TABLE OF CONTENTS
略号一覧表 ... 2 2.6.1 緒言 ... 3 1 非臨床試験計画の概要 ... 3 2 参考文献 ... 6LIST OF FIGURES
Figure 1: ラブリズマブ遺伝子組換え抗体の基本構造 ... 3略号一覧表
略語・用語 内容(英語) 内容(日本語)
ADCC antibody-dependent cell-mediated cytotoxicity 抗体依存性細胞傷害
aHUS atypical hemolytic uremic syndrome 非典型溶血性尿毒症症候群
C1q complement component C1q 補体C1q
C5 complement component C5 補体第5 成分
C5a an active metabolite of C5 (proinflammatory anaphylatoxin)
C5 の活性生成物(炎症誘発性アナフィラ トキシン)
C5b an active metabolite of C5 (initiating subunit of the terminal complement complex [C5b-9])
C5 の活性生成物[終末補体複合体(C5b-9) の開始サブユニット]
CAHA cynomolgus anti-human antibody/ies カニクイザル抗ヒト抗体
CDC complement-dependent cytotoxicity 補体依存性細胞傷害
CDR(s) complementarity-determining region(s) 相補性決定領域 CH constant domain, heavy chain 重鎖定常領域
FcR Fc receptor Fc 受容体
FcRn neonatal Fc receptor 胎児性Fc 受容体
FcγR(s) Fc gamma receptor(s) Fcγ 受容体
FcγRI Fc gamma receptor Type I Fcγ 受容体 I
FcγRIIb/c Fc gamma receptor Type IIb/c Fcγ 受容体 IIb/c
FcγRIIIa Fc gamma receptor Type IIIa Fcγ 受容体 IIIa
FcγRIIIb Fc gamma receptor Type IIIb Fcγ 受容体 IIIb
GLP Good Laboratory Practice 医薬品の安全性に関する非臨床試験の実
施の基準
gMG generalized myasthenia gravis 全身型重症筋無力症
NOD-scid non-obese diabetic/severe combined immunodeficient
非肥満糖尿病/重症複合免疫不全
PK pharmacokinetic(s) 薬物動態
PNH paroxysmal nocturnal hemoglobinuria 発作性夜間ヘモグロビン尿症
TK toxicokinetics トキシコキネティクス
2.6.1 緒言
1 非臨床試験計画の概要
ラブリズマブ(遺伝子組換え)は、補体経路の変換酵素により活性化される補体第5 成分(C5) に結合して、その活性化を阻害するヒト化モノクローナル抗体である。その活性化阻害作用によ り、炎症誘発性アナフィラトキシンC5a の遊離及び C5b を介した終末補体複合体の形成を阻害す る。エクリズマブは、発作性夜間ヘモグロビン尿症(PNH)、非典型溶血性尿毒症症候群(aHUS) 及び全身型重症筋無力症(gMG)を適応として承認され、ソリリス®の商品名で製造販売されて いる。ラブリズマブは、エクリズマブと構造的に関連しており、投与間隔を延長するために、エ クリズマブに比較して血清半減期を延長しそれに伴う薬理活性持続時間を延長するよう改変され ている。これにより、制御不能な終末補体によりもたらされるPNH、aHUS 及び他の疾患に対し て有効な治療薬としての新たな選択肢を提供する。ラブリズマブの申請用法・用量は、成人及び 小児aHUS[(補体介在性の血栓性微小血管障害(TMA)]患者に対し、体重に基づき 600~3600 mg の維持用量を4 週又は 8 週に 1 回静脈内投与するものである。 Figure 1: ラブリズマブ遺伝子組換え抗体の基本構造 上記Figure 1に示すように、ラブリズマブは、エクリズマブの重鎖の4 個の固有のアミノ酸を 置換して改変した[27 番目のチロシンをヒスチジンに(Y27H)、57 番目のセリンをヒスチジンへ (S57H)、428 番目のメチオニンをロイシンに(M428L)、並びに 434 番目のアスパラギンをセリ ンに(N434S)置換](重鎖アミノ酸の既知の番号はKabat, 1991に基づく。ラブリズマブの実際 のアミノ酸置換位置はH27、H57、L429 及び S435 である。)。重鎖可変領域の相補性決定領域(CDRs)のCDR1 及び CDR2 の 2 個のアミノ酸置換(Y27H 及び S57H)は、血管コンパートメント(pH7.4) での C5 に対する十分な結合親和性を維持し、終末補体活性の完全な阻害作用を確保すると同時 に、早期エンドソームの酸性化環境(約pH6.0)での C5 への結合を不安定化するために導入され た。第3 重鎖定常領域(CH3)の 2 個のアミノ酸置換(M428L 及び N434S)は、胎児性 Fc 受容 体(FcRn)に対する結合を増強し、FcRn による早期エンドソームから血管コンパートメントへ の抗体リサイクル率を高めるために導入された。これらのFc 置換により、ラブリズマブの FcRn に対する親和性は、pH6.0 でエクリズマブと比較して約 10 倍に増加した。ヒト C5、FcRn 並びに 補体依存性細胞傷害(CDC)及び抗体依存性細胞傷害(ADCC)の開始因子{それぞれ補体 C1q 及び Fcγ 受容体[FcγR(s)]}への結合に対するこれらのアミノ酸置換の影響については、in vitro で検討した(RTR-0003v1.0 及び RTR-0005v1.0)。これらの結果は、ラブリズマブがエクリズマブ よりもC5 及び FcRn への結合において高い pH 依存性を示し、抗原介在性消失の減少及び FcRn 介在性リサイクリングが向上することと一致する。さらに、ラブリズマブに導入されたアミノ酸 置換は、エクリズマブと同様に、FcγRI、FcγRIIb/c、FcγRIIIa、FcγRIIIb 又は C1q に対する結合親 和性をほとんど示さなかった。可溶性ヒト C5 に対するラブリズマブの結合特異性を考え合わせ ると、ラブリズマブがin vivo で ADCC、又は CDC を介した細胞毒性を誘導することは予測され ない。 ラブリズマブのヒト以外の動物種のC5 に対する交差反応性の有無を評価するため、in silico 評 価及びin vitro で C5 結合試験及び C5 活性試験を実施した(RTR-0006v1.0)。その結果、ラブリズ マブはエクリズマブと同様に、in vitro でヒト C5 活性に対する強力な阻害活性を示したが、ヒト 以外の動物種(カニクイザルを含む)のいずれに対しても薬理活性又は交差反応性を示さなかっ た。これらの結果に加えて、ヒト及びカニクイザル組織標準パネルを用いた交差反応性試験[ラ ブリズマブ(PNH)の CTD M2.6.6.8]を GLP 適用下で行ったところ、交差反応性は認められな かった。 ラブリズマブは、一貫してヒト C5 に極めて高い特異性を示すことから、ラブリズマブの非臨 床試験の薬物動態/トキシコキネティクス(PK/TK)データのヒトへの外挿性は限定的である。 さらに、ラブリズマブに対する顕著な免疫反応性がカニクイザル及びウサギに認められた(試験 番号1727-009 及び試験番号 1727-050)ため、ラブリズマブについては、限られた非臨床 PK 評価 のみを実施した。 非臨床PK 試験のいくつかはヒト C5 を用いて実施され、in vivo でラブリズマブの半減期をエク リズマブと比較した(RTR-0008v1.0)。ラブリズマブの PK 試験は、遺伝的に C5 を欠損し、外因 性のタンパク質に対して免疫応答を開始できない非肥満糖尿病/重症複合免疫不全(NOD-scid) マウスで実施された(M2.6.2)。C5 の恒常的な発現をシミュレーションし、抗 C5 モノクローナル 抗体の抗原介在性消失モデルを作成するために、ヒト C5 が連日投与された。本試験結果から、 ヒトC5 存在下でのマウス PK モデルのラブリズマブの半減期は、エクリズマブと比較して顕著な 延長が認められ、ラブリズマブの抗原介在性消失が減少したことが示された。さらに、ラブリズ マブの曝露時間の延長は、エクリズマブに比較して薬理活性持続時間の延長をもたらした。 さらに、GLP 非適用下でカニクイザルを用いた単回投与薬物動態試験を実施した(試験番号 1227-009)。カニクイザル抗ヒト抗体(CAHA)が、ラブリズマブのクリアランスを増加させ、全 身の曝露を減少させたが、血漿中濃度推移から、ラブリズマブの全身循環時間が長くなっている ことが示された。ヒト以外の動物種でのヒトタンパク質に対する全身性免疫反応は、ヒトでの免 疫原性リスクとは通常関係はないと考えられている(Leach, 2014; Vugmeyster, 2012)。
ラブリズマブはヒト以外の動物種の C5 と交差反応性を示さないことから、非臨床安全性試験 ではサロゲート分子(抗体)を使用することを計画した。C5 の長期間阻害による毒性の有無を検 討し、生殖機能に対する影響を評価するため、マウスでの26 週間反復投与毒性試験、並びに生殖 発生毒性試験(Segments I、II 及び III)をサロゲート抗体であるマウス抗マウス C5 抗体 BB5.1 を用いて実施した。ラブリズマブと同様に、サロゲート抗体であるBB5.1 は C5 に直接結合して 補体変換酵素によるタンパク分解活性を阻害し、マウスの C5a 及び C5b-9 の生成を抑制するが、 これらの試験からはC5 を長期間阻害する用量での毒性は認められなかった[ラブリズマブ(PNH) のCTD M2.6.6]。 さらに、高濃度のラブリズマブ皮下投与製剤を用いて、局所刺激性試験をウサギを対象として 実施した。ラブリズマブをウサギに単回皮下投与したところ、局所反応性を示す所見は認められ ず、忍容性は良好であった。しかし、ラブリズマブをウサギに反復皮下投与したところ、免疫反 応が誘発され、免疫複合体に起因する全身性の III 型過敏症反応が示唆されたが、毒性所見とは 判断されなかった(試験番号 1727-050)。このようなヒトタンパク質の投与に起因する免疫学的 反応は、動物では一般的に認められている。ウサギや他の動物種でのヒトタンパク質への全身性 免 疫 反 応 は 、 通 常 、 ヒ ト に お い て 問 題 と な る 免 疫 原 性 リ ス ク と は な ら な い と 考 え ら れ る (Vugmeyster, 2012)。これらのことから、本試験でみられた毒性所見は、ヒトでのラブリズマブ の安全性には関連しないと考えられる。 非臨床試験では、マウスサロゲート分子BB5.1(抗 C5 抗体)を用いた長期 C5 阻害の安全性評 価のための非臨床毒性試験と同時にラブリズマブ分子で予想されるC5 阻害を確認する種々の PK 及び薬力学(PD)評価が計画された。これらの試験では、ラブリズマブの投与により長期間 C5 が完全に阻害されることに関連する有害作用は認められず、ラブリズマブは血清半減期及び関連 する薬理活性持続時間をエクリズマブに比較して延長し、投与間隔の延長を可能にするという結 論が支持された。
2 参考文献
Kabat EA, Wu TT, Perry HM, Gottesman KS. Sequences of proteins of immunological interest. No 91:3242. NIH Publication1991.
Leach MW, Rottman JB, Hock MB, Finco D, Rojko JL, Beyer JC. Immunogenicity/hypersensitivity of biologics. Toxicol Pathol. 2014;42(1):293-300.
Vugmeyster Y, Xu X, Theil FP, Khawli LA, Leach MW. Pharmacokinetics and toxicology of therapeutic proteins: Advances and challenges. World J Biol Chem. 2012;3(4):73-92.
TABLE OF CONTENTS
略号一覧表 ... 3 2.6.2 薬理試験の概要文 ... 4 1 まとめ ... 4 2 効力を裏付ける試験 ... 7 2.1 ラブリズマブ及びエクリズマブに対するpH の効果 ... 7 2.2 ラブリズマブのエピトープマッピング ... 8 2.3 ヒトC5 存在下でのラブリズマブとエクリズマブの in vivo での作用の比較 ... 13 3 副次的薬理試験 ... 16 3.1 多価抗体複合体のFcγRs に対する結合 ... 17 3.2 一価抗体ラブリズマブのFcγRs に対する結合 ... 18 3.3 バイオレイヤー干渉法を用いた抗体に対するC1q 結合の測定 ... 18 3.4 結論 ... 19 4 安全性薬理試験 ... 21 4.1 In vitro 安全性薬理試験 ... 21 4.2 In vivo 安全性薬理試験 ... 21 5 薬力学的薬物相互作用 ... 22 6 考察及び結論 ... 23 7 参考文献 ... 25LIST OF TABLES
Table 1: ラブリズマブ及びエクリズマブの精製ヒトC5 に対する pH 依存的結合カイネ ティクスパラメーター ... 7 Table 2: C5 変異体に対するラブリズマブの結合カイネティクスパラメーター ... 10 Table 3: ラブリズマブ、エクリズマブ及びh5G1.1-IgG1 の FcγRs 又は C1q との相互作 用のまとめ ... 20LIST OF FIGURES
Figure 1: ラブリズマブ及びエクリズマブのFcRn に対する結合カイネティクス ... 8 Figure 2: ヒト以外の霊長類10 種及び非霊長類哺乳動物種 13 種の C5 タンパク質の MG7 ドメインのアミノ酸配列比較 ... 10 Figure 3: 遺伝子組換えヒトC5 変異体を加えた C5 除去血清のニワトリ赤血球溶血に対 するラブリズマブの阻害作用 ... 11 Figure 4: チンパンジー、ヒヒ、アカゲザル及びカニクイザル血清の補体介在性溶血に 対する抗C5 抗体の作用 ... 12 Figure 5: イヌ、ウサギ、モルモット及びラット血清の補体介在性溶血に対する抗 C5 抗体の作用 ... 13 Figure 6: C5 除去ヒト血清を補強したマウス血清の補体介在性溶血に対する阻害作用 ... 13 Figure 7: ヒト C5 の存在下及び非存在下でのラブリズマブ及びエクリズマブの血清中 濃度比率(%)... 14 Figure 8: NOD-scid マウスでのラブリズマブ又はエクリズマブの存在下及び非存在下の ヒトC5 介在性 cRBC 溶血 ... 15Figure 9: FcγRI、FcγRIIa、FcγRIIb/c、FcγRIIIa 及び FcγRIIIb に対する多価抗体複合体の 結合 ... 17
Figure 10: h5G1.1-IgG1 の FcγRI への結合のセンサーグラム ... 18
Figure 11: ラブリズマブのFcγRIIa への結合のセンサーグラム ... 18
略号一覧表
略語・用語 内容(英語) 内容(日本語)
Ab antibody 抗体
ADCC antibody-dependent cell-mediated cytotoxicity 抗体依存性細胞傷害
C1q complement component C1q 補体C1q
C5 complement component C5 補体第5 成分、補体 C5
C5a an active metabolite of C5 (proinflammatory anaphylatoxin)
C5 の活性生成物(炎症誘発性アナフィラ トキシン)
C5b an active metabolite of C5 (initiating subunit of the terminal complement complex [C5b-9])
C5 の活性生成物[終末補体複合体(C5b-9) の開始サブユニット]
C5b-9 terminal complement complex 終末補体複合体
CCP complement classical pathway 補体古典経路
CDC complement-dependent cytotoxicity 補体依存性細胞傷害
CDR(s) complementarity-determining region(s) 相補性決定領域
CH constant domain, heavy chain 重鎖定常領域
CSR clinical study report 治験総括報告書
cRBCs chicken red blood cells ニワトリ赤血球
ELISA enzyme-linked immunosorbent assay 酵素結合免疫吸着測定法
FcR Fc receptor Fc 受容体
FcRn neonatal Fc receptor 胎児性Fc 受容体
FcγR(s) Fc gamma receptor(s) Fcγ 受容体
FcγRI Fc gamma receptor Type I Fcγ 受容体 I
FcγRIIa Fc gamma receptor Type IIa Fcγ 受容体 IIa
FcγRIIb/c Fc gamma receptor Type IIb/c Fcγ 受容体 IIb/c
FcγRIIIa Fc gamma receptor Type IIIa Fcγ 受容体 IIIa
FcγRIIIb Fc gamma receptor Type IIIb Fcγ 受容体 IIIb
IgG immunoglobulin G 免疫グロブリンG
Ka association rate 結合速度
KD dissociation constant 解離定数
Kd dissociation rate 解離速度
NOD-scid non-obese diabetic/severe combined immunodeficient
非肥満糖尿病/重症複合免疫不全
PD pharmacodynamics 薬力学
PK pharmacokinetic(s) 薬物動態
2.6.2 薬理試験の概要文
1 まとめ
ラブリズマブは、エクリズマブの誘導体で、補体経路の変換酵素により活性化される補体第 5 成分(C5)に結合してその活性化を阻害するヒト化モノクローナル抗体である。その活性化阻害 作用により、炎症誘発性アナフィラトキシンC5a の遊離及び C5b を介した終末補体複合体の形成 を阻害する。ラブリズマブの C5 に対するエクリズマブと同様の結合特異性を確保するために、 C5 と直接接触する可変領域のアミノ酸残基はエクリズマブから改変しなかった。さらに、親和性 に関係する遺伝子改変は、抗原介在性消失を十分に低減し、生理的条件で終末補体の活性化を完 全に阻害する機能も維持するよう調整された。 効力を裏付ける試験は、毒性試験での動物種の選択に役立てるため、並びにin vitro でのラブリ ズマブのヒトC5 及びヒト胎児性 Fc 受容体(FcRn)との結合カイネティクスに対する pH の影響 を評価するために実施した。ヒト及びヒト以外の動物種の C5 分子のエピトープマッピング試験 及び配列比較から予測され、in vitro 結合試験及び活性試験から確認されたとおり、ラブリズマブ 及びエクリズマブで試験をしたヒト以外のいずれの動物種に対する薬理活性も全く検出されなか った。交差反応性が認められないことからラブリズマブのin vivo 非臨床試験は制限を受けた。し かし、C5 に関連する毒性の有無の検討は、マウス C5 に特異的なサロゲート抗体を用いてマウス で実施した[ラブリズマブ(PNH)の CTD M2.6.6 及び 2.6.7 参照]。ラブリズマブの薬物動態(PK) 及び薬力学(PD)試験は、恒常的なヒトでの C5 の生成をシミュレーションする非肥満糖尿病/ 重症複合免疫不全(NOD-scid)マウスモデルを用いて実施した。 ラブリズマブの動物種との交差反応性の有無の評価及び毒性評価動物種の選択のために、C5 エピトープマッピング試験を実施し、ラブリズマブの結合部位を確認した(RTR-0006v1.0)。ヒト C5 の MG7 ドメインの 4 残基(K879、K882、R885 及び W917)がラブリズマブの結合に重要で あることが明らかにされた。ヒト以外の動物種の交差反応性に関して、ヒトに特有である W917 が特に重要であった。ヒト以外の霊長類種10 種及び非霊長類哺乳動物種 13 種から確認、又は予 測されたC5 タンパク質の MG7 ドメインのアミノ酸配列比較を行ったところ、K879、K882 及び R885 はチンパンジー、ヒヒ及びアカゲザルに保存され、ヒト及びチンパンジーの MG7 ドメイン のアミノ酸配列は、E904(グリシン)及び W917(セリン)の 2 ヵ所のみが異なっていた。 ラブリズマブの結合に対するK879、K882、R885 及び W917 アミノ酸の重要性を評価するため に、遺伝子組換えヒト C5 変異体を用いて、in vitro での結合活性及び溶血活性を評価した (RTR-0006v1.0)。ラブリズマブは、遺伝子組換えヒト C5 の W917S 変異体に対する結合活性を全 く示さず、また、W917S 変異体を含む反応系の in vitro での溶血も阻害しなかった。これらの試 験結果はラブリズマブの結合に対するヒト C5 の W917 アミノ酸残基の重要性を裏付けており、 ラブリズマブは、対応するアミノ酸が保存されていない動物種の C5 には結合しないと考えられ た。結果的に、ラブリズマブは、マウス、ヒト以外の霊長類4 種(チンパンジー、ヒヒ、アカゲ ザル及びカニクイザル)、あるいは毒性試験に一般的に使用されるその他の非霊長類哺乳動物(イ ヌ、ウサギ、モルモット及びラット)の血清による抗体で感作したニワトリ赤血球(cRBCs)の in vitro での溶血を検討したいずれの濃度においても阻害しなかった。これらの試験結果から、試 験を実施したヒト以外の全ての動物種の C5 に対するラブリズマブの結合は、ラブリズマブの毒 性、PK 及び PD を試験するためには不十分であることが示された。 ラブリズマブは、エクリズマブのin vivo での半減期を延長するようデザインされたエクリズマ ブの誘導体である。ラブリズマブは、エクリズマブの相補性決定領域(CDRs)及び重鎖定常領域にそれぞれC5 及び FcRn との結合において pH 感受性を付与するアミノ酸置換をもつ。これらの 置換は、抗原介在性消失を低減し、早期エンドソームから血管コンパートメントへの抗体リサイ クル率を高め、エクリズマブと比較して血中半減期を延長することを目的としてデザインされた。 in vitro でのヒト C5 及び FcRn に対する結合に関するこれらの置換の影響について検討した (RTR-0003v1.0)。その結果、CDRs 領域の 2 個のアミノ酸の置換(Y27H 及び S57H)により、ヒ トC5 に対するラブリズマブの親和性はエクリズマブと比較して pH7.4 では約 17 倍及び pH6.0 で は約36 倍減少した。また、重鎖定常領域の 2 個のアミノ酸の置換(M428L 及び N434S)により、 FcRn に対するラブリズマブの親和性はエクリズマブと比較して pH6.0 で約 10 倍増加した。以上 から、これらの変化により、エンドソーム内でのpH6.0 の条件下での抗体・C5 複合体の解離が加 速し、その結果エンドソームから血管コンパートメントへの抗体リサイクリングの効率が高まる ことにより、ラブリズマブの循環血中半減期及び薬理活性持続時間が延長すると予測された。 ラブリズマブの PK 及び PD は、抗原介在性消失のマウスモデルを用いた試験で評価した。遺 伝的にC5 を欠損し、さらに外因性のタンパク質に対して免疫応答を開始できない NOD-scid マウ スにヒト C5 を試験期間を通して投与することにより C5 の恒常的発現をシミュレーションした (RTR-0008v1.0)。本モデルは、ラブリズマブの PK に対する重鎖定常領域の(ヒト FcRn に対す る親和性が増加する)アミノ酸置換の影響を評価するためのものではない。本試験結果から、用 いたマウス PK モデルではエクリズマブと比較してラブリズマブの有意な半減期の延長が認めら れ、ラブリズマブの抗原介在性消失が低減したことが示された。さらに、ラブリズマブの長時間 の曝露は、エクリズマブと比較して薬理活性がより長時間持続することと一致した。すなわち、 ラブリズマブのY27H 及び S57H の置換により、NOD-scid マウスにおけるヒト補体 C5 存在下で のPK/PD 特性がエクリズマブと比較して改善された。 ラブリズマブのエフェクター機能の有無を評価するため、副次的薬理試験としてのin vitro 試験 を実施した(RTR-0005v1.0)。ラブリズマブは、安定なジスルフィド結合でペアリングした IgG2 をエフェクター機能を持たないIgG4 と組み合わせたキメラヒト免疫グロブリン G(IgG)2/G4 の Fc 領域を有する。ラブリズマブは可溶性抗原を標的としているため、抗体依存性細胞傷害(ADCC) 又は補体依存性細胞傷害(CDC)誘発能の直接的な評価はできなかった。そのため、代替えとし て、ラブリズマブ又はエクリズマブのFcγ 受容体[FcγR(s) ]及び補体 C1q への結合を直接測定 した。ラブリズマブは、C1q、FcγRI、FcγRIIb/c、FcγRIIIa 又は FcγRIIIb への結合を示さなかった が、FcγRIIa に対してはアイソタイプ・コントロール抗体である IgG1 より約 3 倍弱い親和性を示 した。これらの結果は、ラブリズマブはFcγR 又は C1q を介して、それぞれ ADCC 又は CDC を 誘発するエフェクター機能を発現する可能性はほとんどないという仮説を支持している。可溶性 抗原であること、及びエフェクター分子に対する結合が弱いことから、ラブリズマブはin vivo で ADCC 又は CDC を誘発する可能性はないと考えられる。 これらの結果は、ラブリズマブにおける遺伝子改変によるアミノ酸の置換は、ヒトにおいて、 ラブリズマブの半減期及び対応する薬理活性をエクリズマブと比較して有意に延長し、in vivo で、 ラブリズマブはエクリズマブと同様にADCC 又は CDC を誘発しないという仮説を支持する。 以上をまとめると、ラブリズマブは pH7.4 で C5 に対して高い親和性を維持し、エクリズマブ と同一のエピトープを認識する。エクリズマブの作用機序、すなわちC5 への結合は pH7.4 で血 管コンパートメントで主に生じる。pH7.4 での C5 に対するラブリズマブの親和性(解離定数 [KD] = 490 pM、RTR-0003v1.0)は、循環血中での C5 ベースライン濃度を 3 オーダー以上下回っ ている[ALXN1210-HV-101 CSR Table 14.2.2.1;(Rawal, 2001)]。すなわち、エクリズマブ
(KD = 30 pM、RTR-0003v1.0)と同様に、飽和濃度のラブリズマブ存在下で C5 濃度が増加して も、in vitro 試験及び臨床試験結果(ALXN1210-HV-PNH-201)で明らかにされているとおり、そ の増加した C5 濃度は、終末補体活性を効果的に阻害できる程度であり、その限界点を十分に下 回る。ラブリズマブとエクリズマブを飽和濃度で直接比較したin vitro 試験において、両者の終末 補体阻害活性は同等であることが示されている(RTR-0006v1.0)。
2 効力を裏付ける試験
ラブリズマブは、エクリズマブの遺伝子組換え体であり、同様の抗原結合特異性を有するが、 エクリズマブと比較して半減期の延長が認められる。ラブリズマブは、重鎖のY27H、S57H、M428L 及びN434S(エクリズマブ重鎖アミノ酸のナンバリングはKabat, 1991に従った。ラブリズマブの 実際のアミノ酸置換位置はH27、H57、L429 及び S435 である。)の 4 個のアミノ酸がエクリズマ ブと異なる。これらの置換は、抗原介在性消失による抗体の消失の低減、及び FcRn による抗体 リサイクリングの効率の増大という2 つの異なる機序によって、循環血中のラブリズマブの半減 期を延長することにより、投与間隔を延長できるようデザインされている。2.1 ラブリズマブ及びエクリズマブに対する pH の効果
RTR-0003v1.0 試験は、ラブリズマブのヒト C5 及び FcRn への結合カイネティクスにおけるア ミノ酸置換の影響を評価するために実施した。 ラブリズマブの重鎖可変領域のCDR1 及び CDR2 の 2 個のアミノ酸置換(Y27H 及び S57H)は、 早期エンドソームの酸性化環境(約pH6.0)で C5 に対する結合を不安定化する一方で、終末補体 経路の完全な阻害作用を確保するため、血管コンパートメント(pH7.4)で C5 に対する十分な結 合親和性を維持する目的で導入した。これらのヒスチジン置換の導入は、ピノサイトーシス後の 早期エンドソームの酸性化環境で遊離型抗体に対する抗体・C5 複合体の解離率が著しく増加する ことにより、抗原介在性消失を顕著に低減するようデザインされた。これらのアミノ酸置換は、 ラブリズマブのC5 に対する KDをエクリズマブと比較してpH7.4 で約 17 倍、及び pH6.0 で約 36 倍増大させた(Table 1)。 Table 1: ラブリズマブ及びエクリズマブの精製ヒトC5 に対する pH 依存的結合カイネ ティクスパラメーター pH Association Rate: Ka (1/M*s) Dissociation Rate: Kd (1/s) Dissociation Constant: KD (M) Chi2 ラブリズマブ 8.0 6.25 × 105 1.33 × 10−4 2.13 × 10−10 0.055 7.4 4.62 × 105 2.27 × 10−4 4.91 × 10−10 0.045 7.0 4.28 ×105 3.90 × 10−4 9.11 × 10−10 0.028 6.5 4.08 × 105 8.94 × 10−4 2.19 × 10−9 0.172 6.0 1.63 × 105 3.54 × 10−3 2.18 × 10−8 0.373 Eculizumab 8.0 1.39 × 106 2.04 × 10−5 1.47 × 10−11 0.104 7.4 1.10 × 106 3.23 × 10−5 2.93 × 10−11 0.094 7.0 8.86 × 105 6.34 × 10−5 7.15 × 10−11 0.032 6.5 8.41 × 105 1.73 × 10−4 2.06 × 10−10 0.037 6.0 7.05 × 105 4.28 × 10−4 6.06 × 10−10 0.092 Source: RTR-0003v1.0 ラブリズマブの第3 重鎖定常領域(CH3)における 2 個のアミノ酸の置換(M428L 及び N434S) は、FcRn に対する結合を促進し、FcRn による早期エンドソームから血管コンパートメントへの 抗体のリサイクル率を高めるために導入した。これらFc 領域のアミノ酸置換により、ラブリズマ ブのFcRn に対する親和性は、エクリズマブと比較して pH6.0 では約 10 倍増大した。ラブリズマ ブ及びエクリズマブの FcRn に対する結合カイネティクスのセンサーグラム及びラングミュアモ デルをFigure 1に示した。ラブリズマブ及びエクリズマブは、いずれもpH7.4 で FcRn に対する特異的結合性を示さなかった。 これらの結果、すなわちC5 及び FcRn への結合に対するこれらの変化から、ピノサイトーシス 後の早期エンドソームの酸性化環境で抗体・C5 複合体の解離、及び早期エンドソームから FcRn を介した血管コンパートメントへリサイクルされる抗体分画の両方が増大することにより、抗原 介在性消失が顕著に低減することが予測された。本試験報告書 RTR-0003 は M5.3.2.3.1 に記載し た。 Figure 1: ラブリズマブ及びエクリズマブのFcRn に対する結合カイネティクス
Abbreviations: FcRn = human neonatal Fc receptor、KD = dissociation constant、M = molar
Source: RTR-0003v1.0
2.2 ラブリズマブのエピトープマッピング
RTR-0006v1.0 試験は、ラブリズマブの毒性、PK 及び PD 試験で使用可能な動物種の適合性を 評価するために実施した。 ラブリズマブの親抗体であるエクリズマブの親抗体であるマウス型抗体(m5G1.1)で以前実施 したエピトープマッピングでは、マウス型抗体に対するエピトープは、ヒト C5 の α 鎖のアミノ 酸822~933 であった(Alexion GTR-0055.00-AD01)。ペプチドマップ及びアミノ酸置換より、ア ミノ酸879~883 に対するエピトープをさらに絞り込み、K879 及び K882 のリジン残基が結合に 関与していることを明らかにした。臨床試験結果から、ヒト C5 のアルギニン 885(R885)の変 異もラブリズマブの親抗体であるエクリズマブの結合を崩壊すると報告されている(Nishimura, 2014)。ラブリズマブに対するC5 のエピトープについて明らかにするため、本試験に引き続いて、 複 数 の 論 文 で 、 エ ク リ ズ マ ブ と C5 と の 結 合 特 異 性 が 確 認 さ れ て い る (Brachet, 2016; Schatz-Jakobsen, 2016)。本試験では、ラブリズマブがm5G1.1 及びエクリズマブと同じ C5 のエピトープを認識することを確認するために、主要アミノ酸を選択的に置換した遺伝子組換えヒトC5 変異体への結合カイネティクスパラメーターを、表面プラズモン共鳴法(SPR)を用いて測定し た。ラブリズマブの遺伝子組換えヒトC5 変異体に対する in vitro での薬理活性についても、cRBCs の補体介在性溶血に対する阻害能を測定することより評価した。さらに、動物モデルでのラブリ ズマブの薬理活性の有無について、チンパンジー、ヒヒ、アカゲザル、カニクイザル、イヌ、ウ サギ、モルモット、ラット及びマウス血清を用いたin vitro の補体介在性溶血活性に対する阻害作 用を測定することより評価した。 ヒトC5 並びにヒト以外の霊長類 10 種及び非霊長類哺乳動物 13 種に関して公表されている C5 のアミノ酸配列のMG7 ドメインの比較を行った結果、K879、K882 及び R885 がチンパンジー、 ヒヒ及びアカゲザルに保存されており(Figure 2)、並びに m5G1.1 及びエクリズマブは、チンパ ンジー及びヒヒ血清で非常に限られた薬理活性を示すものの、アカゲザル血清では全く活性を示 さないことを明らかにした(Alexion GTR-0050.00-AD01)。ヒト及びチンパンジーMG7 ドメイン のアミノ酸配列は、E904(グリシン)及び W917(セリン)の 2 ヵ所のみが異なっている(括弧 内はチンパンジーのアミノ酸)。E904 のグリシンへの置換はチンパンジー及びボノボ(ヒト科チ ンパンジー属に分類される霊長類)に特異的であり、ヒヒ及びアカゲザルの MG7 ドメインのア ミノ酸配列は、I839(バリン)、S893(アスパラギン)、H908(グルタミン)、W917(セリン)及 びT926(セリン)の 5 個のアミノ酸残基がヒトと異なっている(括弧内はヒヒ及びアカゲザルの アミノ酸)。これらE904、I839、S893、H908 及び T926 の相異はおおむね動物種間で保存されて いないが、W917(セリン)の置換は、m5G1.1 及びエクリズマブで in vitro 薬理活性を評価した全 てのヒト以外の動物種に保存されている。 Laursen らによって報告されたヒト C5 の MG7 ドメインの構造解析(ヒト C5 とコブラ毒因子の 複合体の結晶構造解析結果)から(Laursen, 2011)、K882、R885 及び W917 はそれぞれ 2Ǻ 以内の クラスターを形成して、複合体中でコブラ毒因子と直接接触していることが明らかとなった。こ の試験結果は、当該3 アミノ酸は全て m5G1.1 及びエクリズマブのヒト C5 との相互作用に重要で あり、ヒト以外の相同分子種においてW917 がセリンに置換されていることが、ヒト以外のいず れの動物種においても薬理活性を顕著に減少、又は消失させる決定的な違いになるという仮説と 一致する。 次に、ラブリズマブの結合に対するアミノ酸K879、K882、R885 及び W917 の重要性を評価す るために、ヒト C5 遺伝子組換え変異体クローンを発現させ、SPR を用いて、精製した天然型ヒ トC5 及び遺伝子組換えヒト C5 変異体に対するラブリズマブの結合カイネティクスパラメーター を測定した(Table 2)。その結果、ラブリズマブは、天然型ヒト C5 及び遺伝子組換えヒト C5 (RAA031)にそれぞれ、500 pM 及び 800 pM の KDで結合した。ラブリズマブは、C5 の遺伝子 組換えK879R_K882R(RAA049)に 2 nM のやや弱い親和性で結合した。ラブリズマブは、C5 の 遺伝子組換えR885H(RAA047)、W917S(RAA048)又は K879E_K882E(RAA050)のいずれに 対しても特異的な結合活性を示さなかった。RAA047、RAA048 及び RAA050 に関して非特異的 結合が幾分認められた。
Figure 2: ヒト以外の霊長類10 種及び非霊長類哺乳動物種 13 種の C5 タンパク質の MG7 ドメインのアミノ酸配列比較 Source:RTR-0006v1.0 Table 2: C5 変異体に対するラブリズマブの結合カイネティクスパラメーター Analyte Association Rate: Ka (1/M × s) Dissociation Rate: Kd (1/s) Dissociation Constant: KD (M) Chi2 Native human C5 (hC5) 4.26 × 105 2.13 × 10−4 5.01 × 10−10 0.0259
Recombinant wild type C5
(RAA031) 4.33 × 10 5 3.49 × 10−4 8.06 × 10−10 0.0591 R885H C5 (RAA047) ND W917S C5 (RAA048) ND K879R_K882R C5 (RAA049) 1.18 × 105 2.09 × 10−4 1.77 × 10−9 2.6 K879E_K882E C5 (RAA050) 8.36 × 103 1.06 × 10−4 1.27 × 10−8 0.0772
Abbreviations: Ka = association rate、Kd = dissociation rate、KD = dissociation constant、ND = not detectable
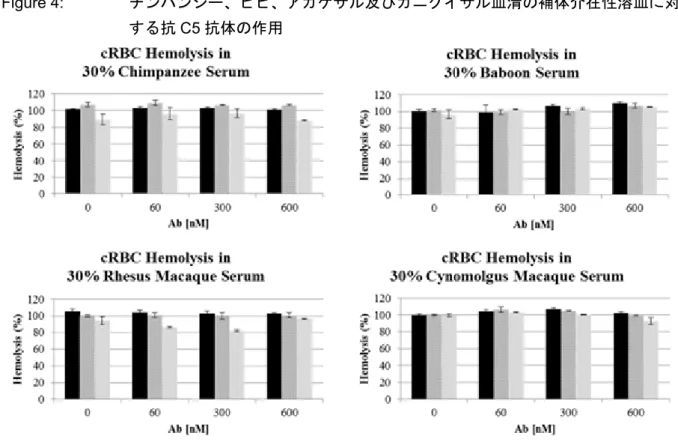
Source:RTR-0006v1.0 次に、in vitro でのラブリズマブの薬理活性を、抗体で感作した cRBCs の補体古典経路(CCP) 介在性溶血の阻害作用の測定より評価した。遺伝子組換えヒト C5 変異体に対する薬理活性は、 正常ヒトC5 除去血清に精製した遺伝子組換え C5 を加えて補体活性を再構築することにより評価 した。遺伝子組換えヒトC5 変異体 RAA047(R885H)及び RAA048(W917S)に対するラブリズ マブの結合が認められなかったことと一致して、ラブリズマブは、これらより再構築した反応系 の溶血を阻害しなかった(Figure 3)。 次に、抗体で感作したcRBC 溶血を、ヒト以外の霊長類 4 種(チンパンジー、ヒヒ、アカゲザ ル及びカニクイザル)及び毒性試験で一般的に使用される非霊長類哺乳動物4 種(イヌ、ウサギ、 モルモット及びラット)の30%血清を用いて、エクリズマブ、ラブリズマブ、又は BNJ430 存在 下で検討した。BNJ430 抗体は、ラブリズマブと同一のヒト IgG2/G4 Fc をもつが、マウス C5 にの み選択的に結合し、マウス血清に対しては陽性対照となるが他の動物種の血清では陰性対照とな る。これらの抗体(エクリズマブ、ラブリズマブ及びBNJ430)は検討した全ての濃度において、 ヒト以外の動物種のいずれの血清の溶血にも効果を示さなかった(Figure 4、Figure 5)。マウス血 清はcRBC の CCP 介在性溶血誘導能が低いため、ヒト C5 除去血清(25%)をマウス血清(25%) に加え、測定において確実に補体を活性化するために必要な(C5 以外の)他の補体カスケードタ ンパク質を補強した。BNJ430 は C5 分子に対して過剰なモル濃度で、溶血を効果的に阻害したが、
ラブリズマブはマウス血清に対して薬理活性を全く示さなかった(Figure 6)。 ラブリズマブは、C5 に対する抗原結合部位が約 8 倍の過剰となるようなモル濃度で、検討した いずれのヒト以外の血清に対してもin vitro で薬理活性を示さなかったことから、本試験結果より、 利用できるヒト以外の動物種のいずれに対しても、ラブリズマブはヒトの PK 又は PD 試験の動 物モデルとして適合する適切な薬理活性を示さないことが裏付けられた。 Figure 3: 遺伝子組換えヒトC5 変異体を加えた C5 除去血清のニワトリ赤血球溶血に対 するラブリズマブの阻害作用 Source:RTR-0006v1.0
Figure 4: チンパンジー、ヒヒ、アカゲザル及びカニクイザル血清の補体介在性溶血に対 する抗C5 抗体の作用
Abbreviations: Ab = antibody、cRBC = chicken red blood cell Source:RTR-0006v1.0
Figure 5: イヌ、ウサギ、モルモット及びラット血清の補体介在性溶血に対する抗C5 抗 体の作用
Abbreviations: Ab = antibody、cRBC = chicken red blood cell Source:RTR-0006v1.0
Figure 6: C5 除去ヒト血清を添加したマウス血清の補体介在性溶血に対する阻害作用
Abbreviations: Ab = antibody、cRBC = chicken red blood cell Source:RTR-0006v1.0
2.3 ヒト C5 存在下でのラブリズマブとエクリズマブの in vivo での作用の比較
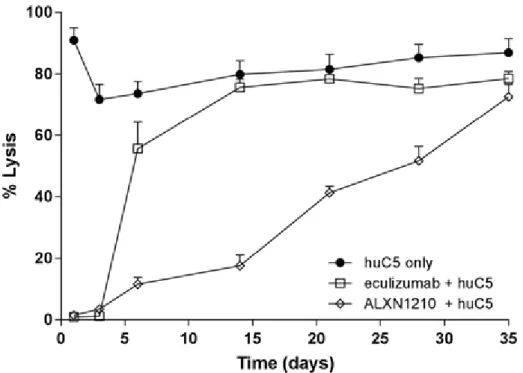
RTR-0008v1.0 試験は、ラブリズマブの抗原介在性消失、並びに PK 及び PD をエクリズマブと 比較評価するために実施した。NOD-scid マウスは遺伝的に C5 を欠損していること、及びヒト C5 又はヒト化抗体に対する免疫応答を開始できないことから、ラブリズマブ及びエクリズマブの抗原介在性消失及び対応する薬力学的効果を検討するモデル開発のために選択された。ヒト C5 の 恒常的発現による抗原介在性消失効果のモデル作成のために、マウスにDay 0 から Day 35(試験 終了日)までの間ヒトC5を皮下投与した。マウスはDay 0にラブリズマブ又はエクリズマブ100 μg を単回静脈内投与された。ラブリズマブ及びエクリズマブの血清中濃度並びにex vivo 血清溶血活 性を評価した。 C5 非存在下においてラブリズマブ及びエクリズマブは、同様の半減期(ラブリズマブでは 25.37 ± 1.02 日及びエクリズマブでは 27.65 ± 2.28 日)を示し、同様の PK プロファイルを示した。 C5 存在下においてラブリズマブは Day 28 まで抗原介在性消失をほとんど示さず、Day 35 までエ クリズマブより半減期(ラブリズマブでは13.40 ± 2.18 日及びエクリズマブでは 3.93 ± 0.54 日) を3 倍以上延長した(Figure 7)。両抗体はDay 3 まで、C5 介在性の溶血活性をほぼ完全に阻害し た。(Figure 8)。最も重要なことは、Day 14 に、エクリズマブは C5 阻害作用を示さなかったにも かかわらず、ラブリズマブは約83%の阻害を示し、Day 28 まで部分的な阻害(約 50%)を維持し たことである。 Figure 7: ヒトC5 の存在下及び非存在下でのラブリズマブ及びエクリズマブの血清中濃 度比率(%) Source:RTR-0008v1.0
Figure 8: NOD-scid マウスでのラブリズマブ又はエクリズマブの存在下及び非存在下の ヒトC5 介在性 cRBC 溶血
Abbreviations: cRBC = chicken red blood cell、NOD-scid = non-obese diabetic/severe combined immunodeficient Source:RTR-0008v1.0 ラブリズマブは、抗原介在性消失をほとんど示さず、本マウスモデルにおいてエクリズマブよ り半減期が有意に延長した。さらに、ラブリズマブの曝露時間の延長は、エクリズマブに比較し て薬力学的効果を示す期間を延長した。すなわち、ラブリズマブのY27H 及び S57H のアミノ酸 置換は、ヒトC5 存在下の NOD-scid マウスにおいて、エクリズマブより改善された PK/PD プロ ファイルをラブリズマブにもたらすことが示された。M428L 及び N434S の置換は、ヒト FcRn に 対する結合親和性を促進するよう改変されており、マウス FcRn への結合に対する効果は不明で ある。そのため、本モデルは、ラブリズマブの M428L 及び N434S の変異の PK に対する影響を 評価することを目的としていない。
3 副次的薬理試験
FcγRs は、ADCC を誘発する抗体のエフェクター機能制御において重要な役割を果たしている
(Sisto, 2009)。機能的及び構造的に異なる3 種類の FcγR すなわち、FcγRI(CD64)、 FcγRII(CD32)
及びFcγRIII(CD16)がヒト白血球に発現している。FcγRI は、高親和性受容体とされており、IgG 単量体に結合活性をもつ(Allen, 1988; Gessner, 1998)。対照的に、FcγRII 及び FcγRIII は IgG に対 して低親和性であり、これらの受容体は多重結合性免疫複合体と会合することによりもたらされ るアビディティによって活性化するのに適している(Shields, 2001)。FcγRs のライゲーションは、 FcγRI及び FcγRIIIのように活性化シグナルを産生、又は FcγRIIb のように阻害シグナルを産生し、 両者は安定した免疫応答に不可欠である(Nimmerjahn, 2005)。抗体が C1q に結合すると、CDC が開始される。 RTR-0005v1.0 試験は、ラブリズマブのエフェクター機能の有無を評価するために実施された。 ラブリズマブのキメラ Fc 領域は、CH2 及び CH3 ドメインにヒト IgG4 の定常領域を含み、エク リズマブと同一のキメラIgG2/G4 Fc 領域を有し、エクリズマブと同様にエフェクター機能を持た ないことが予測される。 ラブリズマブ及びエクリズマブの両者はヒトIgG2/G4 キメラ Fc を有しており、CH1 領域、ヒ ンジ部及びヒトIgG2 の CH2 領域の最初の 6 アミノ酸残基をヒト IgG4 の CH2 及び CH3 領域の残 りの部分と融合したものからなる。このキメラFc 領域では、安定なジスルフィド結合でペアリン グしたIgG2 はエフェクター機能を持たない IgG4 と融合している。ラブリズマブ及びエクリズマ ブは、可溶性抗原を標的としていることから、ADCC 又は CDC の開始活性を直接評価すること ができなかった。そのため、代替えとしてラブリズマブ及びエクリズマブのFcγRs 及び C1q に対 する結合を直接測定することにより、ラブリズマブ及びエクリズマブは、FcγRs 及び C1q への結 合を示さず、ADCC 又は CDC を誘発できないことが推測された。エクリズマブと同じヒト化可 変領域をもつIgG1 アイソタイプ・コントロール抗体(h5G1.1-IgG1)を対照抗体として用いた。 h5G1.1-IgG1 は、それ自体は細胞結合型抗原をもたないことから、ADCC 又は CDC を誘発するこ とは予測されないが、IgG1 アイソタイプの Fc 領域は十分にエフェクター機能分子を結合するこ とが予測された。 RTR-0005v1.0 試験において、FcγR サブクラス(FcγRI、FcγRIIa、FcγRIIb、FcγRIIb/c、FcγRIIIa 及びFcγRIIIb)の 3 抗体(ラブリズマブ、エクリズマブ及び h5G1.1-IgG1)全てとの多重結合を、 酵素結合免疫吸着測定法(ELISA)で評価し、FcγRs との単結合は SPR で評価した。バイオレイ ヤー干渉法及びSPR を用いて、3 抗体に対する C1q の結合を測定した。 ELISA で免疫複合体の抗体と FcγRs とのアビディティ主導の多重結合を検出し、ラブリズマブ 及びエクリズマブはそれぞれ、FcγRI、FcγRIIb/c、FcγRIIIa 又は FcγRIIIb に対する結合性を示さず、 FcγRIIa に対しては IgG1 アイソタイプ・コントロール抗体(h5G1.1-IgG1)より 4~8 倍弱い結合 を示した。
SPR で測定したラブリズマブ及びエクリズマブの FcγR 単量体結合に対する解離定数は、FcγRI (約4 μM)、FcγRIIa(約 2 μM)、FcγRIIb/c(約 9 μM)、FcγRIIIa(約 7 μM)及び FcγRIIIb(約 3 μM) で、FcγRs との相互作用はかなり弱く、2 抗体間でほぼ類似していることが確認された。IgG1 ア イソタイプ・コントロール抗体(h5G1.1-IgG1)の解離定数は、FcγRI(123 pM)に対して高親和 性を示し、FcγRIIa(約 1 μM)、FcγRIIb(約 3 μM)、FcγRIIIa(約 1 μM)及び FcγRIIIb(約 2 μM) に対しては、IgG2/G4 アイソタイプ抗体に比較して低親和性の FcγRs への結合を軽度に増強した。 C1q とラブリズマブ又はエクリズマブとの間に、バイオレイヤー干渉法による相互作用は検出さ
れなかった。これらの試験結果は、エクリズマブのヒト IgG2/G4 キメラ Fc 領域には、それぞれ ADCC、又は CDC を誘発する FcγRs 又は C1q を介したエフェクター機能をほとんど又は全く発現 する能力がないという仮説を支持する。さらに、ラブリズマブに組み込まれている重鎖アミノ酸 置換により、エクリズマブに比較して有意な結合変化は認められなかった。
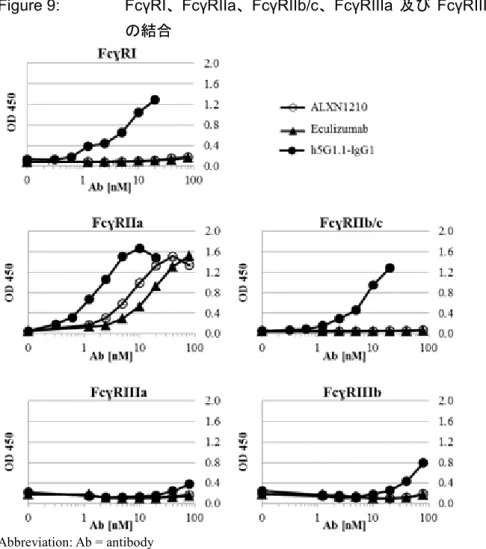
3.1 多価抗体複合体の FcγRs に対する結合
抗体(ラブリズマブ、エクリズマブ及びh5G1.1-IgG1)を、抗ヒト IgG ポリクローナル抗体 F(ab’)2 フラグメントを用いて多重結合性免疫複合体に会合し、FcγRs をマイクロプレートに高密度に結 合させた。これらの条件は、免疫複合体と低親和性受容体とのin vivo での相互作用によって生じ るアビディティ主導応答をよりよく再現するために選択された。対照抗体である IgG1 アイソタ イプ・コントロール抗体(h5G1.1-IgG1)の多価複合体は、FcγRI、FcγRIIa、FcγRIIb/c 及び FcγRIIIb に対してバックグラウンドの3~10 倍の結合性を示した(Figure 9)。ラブリズマブ及びエクリズ マブは、FcγRI、FcγRIIb/c 又は FcγRIIIb に対してバックグラウンドを超える結合シグナルを示さ ず、FcγRIIa に対しては、それぞれ、h5G1.1-IgG1 より 4~8 倍低い結合シグナルを示した。FcγRIIIa に対する結合シグナルは、3 抗体全てバックグラウンドの 2 倍より低かった。
Figure 9: FcγRI、FcγRIIa、FcγRIIb/c、FcγRIIIa 及び FcγRIIIb に対する多価抗体複合体 の結合
Abbreviation: Ab = antibody
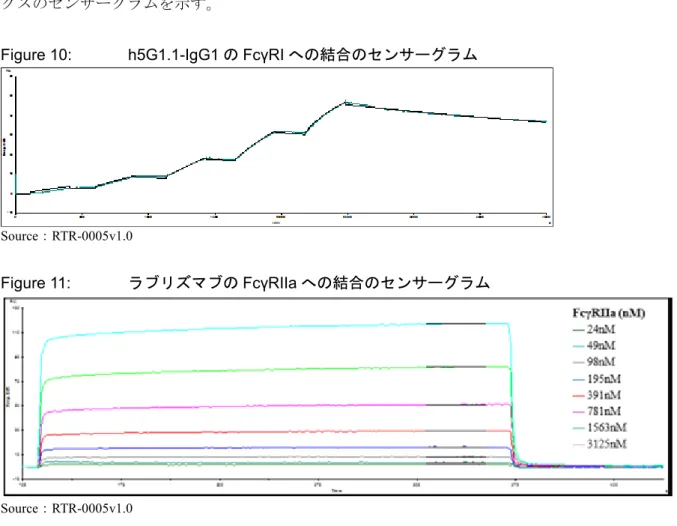
3.2 一価抗体ラブリズマブの FcγRs に対する結合
Figure 10 は、IgG1 アイソタイプ・コントロール抗体(h5G1.1-IgG1)の「高親和性」受容体で
あるFcγRI に対する結合のセンサーグラムを示す。「低親和性」受容体(FcγRIIa、FcγRIIb/c、FcγRIIIa 及び FcγRIIB)に対する抗体の結合のセンサーグラムは、迅速な結合及び解離の「箱型」プロフ ァイルを示す。Figure 11 はラブリズマブのFcγRIIa との相互作用でみられた定常状態カイネティ クスのセンサーグラムを示す。
Figure 10: h5G1.1-IgG1 の FcγRI への結合のセンサーグラム
Source:RTR-0005v1.0 Figure 11: ラブリズマブのFcγRIIa への結合のセンサーグラム Source:RTR-0005v1.0
3.3 バイオレイヤー干渉法を用いた抗体に対する C1q 結合の測定
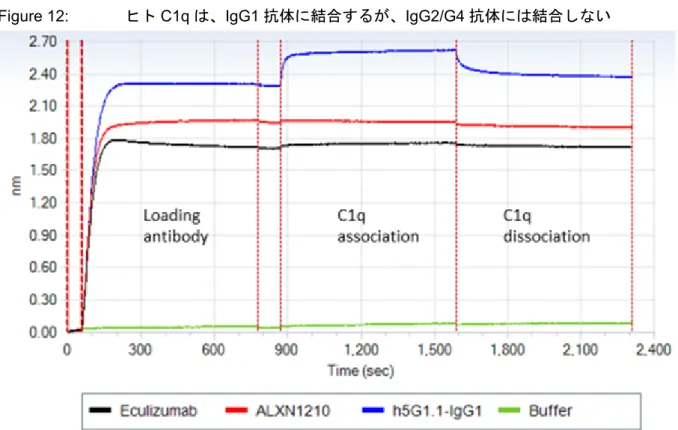
バイオレイヤー干渉法により、ラブリズマブ又はエクリズマブに CDC の開始ステップである C1q 結合は検出されなかった。Figure 12は、ビオチン化抗体[ラブリズマブ、エクリズマブ又は IgG1 アイソタイプ・コントロール抗体(h5G1.1-IgG1)]をストレプトアビジンセンサーにカップ リングし、20 nM のヒト C1q と反応させたバイオレイヤー干渉法でのセンサーグラムを示す。C1q は、h5G1.1-IgG1(青)でコートしたセンサーには結合するが、IgG2/G4 Fc[ラブリズマブ(赤) 又はエクリズマブ(黒)]抗体でコートしたセンサー又は抗体が負荷されていない[溶媒(緑)] センサーには結合しなかった。Figure 12: ヒトC1q は、IgG1 抗体に結合するが、IgG2/G4 抗体には結合しない Source:RTR-0005v1.0
3.4 結論
ELISA、SPR 及びバイオレイヤー干渉法を用いた結合試験より(Table 3 に結果をまとめた。)、 ラブリズマブに組み入れられたアミノ酸置換は、エクリズマブに比較して FcγRI、FcγRIIb/c、 FcγRIIIa、FcγRIIIb 又は C1q に対する結合親和性を変えないことが示された。FcγRIIa に対する多 価の結合を測定するためにデザインされたELISA で、ラブリズマブのシグナルがエクリズマブよ り約2 倍増加したことは、一価結合の親和性の増加を裏付けるものではない。一価の FcγRs に対 して全般的に低親和性を示したこと、多価結合の IgG1 アイソタイプに比較してシグナルが減少 したこと、C1q に対する結合が検出されなかったこと、及び抗原それ自身は可溶性であることよ り、ラブリズマブがこれらのエフェクター分子を介してADCC 又は CDC を開始する可能性は極 めて低いと考えられた。Table 3: ラブリズマブ、エクリズマブ及びh5G1.1-IgG1 の FcγRs 又は C1q との相互作 用のまとめ FcγRI (KD) FcγRIIa (KD) FcγRIIb/c (KD) FcγRIIIa (KD) FcγRIIIb (KD) C1q (KD) ラブリズマブ ELISA − ++ − − − NA SPR
Steady state kinetics 3.75 μM 2.31 μM 8.09 μM 7.23 μM 3.33 μM NA
Biolayer Interferometry −
Eculizumab
ELISA − ++ − − − NA
SPR
Steady state kinetics 3.78 μM 2.58 μM 9.84 μM 6.78 μM 3.49 μM NA
Biolayer Interferometry −
h5G1.1-IgG1
ELISA ++ +++ ++ − +
SPR
Steady state kinetics / single cycle kinetics
0.123 μM 0.80 μM 3.06 μM 0.85 μM 1.89 μM NA
Biolayer Interferometry +++
Abbreviations: ELISA = enzyme-linked immunosorbent assay、FcγRs = Fc-gamma receptors、KD = dissociation constant、
SPR = surface plasmon resonance Source:RTR-0005v1.0
4 安全性薬理試験
4.1 In vitro 安全性薬理試験
ラブリズマブに対するin vitro 安全性薬理試験は実施していない。ラブリズマブの薬理作用の標 的であるヒトC5 は、細胞の外側でのみ機能を発現することから、ヒト細胞株を用いた in vitro 評 価は適切ではないと考えられた。さらに、エクリズマブでの長期間の C5 阻害の臨床経験から中 枢神経系、心血管系又は呼吸器系への影響に対するリスクの増大は認められていない。4.2 In vivo 安全性薬理試験
ラブリズマブに対するin vivo 安全性薬理試験は実施していない。ラブリズマブは、ヒト C5 に 特異的で、他の毒性学的に関連する動物種は存在しないため、in vivo 安全性薬理試験からヒトで の毒性は予測できないと考えられた。マウス抗マウスC5 抗体(BB5.1)を用いたマウスの 26 週 間投与毒性試験で、高用量(60 mg/kg/週)の抗体を投与したマウスは全試験期間を通して完全な 補体阻害を示したが、心血管系、呼吸器系及び中枢神経系に対する毒性徴候は認められなかった [ラブリズマブ(PNH)の CTD M2.4.4.1 及び M2.6.6.3.2 を参照]。さらに、エクリズマブでの長 期間の C5 阻害の臨床経験から中枢神経系、心血管系又は呼吸器系への影響に対するリスクの増 大は認められていない。5 薬力学的薬物相互作用
薬力学的薬物相互作用試験は実施していない。ラブリズマブは、エクリズマブから改変され、 エクリズマブと同様に唯一の標的であるヒト C5 を共有する。エクリズマブに対して他の標的は 同定されていない。さらに、エクリズマブでの長期間の C5 阻害の臨床経験から中枢神経系、心 血管系又は呼吸器系への影響に対するリスクの増大は認められていない。
6 考察及び結論
ラブリズマブは、急速、完全かつ持続的な C5 阻害が得られるよう、エクリズマブを改変し半 減期を延長させたものである。ラブリズマブ及びエクリズマブは、どちらもヒト補体 C5 に対す るヒト化モノクローナル抗体であり、両者は99%以上のアミノ酸配列の相同性を共有する。ラブ リズマブは標的遺伝子の必要最小限の組換えにより誘導され、エクリズマブ重鎖の4 アミノ酸残 基が置換されている。これらの改変により、ラブリズマブの半減期がエクリズマブに比較して延 長し、終末補体阻害時間を延長すると共に、エクリズマブの有する C5 結合に対する高い特異性 を有し、且つ抗体のエフェクター機能をほとんど示さないという2 つの特性を保持するようデザ インされている。 そのため、エクリズマブと同様に、ラブリズマブの作用機序は全て、C5 への結合を介している。 C5 に結合して、補体介在性活性生成物である C5a 及び C5b[終末補体複合体(C5b-9)の開始サ ブユニット]への開裂を阻害する働きをする。非臨床試験より、これらの目的が達成されたこと が裏付けられた。 ラブリズマブのin vitro 及び in vivo での効力を裏付ける薬理作用及び副次的薬理作用を明らかに するため、多くの試験が実施された。ラブリズマブがヒト以外で検討した動物種全てと交差反応 を示さないことから、ラブリズマブを評価できるin vivo 非臨床モデルは制限されたが、標的であ るC5 に関連する毒性の有無は、マウス C5 に特異的なサロゲート抗体を用いてマウスで評価した [ラブリズマブ(PNH)の CTD M2.6.6 及び 2.6.7 参照]。 ラブリズマブのヒトC5 結合部位を決定しラブリズマブがヒト以外の動物種の C5 と交差反応す る可能性を明らかにするために、以前実施したエクリズマブの試験結果をもとにエピトープマッ ピング試験を実施した。ヒト以外の霊長類10 種及び非霊長類哺乳動物 13 種の C5 タンパク質の 配列比較により、ヒトC5 の 4 残基がラブリズマブとの結合に重要であること、4 残基のうち W917 がヒトC5 に特異的であることが明らかにされた。In vitro 結合試験及び溶血試験において、ラブ リズマブは、C5 の W917S 変異体に対して結合活性を全く示さず、ラブリズマブは、W917S 変異 体を含むin vitro 反応系の溶血を阻害しなかった。これらの結果は、検討した動物のいずれにおい ても、ヒト以外のC5 に対するラブリズマブの結合活性は、ラブリズマブの毒性、PK 及び PD を 試験するためには不十分であることを示唆した。 ラブリズマブは、エクリズマブに比してin vivo での半減期を延長するよう改変されたエクリズ マブの誘導体である。ラブリズマブは、エクリズマブの CDRs 及び重鎖定常領域にそれぞれ C5 及びFcRn との結合に対して pH 感受性を与えるアミノ酸置換をもつ。これらの置換により、抗原 介在性消失を低減し、早期エンドソームから血管コンパートメントへの抗体のリサイクル率を増 大して、その結果、エクリズマブと比較して血中半減期を延長できるようデザインされた。ラブ リズマブのin vivo 非臨床評価は、交差反応性がないことから制限を受けたが、PK 試験は、遺伝 的にC5 を欠損し、外因性のタンパク質に対して免疫応答を開始できない NOD-scid マウスモデル で実施した。ヒトC5 をマウスに投与することにより C5 の恒常的発現をシミュレートし、本モデ ルにおける抗原介在性消失に関する影響を評価した。試験結果からラブリズマブは抗原介在性消 失をほとんど示さないことが明らかとなり、本マウス PK モデルにおいてエクリズマブに比較し て有意な半減期の延長が認められた。 安定なジスルフィド結合でペアリングしたIgG2 を、エフェクター機能を持たない IgG4 と融合 させたヒト IgG2/G4 キメラ Fc 領域を含むラブリズマブのエフェクター機能を評価するために、 FcγRs 及び C1q に対するラブリズマブ又はエクリズマブの結合の直接的な測定を実施した。ラブリズマブは、C1q、FcγRI、FcγRIIb/c、FcγRIIIa 又は FcγRIIIb に対する結合性を示さず、また、FcγRIIa に対しては対照抗体であるIgG1 アイソタイプ・コントロール抗体(h5G1.1-IgG1)より約 3 倍弱 い結合が認められたことより、CDC 又は ADCC を誘発できないことが推測された。可溶性抗原 であること及びエフェクター分子に対する結合が弱いことから、ラブリズマブはin vivo で ADCC 又はCDC を誘発できないと考えられた。 エクリズマブと同様に、ラブリズマブは、C5 に結合することにより薬理作用及び完全な終末補 体阻害を飽和濃度で示す。さらにラブリズマブは、エフェクター機能を有さないと予測され、C1q 又はほとんどのFcγ 受容体に対する結合性を示さない。これら試験結果は、ラブリズマブで実施 された遺伝子改変は、ヒトでのラブリズマブの半減期及び関連する薬理活性をエクリズマブに比 較して顕著に延長するという仮説を支持する。
7 参考文献
Allen JM, Seed B. Nucleotide sequence of three cDNAs for the human high affinity Fc receptor (FcRI). Nucleic Acids Res. 1988;16(24):11824.
Brachet G, Respaud R, Arnoult C, et al. Increment in Drug Loading on an Antibody-Drug Conjugate Increases Its Binding to the Human Neonatal Fc Receptor in Vitro. Mol Pharm. 2016;13(4):1405-1412. Gessner JE, Heiken H, Tamm A, Schmidt RE. The IgG Fc receptor family. Ann Hematol. 1998;76(6):231-248.
Kabat E, Wu T, Perry H, Gottesman K. Sequences of proteins of immunological interest. No 91:3242. NIH Publication 1991.
Laursen NS, Andersen KR, Braren I, Spillner E, Sottrup-Jensen L, Andersen GR. Substrate recognition by complement convertases revealed in the C5-cobra venom factor complex. EMBO J. 2011;30(3):606-616. Nimmerjahn F, Ravetch JV. Divergent immunoglobulin g subclass activity through selective Fc receptor binding. Science. 2005;310(5753):1510-1512.
Nishimura J, Yamamoto M, Hayashi S, et al. Genetic variants in C5 and poor response to eculizumab. N Engl J Med. 2014;370(7):632-639.
Rawal N, Pangburn MK. Structure/function of C5 convertases of complement. Int Immunopharmacol. 2001;1(3):415-422.
Schatz-Jakobsen JA, Zhang Y, Johnson K, Neill A, Sheridan D, Andersen GR. Structural Basis for Eculizumab-Mediated Inhibition of the Complement Terminal Pathway. J Immunol. 2016;197(1):337-344. Shields RL, Namenuk AK, Hong K, et al. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J Biol Chem. 2001;276(9):6591-6604.
Sisto M, Lisi S, D’Amore S, D’Amore M. Autoantibodies, human Fcγ receptors, and autoimmunity. Journal of Receptor, Ligand and Channel Research. 2009;2:45-57.
TABLE OF CONTENTS
2.6.3 薬理試験概要表 ... 2 2.6.3.1 薬理試験 ... 2 2.6.3.2 効力を裏付ける試験 ... 3 2.6.3.3 副次的薬理試験 ... 4 2.6.3.4 安全性薬理試験 ... 5 2.6.3.5 薬力学的薬物相互作用試験 ... 52.6.3 薬理試験概要表
2.6.3.1 薬理試験
Overview Test Article: ALXN1210 (Ravulizumab)
Type of Study Test System Method of
Administration Testing Facility Study Number Location (eCTD)* Primary Pharmacodynamics
An in vitro assessment of the effects of pH on the kinetics of
ALXN1210 and eculizumab binding to human C5 and FcRn Surface plasmon resonance In vitro Alexion Pharmaceuticals RTR-0003v1.0 5.3.2.3.1 Epitope mapping of the ALXN1210 binding site on human C5
and predicted cross-reactivity of ALXN1210 with non-human C5
Surface plasmon resonance,
complement-mediated hemolysis assays, in
silico amino acid
sequence comparisons
In vitro Alexion
Pharmaceuticals RTR-0006v1.0 4.2.1.1.1
A comparison of single dose pharmacokinetics and pharmacodynamics of two anti-human C5 monoclonal antibodies, ALXN1210 and eculizumab, in the presence and absence of human C5 in NOD-scid mice
NOD-scid mice Intravenous Alexion
Pharmaceuticals RTR-0008v1.0 4.2.2.7.2 Secondary Pharmacodynamics
A comparative assessment of ALXN1210, eculizumab and
h5G1.1-IgG1 binding to Fc-gamma receptors and C1q in vitro ELISA, surface plasmon resonance and biolayer interferometry
In vitro Alexion
Pharmaceuticals RTR-0005v1.0 5.3.2.3.2
Safety Pharmacology No studies performed Not applicable Not applicable Not applicable Not applicable
Pharmacodynamic Drug Interactions No studies performed Not applicable Not applicable Not applicable Not applicable * Ravulizumab eCTD for PNH indication (eCTD No. 300904001)
2.6.3.2 効力を裏付ける試験
Test Article: ALXN1210 (Ravulizumab) Type of Study Species/Strain Method of
Admin. Doses (μg/kg) Gender and No. per Group
Noteworthy Findings Study Number
An in vitro assessment of the effects of pH on the kinetics of ALXN1210 and eculizumab binding to human C5 and FcRn
Human In vitro NA NA Two amino acid substitutions in the complementarity- determining regions (Y27H and S57H) decrease the affinity of ALXN1210 for human C5 by ~ 17-fold at pH 7.4 and ~36-fold at pH 6.0, compared to eculizumab. Two amino acid substitutions in the heavy chain constant region (M428L and N434S) increase the affinity of ALXN1210 for FcRn by ~10-fold compared to eculizumab. Together, these substitutions were expected to significantly attenuate target-mediated drug disposition and increase the fraction of antibody recycled from the early endosome back into the vascular compartment.
RTR-0003 v1.0
Epitope mapping of the ALXN1210 binding site on human C5 and predicted cross-reactivity of ALXN1210 with non-human C5 Human, various non-human primate, various non-primate mammalian species
In vitro NA NA Four residues in the MG7 domain of human C5,
including W917, were demonstrated to be critical for ALXN1210 binding and activity. ALXN1210 did not block in vitro hemolytic activity of sera from any animal species tested. The results suggest that ALXN1210 binding to non-human C5 is insufficient to study the toxicology, pharmacokinetics and pharmacodynamics of ALXN1210 in any animal species tested.
RTR-0006 v1.0 Comparison of single dose pharmacokinetics and pharmacodynamics of two anti-human C5 monoclonal antibodies, ALXN1210 and eculizumab, in the presence and absence of human C5 in NOD-scid mice
Mouse/NOD-scid IV for Abs, SC for hC5 250 μg of hC5 loading dose on day −1, followed by 50 μg twice daily Females;
8/group ALXN1210 had a significantly longer half-life than eculizumab in this mouse model. The extended exposure of ALXN1210 corresponded to extended duration of pharmacodynamic effect relative to eculizumab.
RTR-0008 v1.0
Abbreviations: Ab = antibody, ELISA = enzyme-linked immunosorbent assay, FcRN = Fc receptor, hC5 = human C5, IV = intravenous, NA = not applicable, NOD-scid = non-obese diabetic/severe combined immunodeficient, SC = subcutaneous
2.6.3.3 副次的薬理試験
Test Article: ALXN1210 (Ravulizumab) Type of Study Species/Strain Method of
Admin. Doses (μg/kg) Gender and No. per Group
Noteworthy Findings Study Number
A comparative assessment of ALXN1210,
eculizumab and
h5G1.1-IgG1 binding to Fc-gamma receptors and C1q in vitro
Human In vitro NA NA ALXN1210 displayed no detectable binding to FcγRI,
FcγRIIb/c, FcγRIIIa, or FcγRIIIb, and a ~3-fold weaker association with FcγRIIa than an IgG1 isotype control. In addition, no interactions between ALXN1210 and C1q were detected. The results are consistent with the hypothesis that ALXN1210 has little to no capacity to elicit effector function through FcγRs or C1q to mediate ADCC or CDC, respectively.
RTR-0005 v1.0
2.6.3.4 安全性薬理試験
安全性薬理試験は実施していない。