Chemotherapy 〇f nelmmtniasis‑the molecular basis of drug a℃tion. pp. 1‑68. In A. B., Sen, J・ ℃. Katiyar & P. Y. Guru (ed.). Perspective in Parasitoloμr ℃BS Publishers & Distributors, New Delhi.
24) Shonk, ℃. E. & Boxer, G. E. (1964): Enzyme patterns in human tissues I. Methods for the deter‑
minati〇n of μ1y℃olyti℃ enzymes. ℃an℃er Research, 24, 7〇9‑721・
25) Shukla, A., Roy, R・, Bhaduri, A. P. & ℃hatterjee, R. K. (1995): A new approach in study 〇f bioenerμeti℃s of ma℃rofilariae using nuclear maμneti℃ res〇nan℃e (NMR) spe℃tros℃opy. Ind. J. Phar‑
ma℃ol., 27, 106‑110.
26) Sinμ h, D. P., Misra, S. & ℃hatterjee, R. K. (1989): Acanthochtelonema viteae: Infection in Mastomys natalensis and other rodent species. Ind. J. ParasitoL, 13 (1), 101‑110.
27) Srivastava, V. M・ L. (1995): Surface t〇poμrapny 〇f helminths in relation to drug action. pp. 165‑20〇.
In J. C. Katiyar, S. Gupta, & S. Satyavan (ed)., Approaches to Helminth Chemotherapy, CBS Pub‑
1ishers & Distributors. New Delhi.
28) Thomps〇n, S. N., Platzer, E. G. & Lee, R. W. K・ (1987): In vivo 31P‑NMR spectrum of Hymenolepis