Title
よび溶菌物質の精製とその性状
Author(s)
吉元, 房枝; 今村, 禎祐
Citation
琉球大学保健学医学雑誌=Ryukyu University Journal of
Health Sciences and Medicine, 1(3): 185-197
Issue Date
1978
URL
http://hdl.handle.net/20.500.12001/2234
琉大保医誌1(3) : 185-197, 1978
ウェルシュ菌K29株の産生するバクテリオシンおよび
溶菌物質の精製とその性状
琉球大学保健学部微生物学教室吉 元 房 枝
は じめに バクテリオシンはある種の細菌により産生され, 同種または類縁の菌にのみ殺菌作用を示し,蛋白鷺 を主成分とする物質に対して稔称されている。ウェ ルシュ菌の産生するバクテリオシンは最初Smithil によって漆原株分離時に,感受性菌で増強されない 菌株間阻害物質としてその存在が認められた。その ご,多くの研究者により,ウェルシュ菌のバクテリ オシンが分放され,その性状についても明らかにさ れている02卜8) 最近Mahony ら9)は作用様式に関する研究にお いて,バクテリオシンの作用により感受性菌の細胞 壁が溶解され,溶菌が起こることを示唆す.るととも に, 0.3M庶糖溶液存在下において spheroplast-formationが誘発されることを報告している。また Hirano ら10)も Cl- perfringens K29株の培養上清 は殺菌と溶菌の両作用を示すことを報告している。 しかしMahony らや Hirano らの報告は粗バク テリオシンについてなされたものであり,両作用が 同一因子によって起こるのかそれとも別々の因子に よって起こるのか不明である。 そこで著者らは Hsrano らと同一株を用いて両 作用の作用様式を調べる目的でバクテリオシンの精 製を試みた。その結果,殺菌物質と溶菌物質がほぼ 分離・精製できたので,それらの産生と性状につい て比較検討を加えた。 実験材料と方法 使用菌株:バクテリオシン産生株としてCl.カer-fringens K29,感受性菌として同108株を使用した (いずれも尿由来株である)0 使用培地:バクテリオシン産生用培地としてブレ イン--トインヒュ-ジョンブイヨン(栄研,以下 BHI)培地を,バクテリオシン定量用平板培地とし今 村 禎 祐
185 てGAM寒天培地(日水)と普通寒天培地との混合 培地i : i)を使用した。まt=菌継代用培地は, GAM半流動高層培地(日水)を,保存用培地とし てクックドミート培地(日水)を使用した。またバ クテリオシン希釈培地は普通ブイヨンを使用した。 バクテリオシンの産生:bhi培地に18時間予備 培養したK29株培養液4.5m」を3,000rpm, 30分間遠 心し,沈査に等量の新鮮BHI培地を加え ioom.e のBHI培地の入った三角コルベンへ接種し, 37℃ で培養した。その後,経時的に4mEずつ取出し, 3,000rpm, 30分間遠心した。上清を取り,ミリポ アフィルター(孔径0.45JJ)で濾過し,櫨液中の発 育阻止(以下GI)活性および溶菌(以下L)活性 を測定した。 活性の測定: 1) GI活性-感受性菌Cl.カ・erfrin-ens 108株の5時間培養液をGAM混合寒天培地に 拡げ,表面を乾燥後これにあらかじめ希釈した液を 1白金耳スポットし, 37℃, 1夜嫌気培養後に透明 な阻止円を形成する貴高希釈度を求め,その逆数を 試液0.01m」あたりのGI活性 A.U.とした0 2) L活性-Hiranoら11)の方法に準じて行った。 同様の感受性菌をGAM混合寒天培地に拡げ, 1夜 嫌気培養後,形成された感受性菌の菌苔上に試液を 同様にスポットし, 37℃, 1時間静置後,透明な溶 菌円を形成する最高希釈度を求め,その逆数を試液 O.Olrn^あたりのL活憧(A.U.)とした。 バクテリオシンの分離・精製:Illに示すように, 培養上清は37℃, 18時間培養菌液を新鮮BHI培地 に接種し, 37℃, 5時間培養後, 10,000rpm, 20分 間の遠心上清をミリポアフィルタ- (孔径0.45ju) で痛過し,滅液として得た。この培養上清に硫酸ア ンモニウム(和光)を50%飽和になるように加え, 1夜冷室に放置した。生じた沈澱物をIO.OOOrpm, 30分間遠心して集め,少量の0.025M Tris-HCl緩 衝液pH7.7に溶解し,十分透析後,粗バクテリオシンとした。粗バクテリオシンはGI活性, L活性 およびホスホリパーゼC活性などを示した。つぎに 使用するまで-20℃で凍結保存した。 Sephadex G-100ゲルクロマトグラフィーによる 分画:粗バクテリオシン液10m」をあらなじめ0.025 M Tris-HCl緩衝液pH 7.7で平衡化したSepha-dex G- 100 (Pharmacia Fine Chemicas)長さ 3・7×75cmのカラムでクロマトグラフィーを行ったo フラクションは5mゼずつ採取し,それぞれについて GI活性, L活性および280m//における吸光度を測 定した。同様な分画操作を数回に分けて行い,いず れにおいても8 A.U.以上の活性部をそれぞれ集め, 50%飽和硫酸アンモニウム塩折を行い,沈澱を10,000 rpm, 40分間遠心して濃縮し, GI画分とL画分と した。それぞれ0.025M Tris-HCl緩衝液で1夜透 析を行い,次回使用するまで-20℃で凍結保存した。
Diethylaminoethyl (DEAE) Sephadex A-50 イオン交換クロマトグラフィーによる分画:Sepha-dex G-100で得た試料3m且をあらかじめ0.025 M Tris-HCl緩衝液pH7.7で平衡化したDEAE
Se-phadex A-50 (Pharmacia Fine Chemicals)イ
オン交換クロマトグラフィーにより分画した。溶出 は同緩衝液に 0.3M NaClを加え0-0.3MNaCl linear gradientで行った。得られたそれぞれの分 画についてGI活性, L活性,ホスホリパーゼC 活性および280m〟における吸光度を測定した。 8 A.U.以上のGI活性画分, L活性画分をそれぞ れ集め,透析後部分精製標晶とした。 蛋白質の定量: Lowry ら12)の方法で牛血清アル ブミンを標準物質として定量した。 部分精製標品の紫外部吸収:部分精製GI物質お よびL物質の紫外部吸収は日立分光光度計139で測 定した。また本機器は菌の濁度の測定にも使用した。 ポリアクリルアミド電気泳動:電気泳動は永井13) によって述べられた方法を使用した。泳動は冷室で 行い,ゲル濃度は7.5%,電流は管ォux8cm)あた り2mAを流した。蛋白質,脂質化合物の染色はそれ ぞれアミドブラック10B (Merck),スタンプラッ クB (Merck)を使用した。また炭水化物はPAS 染色法により検出した。 GI活性およびL活性の検 出はJetten ら14)の方法を応用して行った。 GI 活 怪はGAM混合寒天培地にあらかじめスライスし
Fig. 1. Separation and purification of bacteriocin and
ウェルシュ菌のノ{-クテリオシンと癖菌物質 た泳動ゲルディスクを載せ, 4℃, 1夜静置後感受 性菌を拡げ, 37℃, 1夜嫌気培養後,阻止円の出現 した位置より決定した。一方同平板に感受性菌を あらかじめ拡げ, 37℃, 1夜嫌気培養後生じた菌苔 上に前述の泳動ディスクを載せ, 37℃, 1夜静置後, 溶菌円の形成が観察された位置をL活性の位置とL vサ、 各種試薬の調製および実験条件:トリプシン(Di-fco, 1 :250), α-キモト1)プシン4P-L Bioch-emicals, Inc.),ペプシン(石津製薬, i : lo.ooo) による作用は以下のように調製し, 37℃, 30分間静 置して調べた。トリプシン250^g/mォ, α一キモト リプシン250tig/mlをそれぞれ0.025M Tris-HCl 緩衝液pH 8.2に,ペプシン2.5mg/m-eは0.05M酢 酸衝液pH 4.5に溶解した。クロロホルムとエチル エーテルによる影響は37℃, 30分静置後,クロロホ ルム,エチルエーテルを除去し残存活性を測定した。 クロロホルム処理細胞の調製:感受性菌108株の 37-C,18時間培養液0.2m」を新鮮BHI培地4 m居へ接' 種し, 3時間さらに培養した。培養液にクロロホル ムを最終濃度が5%になるように加え,室温に30分 間放置した。処理した細胞は, 3,000rpnl, 30分間の 遠心を3回繰返し行うことにより洗浄した。最後に 660mβにおける濁度が0.6になるように BHI を沈 査に加えて菌浮遊液とした。 実 験 結 果 バクテリオシンの産生:バクテリオシン産生の経 時変化を図2に示した。 GI活性, L活性とも培養 後1時間までは明らかでないが, 2時間後から培養 液中に現われ, 3時間で最大となる。また本結果は バクテリオシンの藤生と菌の増殖が平行しているこ とを示し,対数増殖期から定常期の初期にかけてバ クテリオシンが産生され,定常期の中・後期には産 生されないことを示している.一方濁度の低下や生 菌数の有意な減少は認められなかった。本結果に基 づき,バクテリオシンの精製材料として5時間培養 菌液を使用した。 バクテリオシンの分離・精製: Sephadex G-100 ゲル滅過により粗バクテリオシンの吸収曲線は二つ のピークにわかれた(回3 。GI活性はvoid-volu-meより少し遅れて第1ピーク(灰白色を呈し高分 子成分と思われる)の屑に溶出した0 -方L活性は GI活性とオーーバーラップして出現し,第1ピーク と第2ピーク(黄色を里し,おそらく培地中のペプ
Fig. 2. Production of the bacteriocin and the bactenolytic substance by C/. perfnngens strain K29 in BHI broth. Optical density(サ-・), viable count(○-○), inhibitory activity
(□ - □), and bactenolytic activity
(▲--▲) were assayed every 1 hour.
チド成分が主成分と思われる)との中間に溶出した。 このようにGI活性と L活性は分離し,本所見よ りこれらの活性がそれぞれ別個の物質によって生じ ていることが考えられ,著者らはGI 活性を示す物 質を発育阻止物質(以下GI物質), L活性を示す 物質を溶菌物質(以下L物質)と呼び,二つの活性 画分をそれぞれ集め,溺1に示したようにさらに精 製を行った。 図4はSephadex G-100濃縮画分液をDEAE Sephadex A-50イオン交換クロマトグラフィーに より分牡したものである。 GI活性は溶出量180-320i< NaCl潰度0.1-0.2M間で幅広く溶出したo LかL GI活性の一部は0.4M NaCl溶液で押出 したあとも DEAE Sephadex A-50の上端に吸着 されたままであった。ここで得られたGI 画分を pH 8.3用ゲルポリアクリルアミド電気泳動を行い, 純度を検討した。その結果,二本の隣接したバンド がアミドブラック10Bで染色され, GI活性もその 部位に一致したo Lかし二本のいずれかが本物質で あるのか現在まで同定できていなし!。またこの部位 はPAS染色やスタンプラックBで染色されなかっ 187
た。 L画分についても同様な方法で精製した(図5)0 ほとんどのL活性がDEAE Sephadex G-100に吸 着されないで溶出した。非吸着画分についてpH4,0 用ゲルアクリルアミド電気泳動を行い純度を検討し た。その結果,一本のバンドがアミドブラック染色 により出現し, L活性もそこに一致した。また GI 活性と同様に炭水化物や脂質化合物は検出されなか am このようにして得られたGI物質はまったくL活 性を示さず,またL物質もL活性が32A.U.以下 ではGI活性を示さなかった。さらに両物質の部分 精製標品について卵黄浮遊液に両物質をそれぞれ加 え混濁を生ずるか否かで定性的にホスホリパーゼC の検出を検討したが,検出されなかった。 これらの所見より粗バクテリオシン液中には GI 活性を示すGI物質とL活性を示すL物質とが混在 することが明らかになった。 表1, 2は種々の段階における精製の結果をまと めたものである。 部分精製バクテリオシンのpH,温度安定性およ び透析の影響:図6 aに示すようにGI物質はpH7 -8で安定であるが, pH5.0で50%, pH 9.0で約 90%が不括化し, PH 4以下およびpHIO以上で完 全に不活化した。一方L物質はGI物質とわずかに 異なり, pH5-8で安定,pH4.0および9.0で50% が不括化し, pH 3以下およ一びpH IO以上で完全に 不括化した。 16bはGI, L物質の温度安定性を示したもの である。 GI物質は45℃で75%が不括化し, 50℃で 完全に不括化した。またL物質は45℃で約25%の活 性が低下し, 50℃で完全に不括化した。この所見よ り両物質とも易熱性であることが明らかになった。 蒸留水および0.025M Tris-HCl緩衝液pH 7.2 に対して透析すると,活性は透析内液に残っており, 両物質とも非透析性であった(表3)0
Fig. 3. Gel filtration of crude bacteriocin on Sephadex G-100.
Column size : 3.8×70cm; eluting buffer : 0.025M Tris-HCl buffer, pH 7.7; flow rate: 30 ml/hr;
fraction: 5 ml ; symbols : -・ ・- absorption at 280mu, -o-o- inhibitory activity,
ウェルシュ菌のバクテリオシンと溶菌物質
Fig. 4. Chromatography of Cl. perfringens strain K29 bactenocin on DEAE Sephadex A-50.
Column size : 0.9×25cm ; gradient buffer : from 0.025M Tris-HCl buffer, pH 7.7 to 0.3M NaCl in the same buffer; flow rate : 15 ml/hr; fraction : 3 ml; symbols : -●- absorbance at 280 mu,
-0-I-○--inhibitory activity, and NaCl concentration.
Fraction number
Fig. 5. Chromatography of Cl. perfringens strain K29 bacte-riolytic substance on DEAE Sephadex A-50.
Column size : 1.5×30cm; gradient buffer : from
O.025M Tris-HCl buffer, pH 7.7 to 0.3M Nad in the same buffer; flow rate: 15 ml/hr; fraction: 3 ml; symbols : absorbance at 280 m/i, ●‥1●… bacteriolytic activity,
-- NaCl concentration.
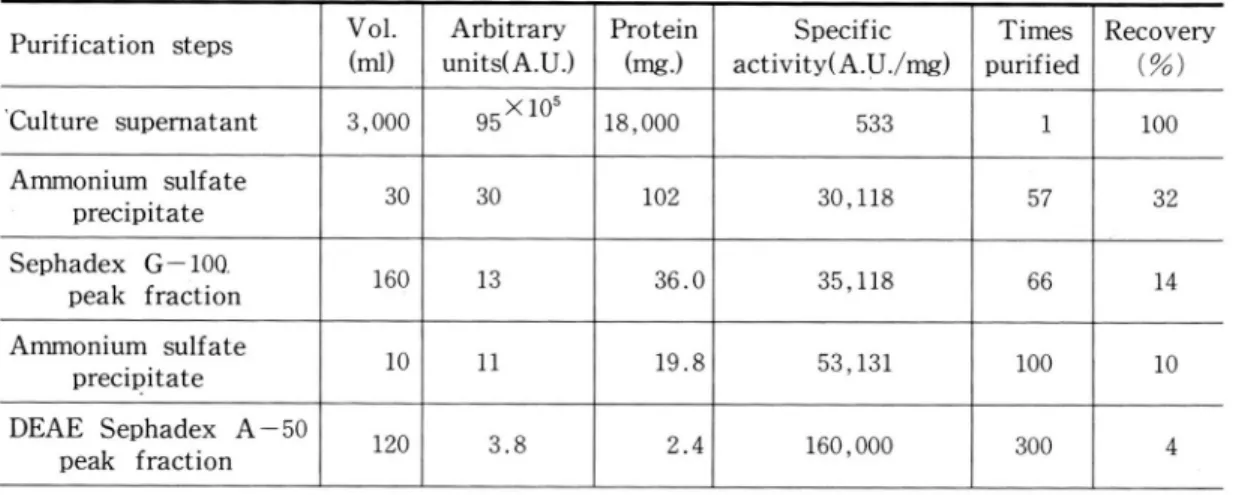
Table 1. Purification of the bacteriocin produced by Cl. perfringens strain K29
P urification step s V ol. A rb itrary Protein S pecific T im es R ecovery m l units(A .U .) (m g.) activ ityCA .U ノm g ) purified (% )
、Cu lture supern atant 3,000 95×105 18 ,000 533 1 100
A m m onium sulfate precipitate 30 30 102 30,118 57 32 Sephadex G ー100一 peak fraction 160 13 36 .0 35,118 66 14 A m m onium sulfate precipitate 10 ll 19 .8 53,131 100 10 D EA E S ephadex A ー50 peak fraction 120 3.8 2.4 160 ,000 300 4
Table 2. Purification of the bacteriolytic substance produced by Cl. ♪erfringens strain K29
P u rific ation step s V o l. A rb itra ry P ro te in S p ec ific T im es R eco v e ry m l u n its(A .U 一) (m g .) a ctiv ity (A .U ノm g ) p u rifie d (% )
C u ltu re sup e rn a tan t 14 ,0 00 44 8 ×105 23 ,100 193 1 100
A m m o n iu m su lfa te p rec ip itate 172 3 76 3 ,199 ll ,758 67 84 S e p h ad e x G - 100 pe a k fra ction 667 8 5 .4 267 3 2 ,000 165 19 A m m o n iu m su lfa te p rec ip itate 8 4 .1 10 .4 39 ,000 204 0 .9 D E A E S ep h a d ex A ー50 p e a k fra ction 30 2 3 .4 5 7 ,143 29 6 0 .2 各種試薬の影響:表3は理化学的処理の結果をま とめたものである GI, L物質とも蛋白分解酵素処 理により不活化した。また両物質は有機溶媒処理に よっても不括化した。この不活化は蛋白質の変性に よるものであった。尿素処理により両物質とも不活 化した。 バクテリオシンの紫外部吸収: GI物質はまだ精 製は十分とは言えなかったが,吸収スペクトルにつ いて検討した(図7)O吸収パターンは緩やかなカー ブを示し,最大吸収が275m/l付近に,最小吸収は 260m;uにあった。一方L物質は280m/uに最大を250 rruに最小吸収を有する典型的な蛋白質のパターン を示した(図7)0 クロロホルム処理細胞に対する両物質の影響:那 分精製したGI, L物質の感受性菌に対する作用を 検討するため,クロロホルム処理細胞に両物質をそ れぞれ作用させ,経時的に濁度の変化を観察した (図8 。GI物質を添加したものは濁度は一定で変 化が認められなかったのに対し, L物質を加えたも のは,作用後10分から濁度の低下が認められた。ま た対数増殖期および定常期の生薗細胞,加熱処理細 胞についても両物質の作用を検討したが,同様な結 果を得た。以上の所見よりL物質の溶菌活性が再確 認され,またGI物質は溶菌活性のないことが明ら かになった。 感受性菌に対するGI物質の影響:溶菌活性のな いGI物質が感受性菌に対してどのような作用を示 すかについて検討するため,対数増殖期の菌に16A.
ウェルシュ菌のバクテリオシンと溶菌物質
Fig. 6. Effect of pH(a) and temperature(b) on the bactenocin and bacteriolytic substance.
(a) : The activity was tested in following buffers; pH 2.0 : 0.05M acetate buffer adjusted with IN HCl, pH 3.0-6.0 : 0.05M acetate buffer, pH pH 3.0-6.0-8.0 : 0.05M phosphate buffer, pH 7.0-9.0 : 0.05M Tris-HCl buffer, pH 10.0-ll.0 : 0.05M Tris-HCl buffer adjusted with IN NaOH. After
dialysed for 24 hours in refrigirator against the buffers indicated, the pH was brought back to pH 7.0 with HCl or NaOH and was assayed for activities.
(b) : These samples were heated for 5 minutes at tern-peratures indicated, and chilled in ice immediately, and residual activity was assayed.
U.のGI物質を加え37℃で作用させた。その結果 を図9に示した。この図からもまたGI物質が感受 性菌の濁度に直接影響しないことが再確認されたが, GI物質により著しく生菌数が減少した。この結果は GI物質が感受性菌に対して溶菌的でなく殺菌的に 作用することを示している。 考 察 ウェルシュ菌K29株の他の菌株に対する発育阻止 (GI)活性および洛菌(L)活性は自然に培養液中 に他の毒素とともに産生された。この培養上酒場液 を硫酸アンモニウム塩析濃縮後,透析, Sephadex G-100およびDEAE Sephadex A-50カラムクロ マトグラフィーとを組合せて精製を行った。精製し たGI物質はアクリルアミド電気泳動により二本の
Symbols : -o bacteriocin,一一‥●-一一- bac-teriolytic substance. 191 隣接したバンドが混在し,一方L物質は一本のバン ドを示し,両物質とも此活性が約300倍上昇し,かな り精製されているものの収量は少なく,精製過程に ついてはさらに検討が必要である。 cl. ♪erfringens K29株の粗バクテ1)オシン中の GI活性およびL活性は本実験結果よりそれぞれ異 なった物質すなわちGI物質とL物質とによって発 現されていることが明らかになり,本論文は粗バク テリオシン活性が単一物質によって発現されている のではないことを指摘するものである。 Tagら15)はバクテリオシンの基準についてつぎ のように述べている。 1)同種または近縁種の細菌 に活性があり,活性スペクトル城が狭い, 2)生物 学的に活性のある蛋白質部分が存在する, 3)殺菌 的に作用する, 4)特異的にリセプターに付着する,
Table 3. Effect of various treatments on the activity of bacteriocin preparation and bacteriolytic preparation
T rea tm e nt C o n d itio n B a cten o c in p re p a ra tio n B a cte n o ly tic p re p a ra tio n C on tro l T rea ted C o n tro l T re ated T ry p sin 25 0ug 1 : 2 5 0 ) 37℃, 30m in . 32 (A .U .) (A .U .) 3 2 (A .U .) (A .U .) αch y m o try p sin 250 ug ′′ 32 3 2 P ep sin 2 .5m g ( 1 ‥10 ,000 ′′ 32 0 32 0 C h lo ro fo rm 50 % ′′ 3 L 32 E th y leth e r 5 0% ′′ 3 2 0 32 U re a 6 M 4 ℃, 24 h r. 32 32 0 D ia ly sis ′′ 32 32 32 32
Fig. 7. Absorption spectra of the bacte-riocin and the bacteriolytic sub-stance. Symbols :一・ ・- bacte- nocin, -0-○- bacten-olytic substance. 5)バクテリオシンの産生はプラスミドにより決定 され,産生菌は自分自身の作るバクテリオシンに対 して免疫である, 6)バクテリオシンの合成は致死 的である。しかしこれらの基準のなかで2)および3) を満足するものはバクテリオシンと考えられてきた。 今回著者らが精製したGI物質はイ a. ♪erfring-ensの他の菌株にのみ発育阻止活性があり,産生菌 の発育を阻止しない,ロ)易熱性で蛋白分解酵素処 理により不括化される, -)感受性菌に対し殺菌的 に作用する。以上の所見より著者らはGI物質をバ クテリオシンと判断した。 近年多くのバクテリオシンが精製されてきたが, ウェルシュ菌バクテリオシンの精製についてはper-fringocin lll0516'やバクテリオシンN 517>の研究 がある。これらのバクテリオシンはそれぞれ分子量 76,000, 82,000の単純蛋白質として報告されている。 著者らが今回報告するバクテリオシン(GI物質)は 完全には精製されていないが,ゲル櫨過の結果より 10万以下の分子量であることが推測され,また染色 の結果,脂質化合物や炭水化物が検出されないので 単純蛋白質の可能性を示唆するものである。そのほ か本バクテl)オシンとこれらのバクテリオシンとの 間には種々の性質の異同がみられる。すなわち1) バクテリオシンの産生様式-perfringocin 11105は 定常期以後溶菌とともに培養液中に産生され,また バクテリオシンN5も同様に産生されるが産生量が 著しく少ない。しかしUV照射により誘発され産 生量は増加する。一方本バクテリオシンは発育と平 行して培養液中に産生される。 2)温度感受性-perfrngocin l1105は耐熱性であるがバクテ))オシ ンN5および本バクテリオシンは易熱性である。 3) pH安定性-perfringocin 11105はpH 2-12で安定
ウェルシュ菌のバクテリオシンと溶菌物質
Fig. 8. Action of the bacteriocin and the bac-tenolytic substance from Cl. perfringens strain K29 on chloroform-treated sen-sitive cells Cl. perfringens strain 108 was grown m BHI broth. Cells from exponential phase were harvested By centnfugation and were exposed for 30 minutes at room temperature to BHI broth containing 5%(v/v) chloroform. The treated cells were centrifuged, washed twice with BHI broth, and resuspended in the same broth. These samples were treated with the bacteri-ocin(- - △‥) or bacteriolytic substance (--●‥), or with 0.025M Tris-HCl buffer, pH 7.7 (- ○-), and were incubated at 37 C. The OD at 660mォ.
was determined every 15 minutes. Each activity : 16 A.U. であるが本バクテリオシンはpH 7-8で安定であ り,前者に比べてpH安定城が狭い。以上のように ウェルシュ菌により産生されるバクテリオシンは単 純蛋白質という点で一致すると考えられるが,その 性状には差異があり,それぞれ異ったバクテリオシ ンと思われる。
megacin A-216 enterococcin
X-14J9'perfrin-gens 11105,バクテリオシン28およびCl. ♪・erfrin-gens K29株によって産生されるバクテリオシンは 溶菌作用を示すバクテリオシンとされている。現在 のところ溶菌作用を示すバクテリオシンの報告は少 なく, colicin M20>を除いてはいずれもグラム陽性 菌の産生するものである megacin A-216は精製 され,その本体はホスホリパーゼA2サであり,また 193 enterococcin X-14は遺伝学的研究によりhemoly-sinであることが明らかになっている。これらのバ クテリオシンの作用により,感受性菌の細胞膜は障 害を受け,その結果内容物が漏出し溶菌が起こると されている。前述のように本菌の産生するバクテリ オシンは溶菌作用がなく,これらのバクテリオシン とは異なると考えられる。また本菌の産生するL物 質は,イ) Cl. perfringensのみに溶菌活性を示し, 産生菌自身は溶菌さえない,ロ)易熱性で蛋白分解 酵素感受性, -) L物質の作用は溶菌的で生菌, 加熱,ホルマリンおよびクロロホルム処理感受性菌 を溶菌した。また0.3M庶糖を含むBHI培地に浮 遊した感受性菌108株にL物質を加えて培養すると, 感受性菌の溶菌は認められず,光学顕微鏡観察によ り, spheroplast形成が観察された。この所見はL 物質が細胞表面に作用することを示唆するものと考 えている。以上の知見よりL物質は細胞膜に作用す るmegacm A-216やenterococcin X-14などのバ クテリオシンと異なり,細胞壁を溶解するリゾチー ム様溶菌酵素の一種と考えている。 L物質はsphe-roplast形成を誘発する点でウェルシュ菌バクテリ オシン28に類似する。しかしバクテリオシン28の実 験は粗バクテリオシンを使用して行われた実験であ り,それには著者らの分離したL物質様物質が混在 するのではないかと推測される perfringocin11105 の溶菌作用の詳細については明らかにされていない。 細菌が産生する溶菌酵素については細胞外港菌酵 秦,自己融解酵素ならびにphage感染によって新 たに合成されるphage-associated lysin もしくは virolysina>および不完全phage型バクテリオシン 産生に伴って合成されるendolysin23'などが報告さ れ,基質により名称が与えられている。現在までに 本菌からはphageは分離されておらず,また電子 顕微鏡観察においてもphageやphage-tail様構 造物は観察されていない。しかし本菌がvirolysin のみを産生する defective lysogenic bacteriaで ある可能性も考えられ, virolysinを産生している のかendolysinを産生しているのか本研究からは 不明である。またウェルシュ菌の産生する溶菌酵素 については市販のホスホリパーゼC標品中に混在す るendo-N-acetyl-glucosaminidase細菌体外酵素 と芽胞およびエンテロトキシン放出に作用する N-acetyl-muramyl-L-alanine amidase25'自己融解酵 素が報告されている。本菌の産生するL物質がこれ
らの酵素である可能性もあり,本物質が細菌geno-Fig. 9. Bactericidal effect of the bacteriocin on sensitive cells. Cl. ♪erfringens strain lO8 was grown in BHI broth. Cells from the exponential phase were cen-trifuged, washed, and resuspended in fresh BHI broth. Samples of this sus-pension were treated with bacteriocin (・) or with 0.025M Tris-HCl buff-er( ○ ) and were incubated at 37。C. The OD at 660 m/∫(broken line) and the number of viable cells(solid line) were determined every 15 minutes. Bacteriocin activity : 16 A.U.
Vo : viable count at time zero, Vt : viable count at any later time t.
me由来の酵素なのかphage-genome由来の酵素 であるのか不明である。これらの問題の解明にはK 29株に感染するphageの発見 し物質の本体およ びその役割についてより深い研究を必要とする。 過去に報告されている溶菌活性を示すバクテリオ シンの殺菌活性と溶菌活性は同一物質によって発現 される。しかし著者らの報告する易熱性バクテリオ シン(GI物質)はこれまで論議してきたように, 殺菌作用のみを有し,溶菌作用は別の物質(L物質) によって起こる現象といえる。 要 約 Clostridiumヵerfringens K29株の培養上清中に 出現する発育阻止(GI)物質,ならびに溶薗(L) 物質を硫酸アンモニウム塩析浪縮後, Sephadex G-100およびDEAE Sephadex A-50カラムクロマ トグラフィーとの組合せにより分離・精製し,両物 質の理化学的性状および感受性菌に対する溶菌作用 の有無について比較研究を行い,以下の結果を得た。 1. Sephadex G-100カラムクロマトグラフィ ーにより,発育阻止を示すGI物質と溶菌活性を示 すL物質の二種の生物活性物質が分離された。さら にDEAE Sephadex A-50イオン交換クロマト グラフィーにより精製すると,両物質とも培養上清 より比活性が約300倍上昇した。 2. GI物質はpH7-8で安定, pH4以下およ びpHIO以上で不括化された。一方L物質はpH 5 -8で安定であり, pH 3以下およびpHIO以上で不 活化されたが, GI物質に比較してややpH安定城が 広い。 3・両物質とも易熱性(50℃, 5分で不括化),お よび非透析性である。 4.両物質ともクロロホルム,エチルエーテルに より不括化した。 5・蛋白分解酵素であるペプシン,トリプシン, α-キモトリプシンにより不活化された。 6. L物質は感受性菌に対して溶菌的に作用する が, GI物質には本作用がなく,殺菌的にのみ作用 する。 以上のように Cl. fierfringens K29株の培養上 清中にはGI物質とL物質が混在することが明らか になり, GI物質が従来のバクテリオシンに適合す ることが論議された。 この研究費の二部は文部省科学研究費を用いた。 本論文の要旨は第28回(1975年11月),第29回(1976 年11月)日本細菌学会九州支部捻会にお-いて発表し た。 参 考 文 献
1 ) Smith, H.W. : The bacteriophage of Clos-tndium ♪erfringens. J. Gen. Microbiol. 21,
622-630, 1959.
2 ) Sasarman, A., Antohi, M. : Presence des bacteriocines chez le Cl. ♪・erfringens. Arch. Roum. Path. Exp. Microbiol. 22,
377-380, 1963.
3 )内山一雄:ウェルシュ菌が産生する"bacteriocin" よう物質に関する研究, I,ウェルシュ菌々株 における本物質とactivity spectrumについて,
ウェルシュ菌のバクテリオシンと溶菌物質
鹿大医誌18, 131-144, 1966.
- 4 )内山一雄:ウェルシュ菌が産生するHbacteriocin よう物質に関する研究II,その性状について, 鹿大医誌18, 145-156, 1966.
5 ) Tubylewicz, H. : Experimental studies on the bacteriocinogeity in Clostridium per-fringens type A. I. Isolation of bacterio-cms and their antibacterial spectrum. Bull. Acad. Polon. Sci., Ser. sci. biol., 14, 31-36, 1966.
6 ) Tubylewicz, H. : Experimental studies on the bacteriocinogeneity in Clostridium ♪er-fringens type A. II. Properties of isolated bacteriocines. Bull. Acad. Polon. Sci., Ser. sci. biol. 14, 467-472, 1966.
7) Mahony, D.E., Butler, M.E.
:Bacterio-ernes of Clostridiumカerfringens. 1. Iso-lation and preliminary studies. Can. J.
Microbiol. 17, 1-6, 1971.
8) Sasarman, A., Antohi, M. : Une nouvelle classe de Welchicines. Rev. Can. Biol.
30, 183-189, 1971.
9) Mahony, D.E., Butler, M.E., Lewis, R.G.
: Bacteriocins of Clostridium perfringens. 2. Studies on mode of action. Can. J. Microbiol. 17, 1435-1443, 1971.
10) Hirano, S., Imamura, T. :Two types of bactenocins produced by Clostridium ♪er-fnngens. Acta Med. Univ. Kagoshima 14,
115-118, 1972.
ll) Hirano, S., Imamura, T. :A phage-asso-ciated lysin Clostridium ♪vrfringens. Acta Med. Univ. Kagoshima 14, 91-101, 1972. 12) Lowry, O.H., Rosebrough, T., Fair, A.L.,
Randall, R.J. : Protein measurement with the Folin phenol reagent. J. Biol. Chem.
193, 265-275, 1951.
13)永井裕:別冊・蛋白質・核酸・酵素:生物化学 実験法Ⅸ :物理化学的実験法(3), 3-14,共
立出版株式会社,東京, 1967.
14) Jetten, A.M., Vogels, G.D., de windt, F. '. Production and purification of Staphy-lococcus epidermidis bacteriocin. J. Bacte-nol. 112, 235-242, 1972.
15) Tagg, J.R., Dajani, A.S., Wannamaker,
195 L.W. : Bacteriocins of Gram-positive bac-term. Bact. Rev. 40, 722-756, 1976. 16) Clerk, D.J., Robson, R.M., Morris, ∫.G∴
purification of two Clostridium bacterio-cms by procedures appropriate to hydro-phobic proteins. Antimicrob. Agents Chemother. 7 , 256-264, 1975.
17) Wolff, A., Ionesco, H. :Purification et caractensation de la bacteriocine N 5 de Clostndiumperfringens BP6K-N5 type A. Ann. Microiol.Unst. Pasteur) 126B, 343-356, 1975.
18) Holland, I.B. : The purification and prop-erties of Megacin, a bacteriocin from 励alms megaterium. Biochem. T. 78, 641
-648, 1961.
19) Brock, T., Davie, J.M. : Probable identi-ty of a group D hemolysin with a bac-tenocin. J. Bacteriol. 86, 708-712, 1963. 20) Braun, V., Schaller, K., Wabl, M.R. :
Isolation and action of colicin M. Ant-lmicrob. Agents Chemother. 5 , 520-533,
1974.
21) Ozeki, M., Higashi, Y., Saito, H., An, T., Amako, T. :Identity of megacin A with phospholipase A. Biken J. 9, 201
-213, 1966.
22) Ralston, D.J., Baer, B., Lieberman, M., Kru-eger, A.P. : Staphylococcal virolysin, a phage-induced lysin ; Its differentiation from the autolysin of normal cells. J. Gen. Physiol. 40, 791-807, 1957.
23) Ogata, S., Choi, K.H., Ikeda, Y., Hongo,M. : Bacterial lysis of Clostridium species. HI. properties of premature lysis pro-voked by antibiotic in mytomycin Cin-duced lysis of Clostridium saccharoperbutyl-acetonicum. J. Gen. Appl. Microbiol. 20,
153-168, 1974.
24) Martin, H.H., Kemper, S. : Endo-N-acetyl-glucosammidase from Clostridiump<erfrin-gens, lytic for cell wall murein of gram negative bacteria. J. bacteriol. 102, 347-350, 1970.
au-tolysis in Clostridium petfringens type A. Biochem. Biophys. Acta 338, 605-618, 1974.
ウェルシュ菌のバクテリオシンと溶菌物質
Abstract
Partial
purification
and some properties
of the
bacteriocin
and the
bacteriolytic
substance
produced
by Clostridium
perfringens
strain
K29
197