Ⅰ.はじめに T 細胞,B 細胞はそれぞれ,T 細胞受容体(TCR) およびB 細胞受容体(BCR)遺伝子のランダムな 組み換えによって多様性を得ている.そのため一定 の確率で自己反応性T 細胞,B 細胞が産生される が,免疫寛容として知られるいくつかのメカニズム によってほとんどが排除ないし抑制されることで, 身体は自己免疫疾患から守られている1).このうち 特に重要と考えられているのが胸腺における中枢性 免疫寛容と,末梢において制御性T 細胞によって 制御される末梢性免疫寛容である.昨年の本誌に中 枢性免疫寛容と末梢性免疫寛容の関連を論じる優 れた総説が掲載されたが2),本稿では,中枢性免疫 寛容の異常によって生じるAutoimmune polyendocr-inopathy, candidiasis, ectodermal dystrophy(APECED)
と末梢性免疫寛容の異常によって生じる
Immuned-ysregulation, polyendocrinopathy, enteropathy, X-linked 総 説
中枢性免疫寛容不全と末梢性免疫寛容不全における標的抗原特異性
千田奈津子
*1,小 林 一 郎
*2Difference in target antigens between central tolerance and peripheral tolerance deficiencies
Natsuko Chida*1 and Ichiro Kobayashi*2*1Department of Dentistry for Children and Disabled Persons,
Hokkaido University Graduate School of Dental Medicine, Sapporo, Japan
*2Department of Pediatrics, Hokkaido University Graduate School of Medicine, Sapporo, Japan (Accepted April 1, 2015)
summary
Failure of the immunotolerance mechanisms causes multiple organ-specific autoimmune disorders. Mutations of autoim-mune regulator (AIRE) gene result in central immunotolerance deficiency named autoimautoim-mune polyendocrinopathy, candidiasis, ectodermal dystrophy (APECED). Mutations of FOXP3 genes cause regulatory T cell (Treg) deficiency named immune dys-regulation, polyendocrinopathy, enteropathy, X-linked (IPEX) syndrome. Because T cell tolerance influences B cell tolerance, autoantibodies seem to reflect the presence of autoreactive T cells with the same antigen specificity. To date many differences in both clinical features and autoantibody profiles have been described between APECED and IPEX syndrome. In addition to the differences in target organs, we have found differences in the target antigens in the same organ, small intestine, between both disorders; anti-autoimmune enteropathy-related 75 kDa antigen (AIE-75) antibodies are specific to IPEX syndrome, whereas anti-tryptophan hydroxylase-1 (TPH-1) antibodies are specific to APECED. These facts suggest that immunotolerance to AIE-75 depends on the Treg, whereas the tolerance to TPH-1 depends on the central mechanisms. Furthermore, given the earlier onset and more serious clinical features of IPEX syndrome than APECED, physiological roles of Aire on the selection of Treg may be, if present, limited.
Key words APECED; IPEX syndrome; Aire; FoxP3; tissue-specific antigen 抄 録
中枢性免疫寛容の異常によって生じるAutoimmune polyendocrinopathy, candidiasis, ectodermal dystrophy(APECED) と末梢性免疫寛容の異常によって生じるImmunedysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX 症候群)は共に多彩な自己免疫疾患を合併する.いずれも T 細胞免疫寛容不全であるが,B 細胞レベルの免
疫寛容にも大きな影響をもたらすことから,自己抗体の標的抗原特異性はT 細胞レベルでの自己反応性を反映す
ると考えられる.興味深いことに,APECED と IPEX 症候群では好発罹患臓器に違いが見られる.また,同じ小腸
であってもAPECED ではトリプトファン水酸化酵素,IPEX 症候群では自己免疫性腸症関連 75 kDa 抗原(AIE-75)
に対する自己抗体が疾患特異的である.こうした事実は,抗原により中枢性ないし末梢性免疫寛容への依存度が異
なり,またTreg の分化に対する Aire の役割は,もしあっても限局したものである事を示唆する.
*1 北海道大学大学院歯学研究科小児・障害者歯科学
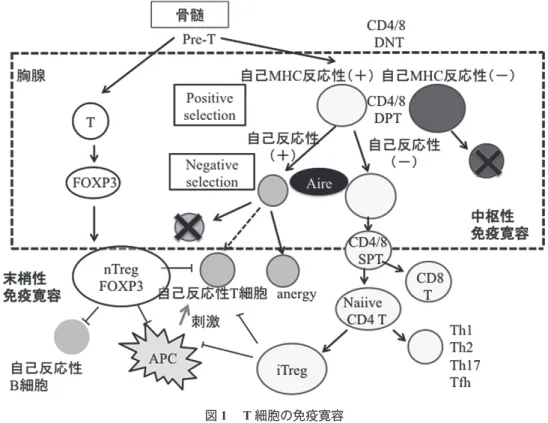
syndrome(IPEX 症候群)の臨床像と自己抗体の検 討から,2 つの免疫寛容機序の抗原特異性を論じる. Ⅱ.T 細胞および B 細胞の免疫寛容機構 1.Autoimmune regulator(Aire)と中枢性免疫寛容 ネガティブセレクションは胸腺髄質において自己 反応性T 細胞を除去するメカニズム(中枢性免疫 寛容)であるが,そのためには胸腺において本来他 の組織に特異的に発現すべき自己抗原(組織特異性 抗原;TSAs)が発現される必要がある.自己免疫 調節因子Aire(Autoimmune regulator)は TSAs の胸
腺内部での発現に関与する545 アミノ酸からなる転 写調節因子で,主に胸腺,特に髄質細胞上皮に発現 しているが,樹状細胞にも多く発現している3−6). 髄質上皮細胞内で転写・翻訳されたTSAs は分解さ れ,MHC class II によって T 細胞に提示される.こ の自己抗原であるTSAs に反応する自己反応性 T 細 胞はアポトーシスにより除去される7). 2.制御性 T 細胞と末梢性免疫寛容 自己反応性T 細胞がネガティブセレクションを 回避して末梢に流出しても,多くはアナジー状態 もしくは制御性T 細胞(Treg)によって不活化され る(図1 ).Treg には胸腺内で発生し安定した制御
機能を有するnaturally occurring Treg(nTreg),末梢 においてnaïve CD4+T 細胞から誘導される inducible Treg(iTreg)の他,Tr1,Th3,CD8+Treg などの種 類が報告されている.Forkhead box 転写因子 FOXP3
はnTreg および iTreg の分化成熟のマスター遺伝子 として重要な役割を果たす8).nTreg は Treg-specific demethylation region の脱メチル化をうけており,エ ピジェネティックに安定性が保たれている9).nTreg 自身は抗原特異性を持つが,抗原提示細胞の抑制, 膜型あるいは分泌型TGF-β,IL-10 などを介して抗 原特異性とは無関係に周囲のT 細胞を不活化する ことから,Treg の作用は優性免疫寛容と呼ばれて いる10). 3.B 細胞免疫寛容 B 細胞の免疫寛容は骨髄内および末梢リンパ組織 で2 段階のステップで行われる(図 2 ).まず,骨 髄内に存在する細胞の表面抗原に反応する細胞は, BCR 軽鎖の入れ替え(レセプター編集)によりそ の抗原特異性が変化し,なお自己反応性のあるもの はclonal deletion を受ける(central checkpoint)11, 12).
さらにB 細胞は末梢リンパ組織のリンパ濾胞で
somatic hypermutation による親和性成熟やクラスス イッチを行う.この際に同じ抗原特異性を持つ濾胞
図1 T 細胞の免疫寛容
APC; antigen presenting cells, DNT; double-negative T cell, DPT; double-positive T cell, FOXP3; Forkhead transcription factor P3, nTreg; naturally occurring regulatory T cells, iTreg; inducible regulatory T cells.
性ヘルパーT 細胞(Tfh)からシグナル伝達が必要 である13, 14).胸腺で作られたnTreg 由来と考えられ る濾胞性Treg は濾胞のサイズを規定し,また Tfh を抑制するとともに,perforin-granzyme 系による直 接破壊などを介して自己反応性B 細胞を制御する (peripheral checkpoint)14−18). B リンパ球の表面マーカーを用いて単一細胞毎の 自己反応性BCR を検討すると,健常人であっても
骨髄中のCD34−CD19+CD10+IgM−のearly immature B
細胞では75%以上が自己反応性である.自己反応
性 比 率 はcentral checkpoint を 受 け た CD34−CD19+ CD10+IgM−のimmature B 細胞では 20−50%,periph-eral checkpoint を受けた後の末梢血 CD19+CD10−IgM+ CD27− mature naïve B 細胞では 20%程度まで減少す る11, 12, 19).
Ⅲ.免疫寛容の破綻と自己免疫疾患
免疫寛容メカニズムが破綻すると,多様な臓器 特異性自己免疫障害が生じ,Aire 遺伝子の変異は APECED を,FOXP3 の変異は IPEX 症候群を来す.
いずれの場合も自己反応性T 細胞除去や抑制の異 常であるが,エフェクターT 細胞のみならず自己 反応性Tfh の生存を来し,結果として同じ抗原特異 性を持つ自己反応性B 細胞の蓄積と自己抗体産生 をもたらすと考えられる14).すなわち,自己抗体の 存在は同じ抗原に対する自己反応性T 細胞の存在 を示唆するものであり,実際に自己抗体によって同 定された自己抗原の多くは,同一患者のT 細胞を 刺激することが知られている20). 1.APECED APECED は,Aire 遺伝子の変異によりネガティ ブセレクションが起こらず,自己反応性T 細胞が 末梢に流出する常染色体劣性遺伝形式の疾患であ り,免疫調節不全を来す原発性免疫不全症に分類さ れている3, 21).APECED における Aire 遺伝子変異は これまでに60 以上が報告されている3).Addison 病 や副甲状腺機能低下症,1 型糖尿病などの内分泌症 状,皮膚粘膜カンジダ症,禿瘡や白斑などの外胚葉 異形成を来し,消化器症状を来す症例も10%程度 存在する(表1 )22, 23).中でもAddison 病,副甲状 腺機能低下症,皮膚粘膜カンジダ症は発症頻度が高 く,これらの組み合わせおよびAire 遺伝子の変異 が診断基準となる.カンジダ症は5 歳までに,内分 泌症状は10−15 歳頃に発症することが多く,IPEX 症候群と比較し遅い22, 23).北欧諸国で1 万に 1 人と 頻度が高いが我が国ではまれである24).治療法は, 合併症に対するホルモン,カルシウム補充療法や抗 真菌剤投与,免疫抑制剤投与である. 図2 B 細胞の免疫寛容
2.IPEX 症候群
IPEX 症候群は,FOXP3 遺伝子の変異による Treg
の機能不全が原因で発症するX 連鎖性の疾患であ る25).これまでに少なくとも国内で7 家系が知られ, 世界で50 以上の変異が同定されている.腸炎の本 態は自己免疫性腸症であり,食事療法・止痢剤・完 全静脈栄養に反応しない難治性下痢症が乳児期に発 症し,小腸上皮細胞に対する自己抗体が血清中に検 出される26).炎症は小腸に強いことが多く,分泌性 下痢となることから,容易に脱水と電解質異常を呈 する.内分泌症状では1 型糖尿病と甲状腺機能低下 症が多く,新生児期に発症,もしくは新生児マス スクリーニングで発見されることもある.一方, APECED に見られる Addison 病や副甲状腺機能低下 症の報告はない(表2 )25).治療としてホルモン補 充療法,免疫抑制剤,副腎皮質ステロイドなどの投 与が行われるが,唯一の根本的治療は造血幹細胞移 植である. Kinnunen らは IPEX 症候群患者において先に触れ たMeffre らの方法で B 細胞の各分化段階での自己 反応性を検討し,CD34−CD19+CD10+IgM−のimmature B 細胞までの自己反応性 B 細胞比率は対照と変わ らないが,CD19+CD10−IgM+CD27− mature naïve B 細
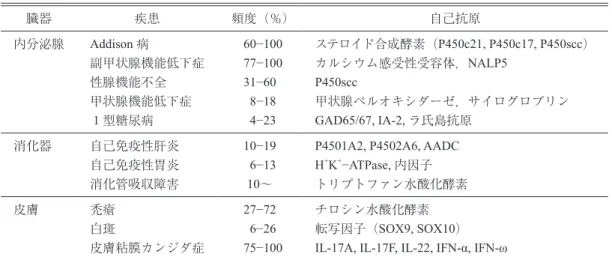
胞でその比率が低下しないことを示した27).この結 表1 APECED における自己免疫疾患と対応抗原 臓器 疾患 頻度(%) 自己抗原 内分泌腺 Addison 病 副甲状腺機能低下症 性腺機能不全 甲状腺機能低下症 1 型糖尿病 60−100 77−100 31−60 8−18 4−23 ステロイド合成酵素(P450c21, P450c17, P450scc) カルシウム感受性受容体,NALP5 P450scc 甲状腺ペルオキシダーゼ,サイログロブリン GAD65/67, IA-2, ラ氏島抗原 消化器 自己免疫性肝炎 自己免疫性胃炎 消化管吸収障害 10−19 6−13 10~
P4501A2, P4502A6, AADC
H+K+−ATPase, 内因子 トリプトファン水酸化酵素 皮膚 禿瘡 白斑 皮膚粘膜カンジダ症 27−72 6−26 75−100 チロシン水酸化酵素 転写因子(SOX9, SOX10) IL-17A, IL-17F, IL-22, IFN-α, IFN-ω 文献22, 23)より
GAD; glutamic acid decarboxylase, IA; insulinoma-2 antigen, NALP5; NACHT leucine-rich-repeat protein 5.
表2 IPEX 症候群における自己免疫疾患と対応抗原 臓器 疾患(頻度) 自己抗原 消化器 自己免疫性腸症(124/136) 食物アレルギー 肝炎(8/136)・胆管炎 AIE-75,villin 抗食物抗原(IgE)
平滑筋,LKM(liver kidney microsome)-1
内分泌腺 甲状腺機能低下症(27/136) 1 型糖尿病(> 83/136) 甲状腺ペルオキシダーゼ,サイログロブリン GAD,ラ氏島抗原 血液 溶血性貧血 血小板減少(42/136) 好中球減少症 クームス 血小板 好中球 腎臓 間質性腎炎(5/136) 糸球体腎炎・MGN, MCNS;(14/136) AIE-75? villin? 基底膜 神経 重症筋無力症 アセチルコリン受容体 皮膚 湿疹≫類天疱瘡様,乾癬様,禿瘡;(95/136) Keratin-14 その他 全身性血管炎 ? 文献25)より
AIE-75; autoimmune enteropathy-related 75kDa antigen, GAD; glutamic acid decarboxylase, MGN; membranous glomerulonephritis, MCNS; minimal change nephrotic syndrome.
果は,Treg 機能不全では central checkpoint は機能す るが,peripheral checkpoint が働かない事を示してい る. 3.APECED と IPEX の臨床像の比較 ⑴ 標的臓器の違い APECED および IPEX 症候群は各々中枢性および 末梢性免疫寛容の障害による自己免疫疾患であり, 内分泌症状,消化管症状,皮膚疾患など一部共通し た臨床症状を来す.しかし,APECED では前述の通 り副甲状腺機能低下症,Addison 病,皮膚粘膜カン ジダ症の頻度が高く,IPEX 症候群では 1 型糖尿病, 自己免疫性腸症,湿疹が多いなどの違いも見られ る22, 23, 25).以上より中枢性寛容不全であるAPECED と末梢性免疫寛容不全であるIPEX の間には重なり はあるものの障害組織の分布や頻度に大きな違いが あることがわかる. ⑵ 同一組織における自己抗原の違い 筆者らは既にIPEX 症候群患者の血清を用いて
AIE-75(Autoimmune enteropathy-related 75kDa antigen)
とvillin が疾患特異性の高い自己抗原であることを 報告している28−30).AIE-75 は 521 アミノ酸からなり, 蛋白結合モジュールであるPDZ ドメインを 3 つ有 する29).小腸上皮細胞や腎近位尿細管上皮細胞の刷 子縁および網膜ならびに内耳上皮細胞の刷子縁類似 構造におけるscaffold protein として機能し,その変 異は症候性難聴の1 つである Usher 症候群 Type-Ic の原因である31).Villin はアクチン結合タンパクの 1 つであり,やはり小腸などの絨毛構造に発現して いる32).こうした抗原分布に一致して,IPEX 症候 群では十二指腸および小腸粘膜上皮に強い炎症と絨 毛萎縮を生じる.一方APECED でも頻度は少ない ながら消化器症状を呈する症例がある33).こうした 症例ではトリプトファン水酸化酵素-1(TPH-1)に 対する自己抗体がみられ34−36),小腸粘膜上皮組織で はTPH-1 を有するエンテロクロマフィン細胞のみ が脱落していることが報告されている36).これら3 つの自己抗体の疾患特異性は正常人および他の自己 免疫疾患や消化管疾患をコントロールとして検討さ れてきたが,APECED と IPEX 症候群の間での検討 はなされていなかった. そこで筆者らは両疾患に共通しながら症状の異 なる腸疾患における自己抗体に着目した.TPH-1, AIE-75,villin の遺伝子組換えタンパクを作製し, APECED 患者 23 名,IPEX 症候群患者 7 名の血清 を用いそれぞれの抗体価をimmunoblot 法で測定 した37).その結果,抗TPH-1 自己抗体は APECED のみで,抗AIE-75 自己抗体は IPEX 症候群のみで 検出され,その疾患特異性が確認された.一方抗 villin 自己抗体については APECED 患者で陽性とな る症例があり,特異性が低いことが判明した.そ れぞれの疾患において消化管病変がTPH-1 および AIE-75 の分布に一致していることから,抗原特異 的機序で組織破壊が生じていると考えられた. ⑶ 自己抗体の病的意義 細胞表面抗原に対する自己抗体,すなわち血球減 少症における血球表面抗原に対する抗体,バセドウ 病における抗TSH 受容体抗体,重症筋無力症にお ける抗アセチルコリン受容体抗体などの病的役割は 理解しやすいが,その数はむしろ限られている.実 際,AIE-75, villin,TPH-1 を含め,APECED や IPEX 症候群における自己抗原の多くは細胞質内蛋白であ る29, 30, 32).したがって自己抗体がこれらの抗原と接 触するためには,先に何らかの刺激で組織の破壊が 生じている必要がある.AIE-75 や villin は腎近位尿 細管にも強く発現しているが,間質性腎炎を来す症 例は多くはない25, 28−30).消化管が常に細菌やウイル スに接しているのに対し腎は基本的に無菌的である こと,消化器症状が感染や食物アレルギーを契機に 発症する症例があることなどを考慮すると38, 39),感 染症などによって組織破壊が生じた際に細胞質内抗 原が抗体と接する可能性が考えられる.ひとたび 抗原が流出すれば,自己抗体との複合体がFcgR を 保有する抗原提示細胞に,また抗原が抗原特異的 BCR をもつ B 細胞により取り込まれ,T 細胞への 抗原提示を介して免疫反応を増幅することが考えら れる40).FOXP3 変異を有し IPEX 症候群のモデルで あるscurfy(Sf)マウスにおいては一部の病態には B 細胞が強く関与し,また MHC class II を欠失させ ることでB 細胞の自己抗体産生が消失することか ら,自己抗体産生はT 細胞依存性であることが示 唆されている41). ⑷ 両疾患における標的抗原の違いに関する考察 こうした両疾患における組織および抗原特異性の 違いを説明するものは何か.胸腺においてAire 依 存性に発現するTSA の場合には説明は容易である. APECED で自己抗原となるチトクローム P450 や プレプロインスリンはマウスにおいてはAire 依存 性に胸腺に発現することが示されている6).一方, Aire ノックアウトおよび野生型のマウス胸腺におけ
るTSAs の発現を DNA アレイで比較すると,TPH-1 およびvillin は mRNA レベルで胸腺に発現している ことが示されているが,予想に反していずれもAire 非依存性である42).AIE-75 のマウスホモログは使 用されたアレイに含まれておらず,検討されていな い.胸腺におけるTSAs の発現が種によって異なる 可能性がある一方,Aire 自身が転写因子としてのみ でなく,抗原提示レベルで何らかの役割を果たして いる可能性も示唆されている3). FOXP3 変異を有する Sf マウスの初発症状は皮膚 や尾の異常や肝炎・胃炎でありIPEX 症候群との違 いが見られる43).腸炎はSf マウスの T 細胞を Rag-1 ノックアウトマウスに移入後に見られるが44),消化 管特異的自己抗体は検討されていない.したがっ て,今後は種の違いを加味した多面的な検討が必要 と思われる. もう一つの疑問は,Aire の Treg 分化における役 割である.Treg の分化には胸腺上皮細胞による自己 抗原の提示が必要とされており,Aire が重要な役割 を果たすとするマウスでのデータや45, 46),APECED 患者のTreg がやや減少しているという報告がある47). 一方で,Aire ノックアウトマウスでは Treg の数は 正常でありin vitro でも機能に問題はないという報 告もある4, 48, 49).もし,Aire による TSAs の胸腺内 発現がTreg の抗原特異性を規定する上で重要であ るとすれば,APECED は IPEX 症候群の症状すべて を網羅し,かつ重症であるはずである.しかし実際 にはIPEX 症候群の方が APECED と比較して発症 が早期で,かつ症状は激烈である.さらに両疾患に おける標的組織,あるいは同じ臓器であっても標的 抗原が異なるという事実は,Treg の分化に対する Aire の役割は,もしあっても限局したものである事 を示唆する. Ⅳ.おわりに 中枢性免疫寛容不全であるAPECED と末梢性免 疫寛容不全であるIPEX 症候群における標的臓器・ 標的自己抗原の違いは,抗原により中枢性ないし末 梢性免疫寛容への依存度が異なる可能性を示唆す る.しかし,そのメカニズムは不明な点が多く,マ ウスとヒトでの違いも踏まえた検討が待たれる. 文 献
1) Boehmer, H., et al.: Checkpoints in lymphocyte development and autoimmune disease. Nat Immu -nol. 11: 14−20, 2010.
2) 花房崇明:中枢性及び末梢性免疫寛容におけ るAire の働き.日本臨床免疫学会誌 37: 133− 138, 2014.
3) Mathis, D., et al.: Aire. Annu Rev Immunol. 27: 287−312, 2009.
4) Anderson, M.S., et al.: The cellular mechanism of Aire control of T cell tolerance. Immunity. 23: 227−239, 2005.
5) Hubert, F.X., et al.: A specific anti-Aire antibody reveals aire expression is restricted to medullary thymic epithelial cells and not expressed in periph-ery. J Immunol. 180: 3824−3832, 2008.
6) Anderson, M.S., et al.: Projection of an immuno-logical self shadow within the thymus by the aire protein. Science. 298: 1395−1401, 2002.
7) Mohan, J.F., Unanue, E.R.: Unconventional recog-nition of peptides by T cells and the implications for autoimmunity. Nat Rev Immunol. 12: 7121− 7128, 2012.
8) Sakaguchi, S., et al.: Regulatory T cells and im-mune tolerance. Cell. 133: 775−787, 2008.
9) Morikawa, H., Sakaguchi, S.: Genetic and epigen-etic basis of Treg cell development and function: from a FoxP3-centered view to an epigenome- defined view of natural Treg cells. Immunol Rev. 259: 192−205, 2014
10) Shevach, E.M., et al.: Mechanisms of foxp3+ T regulatory cell-mediated suppression. Immunity. 30: 636−645, 2009.
11) Wardemann, H., et al.: Predominant autoantibody production by early human B cell precursors.
Science. 301: 1374−1377, 2003.
12) Wardemann, H., et al.: Human autoantibody silenc-ing by immunoglobulin light chains. J Exp Med. 200: 191−199, 2004.
13) Craft, J.E.: Follicular helper T cells in immunity and systemic autoimmunity. Nat Rev Rheumatol. 8: 334−347, 2012.
14) Ueno, H., et al.: Pathophysiology of T follicular helper cells in humans and mice. Nat Immunol. 16: 142−152, 2015.
15) Lim, H.W., et al.: Cutting Edge: Direct Suppres-sion of B Cells by CD4+ CD25+ regulatory T cells. J Immunol. 175: 4180−4183, 2005.
16) Iikuni, N., et al.: Cutting Edge: Regulatory T cells directly suppress B cells in systemic lupus
erythe-matosus. J Immunol. 183: 1518−1522, 2009. 17) Linterman, M.A., et al.: Foxp3+ follicular
regula-tory T cells control T follicular helper cells and the germinal center response. Nat Med. 17: 975−982, 2012.
18) Chung, Y., et al.: Follicular regulatory T (Tfr) cells with dual Foxp3 and Bcl6 expression suppress germinal center reactions. Nat Med. 17: 983−988, 2012.
19) Meffre, E., et al.: B-cell tolerance checkpoints in health and autoimmunity. Curr Opin Immunol. 20: 632−638, 2008.
20) Monneaux, F., Muller, S.: Epitope spreading in systemic lupus erythematosus: identification of triggering peptide sequences. Arthritis Rheum. 46: 1430−1438, 2002.
21) Al-Herz, W., et al: Primary immunodeficiency diseases: an update on the classification from the International Union of Immunological Societies Expert Committee for Primary Immunodeficiency.
Front Immunol. 2014. doi: 10.3389/fimmu.2014.
00162.
22) Betterle, C., et al.: Autoimmune adrenal insuffi-ciency and autoimmune polyendocrine syndromes: autoantibodies, autoantigens, and their applicabili-ty in diagnosis and disease prediction. Endocr Rev. 23: 327−364, 2002.
23) Perheentupa, J.: Autoimmune polyendocrinopathy- candidiasis-ectodermal dystrophy. J Clin
Endocri-nol Metab. 91: 2843−2850, 2006.
24) Björses, P., et al.: Genetic homogeneity of auto-immune polyglandular disease type I. Am J Hum
Genet. 59: 879−886, 1999.
25) Barzaghi, F., et al.: Immune dysregulation, poly-endocrinopathy, enteropathy, x-linked syndrome: a paradigm of immunodeficiency with autoimmuni-ty. Front Immunol. 3: 211, 2012.
26) Unsworth, D.J., Walker-Smith, J.A.: Autoimmunity in diarrhoeal disease. J Pediatr Gastroenterol Nutr. 4: 375−380, 1985.
27) Kinnunen, T., et al.: Accumulation of peripheral autoreactive B cells in the absence of functional human regulatory T cells. Blood. 121: 1595−1603, 2013.
28) Kobayashi, I., et al.: A 75-kD autoantigen recog-nized by sera from patients with X-linked autoim-mune enteropathy associated with nephropathy.
Clin Exp Immunol. 111: 527−531, 1998.
29) Kobayashi, I., et al.: Identification of an autoim-mune enteropathy-related 75-kilodalton antigen.
Gastroenterology. 117: 823−830, 1999.
30) Kobayashi, I., et al.: Autoantibodies to villin occur frequently in IPEX, a severe immune dysregula-tion, syndrome caused by mutation of FOXP3.
Clin Immunol. 141: 83−89, 2011.
31) Verpy, E., et al.: A defect in harmonin, a PDZ domain-containing protein expressed in the inner ear sensory hair cells, underlies Usher syndrome type 1C. Nature Genetics. 26: 51−56, 2000.
32) Khurana, S., et al.: Regulation of cell structure and function by actin-binding proteins: villin’s perspec-tive. FEBS Lett. 582: 2128−2239, 2008.
33) Sayar, E., et al.: Intestinal dysfunction in APECED syndrome could mimic IPEX syndrome. J Pediatr
Gastroenterol Nutr. 56: 2013.
34) Ekwall, O., et al.: Tryptophan hydroxylase auto-antibodies and intestinal diseasein autoimmune polyendocrine syndrome type 1. Lancet. 354: 568, 1999.
35) Dal Pra, C., et al.: Autoantibodies to human tryp-tophan hydroxylase and aromatic L-amino acid decarboxylase. Eur J Endocrinol. 150: 313−321, 2004.
36) Scarpa, R., et al.: Tryptophan hydroxylase autoan-tibodies as markers of a distinct autoimmune gas-trointestinal component of autoimmune polyendo-crine syndrome type 1. J Clin Endocrinol Metab. 98: 704−712, 2013.
37) Chida, N., et al.: Disease specificity of anti-trypto-phan hydroxylase-1 and anti-AIE-75 autoantibod-ies in APECED and IPEX syndrome. Clin
Immu-nol. 156: 36−42, 2015.
38) Satake, N., et al.: A Japanese family of X-linked auto-immune enteropathy with haemolytic anaemia and polyendocrinopathy. Eur J Pediatr. 152: 313− 315, 1993.
39) Torgerson, T.R., et al.: Severe food allergy as a variant of IPEX syndrome caused by a deletion in a noncoding region of the FOXP3 gene.
Gastroen-terology. 132: 1705−1717, 2007.
40) Avalos, A.M., Ploegh, H.L.: Early BCR events and antigen capture, processing, and loading on MHC class II on B cells. Front Immunol. 10: 92, 2014. 41) Aschermann, S., et al.: B cells are critical for
auto-immune pathology in Scurfy mice. Proc Natl Acad
Sci U S A. 110: 19042−19047, 2013.
42) Derbinski, J., et al.: Promiscuous gene expression in thymic epithelial cells is regulated at multiple levels. J Exp Med. 202: 33−45, 2005.
43) Godfrey, V.L., et al.: X-linked lymphoreticular dis-ease in the scurfy (sf) mutant mouse. Am J Pathol. 138: 1379−1387, 1991.
44) Godfrey, V.L., et al.: Transplantation of T cell- mediated, lymphoreticular disease from the scurfy (sf) mouse. Am J Pathol. 145: 281−286, 1994. 45) Aschenbrenner, K., et al.: Selection of Foxp3+
reg-ulatory T cells specific for self antigen expressed and presented by Aire+ medullary thymic epitheli-al cells. Nat Immunol. 8: 351−358, 2007.
46) Malchow, S., et al.: Aire-dependent thymic de-velopment of tumor-associated regulatory T cells.
Science. 339: 1219−1224, 2013.
47) Kekäläinen, E., et al.: A defect of regulatory T cells in patients with autoimmune polyendocrinopathy- candidiasis-ectodermal dystrophy. J Immunol. 178: 1208−1215, 2007.
48) Kuroda, N., et al.: Development of autoimmunity against transcriptionally unrepressed target antigen in the thymus of Aire-deficient mice. J Immunol. 174: 1862−1870, 2005.
49) Liston, A., et al.: Aire regulates negative selection of organ-specific T cells. Nat Immunol. 4: 350− 354, 2003.