序 文
Streptococcus pyogenes
の菌体表層に存在する M 蛋白は主要な病原因子であり,多核白血球の食作 用に抵抗性を示し
1),抗原性の違いにより 80 種以 上の血清型(provisional type を含む)に分けられ
ている
2).この M 蛋白に対する抗体はオプソニン 抗体として型特異的感染防御の働きを示し
3),ま た特定の M 型がリウマチ熱,急性糸球体腎炎
4), 劇症型溶血性レンサ球菌感染症
5)等との関連を示 唆されており,M 型別は病因との関連や疫学解析 の上で非常に重要である.この M 型別は従来,ゲ ル内沈降反応法で行われていたが,高力価の型別 用抗血清を作製しにくいこと,市販の型別用抗血
Streptococcus pyogenes の M 蛋白遺伝子(emm )型別
1)大阪府立公衆衛生研究所微生物課,2)淀川キリスト教病院小児科,3)城東中央病院小児科
(レンサ球菌感染症研究会:会長 保科 清)
勝川 千尋
1)田丸 亜貴
1)森川 嘉郎
2)小田 公子
3)(平成 13 年 10 月 9 日受付)
(平成 13 年 12 月 28 日受理)
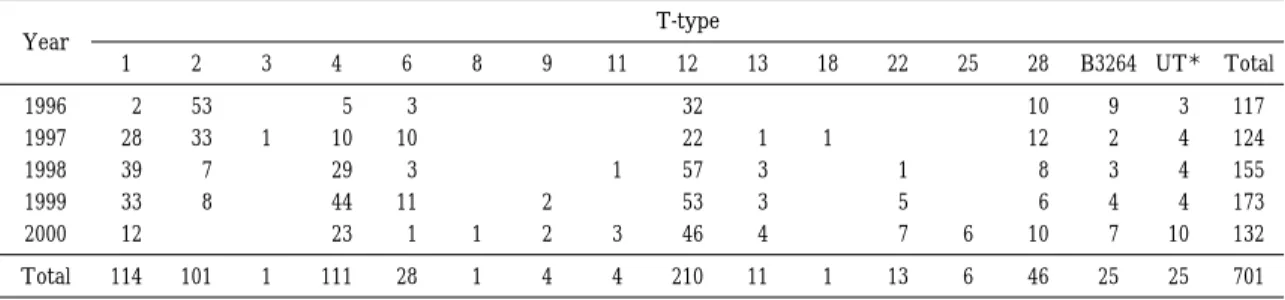
1996 年から 2000 年に大阪府立公衆衛生研究所で分離および収集したStreptococcus pyogenes701 株の 血清型別(T 型別)および M 蛋白遺伝子(emm)型別を実施し,その関連性を調査した.
T 型別では 15 種類の血清型および型別不能に分類され,分離頻度の最も高いのは T12 型,次いで T 1 型,T4 型,T2 型であった.T12 型は若干の増減はあるが,5 年間を通して分離頻度の高い血清型であっ た.これに対して T1 型,T4 型は毎年分離されたがその流行には波が見られ,T1 型は 1997 年〜1999 年にかけて,T4 型は 1998 年〜2000 年にかけて分離頻度が高かった.T2 型は 1996 年および 1997 年に多 く分離されたがその後激減し,2000 年には分離されなくなった.
分離菌株のうち 304 株についてemm型別を実施した.その結果,21 種類のemm型に分類され,T 型別では型別不能であった株も含め全ての菌株が型別可能であった.T1 型,T2 型,T4 型,T6 型,T 9 型,T11 型,T12 型,T22 型,T25 型がそれぞれ単一のemm型に決定され,T1 型:emm1,T2 型:emm 2,T4 型:emm4,T6 型:emm6,T9 型:emm9,T11 型:emm11,T12 型:emm12,T22 型:emm22,T25 型:emm75であった.複数のemm型が検出されたのが T13 型,T28 型,B3264 型,UT であり,T13 型:emm73,emm77,emm101,T28 型:emm28,emm87,TB3264 型:emm89,emm94,UT:emm2,emm12,
emm22,emm58,emm75,emm77,emm112であった.
今回われわれが実施したemm型別は多くの M 型別用抗血清を必要とする血清型別と異なり,1 ペ アーのプライマーを用いた PCR によってすべてのemm遺伝子の増幅が可能であり,増幅産物の塩基配 列を決定することにより容易に型別が行えた.このemm型別は今後レンサ球菌の型別方法として有用な 手段となっていくと考える.
〔感染症誌 76:238〜245,2002〕
要 旨
別刷請求先:(〒537―0025)大阪市東成区中道 1―3―69 大阪府立公衆衛生研究所微生物課
勝川 千尋
Key words: Streptococcus pyogenes, T-typing, M protein gene(emm)typing
清がないことなどの理由で一般にはあまり実施さ れなかった.しかし,近年 M 蛋白に関しては多く の型についてその遺伝子がクローニングされ,塩 基配列が決定されている.そのデータは Centers for Disease Control and Prevention(CDC)にデー タベースとして蓄積,整理され,公開されている ので
6),菌の M 蛋白遺伝子(emm)のシークエンス を行い,そのデータと比較することにより遺伝子 レベルでの M 型別(emm 型別)が可能になった.
われわれは今回 1996 年から 2000 年に大阪府立公 衆衛生研究所で分離および収集した
S. pyogenesの血清型別(T 型別)および
emm型別を実施し,
その関連性を調査したので報告する.
材料と方法
1.試験菌株
1996 年から 2000 年に大阪府立公衆衛生研究所 で分離および収集した
S. pyogenes701 株を供 試 した.その内訳は,劇症型溶血性レンサ球菌感染 症(以下 TSLS)患者由来株 10 株,その他の溶血 性レンサ球菌感染症患者由来株 691 株であった.
その詳細を以下に示す.
1)TSLS 患者由来株
TSLS の診断基準を満たす患者 10 名から分離 された菌株で, 分離材料は血液 4, 腹水 1, 壊死部組 織 1, 関 節 液 1, 正 常 な ら ば 無 菌 部 詳 細 不 明 3 で あった.患者年齢は小児 3 名(3, 5, 6 歳) ,成人 6 名(32, 35, 42, 53, 56, 72 歳),不明 1 名であり,医 療機関受診場所は大阪府:4 名, 近県:6 名であっ た.
2)その他の溶血性レンサ球菌感染症患者由来 株
小児科を受診した
S. pyogenes感染を疑われる 上気道炎患者および発疹を伴う溶連菌感染症患者 の咽頭材料(咽頭ぬぐい液,うがい液)から分離 された菌株 663 株,TSLS の診断基準は満たさな いが重症化した溶連菌感染症患者の正常ならば無 菌部(血液,膿汁,腹水,尿,皮膚病巣各 1)から 分離された菌株 5 株,皮膚感染患者の皮膚病巣か ら分離された菌株 8 株,膣炎患者の膣分泌物から 分離された菌株 13 株, 耳炎患者の耳漏膿から分離 された菌株 1 株,溶連菌感染症患者から分離され
検体の由来不明菌株 1 株であった.患者の医療機 関受診場所別にみると大阪府:665 株(市内 E 病 院:402 株,J 病院:128 株,Y 病院:86 株,府下 M 病院:34 株,S 病院:11 株,P 病院:2 株,N 病院:1 株,K 病院:1 株) ,近県:26 株であった.
2.菌の同定
トリプチケースソイヒツジ血液寒天培地上で大 きな
β溶血を示し,カタラーゼ陰性,グラム陽性 球菌であることを確認後,血清群別キットを用い て A 群抗原を保有することおよびピロリドニル アリルアミダーゼ活性試験(以下 PYR 試験)陽性 であることが確認できた菌を
S. pyogenesと同定 した.
3.T 型別
T 蛋白による型別は市販の T 型別用免疫血清
(デンカ生研) を用いてスライド凝集反応法で実施 した.抗原はデンカ生研の操作法に従いブタ膵エ キス(デンカ生研)処理により作製した.
4.M 蛋白遺伝子(emm)のシークエンスによる
emm型別
型別は Beall ら
7)の方法に準じて
emm遺伝子を PCR で増幅, その増幅産物の 5 末端側の塩基配列 を決定し,CDC のデータベースを利用して型別す る手法を用いた.以下にその詳細を示す.
1)PCR
トリプチケースソイヒツジ血液寒天培地上の 4
〜5 コロニーを TE バッファー (10mM Tris-HCl,
1mM EDTA,pH 8.0) 100
µl に懸濁,ヒートブロッ クで 95℃10 分加熱後, その遠心上清をテンプレー ト DNA とした.
プライマーは各型の
emm遺伝子に共通な塩基 配列が存在するシグナルペプチド領域および保存 領域に設定した以下のものを用いた.
emm-1:5 -TAT T(C!
G)G CTT AGA AAA TTA A-3
emm-2:5 -GCA AGT TCT TCA GCT TGT TT-3
反応液中のプライマーの濃度は 1
µM,増幅酵素 は
TaKaRa LA Taq(宝酒造)を用い,上記のテンプ レート DNA を 0.5
µl 加え,全量を 25
µl とした.増 幅プログラムは 94℃5 分の前熱変性後,熱変性 94
℃30 秒,アニーリング 55℃1 分,伸張反応 72℃2
Table 1 Annual changes of T-type distribution of Streptococcus pyogenes isolated from 1996 to 2000
T-type Year
Total UT * B3264 28 25 22 18 13 12 11 9 8 6 4 3 2 1
117 3 9 10 32
3 5 53
2 1996
124 4 2 12 1
1 22 10
10 1 33 28 1997
155 4 3 8 1
3 57 1 3
29 7
39 1998
173 4 4 6 5
3 53 2
11 44 8
33 1999
132 10 7 10 6 7 4
46 3 2 1 1 23 12
2000
701 25 25 46 6 13 1 11 210 4 4 1 28 111 1 101 114 Total
* : T-untypable
分を 30 サイクル繰り返し,最終伸張反応を 72℃
10 分行った.PCR 産物のうち 5
µl を 1% アガロー スによる電気泳動で
emm遺伝子の増幅を確認, そ の後残り 20
µl を同様に電気泳動,エチジウムブロ マイドで染色し,長波長の紫外線照射のもとで増 幅されたバンドを切り出した.切り出したアガ ロースブロックをサンプルチューブに入れ,−20
℃,overnight または−80℃,30 分放置後,遠心分 離し,その上清をシークエンス用テンプレート DNA とした.DNA 溶液の濃度は 5〜20ng
!µl に 調整し,濃度が低い場合はこの DNA 溶液をテン プレート DNA とし再度 PCR を行って増幅した.
2)シークエンス
シークエンスは ABI PRISM 310(Applied Bio- systems)を用い,Dye Terminator 法で行った.
シークエンス試料作製のプライマーは M 蛋白の N 末の Hypervariable な領域のシークエンスを調 べるためシグナルペプチド領域に設定した以下の プライマーを用いた.
emmseq:5 -TAT TCG CTT AGA AAA TTA AAA ACA GG-3
試料の作製は BigDye Terminator Cycle Seque- ncing FS Ready Reaction Kit ( Applied Biosys- tems)を用い,全量を 4 分の 1 にして行 っ た.
emmseq プライマーを用いてシークエンスがうま く行えない場合は PCR に用 い た emm-1 プ ラ イ マーを用いてシークエンス反応を行った.反応後 の試料の精製は冷却しないエタノール沈殿法で 行った.
3)emm 塩基配列の解析
既報
8)の種々の M 蛋白遺伝子のリーダーシーク エンスの比較を参照し,シークエンスプライマー の 3 末 端 か ら 5 番 目 の 塩 基 と 推 測 さ れ る 位 置
(Hydrophobic region の最初)から 160bp を正確 に シ ー ク エ ン ス を 行 い,そ の デ ー タ を CDC の
Streptococcus pyogenesDatabase(http:
!!www.cdc.
gov
!ncidod
!biotech
!strep
!strepblast.html)へ 送 付,解析結果を電子メールで受け取った.emm 型は Beall ら
7)の定義に従い,95% 以上の相同性を 持つことを確認し決定した.
成 績
1996 年から 2000 年の 5 年間に分離,収集した
701 株の T 型別成績を Table 1 に示した.15 種類
の血清型および型別不能に分類され,分離頻度の
最も高いのは T12 型,次いで T1 型,T4 型,T2
型であった.T12 型は若干の増減はあるが,5 年間
を通して分離頻度の高い血清型であった.これに
対して T1 型,T4 型は毎年分離されたがその流行
には波が見られ,T1 型は 1997 年〜1999 年にかけ
て,T4 型は 1998 年〜2000 年にかけて分離頻度が
高かった. T2 型は 1996 年および 1997 年に多く分
離されたがその後激減し,2000 年には分離されな
くなった.この 4 血清型以外では,T6 型,T28
型,TB3264 型が数は少ないが毎年分離された.T
型別不能(以下 UT)は 1996 年〜1999 年は全体の
2〜3% であったが,2000 年 に は 7.6% に 増 加 し
た.劇症型溶血性レンサ球菌感染症患者由来株は
1998 年:T1 型 1 株,T12 型 1 株,1999 年:T1
型 5 株,T12 型 1 株,T22 型 1 株,2000 年:T1
型 1 株であった.
Table 2a emm types associated with T-types
emm type(No. of isolates)
No. of isolates examined T type
emm1(35)
35 T-1
emm2(25)
25 T-2
emm80(1)
1 T-3
emm4(40)
40 T-4
emm6(28)
28 T-6
emm76(1)
1 T-8
emm9(4)
4 T-9
emm11(4)
4 T-11
emm12(39)
39 T-12
emm73(5),emm77(5),emm101(1)
11 T-13
emm18(1)
1 T-18
emm22(13)
13 T-22
emm75(6)
6 T-25
emm28(45) emm87(1)
46 T-28
emm89(8) emm94(17)
25 T-B3264
emm2(1),emm12(1),emm22(1), emm58(10),emm75(2),emm77(8), emm112(2)
25 UT*
* : T-untypable
Table 2b T-types associated with emm types
T type(No. of isolates)
No. of isolates emm type
T-1(35)
emm1 35
T-2(25),UT*(1)
emm2 26
T-4(40)
emm4 40
T-6(28)
emm6 28
T-9(4)
emm9 4
T-11(4)
emm11 4
T-12(39),UT*(1)
emm12 40
T-18(1)
emm18 1
T-22(13),UT*(1)
emm22 14
T-28(45)
emm28 45
UT*(10)
emm58 10
T-13(5)
emm73 5
T-25(6),UT*(2)
emm75 8
T-8(1)
emm76 1
T-13(5),UT*(8)
emm77 13
T-3(1)
emm80 1
T-28(1)
emm87 1
T-B3264(8)
emm89 8
T-B3264(17)
emm94 17
T-13(1)
emm101 1
UT*(2)
emm112 2
* : T-untypable
emm
型別は分離菌株数の少ない T3 型,T6 型,
T8 型,T9 型,T11 型,T13 型,T18 型,T22
型,T25 型,T28 型,TB3264 型,UT については,
計 165 株全株型別を行った.分離菌株数の多い T 12 型は 1996 年:32 株中 4 株,1997 年:22 株中 3 株, 1998 年:57 株中 12 株, 1999 年:53 株中 6 株,
2000 年:46 株 中 14 株,計 210 株 中 39 株(18.6
%) ,T1 型 は 1996 年:2 株 中 2 株,1997 年:28 株中 3 株,1998 年:39 株中 6 株,1999 年:33 株 中 12 株,2000 年:12 株 中 12 株,計 114 株 中 35 株(30.7%) ,T4 型は 1996 年:5 株中 5 株,1997 年:10 株中 2 株, 1998 年:29 株中 5 株, 1999 年:
44 株中 5 株,2000 年:23 株中 23 株,計 111 株中 40 株(36.0%) ,T2 型 は 1996 年:53 株 中 6 株,
1997 年:33 株中 4 株,1998 年:7 株中 7 株,1999 年:8 株中 8 株,計 101 株中 25 株(24.8%)につい て型別を行った.合計 304 株の型別を実施した結 果,21 種類の
emm型に分類され,T 型別では型別 不能であった株も含め全ての菌株が型別可能で あった.その型別成績を Table 2a および Table 2b に示した.Table 2a は T 型別 に 集 計 し た が,T 型別で複数菌株分離された菌型の中では T1 型, T 2 型,T4 型,T6 型,T9 型,T11 型,T12 型,T 22 型,T25 型がそれぞれ単一の
emm型に決定さ れ,T1 型:emm1,T2 型:emm2,T4 型:emm4,T 6 型:emm6,T9 型:emm9,T11 型:emm11,T12 型:emm12,T22 型:emm22 ,T25 型:emm75 で あった.複数の
emm型が検出されたのが T13 型,
T28 型,TB3264 型,UT であり,T13 型:emm73,
emm 77
,
emm 101,T 28 型 :
emm 28,
emm 87,TB 3264 型 :
emm89,
emm94,UT:
emm 2,
emm 12,
emm22,emm58,emm75,emm77,emm112で あ っ た.Table 2b は
emm型別に集計した.
emm2,emm 12,emm22,emm75,emm77で は 1 種 類 の T 型 お よび UT が含まれていたが,2 種の異なる T 型が 検出された
emm型はなかった.
1 つの T 型で異なる
emm型が検出された T13
型,T28 型,TB3264 型,UT について,
emm型の
年次変化を調べ,その集計結果を Table 3 に示し
た.T13 型は 1997 年から 1998 年にかけて分離さ
れ た 菌 は 全 て
emm77で あ っ た が,1999 年 か ら
emm73が分離されるようになり 2000 年はすべて
emm73になった.T28 型は 1996 年から 1999 年ま
Table 3 Annual changes of emm types of Streptococ- cus pyogenes T-13, T-28, T-B3264, T-untypable isolates from 1996 to 2000
UT* T-B3264
T-28 T-13
Year
emm58(2)
emm2(1)
emm94(9)
emm28(10)
1996
emm58(2)
emm75(1)
emm77(1)
emm89(1)
emm94(1)
emm28(12)
emm77(1)
1997
emm12(1)
emm58(1)
emm77(1)
emm112(1)
emm94(3)
emm28(8)
emm77(3)
1998
emm58(2)
emm77(1)
emm112(1)
emm94(4)
emm28(6)
emm73(1)
emm77(1)
emm101(1)
1999
emm22(1)
emm58(3)
emm75(1)
emm77(5)
emm89(7)
emm28(9)
emm87(1)
emm73(4)
2000
* : T-untypable
ではすべて
emm28であったが,2000 年に
emm87が 1 株分離され た.TB3264 型 は 1999 年 ま で は 1997 年 の 1 株 を 除 き
emm94で あ っ た が,2000 年はすべて
emm89になった.UT は 5 年間を通し て
emm58および
emm77が多く分離され,特に目 立った年次変化は認められなかった.
考 察
M 型別は Swift ら
9)によって毛細管沈降反応法 により始められた.その後,多くの血清型が発見 され,Johnson ら
10)は 1993 年に M81 までの型,約 74 種の血清型があることを報告した.その後も新 しい血清型が確認され現在では provisional type を含めて 86 種の型があるとされている
2).この M 型別は現在も一般にはゲル内沈降反応法で行 われているが,高力価の型別用免疫血清を作製し にくいこと,市販の型別用抗血清がないことなど の理由で一般には普及していない.分離頻度の高 い 1, 3, 4, 6, 12 型の抗血清を準備しておけば 50〜
60% 以上の菌株について型別が行えるが型別不 能菌も多い
11)12).村井ら
13)は 29 種類の M 型別 血 清を用いて 1975 年 5 月から 2000 年 2 月までに松 山市内で分離された
S. pyogenes4,195 株について 型別を行ったが,19.9% の菌が型別不能であった と報告している.さらに多くの
S. pyogenesについ
て M 型を決定するためには,より多くの種類の抗 血清を準備しておく必要があるが,これは現状で は非常に困難である.しかし,M 蛋白に関しては 近年多くの型についてその遺伝子がクローニング され,その塩基配列はデータベースとして存在す る.データベースは CDC より公開されている
6)の で, M 蛋白遺伝子 (emm) のシークエンスを行い,
蓄積されたデータと比較することにより遺伝子レ ベルでの
emm型別が可能になった.
成熟 M 蛋白は N 末より Hypervariable region,
Variable region,Conserved region に分けられ,N 末の特異性が高く,C 末は各型に共通なアミノ酸 配列が存在する構造になっている
14).emm 遺伝子 を PCR で 増 幅 す る 場 合,N 末 の Hypervariable region には各型に共通な塩基配列は存在しない のでこの領域にプライマーは設定できない.しか し M 蛋白は菌体外に突出した蛋白であり,菌体内 で作られたときは M 蛋白の細胞膜通過に重要な 役割を果たすシグナルベプチドが結合している.
このシグナルペプチドは異なる M 型でもアミノ 酸配列が非常に類似している.シグナルペプチド は N 末か ら Basic region,Hydrophobic region,
Cleavage region に分けられるがその中で も Ba- sic region は各型に共通な塩基配列が存在する
8). 従ってシグナルペプチドの Basic region お よ び M 蛋白の Conserved region にプライマーを設定 することにより,多くの M 型の
emm遺伝子の増 幅が可能となる
7).
このプライマーによって増幅された PCR 産物 について,5 末端側のシークエンスを行うと,約 60bp のリーダーシークエンスとそれに続く特異 性の高い Hypervariable region の塩基配列を決定 することが出来る.Beall ら
7)はこの
emm遺伝子の 5 末端側の塩基配列と血清学的 M 型が高い相関 を示すことを示し,5 末端側の 160bp を決定する ことにより,
emm型別を行えることを報告した.
今回われわれは Beall ら
7)の方法に準じて
emm型 別を行ったところ,T 型別では型別不能であった 菌も含め実施した 304 株すべてが型別可能であっ た.
M 蛋白分子の Hypervariable region および Va-
riable region のアミノ酸配列は異なる M 型間で は相同性がほとんどないか非常に低く,M 蛋白分 子のサイズも異なることが知られている
15).また 同一 M 型間でも DNA の組換え,複製の過程での 変異に基づくと推測される M 蛋白分子のサイズ の変異が観察されている
14).そのため M 蛋白分 子には型特異的なサイズはなく,
emm遺伝子の増 幅を行った場合もその PCR 産物のサイズから M 型を推測することはできない.今回供試した菌株 についても,1Kb 前後の PCR 産物が増幅されて きたが, 同一
emm型であってもサイズの異なる菌 株が存在した.
また,S. pyogenes の菌体表層に存在し,M 蛋白 と類似構造を持つ M 蛋白様蛋白遺伝子(M-like protein 遺伝子,以下
emml遺伝子)
16)が
emm遺伝 子と同時に増幅されてきた菌株も存在したが,今 回実施した条件では
emml遺伝子は増幅効率が悪 いため,アガロース電気泳動後,エチジウムブロ マイド染色を行うと,
emm遺伝子のバンドが濃く 染色されるのに対して,emml 遺伝子のバンドは 薄く,明確に判別できた.最も濃く明瞭なバンド をアガロースから切り出し,シークエンスを行う ことで
emm型別が実施できた.
今回検査した T1 型,T4 型,T6 型,T12 型菌は そ れ ぞ れ
emm1,emm4,emm6,emm12に 型 別 さ れ,T 型別からみた場合の一致率は 100% であっ た.これらの血清型はわれわれが 1988 年および 1989 年に分離した
S. pyogenesについて実施した M 型別用抗血清を用いた型別
12)でもそれぞれ M1 型,M4 型,M6 型,M12 型に型別される確率が高 い菌型であった.しかし一致率は 100% でなくそ れぞれ 98.1%,89.4%,91.7%,94.3% であった.
Beall ら
2)7)17)18)は世界各国から収集した
S. pyogenesの T 型と M 型の関連性について,T1 型は
emm1以外に
emm28,emm68,T4 型は
emm4以外に
emm 22,emm46,
emm48,
emm58,
emm60,
emm63,
emm 81,pt2841,st213,st4936,T6 型 は
emm6以 外 に
emm70,
emm75,
st64!14,st3018,
st4935,
st448,T 12 型は
emm12 以外に
emm22,
emm59,
emm62,
emm66,emm72,emm76,
pt4245,
st2967,stns14xに 型別される菌株が存在したことを報告しており
(T 型を complex として型別した菌株を除く) , 1988 年および 1989 年に 分 離 し た
S. pyogenesT1 型,T4 型,T6 型,T12 型菌の中に
emm1,emm4,emm6,emm12
以外の型が存在した可能性も考え られる.また,M 型別用抗血清を用いた型別では 沈降反応用抗原の抽出に用いる菌量が少ない場合 には反応が認められず, 菌量を 10 倍にすることに よって型別できた菌株があった
12)ことから,M 蛋 白の産生量に菌株による差が認められ,産生量の 少ない菌株は型別できなかった可能性もある.ま た遺伝子を保有するにも関わらず実際に M 蛋白 を産生しない菌株があることも否定できない.
これらの 4 菌型以外でも T2 型菌が
emm2に,
T9 型菌が
emm9に,T11 型菌が
emm11に,T22 型 菌 が
emm22に,T25 型 菌 が
emm75に 型 別 さ れ,それぞれ 1 種類の
emm型に型別された.大阪 近辺という比較的狭い地域の分離菌株について調 べたことおよび 1996 年から 2000 年の 5 年間とい う短い期間に分離された菌株であったことが関連 しているのか, 今回
emm型別を行った菌株に関し ては 1 つの T 型は 1 種類の
emm型に型別される 傾向にあった.
T13 型,TB3264 型,T28 型に関しては複数の
emm型が検出され,T13 型は 3 種類の
emm型に,
TB3264 型および T28 型は 2 種類の
emm型に型 別された.T13 型および TB3264 型では 5 年の間 に
emm型が変化してきており, T 型のみでは判明 しなかった菌型の変化を観察することができた.
溶血レンサ球菌レファレンスセンターでは劇症 型 A 群溶血性レンサ球菌感染症患者から分離さ れたレンサ球菌を全国的に収集し菌の解析を行っ ており,T3 型の
emm型に関しては 1992 年から 1995 年にかけて分離された T3 型 10 株はすべて
emm3に型別されたが,1996 年以降分離された T 3 型 5 株は
emm3(2 株),emm11(1 株) ,emm81
(2 株)に型別されたと報告している
19).今回われ
われが分離した T3 型の
emm型は
emm80であり
溶血レンサ球菌レファレンスセンターの報告して
いる
emm型とも異なる.また Beall ら
18)も T3
!13
!B3264 型菌(complex として型別)は多くの
emm型が検出されることを報告しており, T3 型は T13
型,TB3264 型とともに複数の
emm型が検出され る可能性の高い菌型であるといえる.
UT 25 株の中にはすべての型別用免疫血清に陰 性のものが 24 株,T2 血清または T28 血清に陽性 を示すが結果は血清のロットにより異なり一定し なかったため型別不能としたものが 1 株あった.
前 者 は
emm12:1 株,emm22:1 株,emm58:10株,
emm75:2 株,emm77:8 株,emm112:2 株であったが,これらの菌株が T 蛋白を保有しないの か,現在市販されている抗血清とは反応しない新 しい型であるのか,T 蛋白の産生量が低下し凝集 反応では検査が不可能であったのか推測すること は難しい.後者は
emm2と型別されたが,T 型別 では, T2 型の異なる 4 ロットの型別血清を用いて 試験を行ったところそのうち 1 ロットの型別血清 に陽性反応を示し,T28 型の異なる 5 ロットの型 別血清を用いて試験を行った場合はそのうちの 2 ロットの型別血清に陽性反応を示したため UT と した.市販の T28 型別血清は T28 蛋白および R28 蛋白を保有する菌株を用いて免役し,R28 蛋白に 対する抗体は吸収されずに作製されている.その ため,R 蛋白を保有するレンサ球菌が T 型とは関 係なく凝集反応を示す場合もあり,以前から問題 のある型別血清とされてきた
20).今回の菌株につ いて R 蛋白の保有の有無は調べることができな かったが,このような T 型別で判定が困難な菌株 も
emm型別では確実に型別することができた.
今回われわれが実施した
emm型別は多くの M 型別用抗血清を必要とする血清型別と異なり,1 ペアーのプライマーを用いた PCR によってすべ ての
S. pyogenesの
emm遺伝子の増幅が可能であ り,増幅産物の塩基配列を決定することにより容 易に型別が行えた.この
emm型別は血清 M 型別 との一致率も高く,さらに C 群,G 群,L 群の保有 する
emm遺伝子も順次データベースに追加され ており,今後レンサ球菌の型別方法として有用な 手段となっていくと考える.
謝辞:本研究に多大なるご協力をいただきました掖済 会病院小児科大森善也先生,板金康子先生,坂崎弘美先生 に深謝いたします.
文 献
1)Lancefield RC:Current knowledge of type-speci- fic M antigens of group A streptococci. J Immu- nol 1962;89:307―13.
2)Beall B, Gherardi G , Lovgren M , Facklam RR , Forwick BA, Tyrrell GJ:emmandsof gene se- quence variation in relation to serological typing of opacity-factor-positive group A streptococci . Microbiology 2000;146:1195―209.
3)Rothbard S:Bacteriostatic effect of human sera on group A streptococci. I. Type-specific antibod- ies in sera of patients convalescing from group A streptococcal pharyngitis. J Exp Med 1945;82:
93―105.
4)Bisno AL:Group A streptococcal infections and acute rheumatic fever. N Eng J Med 1991;325:
783―93.
5)五十嵐英夫, 柏木義勝, 遠藤美代子, 奥野ルミ,
榎田隆一:劇症型 A 群レンサ球菌感染症の疫学.
渡辺治雄,清水可方監修,劇症型 A 群レンサ球菌 感染症,近代出版,東京,1997:p. 96―113.
6) Centers for Disease Control and Prevention : Streptococcus pyogenesdatabase. http:!!www.cdc.
gov!ncidod!biotech!strep!strepindex.html.
7)Beall B, Facklam R, Thompson T:Sequencing emm-specific PCR products for routine and accu- rate typing of group A streptococci. J Clin Micro- biol 1996;34:953―8.
8)Katsukawa C:Cloning and nucleotide sequence of type 3 M protein gene(emm3)consisting of an N-terminal variable portion and C-terminal con- served C repeat regions:relation to other genes ofStreptococcus pyogenes. Kansenshogaku Zasshi 1994;68:698―705.
9)Swift HF, Wilson AT, Lancefield RC:Typing of group A hemolytic streptococci by M precipitin reactions in capillary pipettes. J Exp Med 1943;
78:127―33.
10)Johnson DR, Kaplan EL:A review of the correla- tion of T-agglutination patterns and M-protein typing and opacity factor production in the identi- fication of group A streptococci. J Med Microbiol 1993;38:311―5.
11)中島邦夫:A 群溶血レンサ球菌の群別と型別.臨 床と微生物 1988;15:22―7.
12)勝川千尋,原田七寛:大阪府下で分離された A 群溶血レンサ球菌の血清型と薬剤感受性につい て(1988〜1989 年).感染症誌 1991;656:945―
52.
13)村井貞子,石井恵子,徳丸 実:松山市内の一小
児科クリニックにおける溶レン菌感染症の長期
観察.レンサ球菌感染症研究会第 33 回学術講演 会抄録集,2000.
14)Fischetti VA:Streptococcal M protein:Molecu- lar design and biological behavior. Clin Micro Rev 1989;2:285―314.
15)岡田信彦:化膿連鎖球菌のビルレンス因子,M 蛋白の役割とその発現機構.日本細菌学雑誌 1996;51:571―81.
16) Whatmore AM , Kehoe MA : Horizontal gene transfer in the evolution of group A streptococcal emm-like genes:gene mosaics and variation in Vir regulons. Mol Microbiol 1994;11:363―74.
17)Beall B, Facklam R, Hoenes T, Schwartz B:Sur- vey ofemmgene sequences and T-antigen types from systemicStreptococcus pyogenesinfection iso-
lates collected in San Francisco, California;At- lanta, Georgia;and Connecticut in 1994 and 1995.
J Clin Microbiol 1997;35:1231―5.
18)Beall B , Facklam RR , Elliott JA , Franklin AR , Hoenes T, Jackson D,et al.:Streptococcalemm types associated with T-agglutination types and the use of conservedemmgene restriction frag- ment patterns for subtyping group A strepto- cocci. J Med Microbiol 1998;47:893―8.
19)溶血レンサ球菌レファレンスセンター.2001.第 22 回衛生微生物協議会溶血レンサ球菌レファレ ンスセンター報告書.
20)溶血レンサ球菌レファレンスセンター.1997.第 18 回衛生微生物協議会溶血レンサ球菌レファレ ンスセンター会議.
M Protein Gene(emm)Typing of
Streptococcus pyogenesChihiro KATSUKAWA, Aki TAMARU, Yoshiro MORIKAWA & Kimiko ODA
1)Department of Microbiology, Osaka Prefectural Institute of Public Health
2)Department of Pediatrics, Yodogawa Christian Hospital
3)Department of Pediatrics, Joto-Chuo Hospital