The Journal of Physiology
https://jp.msubmit.net
JP-RP-2017-274106
Title: Physiological, synaptic and morphological properties of Phox2b-expressing neurons in the reticular formation dorsal to the trigeminal motor nucleus
Authors: Kouta Nagoya Shiro Nakamura
Keiko Ikeda Hiroshi Onimaru
Atsushi Yoshida Kiyomi Nakayama
Ayako Mochizuki Masaaki Kiyomoto
Fumihiko Sato Kiyoshi Kawakami
Koji Takahashi Tomio Inoue
Author Conflict: No competing interests declared
Author Contribution: Kouta Nagoya: Collection and assembly of data; Data analysis and interpretation; Manuscript Writing; Final approval of manuscript (required) Shiro Nakamura: Conception and design; Collection and assembly of data;
Data analysis and interpretation; Manuscript Writing; Final approval of manuscript (required) Keiko Ikeda: Provision of study materials or patients; Collection and assembly of data; Data analysis and interpretation; Final approval of manuscript (required) Hiroshi Onimaru: Provision of study materials or patients; Final approval of
manuscript (required) Atsushi Yoshida: Collection and assembly of data; Data
analysis and interpretation; Final approval of manuscript (required) Kiyomi
Nakayama: Data analysis and interpretation; Final approval of manuscript (required)
Ayako Mochizuki: Data analysis and interpretation; Final approval of manuscript (required) Masaaki Kiyomoto: Provision of study materials or patients; Final approval
of manuscript (required) Fumihiko Sato: Provision of study materials or patients;
Data analysis and interpretation; Final approval of manuscript (required) Kiyoshi Kawakami: Provision of study materials or patients; Final approval of manuscript (required) Koji Takahashi: Provision of study materials or patients; Final approval of
manuscript (required) Tomio Inoue: Conception and design; Data analysis and interpretation; Manuscript Writing; Final approval of manuscript (required)
Running Title: Phox2b neurons in the region dorsal to the trigeminal motor nucleus Dual Publication: No
Funding: MEXT-Supported Program for the Strategic Research Foundation at Private Universities 2012-2016: Tomio Inoue, No number; MEXT-Supported Program for the
Strategic Research Foundation at Private Universities 2014-2018: Tomio Inoue, No number; JSPS KAKENHI: Tomio Inoue, JP26293397; JSPS KAKENHI: Tomio Inoue,
JP15K15687; JSPS KAKENHI: Shiro Nakamura, JP16K11488
Physiological, synaptic, and morphological properties of Phox2b-expressing neurons in the reticular formation dorsal to the trigeminal motor nucleus
Kouta Nagoya1, 2, Shiro Nakamura1*, Keiko Ikeda3, Hiroshi Onimaru4, Atsushi Yoshida5, Kiyomi Nakayama1, Ayako Mochizuki1, Masaaki Kiyomoto1, Fumihiko Sato5, Kiyoshi Kawakami6, Koji Takahashi2, Tomio Inoue1
1Department of Oral Physiology, Showa University School of Dentistry, 1-5-8 Hatanodai, Shinagawa-ku, Tokyo 142-8555, Japan
2Division of Oral Rehabilitation Medicine, Department of Special Needs Dentistry, Showa University School of Dentistry, 2-2-1 Kitasenzoku, Ota-ku, Tokyo 145-8515, Japan
3Division of Biology, Hyogo College of Medicine, 1-1 Mukogawa-cho, Nishinomiya, Hyogo, 663-8501, Japan
4Department of Physiology, Showa University School of Medicine, 1-5-8 Hatanodai, Shinagawa-ku, Tokyo 142-8555, Japan
5Department of Oral Anatomy and Neurobiology, Osaka University Graduate School of Dentistry, 1-8, Yamada-Oka, Suita, Osaka 565-0871, Japan
6Division of Biology, Center for Molecular Medicine, Jichi Medical University, 3311-1 Yakushiji, Shimotsuke, Tochigi 329-0498, Japan
Running title: Phox2b neurons in the region dorsal to the trigeminal motor nucleus Keywords: Phox2b, patch clamp, jaw movement
Table of Contents category: Neuroscience–cellular/molecular
*Corresponding author:
Shiro Nakamura, D.D.S., Ph.D.
Department of Oral Physiology, Showa University School of Dentistry, 1-5-8 Hatanodai, Shinagawa-ku, Tokyo 142-8555, Japan
Phone: +81-3-3784-8160; FAX: +81-3-3784-8161 E-mail: [email protected]
Key points
Phox2b is abundantly expressed in the reticular formation dorsal to the trigeminal motor nucleus (RdV) in postnatal day 2−7 transgenic rats harbouring
fluorescently-labelled Phox2b-expressing (Phox2b+) neurons.
Almost all Phox2b+ RdV neurons are excitatory, while Phox2b-negative (Phox2b−) RdV neurons consist of a mixed population: many inhibitory and a few excitatory neurons.
A large population of Phox2b+ neurons exhibit low-frequency firing characteristics without spontaneous activity, whereas most of the Phox2b− neurons show
high-frequency firing properties and fire spontaneously.
About half of Phox2b+ and Phox2b− neurons are premotor neurons that target the trigeminal motoneurons.
The distinct neurotransmitter-phenotype and firing properties of Phox2b+ and Phox2b− neurons in the RdV may play important roles in autonomic feeding-related functions including suckling and mastication.
Abstract
Phox2b encodes a paired-like homeodomain-containing transcription factor that is essential for the development of the autonomic nervous system. Phox2b-expressing (Phox2b+) neurons are present in the reticular formation dorsal to the trigeminal motor nucleus (RdV) as well as the nucleus of the solitary tract and parafacial respiratory group. However, the nature of Phox2b+ RdV neurons is still unclear. We investigated the physiological, synaptic, and morphological properties of Phox2b+ RdV neurons using postnatal 2−7 transgenic rats expressing enhanced yellow fluorescent protein under the control of the Phox2b gene. Almost all of Phox2b+ RdV neurons were glutamatergic, whereas Phox2b-negative (Phox2b−) RdV neurons consisted of glutamatergic,
GABAergic, and glycinergic neurons. The majority (48/56) of Phox2b+ neurons showed low-frequency firing (LF), while most of the Phox2b− neurons (35/42) exhibited
high-frequency firing (HF). All, but one, Phox2b+ neurons (55/56) did not fire spontaneously, whereas about three-fourths of the Phox2b− neurons (31/42) were spontaneously active. K+ channel and persistent Na+ current blockers affected the firing characteristics of LF and HF neurons. The majority of Phox2b+ (35/46) and half of the Phox2b− neurons (19/40) did not respond to stimulations of the mesencephalic
trigeminal nucleus, the trigeminal tract, and the principal sensory trigeminal nucleus.
Biocytin labelling revealed that about half of the Phox2b+ (5/12) and Phox2b− RdV neurons (5/10) send their axons to the trigeminal motor nucleus (MoV). These results suggest that Phox2b+ RdV neurons have distinct neurotransmitter-phenotypes and firing properties from Phox2b− RdV neurons, and they might play important roles in
autonomic feeding-related functions including suckling and mastication.
Abbreviations
AHP, afterhyperpolarisation; amp, amplitude; AP, action potential; 4-AP,
4-aminopyridine; BAC, bacterial artificial chromosome; Ccell, cell capacitance; CCHS, congenital central hypoventilation syndrome; DAB, 3,3’-diaminobenzidine
tetrahydrochloride; DAPI, 4,6-diamidino-2-phenylindole; Dor, dorsal; EGFP, enhanced green fluorescent protein; ERT2, oestrogen receptor T2; Erev, reversal potential; EYFP, enhanced yellow fluorescent protein; G, conductance; GAD65, glutamic acid
decarboxylase 65; GAD67, glutamic acid decarboxylase 67; Glyt2, glycine transporter 2; Gmax, maximal conductance; HF, high-frequency firing; INaP, persistent sodium currents; 1st ISI, first inter-spike-interval; KF, Kölliker-Fuse nucleus; LF, low-frequency firing; Med, medial; MesV, mesencephalic trigeminal nucleus; MoV, trigeminal motor nucleus; P, postnatal; PB, phosphate buffer; PBN, parabrachial nucleus; pFRG,
parafacial respiratory group; Phox2b+, Phox2b-expressing; Phox2b−, Phox2b-negative;
PrV, principal sensory trigeminal nucleus; PSCs, postsynaptic currents; RdV, reticular formation dorsal to the trigeminal motor nucleus; Rec, recording electrode; Rinput, input resistance; rMoV, rostral end of the trigeminal motor nucleus; RTN, retrotrapezoid nucleus; scp, superior cerebellar peduncle; Spont., spontaneously active; Stim,
stimulating electrode; SupV, supratrigeminal nucleus; TBS, tris-buffered saline; Vglut1, vesicular glutamate transporter 1; Vglut2, vesicular glutamate transporter 2; Vrest, resting membrane potential; Vt, trigeminal tract.
Introduction
Phox2b encodes a paired-like homeodomain-containing transcription factor, which is required for neural crest differentiation and central/peripheral autonomic nervous system development (Pattyn et al., 1999; Amiel et al., 2003; Dauger et al., 2003).
Heterozygous mutations in the homeobox gene PHOX2B in the form of expansions or frameshifts of a polyalanine tract cause congenital central hypoventilation syndrome (CCHS) in humans (Amiel et al., 2003; Weese-Mayer et al., 2005). Several studies have shown that the neurons expressing Phox2b (Phox2b+ neurons) are extensively
distributed in the hindbrain regions such as the nucleus of the solitary tract, area postrema, and the dorsal motor nucleus of the vagus nerve in embryonic, neonatal, and adult rodents (Brunet & Pattyn, 2002; Dauger et al., 2003; Kang et al., 2007). These regions are related to the autonomic functions in the cardiovascular, digestive, and respiratory systems. Specifically, Phox2b is expressed in the parafacial respiratory group (pFRG), which at least partially overlaps with the retrotrapezoid nucleus (RTN) (hereinafter referred to as pFRG/RTN) (Onimaru et al., 2008), and Phox2b+ neurons aggregate around capillary blood vessels and are postsynaptically CO2-sensitive (Onimaru et al., 2012a, b).
Mastication is an orofacial movement, which is the initial and an important step in the succeeding digestive processes of food. The reticular formation dorsal to the trigeminal motor nucleus (RdV) including the supratrigeminal region described in the rat brain atlas by Paxinos and Watson (1998), is considered to play a critical role in controlling mastication. The reasons for this as follows: 1) the RdV contains premotor neurons that target jaw-closing and jaw-opening motoneurons (Travers & Norgren, 1983; Donga et al., 1990; Inoue et al., 1992; Li et al., 1995; Kolta et al., 2000;
Takamatsu et al., 2005; Yamamoto et al., 2007; Nakamura et al., 2008; Nonaka et al., 2012), 2) the RdV receives abundant inputs from various sites such as the orofacial structures (Donga et al., 1990; Westberg & Olsson, 1991; Inoue et al., 1992; Inoue et al., 1994), masticatory central pattern generator (CPG) (Nakamura & Katakura, 1995; Lund et al., 1998), cerebral cortex (Hatanaka et al., 2005; Yoshida et al., 2009), amygdala (Yasui et al., 2004), and lateral hypothalamus (Notsu et al., 2008). Kang et al. (2007) showed that, in adult rats, the Phox2b+ neurons is densely distributed in the RdV. Thus, it is possible that Phox2b+ RdV neurons may play a certain role in autonomic
feeding-related functions. However, almost nothing is known about the nature of Phox2b+ RdV neurons.
We have recently generated a bacterial artificial chromosome (BAC) transgenic rat line, which harbours a mouse BAC carrying Phox2b gene modified to drive enhanced yellow fluorescent protein (EYFP) and Cre recombinase-ERT2 (oestrogen receptor T2) (Ikeda et al., 2015). In this study, using this rat line, we examined the electrophysiological and morphological properties of Phox2b+ neurons located in the RdV in postnatal (P) 2−7 rats to elucidate the functional roles of Phox2b+ RdV neurons.
Methods
Ethical approval
All experiments and animal care were approved by the Institutional Animal Research Committee of Showa University, which operates in accordance with Japanese
Government Law No. 105. All efforts were made to minimise the number of animals used and their suffering.
Animals
All experiments were conducted using the transgenic rat line that was genetically modified to express EYFP in Phox2b+ neurons (Phox2b-EYFP rats; Ikeda et al., 2015).
Phox2b-EYFP rats expressed EYFP from a BAC transgene. Phox2b-EYFP rats have been previously described in detail, and have been successfully used in a study on pre-inspiratory neurons in the pFRG/RTN (Ikeda et al., 2015). All experiments were carried out in P2−7 rats.
Immunofluorescence
P2−5 rats were deeply anaesthetised with isoflurane until a loss of reflex was observed by tail pinch. The brainstem was rapidly isolated, fixed in 4% paraformaldehyde (WAKO, Osaka, Japan) in 0.1 M phosphate buffer (PB) at 4°C for 12 h, and washed in PBS. The tissue was immersed in 18% sucrose/PBS overnight, embedded in Tissue-Tek optimal cutting temperature (OCT) compound (Sakura Finetek, Torrance, CA, USA), frozen on dry ice, and then cut into 16−20 μm coronal cryosections.
Immunofluorescence was performed on these cryosections. Guinea pig anti-Phox2b [dilution, 1:2000 (Ikeda et al., 2015)] and chick anti-enhanced green fluorescent protein (EGFP) (Abcam, dilution 1:2000) were used for the primary antibody in
immunofluorescence. The secondary antibodies for fluorescent staining (dilution, 1:2000) were Alexa Fluor 546 anti-guinea pig IgG and Alexa Fluor 488 anti-chick (Molecular Probe, Invitrogen). Sections were stained with
4,6-diamidino-2-phenylindole (DAPI, Sigma) for nuclear staining. Images of
immunofluorescent samples were captured at 4X, 10X, or 20X objective magnifications
of a fluorescence microscope (BX51, Olympus), or at 10X or 20X objective magnifications of a confocal microscope (FV1000, Olympus).
In situ hybridisation
In situ hybridisation was carried out on 16−20 μm thick cryosections from P3−5 rats using single-stranded antisense digoxigenin-UTP (Roche Diagnostics, Basel,
Switzerland)-labelled riboprobes, as described previously (Ikeda et al., 2013; Ikeda et al., 2015). We prepared 5−6 sets of alternate sections. Hybridisation was performed at 50ºC. Hybridisation signals were visualised using an anti-digoxigenin antibody conjugated to alkaline phosphatase (Roche) and 4-nitro blue tetrazolium
chloride/5-bromo-4-chloro-3-indolyl-phosphate stock solution (Roche) was used as the chromogen.
Partial cDNAs of rat glutamic acid decarboxylase 65 (GAD65, nucleotide number 5–548 of the Genebank sequence for M72422) and rat glutamic acid decarboxylase 67 (GAD67, nucleotide number 1–475 of the Genebank sequence for M76177) were obtained by RT-PCR using rat brain total RNA, subcloned into pGEM-T easy Vector (Promega, Madison, WI, USA), and confirmed by sequencing. The riboprobe template plasmids containing cDNA of rat glycine transporter 2 (Glyt2, 1,790 base pair fragment between KpnI and ApaI site), rat vesicular glutamate transporter 1 (Vglut1, nucleotide number 815-1292 of the Genebank sequence for EU253553), and rat vesicular
glutamate transporter 2 (Vglut2, nucleotide number 494-3866 of the Genebank sequence for AF271235) were kindly provided by Dr. I. Tanaka, Dr. S. Yokota, and Dr. S.
Yokota, respectively. Proteinase K (1 μg/mL) was applied for 4 min at 26°C. Signals were detected using an anti-digoxigenin antibody conjugated with alkaline phosphatase
(Roche) and NBT/BCIP (Roche) for chromogen, followed by immunofluorescence as described.
Slice preparation for whole-cell recording
Coronal brainstem slices (400 μm thick) including the trigeminal motor nucleus (MoV) and the RdV were prepared from 94 Phox2b-EYFP rats (P2−7). Animals were
anaesthetised deeply with isoflurane (WAKO, Osaka, Japan) inhalation and then rapidly decapitated after ensuring deep anaesthesia with toe and tail pinches. Each brain was quickly removed and placed in ice-cold (4ºC) carbogenated artificial cerebrospinal fluid (ACSF, composition in mM: 130 NaCl, 3 KCl, 2 CaCl2, 2 MgCl2, 1.25 NaH2PO4, 26 NaHCO3, and 10 glucose) bubbled with 95% O2/5% CO2, pH 7.4. Coronal brain stem slices were cut on a Vibratome (VT1200S, Leica Microsystems, Tokyo, Japan) and slices were allow to recover in a holding chamber containing ACSF at 34ºC for 1 h and then kept at room temperature (24−26ºC) in ACSF.
Whole-cell recording
After recovery, the slices were transferred to a recording chamber attached to the stage of an upright microscope (Eclipse FN1, Nikon, Tokyo Japan) and continuously
superfused with carbogenated ACSF at a flow rate of 1−2 mL/min. Whole-cell patch-clamp recordings were made with a Multiclamp 700B amplifier (Molecular Devices, Sunnyvale, CA, USA) from Phox2b+ neurons in the RdV that were visualised under fluorescence and infrared differential interference contrast microscopy on the microscope equipped with a 40X water immersion objective (0.8 NA, NIR Apo, Nikon).
Patch electrodes were pulled from single-filament 1.5-mm-diameter borosilicate
capillary tubing (GD-1.5, Narishige, Tokyo, Japan) with a microelectrode puller (P-97, Sutter Instrument, Novato, CA, USA). The resistance of patch electrodes was range from 2.5 to 5.0 MΩ when filled with the internal solution. For current-clamp experiments, patch electrodes were filled with an internal solution composed of (in mM): 130 K-gluconate, 10 KCl, 10 HEPES, 0.4 EGTA, 2 MgCl2, 2 Mg-ATP, and 0.3 Na2-GTP (pH 7.25, 285−300 mOsm). Biocytin (0.5%; Sigma-Aldrich, St. Louis, MO, USA) was added to the internal solution for morphological analysis. The control
external solution consisted of ACSF of the following composition (in mM): 130 NaCl, 3 KCl, 2 CaCl2, 2 MgCl2, 1.25 NaH2PO4, 26 NaHCO3, and 10 glucose. For voltage-clamp experiments that were designed to isolate the persistent sodium currents (INaP), the internal solution was composed of the following (in mM): 130 CsF, 9 NaCl, 10 HEPES, 10 EGTA, 1 MgCl2, 3 K2-ATP, and 1 Na-GTP. The external solution was contained the following (in mM): 131 NaCl, 10 HEPES, 3 KCl, 10 glucose, 2 CaCl2, 2 MgCl2, 10 tetraethylammonium (TEA)-Cl, 10 CsCl, 1 4-aminopyridine (4-AP), and 0.3 CdCl2. Sodium currents were obtained by subtraction of the currents remaining in 1.0 μM tetrodotoxin (TTX). All experiments were performed at room temperature.
Data acquisition and analysis
Membrane currents, membrane potentials, and stimulus-evoked postsynaptic currents (PSCs) evoked by electrical stimulation, were recorded with the Multiclamp 700B amplifier. The data were low-pass filtered at 10 kHz, sampled at 20 kHz (Digidata 1332A, Molecular Devices), and analysed on a personal computer using pCLAMP 10.2 (Molecular Devices), Origin 2016 (OriginLab, Northampton, MA, USA), and Microsoft Excel. In current clamp recordings, the measured values of liquid junction potentials
between the pipette-filling and bath solutions (12 mV) were subtracted from all membrane potential values. No correction analysis of liquid-junction potential for voltage-clamp recordings was performed. Series resistance compensation was set to 60−80% in the whole-cell patch configuration and monitored periodically throughout the experiments. Neurons selected for recordings had a stable resting membrane potential more negative than -50 mV. In current-clamp experiments, passive and active membrane properties were measured. Input resistance was determined from recordings of the voltage response to 300-ms hyperpolarising, 20-pA current steps. The membrane time constant was estimated by fitting a single-exponential function to the charging transient of the same recording. Membrane capacitance was calculated by dividing the time constant by the input resistance. Single action potentials were elicited by a 3-ms depolarising current injection with 100 pA increments from the resting membrane potential. The firing properties were examined by current injection of a 1-s constant depolarising pulse. Several parameters of firing frequency were measured from them:
the maximum firing frequency, first inter-spike-interval (1st ISI), and average firing frequency over the last half of an injected 1-s depolarising current (steady state).
Spontaneous activity was measured at the resting membrane potential. When spontaneous activity occurred, the membrane potential was set at around -65 mV by current injection during the experiments for the passive and active membrane properties, and firing characteristics. In voltage-clamp experiments, stimulus-evoked PSCs and INaP
were recorded. A cashew-coated stainless steel concentric bipolar electrodes (200 kΩ, outer diameter of 100 μm; USK-10; Unique Medical, Tokyo, Japan) were used for single-pulse stimulation (0.2 ms pulse duration); the electrode was attached to an Iso-flex stimulation isolation unit (AMPI, Jerusalem, Israel) that was triggered by a
Master-8 stimulus controller (AMPI). Stimulating electrodes were placed 300-700 μm from the patch electrode. The intensity (0−100 μA) of stimulating pulses was increased by graduation to obtain certain orthodromic synaptic responses. Synaptic latency was measured from the stimulus artefact onset to the point when PSCs exceeded background noise. For recording of INaP, a positive slow ramp voltage commands (from −90 to +10 mV, 1 s, in every 5 s) was used. Current density was obtained by normalising peak INaP
to membrane capacitance.
Morphological analyses of biocytin-filled neurons
In a subset of experiments, neurons were filled by passive diffusion of biocytin from patch electrodes during whole-cell recording (15−30 min) without current injection.
After recording, the patch electrodes were carefully removed from the neurons, and the slices remained in the recording chamber until biocytin was transported into axons and dendrites (about 30 min). The slices were then fixed at least for 24 h at 4ºC with 4%
paraformaldehyde (WAKO) in 0.1 M PB. All slices were then cut into 70-μm thick slices using a vibrating microtome (Pro 7, Dosaka EM, Kyoto, Japan). After thorough rinsing in 0.05 M PBS, the slices were incubated with avidin-biotin-horseradish peroxidase (HRP) complex (ABC kit; Vector, Burlingame, CA, USA) in 0.05 M Tris-buffered saline (TBS) containing 1% Triton X-100 (WAKO) for 3 h at room temperature or overnight at 4ºC. The slices were then rinsed with TBS, reacted with 0.04% 3,3’-diaminobenzidine tetrahydrochloride (DAB, Sigma-Aldrich), 0.002% H2O2
(WAKO), and 0.07% nickel ammonium sulphate (Sigma-Aldrich) in 0.05 M TBS, and mounted on gelatine-coated slides. After dehydration, the slices were counterstained with Neutral Red. The stained neurons were reconstructed with a camera lucida.
Statistics
Statistical analysis was conducted using SPSS 17.0J (SPSS Japan, Tokyo, Japan). Data are reported as mean ± SEM. Mean value comparisons of each group were performed with Student’s t-test, one-way ANOVA, and χ2 test. ANOVA was followed by Scheffe’s post-hoc multiple comparison test when appropriate. Probability values of < 0.05 were considered statistically significant.
Results
Distribution of Phox2b+ neurons in the RdV
We first confirmed the distribution pattern of Phox2b+ neurons in the brainstem region surrounding the MoV in the P2−5 Phox2b-EYFP rats (n = 5). The expression level and pattern were consistent across the five samples, and a representative example is illustrated in Figure 1. The general expression pattern of Phox2b in P2−5 rats was comparable with that in adult rats, in which the expression of endogenous Phox2b has been previously reported (Kang et al., 2007). The EGFP-positive cells, together with Phox2b immunoreactive nuclei, were widely distributed in the reticular formation surrounding the MoV (Fig. 1A). Especially, they were densely distributed in the RdV (Fig. 1B−I). This region contained a large part of the supratrigeminal nucleus (SupV), the precise location of which has been recently re-identified by Fujio et al. (2016). The region where Phox2b+ neurons were densely distributed was located rostrally
approximately 100 μm rostral to the MoV and caudally at the level of the caudal end of the MoV. The expression of Phox2b was also found in the intertrigeminal area, medial
peritrigeminal area, and the regions around adjacent to the mesencephalic trigeminal nucleus (MesV) in P2−5 Phox2b-EYFP rats. In addition, Phox2b was sparsely expressed in the interneurons within the MoV but not in motoneurons. Phox2b immunoreactivity was absent in the principal sensory trigeminal nucleus (PrV) (Fig.
1A).
Neurotransmitter phenotype of Phox2b+ and Phox2b− RdV neurons We then performed a series of in situ hybridisation experiments in the P3−5 Phox2b-EYFP rats (n = 4) to examine whether Phox2b+ RdV neurons were
predominantly excitatory or inhibitory or a mixture of both (Fig. 2). We detected mRNA expression using either a Vglut1 probe, Vglut2 probe, mixed GAD65/67 probe, or Glyt2 probe (n = 4). We found that almost all of the Phox2b+ RdV neurons contained
detectable levels of Vglut2 mRNA (Fig. 2A, B), whereas Phox2b+ RdV neurons contained neither GAD65/67 (Fig. 2C, D) nor Glyt2 mRNA (Fig. 2E, F). On the other hand, Phox2b− RdV neurons, which were intermingled with Phox2b+ neurons in the RdV, consisted of many GAD65/67- and Glyt2- and few Vglut2-expressing neurons (Fig.
2B4, D4, F4). No mRNA signal for Vglut1 was detected in Phox2b+ and Phox2b− neurons in the RdV (data not shown). These results suggest that Phox2b+ neurons presumed to be primarily glutamatergic, similar to those previously reported in the pFRG/RTN (Stornetta et al., 2006; Onimaru et al., 2008) and the nucleus of solitary tract (Kang et al., 2007). In contrast, Phox2b− neurons might consist of a mixed population of many GABAergic and glycinergic neurons and a few glutamatergic neurons.
Firing characteristics of Phox2b+ and Phox2b− RdV neurons
We previously showed that P1−6 rat premotor neurons, targeting the MoV in the
supratrigeminal regions, were distinguished into two groups according to their discharge patterns during the steady-state firing in response to 1-s depolarising current pulses from the resting membrane potential. The two groups were of neurons firing at a frequency higher (HF neurons) or lower (LF neurons) than 33 Hz (Nakamura et al., 2014). Therefore, we next examined the firing characteristics of Phox2b+ (n = 56) and Phox2b− RdV neurons (n = 42). Example traces of the firing of Phox2b+ neurons are shown in Fig. 3A and 3D. The majority of Phox2b+ neurons (85.7%, 48/56) maintained repetitive firing in response to current pulse near the rheobase (0.04 nA) throughout the current injection with virtually no spike frequency adaptation (Fig. 3A). However, further depolarisation progressively caused a decrease in the action potential amplitude and an increase in the action potential duration, resulting in an increase in
non-overshooting action potentials and a cessation of firing within the 1-s current pulse (Fig. 3A−C). The maximum frequency of the steady state did not exceed 33 Hz (Fig. 3B, C). Based on the criteria described in the aforementioned study, these neurons were categorised as LF neuron type. A small population of the Phox2b+ neurons (14.3%, 8/56) showed sustained repetitive firing discharge at an intensity of 0.04 nA, near rheobase with the firing frequency similar to that of LF neurons at 0.04 nA (Fig. 3D).
However, the firing frequency progressively increased with current intensity, and the firing frequency of the steady state was beyond 33 Hz at further depolarisation, in contrast to LF neurons (Fig. 3E, F). These neurons were classified as HF neurons. The majority of Phox2b− neurons (83.3%, 35/42) were categorised as HF neurons and the remaining Phox2b− neurons (16.7%, 7/42) categorised as LF neurons (Fig. 3G). There
was a significant difference in the proportion of neurons with LF or HF firing characteristics between Phox2b+ and Phox2b− neurons (χ2 test, P < 0.01). When constructing a histogram of the maximum firing frequency of the steady state in the Phox2b+ and Phox2b− neurons, the peak of the histogram of the Phox2b+ neurons was located to the left compared to that of the Phox2b− neurons (Fig. 3H).
We have recently reported that membrane depolarisation induced a rhythmical burst discharge in a subset of supratrigeminal premotor neurons in P1−6 rats (Nakamura et al., 2014). Therefore, we then examined whether Phox2b+ and Phox2b− RdV neurons produced rhythmical burst firing in response to sustained depolarising current injection in 33 Phox2b+ and 27 Phox2b− neurons. All except one Phox2b+ neurons (55/56) did not fire spontaneously at rest, 85% (47/55) of which were categorised as LF neurons. In 42.4% (14/33) of the Phox2b+ neurons, sparse and irregular spike discharges were produced by prolonged membrane depolarisation (Fig. 4A). About 30% (10/33) of the Phox2b+ neurons showed irregular spike discharges at a lower membrane potential (−43 mV in this example); however, their firing characteristics changed to a rhythmical burst discharge at further depolarisation (−38 mV in this example, Fig. 4B). In 27.3% (9/33) of Phox2b+ neurons, depolarisation elicited a distinct tonic burst discharge (Fig. 4C).
In contrast to the Phox2b+ neurons, the majority of the Phox2b− neurons (74%, 31/42) were spontaneously active (mean firing rate: 6.0 ± 0.8 Hz, n = 12, range: 1.8−11 Hz). The spontaneous firing pattern was significantly different between two groups (χ2 test, P < 0.01) (Fig. 4D, Table 1). Eighty-seven per cent (27/31) and 13% (4/31) of spontaneously-active Phox2b− neurons showed HF and LF firing characteristics, respectively. Most of the Phox2b− neurons (82%, 22/27) showed tonic burst discharge (Fig. 4D), and 11% (3/27) and 7% (2/27) exhibited the irregular and rhythmical burst
discharge, respectively. There was a significant difference in the proportion of the bursting patterns (tonic, irregular, rhythmic) between Phox2b+ and Phox2b− neurons (χ2 test, P < 0.01; Fig. 4E). Significantly greater number of Phox2b+ neurons showed irregular and rhythmical firing characteristics than Phox2b− neurons. Whereas,
significantly greater number of Phox2b− neurons showed tonic discharge characteristics than Phox2b+ neurons. These results indicate that the firing profiles in response to sustained current injection in Phox2b+ RdV neurons differ from those in Phox2b− RdV neurons; almost all Phox2b+ neurons are silent, while the majority of Phox2b− neurons are spontaneously active at the resting membrane potential, and more Phox2b+ neurons have the capability to generate rhythmical burst discharge than Phox2b− neurons when depolarised.
Passive and active membrane properties of Phox2b+ and Phox2b− RdV neurons We compared passive and active membrane properties of the four different neuron types i.e., Phox2b+ LF (n = 48) and HF neurons (n = 8), and Phox2b− LF (n = 7) and HF neurons (n = 35). The results are summarised in Table 1. Cell capacitance was not significantly different among the four neuron types, whereas resting membrane potential of Phox2b+ LF neurons was significantly lower than that of Phox2b− HF neurons, and the input resistances of Phox2b+ LF and HF neurons were significantly smaller than those of Phox2b− LF and HF neurons. Single action potentials were evoked by a 3-ms depolarising current injection from the resting membrane potential. The amplitude of the action potentials was not significantly different among four neuron types. However, the firing threshold of Phox2b+ LF neurons was significantly higher than that of
Phox2b− HF neurons. Both Phox2b+ and Phox2b− neurons exhibited only monophasic
afterhyperpolarisation (AHP); however, the AHP amplitude of Phox2b− HF neurons was significantly larger than that of Phox2b+ LF neurons and Phox2b− LF neurons, and the AHP half-width of Phox2b+ LF neurons was significantly shorter than that of Phox2b− LF neurons.
When comparing the membrane properties between Phox2b+ (n = 56) and Phox2b− neurons (n = 42), the cell capacitance was not significantly different between Phox2b+ and Phox2b− neurons. In contrast, Phox2b+ neurons had lower resting membrane potential and input resistance, higher firing threshold, smaller spike, and AHP amplitude, and shorter AHP half-width than Phox2b− neurons.
Ionic basis for the firing characteristics of Phox2b+ and Phox2b− RdV neurons Previous studies have shown that 4-aminopyridine (4-AP) sensitive K+ current plays a pivotal role in the rapid adaptation and spike termination in interneurons in the
supratrigeminal region and mesencephalic trigeminal neurons in response to a
depolarising current pulse (Del Negro & Chandler, 1997; Hsiao et al., 2007). Thus, to determine whether similar mechanisms are found in Phox2b+ LF RdV neurons, we next examined an effect of 4-AP, a blocker of A-type K+ currents, on the firing characteristics.
Figure 5A and B shows examples of firing of a Phox2b+ LF neuron before and after bath application of 50 μM 4-AP at injected current intensities of 0.20 and 0.30 nA. Before 4-AP application, this neuron ceased to fire within the 1-s current pulse at the intensity of 0.30 nA (Fig. 5A), whereas the firing was maintained throughout the 1-s current pulse at the same intensity during 4-AP application (Fig. 5B). The onset of reduction in the number of spikes with increasing current intensity at the steady state occurred at significantly higher current intensities after 4-AP application than before 4-AP
application (before 4-AP: 0.25 ± 0.03 nA, n = 8; after 4-AP: 0.30 ± 0.02 nA, n = 8, P <
0.05) (Fig. 5C). However, the maximum frequency of the steady-state firing after 4-AP application did not significantly differ from the maximum frequencies before 4-AP application (before 4-AP: 18.8 ± 2.3 Hz vs. after 4-AP: 22.5 ± 0.9 Hz, P = 0.20). The maximum frequencies of the steady-state firing did not exceed 33 Hz in all LF neurons tested in the presence of 4-AP. The average 1st ISI was significantly lower after 4-AP than before 4-AP at current injection of more than 0.18 nA (P < 0.05, Fig. 5C). Since the spike duration was significantly increased after 4-AP (before 4-AP: 1.86 ± 0.20 ms;
after 4-AP: 2.69 ± 0.32 ms, P < 0.05), the steady-state firing less than 33 Hz even in the presence of 4-AP may be in part due to an increase in the duration of spikes by blocking of a 4-AP sensitive K+ current during spike repolarisation. Furthermore, if LF neurons only have small INaP sensitive to riluzole, a blocker of a persistent components of Na+ current (Urbani & Belluzzi, 2000), it might also lead to the steady-state firing at a frequency < 33 Hz. This is because the riluzole-sensitive INaP has been shown to play a crucial role in sustained high-frequency firing characteristics in rat substantia gelatinosa neurons (Cho et al., 2015), the interneurons in the mouse spinal cord (Ziskind-Conhaim et al., 2008), and mouse mesencephalic trigeminal neurons (Enomoto et al., 2007).
Therefore, we next examined the properties of INaP in Phox2b+ and Phox2b− RdV neurons.
We measured the INaP in Phox2b+ and Phox2b− neurons by a slow-ramp voltage commands (see Methods). The difference between the voltage-ramp evoked currents in the absence and presence of 1 μM TTX represented the INaP (Fig. 6A). Representative traces obtained from Phox2b+ and Phox2b− neurons are shown in Figure 6B. In Phox2b+ neurons, INaP activated at around −55.6 ± 1.4 mV and reached a peak amplitude at −33.1
± 1.1 mV (n = 21), and in Phox2b− neurons, it activated at around −60.1 ± 1.6 mV and peaked at −34.8 ± 0.7 mV (n = 21). The onset of activation was significantly higher in Phox2b+ neurons than in Phox2b− neurons (P < 0.05), whereas the voltage at the peak of activation was not significantly different between Phox2b+ and Phox2b− neurons. When calculating the conductance of INaP, which were fitted to the Boltzmann function in Phox2b+ and Phox2b− neurons, the midpoint for activation (Phox2b+ neurons: −40.5 ± 2.1 mV; Phox2b− neurons: −44.5 ± 0.83 mV) and slope factor (Phox2b+ neurons: 3.83 ± 0.42; Phox2b− neurons: 3.28 ± 0.17) were not significantly different (Fig. 6C). However, the maximum current density of INaP, which were obtained by dividing the current amplitude by the cell capacitance, was significantly smaller in Phox2b+ neurons than in Phox2b− neurons (Phox2b+ neurons: 3.06 ± 0.47 pA/pF; Phox2b− neurons: 5.12 ± 0.44 pA/pF, P < 0.01) (Fig. 6D). The peak of the histogram for the current density of Phox2b+ neurons was located to the left of that of Phox2b− neurons (Fig. 6E). These results indicate that most of the Phox2b+ RdV neurons had smaller INaP compared with Phox2b− RdV neurons.
We further assessed whether the INaP contributed to the HF firing behaviour in Phox2b− HF neurons. Figure 7 shows examples of firing properties of a Phox2b− HF neuron before and during bath application of 5 μM riluzole at current intensities of 0.10 and 0.30 nA. Before riluzole application, this example neuron showed sustained HF firing in the steady state at intensities of 0.10 nA (22 Hz) and 0.30 nA (46 Hz) (Fig. 7A).
In the presence of riluzole, this neuron showed sustained firing in the steady state at the intensity of 0.10 nA (20 Hz); however, further depolarisation caused a spike cessation at the steady state at the intensity of 0.30 nA (Fig. 7B). The onset of reduction in the number of spikes with increasing current intensity at steady state appeared at
significantly lower current intensities during riluzole application than before riluzole application (before riluzole: 0.4 ± 0.04 nA, n = 8; during riluzole: 0.27 ± 0.03 nA, n = 8, P < 0.01) (Fig. 7C). Furthermore, the maximum frequency of the steady state was lower than 33 Hz for all neurons tested in the presence of riluzole (before riluzole: 48.5 ± 3.2 Hz vs. during riluzole: 27.8 ± 2.3 Hz). These results indicate that the blockade of INaP
results in the shift of the firing characteristics from HF to LF in Phox2b− HF neurons.
Taken together, these results suggest that presumed large 4-AP sensitive K+ currents and small INaP might account for the low frequency firing characteristics of Phox2b+ RdV neurons.
Inputs to Phox2b+ and Phox2b− RdV neurons
We next examined the properties of synaptic input to Phox2b+ and Phox2b− RdV neurons evoked by single-pulse electrical stimulation of the MesV, the trigeminal tract (Vt), and the dorsal parts of the PrV at a holding potential of −60 mV (Fig. 8A, B).
Although the majority of Phox2b+ neurons (76%, 35/46) and about half of the Phox2b− neurons (48%, 19/40) did not receive synaptic inputs from any of these three regions, several neurons received synaptic inputs from one (Phox2b+: n = 7; Phox2b−: n = 18) or two (Phox2b+: n = 4; Phox2b−: n = 3) of these nuclei and tract. The percentages of neurons showing PSCs in response to Vt and PrV stimulation were not significantly different between Phox2b+ and Phox2b− neurons; however, the percentage of Phox2b+ neurons that responded to MesV stimulation was significantly lower than that of the Phox2b− neurons (χ2 test, P < 0.05; Fig. 8C). The PSCs were evoked at relatively long latencies of around 5 ms (Table 2). These results suggest that the majority of Phox2b+ RdV neurons do not receive the inputs from any of these nuclei and tract, and a fewer
number of Phox2b+ neurons receive the inputs from MesV compared with Phox2b− RdV neurons in our slice experiments.
The Phox2b+ pre-inspiratory neurons in the pFRG/RTN have shown to respond to hypercapnic stimulation postsynaptically, suggesting a role in central CO2
chemoreception (Onimaru et al., 2012a). Thus, we examined whether Phox2b+ RdV neurons also respond to CO2 elevation or not. We found that no Phox2b+ neurons were depolarised in response to hypercapnia (5% to 8% CO2) in current-clamp mode in the presence of TTX (n = 10, data not shown). This result suggests that Phox2b+ RdV neurons do not seem to be CO2-sensitive.
Morphological properties of Phox2b+ and Phox2b− RdV neurons
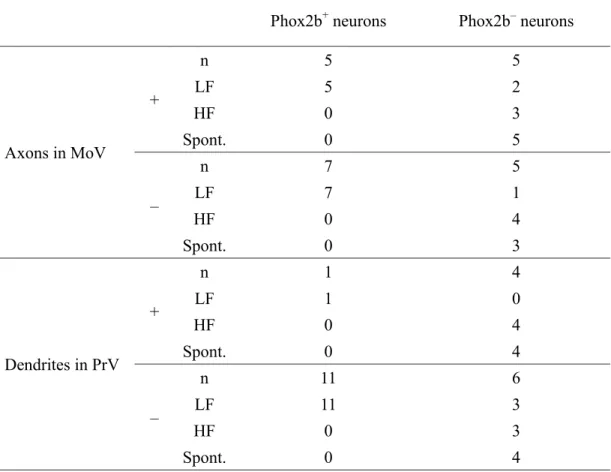
Of the 56 Phox2b+ neurons and 42 Phox2b− RdV neurons with electrophysiological properties characterised by whole-cell recordings, 12 Phox2b+ and 10 Phox2b− RdV neurons were successfully stained with biocytin. An example of reconstruction of a Phox2b+ RdV neuron is shown in Figure 9. This neuron had LF firing characteristics (Fig. 9C) without spontaneous activity and did not respond to the stimulation of MesV, Vt, and PrV. The cell body of this neuron was situated in the SupV described by Fujio et al. (2016) (Fig. 9A, B). The primary dendrites extended in various directions. A stem axon originated from a primary dendrite (Fig. 9A), ran dorsally for some distance, and then emanated two main branches. They ran ventrally to the MoV, entered it from its dorsal aspect, ramified extensively, and terminated within it. Under high magnification, the terminal arbours with boutons terminaux and en passant were identified within the MoV, especially its dorsolateral and central parts (Fig 9A, E). All Phox2b+ neurons except for one Phox2b+ neuron had relatively small and restricted dendritic trees that
were confined within the RdV in which the soma were situated. The dendrites of only one Phox2b+ neuron extended in the lateral direction and reached to the dorsal part of the PrV (Table 3). Of the Phox2b+ neurons examined, 42% (5/12) sent their axon terminals to the dorsal, central, or both parts of the MoV (Table 3). All of
biocytin-labelled Phox2b+ neurons projecting to the MoV had LF firing characteristics and non-spontaneous activity.
Figure 10 shows an example of a Phox2b− RdV neuron. The neuron was
classified as an HF firing neuron. It fired spontaneously (Fig. 10C) and did not respond to stimulation from the MesV, Vt, and PrV. The soma of this neuron was located in the SupV (Fig. 10B). The primary dendrites extended in various directions and penetrated into the dorsal part of the PrV (Fig. 10A, B). A stem axon originated from the soma (Fig.
10A) ran dorsomedially to some extent turned its direction of extension laterally, and then emanated three main branches. The first branch ran ventrolaterally, issued collaterals entering the MoV from its dorsolateral aspect, ramified extensively, and terminated within it. The second branch coursed laterally through the SupV to the PrV and issued collaterals terminating in the SupV, PrV, and the ventrolateral part of the parabrachial nucleus. The third branch coursed medially and issued a main collateral and was severed on the way. The main collateral ran ventromedially along the dorsomedial border of the MoV and sent terminals to the MoV. Under high magnification, the terminal arbours with boutons terminaux and en passant were identified within the lateral, central, and dorsomedial parts of the MoV (Fig 10A, E).
Sixty per cent of Phox2b− neurons (6 of 10 neurons) showed relatively small and restricted dendrites and soma within the RdV; however, the dendrites of the other four Phox2b− neurons extended in the lateral direction and reached to the dorsal part of the
PrV (Table 3). Half of the Phox2b− neurons (5/10) sent their axon terminals to the dorsal, central or both parts of the MoV (Table 3). The ratio of the neurons with the axon terminals in the MoV was not significantly different between Phox2b+ and Phox2b− neurons (P =1). Three of five Phox2b− neurons projecting to the MoV showed HF firing pattern with spontaneous activity, and the remaining two neurons were spontaneously-active LF neurons.
Discussion
In the present study, we demonstrated for the first time the electrophysiological and morphological properties of Phox2b+ neurons located in the RdV using the transgenic rats bearing EYFP-labelled neurons. Comparative analysis revealed that Phox2b+ RdV neurons differ from Phox2b− RdV neurons based on the neurotransmitter phenotypes and electrophysiological properties. Phox2b+ RdV neurons were predominantly glutamatergic, whereas Phox2b− RdV neurons contained multiple ensembles of many GABAergic and glycinergic and a few glutamatergic neurons. In addition, the majority of Phox2b+ RdV neurons had LF firing characteristics and no spontaneous activity, whereas most of the Phox2b− RdV neurons showed HF firing with spontaneous activity.
Furthermore, Phox2b+ RdV neurons received scarce inputs from the MesV compared with Phox2b− RdV neurons. On the other hand, about half of both neuron types provided direct axonal projections to the MoV, i.e., premotor neurons for the MoV. We propose that Phox2b+ and Phox2b− RdV neurons may play important roles in autonomic feeding-related functions including suckling and mastication.
Phox2b+ RdV neurons as excitatory premotor neurons
We have previously reported the existence of excitatory premotor neurons targeting the jaw-closing motoneurons in the supratrigeminal regions, i.e., electrical stimulation of the supratrigeminal regions in anaesthetised rats elicits masseter electromyographic responses at short latencies (Takamatsu et al., 2005). Furthermore, electrical stimulation and photolysis of cage glutamate of the supratrigeminal regions in the rat slice
preparations evoke excitatory PSCs in the masseter motoneurons in the presence of GABAA and glycine receptor antagonists (Nakamura et al., 2008; Nonaka et al., 2012).
Recently, Stanek et al. (2016) confirmed the existence of the excitatory premotor neurons in the supratrigeminal region by demonstrating that acute photostimulation of bilateral projecting neurons in the supratrigeminal regions using channelrhodopsin-2 evokes bilateral masseter activations time-locked to the stimulus pulse in awake behaving mice. In the present study, the RdV including the supratrigeminal regions contained Phox2b+ neurons that were predominantly glutamatergic, and approximately half of the Phox2b+ neurons sent their axons terminating the dorsal and central parts of the MoV, where jaw-closing motoneurons are located (Mizuno et al., 1975; Bourque &
Kolta, 2001). Thus, our present findings provide new evidence that many of the excitatory glutamatergic premotor neurons in the RdV targeting the jaw-closing motoneurons express Phox2b.
We also identified a subset of Phox2b+ RdV neurons that exhibited a rhythmical burst discharge in response to sustained depolarisation. These rhythmical bursting was more frequently observed in Phox2b+ than in Phox2b− neurons. Recent studies have reported the presence of supratrigeminal premotor neurons and PrV interneurons located adjacent to the supratrigeminal regions with similar intrinsic bursting activity (Kolta et
al., 2006; Nakamura et al 2014). It has been proposed that these supratrigeminal regions and PrV neurons may be involved in the generation of suckling and/or masticatory rhythm (Morquette et al. 2012). If this were the case, then Phox2b+ RdV neurons would play a role in suckling and/or masticatory rhythm generation and directly send
rhythmical excitatory inputs to the MoV.
Phox2b− RdV neurons as inhibitory premotor neurons
In contrast to Phox2b+ RdV neurons, in situ hybridisation revealed that Phox2b− RdV neurons were comprised a mixture of many GABAergic and glycinergic and a few glutamatergic neurons. Furthermore, the majority of Phox2b− RdV neurons were spontaneously active. Moreover, our intracellular labelling showed that half of the Phox2b− RdV neurons, all of which were spontaneous active, sent their axons terminating in the lateral, central, and dorsomedial parts of the MoV. Similar
spontaneous firing properties of the inhibitory neurons have been previously described in a large proportion of GAD67-GFP+ tonic-firing neurons in the mouse rostral solitary tract nucleus (Wang & Bradley, 2010), the nucleus of Roller (van Brederode et al., 2011), and a subset of GlyT2-EGFP neurons in the lateral cerebellar nucleus (Uusisaari
& Knopfel, 2010). Furthermore, Bourque and Kolta (2001) reported the existence of spontaneously-active interneurons in the lateral pontine reticular formation in which the supratrigeminal regions was included, although their neurotransmitter phenotypes were not identified. Chronic silencing with tetanus toxin light chain and optogenetic
inactivation of the bilaterally projecting premotor neurons in the supratrigeminal regions has shown to increase bilateral masseter activation during chewing. This suggests a role of bilaterally projecting premotor neurons in the supratrigeminal regions
in maintaining an appropriate level of masseter muscle activities by persistently inhibiting the masseter motoneurons during resting and chewing bouts (Stanek et al., 2016). Taken together, a large population of Phox2b− RdV neurons with spontaneous activity are more likely to be inhibitory premotor neurons, and their spontaneous activities are likely to provide a tonic inhibitory drive onto the masseter motoneurons under resting and chewing periods.
Roles of Phox2b+ RdV neurons in feeding-related functions
In the present study, we demonstrated that a subset of Phox2b+ RdV neurons is possible excitatory glutamatergic premotor neurons for the jaw-closing motoneurons, as
mentioned above. Furthermore, all of Phox2b+ RdV neurons projecting to the MoV showed LF firing characteristics without spontaneous activity. Similar low-frequency firing of excitatory neurons compared with inhibitory neurons has been reported for the neurons in the rat prepositus hypoglossi nucleus (Shino et al., 2008) and the neurons in the superficial and deep dorsal horn (Punnakkal et al., 2014). Mastication is a
rhythmical movement in which the jaw-closing and jaw-opening muscles are alternatively activated. The jaw-closing muscles are activated when biting food. The duration of the jaw-closing phase is quite short during mastication in rodents. Thus, the long-lasting and spontaneous activities of jaw-closing muscles might not be needed for mastication. Therefore, it is reasonable that Phox2b+ neurons with LF firing
characteristics and without spontaneous activity may be suitable for such jaw-closing behaviour.
Phox2b+ RdV neurons may be also involved in controlling the activities of the other motor-related brainstem nuclei caudal to the MoV. Neuroanatomical tracing