九州大学学術情報リポジトリ
Kyushu University Institutional Repository
水稲の光合成・蒸散および乾物生産におけるケイ酸 の生理的作用に関する研究
東江, 栄
九州大学農学研究科農学専攻
https://doi.org/10.11501/3097516
出版情報:Kyushu University, 1994, 博士(農学), 課程博士 バージョン:
φ
水稲の光合成 ・ 蒸散および乾物生産にお けるケイ酸の生理的作用に関する研究
東江 栄
1 994
第1章 緒 論ーーーー
目 と欠
第2章 生長 ・ 乾物生産に及ぼすケイ酸の影響 1 . はじめに- -
2. 材料と方法
第1実験. 生長 ・ 乾物生産及び光合成速度に対する ケイ酸施用効果の品種間差一一ーー
第2実験. 異なる光環境下で生育させた水稲の生長 ・
乾物生産に及ぼすケイ酸の影響ーーーーーーーーーーーーー-12 3. 結果
第1実験. 生長 ・ 乾物生産及び光合成速度に対する
ケイ酸施用効果の品種間差-ーーーーーーー一一ーーーーーーー-14 第2実験. 異なる光環境下で生育させた水稲の生長 ・
乾物生産に及ぼすケイ酸の影響一一ーーーーーー--ーーーー25
4. 考察-ーーーーーーーーーーーーーーー一ーーーーーーー一一一一一一一ー一一ーーーーー一一一一ーーーーーー-32
5. 摘要ーー一ーーーーーーーーーー一ー一一ーーーーーーーーー一一一一一ー一ーーーーーーー一一ーーーーーーー-41
第3章 個葉光合成 ・ 蒸散速度及び拡散伝導度に及ぼすケイ酸の
影響ー一ー一一ーーーーーーーーーーーーー一ーーーーーーーーーーーーーーーーで一ーーーーーー一ーーーーー-44 第1節 生育光環境及びケイ酸濃度が異なる条件下で栽培した
葉身の光合成反応に及ぼすケイ酸の影響
1 . はじめにーー一一ー一ー一ーーーーーーーー一ーー一一一一一一一ーーーーーー一一ー一一ーーーーーーーー-46 2. 材料と方法
第1実験. 生育光環境が異なる葉身の光合成 ・ 蒸散速度 及び舷散伝導度に及ぼすケイ酸の影響ーーーーーーー-48 第2実験. ケイ酸濃度の異なる葉身の光反射率, 吸収率,
3. 結果
透過率, 光エネルギ一利用効率, 光量子収率 及び葉肉活性に及ぼすケイ酸の影響--ー一ー一一一一-49
第1実験. 生育光環境が異なる葉身の光合成 ・ 蒸散速度 及び拡散伝導度に及ぼすケイ酸の影響一一ーーー一一一52 第2実験. ケイ酸濃度の異なる葉身の光反射率, 吸収率,
透過率, 光エネルギ一利用効率, 光量子収率 及び葉肉活性に及ぼすケイ酸の影響ーーー一ーーーーー-54
4. 考察-ーーーーーーーー一一ーーーーーーーーーーーー一一ーーーーーー一一一ー一一ーー一ー一ー一一一ー一--65
5. 摘要-ーーーーーーーーーーーーーーーーーーーーーーーーーーーーー一一一四ーーーー一ーーーーーーーーーー-71
第2節 環境要因の変動に対する気孔反応並びに水ストレス下で
の蒸散速度, 気孔伝導度及び水ポテンシャルの日変化に 及ぼすケイ酸の影響
1 . はじめにーーーーーーーーーーーーーーーーーーーーーーー一ーーーーーーーーーーーーーーーーーーーーーー73 2. 材料と方法
第1実験. 環境要因の変動に対する気孔反応に及ぼす
ケイ酸の影響ーー一一ーーーーーーー一ーーーーーーー一ーーーーーーーーーー75 第2実験. 水ストレス下での個体蒸発散速度, 個葉
水ポテンシャル, 蒸散速度及び気孔伝導
度の日変化に及ぼすケイ酸の影響一ーーーーーーーーーーー77 3. 結果
第1実験. 環境要因の変動に対する気孔反応に及ぼす
ケイ酸の影響ーーーーーーーーーーーーー一ーーーーーーー一ーーーーー一ーー78 第2実験. 水ストレス下での個体蒸発散速度, 個葉
水ポテンシャル, 蒸散速度及び気孔伝導
度の日変化に及ぼすケイ酸の影響ー一一一ーーー一ー一一一89 4. 考察- -ーーーーーーーー一一ーーーーーーーーーーーーーーーーーーーー一一一ーーーーーーーーーーーーーー-89
5. 摘要-一一一ーー一一ー一一一一一一一一一ー一ー一一一ー一一一一一一一一一一ー一一一一ーーーーーーーーー100
第4章 ストレス耐性に関与した細胞強度に及ぼすケイ酸の影響
1 . はじめにーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー一ーーー一一ーーー一一102 2. 材料と方法一ーーーーーーーーー一一一一ーーーーーー一ー一一ーーーーーー一一ーーーーー一一ーーー-104
3. 結果
1 ) 測定法の検討--ー一ー一一ーーーーー一ーーーーーーーーーーーー一ー一一一一一ーー-106 2 ) 漏出に対するケイ酸の影響ー一一ー一一一一一一一一一一ーー一一ー一一一一108 3 )漏出を左右する要因に及ぼすケイ酸の影響ーーーーーーーー-114 4. 考察
1 ) 最適測定条件の決定一一一一一一一一ー一一一一一ー一一一一一一一一一一一一一ー123 2 )膜安定性のケイ酸処理区間での比較ーーー一ー一一ーーーーー一一一124
3 )漏出を左右する要因に及ぼすケイ酸の影響ー一一ーーー一一一126
5. 摘要一一一一ーー一一一一一一ー一一一一ーー一一ーーー一一一一一一一一一一ー一一一ー一一一一ー一一一一-130
第5章 葉身の生理的活性の維持に対するケイ酸の影響
1 . はじめに一一ー一一ーー一一ーーー一一一一ーー一ー一一一一一一一一一一一一一一一一一一一一一一ー一一133 2. 材料と方法
第1実験. 個体老化誘導実験ーー一一ーーーー一一一ーーー一一一一ー一一ーーーー134 第2実験. 葉身老化誘導実験ー一ー一一一ーーーーーー一ーー一一一一一ーー---136 3. 結果
第1実験. 個体老化誘導実験ーーーーーーーー一一一ー一一ー一ーーーー一ーーー-137 第2実験. 葉身老化誘導実験-ーーー一ーーー一ーー一ー一一ーー一一一一一ーー-1 45 4. 考察一一ー一一一一一一一一ー一一ーーーー一一一一一ー一一一一一ー一一一一一一ー一一一一一ーー一一一一一145
5. 摘要一一ーーーー一ーーーー一一ーーー一ーーーーーーーー一一ーーーーーーーーー一ーーーーーーーーーーー-1 55
第6童 総合考察-ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー一一ーーーー- -1 57
要 約一ー一ー一ーー一ーーーーーーー一ーーー一一ー一ーーー一ーー一ーーー一ーーーーーーーーーーー一ーーー一ーー-176
謝 辞-ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー-180 引用文献ーーーーーーー一ーーーーーーーーー-ーーーーーーーーーーーーー一ーーーーーーーーーーー一ーーーー-ーーー-181
多誇 1 � 系者 三壬'.,..ð...
函仔司
世界人口の半数以上がアジアに住んでおり, 米を常食としている(小
田, 1993). 様々な政策努力にも関わらず爆発的な人口増加の抑制は困 難であり, 世界的な食糧(穀類)の絶対量の不足に対する危倶から, 食 糧自給の観点にたった米生産量の増大が重要な課題であると考えられて いる. 生産量の増大対策として耕地面積の拡大がまず考えられるが, 耕 地の外延的拡大は物理的にほぼ限界に達しており, 加えて, 人口の増加 及び経済の発展にともなう工業化や公共施設への整備拡張が農地の工場
・ 住宅などへの用地転用を余儀なくし, 耕地面積は逆に減少する傾向に ある(小田, 1993). したがって, 食糧生産の向上のためには, 高収量 品種の導入, 肥料 ・ 農薬の投入及び基盤整備等, 農業技術の展開を含む 内延的拡大による土地生産性の向上が急務である.
現在, 零細農業が主体のアジアの国々において, 土地生産性の増大は 化成肥料の増授によって図られているが(小田, 1993), 近代化路線に よる労働力の流出及び兼業化は, 有機物の還元投与を減少させ, 水田の 老朽化による生産性の低迷を招いている(高橋ら, 1990). 水稲は後述 するようにケイ酸植物といわれ, その吸収量の多少が収量と密接な関係 をもっ(Elawad and Green jr, 1979; 高橋, 1975). よって, 高収を得 るためにはその要求量も必然的に増大すると考えられ, 老朽化による生 産性の低迷は, 一つには有機物還元投与の減少に寄因する有効態ケイ酸 の低下が原因であると考えられる(奥田, 1958;馬場, 1990; 高橋ら,
1990 )
このような社会情勢の変化に伴う農業生産システムの変動は, 経済発 展過程において日本稲作が経た系譜と酷似する. わが国では, 水田老 朽 化による生産性の低迷問題に対し, 独自に鉱さいを原料とするケイ酸質 肥料を開発し, ケイ酸の還元投与による土壌改良によって増収を実現さ
せた(奥田, 1958). 本肥料 の施用量は, 工業化の進行にともない増加
する廃棄鉱さいの有効利用という経済的目的とも合致し, 1957年に48万 トン, 1968年には130万トンにも達した. その後は水田の作付面積の減少 と共に減少はしたが依然45万トンの高水準を保っており(農林水産省肥 料機械課, 1993), 水田面積の増減とほぼパラレルに動いている. この ことは, 多肥集約型稲作の施肥技術の面でケイ酸肥料がいかに重要な位
置を占めているかを示している(高橋, 1975). 上述した近代化路線に よって必然、的に生じる問題点を抱える国々は, 最終的に日本と同様な系 譜をたどる可能性を有しており, ケイ酸肥料を施用することは必至であ ると考えられる. 韓国及び台湾ではすでにケイ酸肥料が実用され, 中国 においても近年新たにケイ酸質肥料が公定され(高橋ら, 1990), 今後 施用量が増加するものと考えられる.
わが国は, ケイ酸質肥料を世界で初めて公定規格に加えたことからも 明らかなように, ケイ酸の肥料学的研究では先鞭をとった. しかし, 圏 内の土壊肥料学的問題の解決に根ざした 肥料 の応用面に関する実証的検 討が主であり, 施用効果発現のメカニズムに関与した基礎的研究はか な り立ち遅れているように見受けられる. 基礎と応用は相伴って発展する もので, 現在 の慣行 法的な施用法が, 基礎的研究の進展で新たに得られ た情報を基に改善される可能性もある. 今後の基礎的研究の進展は, わ が国に 対して のみならず, 上述の問題点を抱える国々に有益な情報を提
供すると期待される.
ところで, ケイ素は自然界では, ケイ酸あるいはケイ酸塩として多量 に存在し, 土壌中無機成分の約50�70%を占める. そのため, 土壌に生
育する植物でケイ酸を含有しない種 はない. しかし, 土壌中の可給態ケ イ素は少なく, その含量は植物種により大きく異なる. ケイ素を積極的 に吸収蓄積する植物はísilica accumulatorsJあるいは「ケイ酸植物」
と呼ばれ, ísilica non-accumulatorsJ及び「石灰植物Jと対比される
内/臼
( T a k a h a s h i a n d M i y a k e, 1 9 7 7) . 例えば, マツ科, イネ科及びカヤツ
リグサ科に属する植物は, 茎葉中にケイ酸を%レベル(乾物重当り) で 蓄積するが, マメ科植物では微量である(Werner and Roth , 1983) 水稲は代表的なsi1ica-accumu1atorで, ケイ酸をイネ科植物の中で最も
多量に吸収蓄積する(Lewin and Reimann, 1969) .
ケイ酸は水稲の正常な生育に不可欠であり(Lewin and Reimann. 196 9 ; Takahashi and Miyake, 1977; E1awad and Green jr, 1979), ケイ
酸欠乏は, 葉身の下垂及び先端枯死を誘起し, 植物体全体を弱体化させ
る(Werner and Roth, 1983) .
集積植物における欠乏症や高等植物体内含有の共通性は, 植物の生長 に対するケイ酸の作用解明に対する検討を促し, 必須元素の候補として もしばしば検討された. 例えば, 戦前ではSommer(1926) , Lipman (19 38) , Ra1eigh (1938) , 大川(1936)によっていくつかの植物の生育に ケイ酸が良好な影響を与えることが報告されており, 戦後においては,
わが国の土壌肥料研究者によって, 水稲に対するケイ酸質資材の施用効 果の面から集中的に研究が行 われた. その結果, 組 織の物理的強化を介 した病害虫に対する機械的抵抗性の増加, 葉身の直立化にとも なう個体 光合成の増加 (Jones and Handreck, 1967), 過剰j蒸散の抑制(Yoshid a e t a1� , 1959)及び根の酸化力の増大(奥田 ・ 高橋, 1962b)等が明
らかとなり, ケイ酸は水稲にとって農業的に必須な元素と評価されるに 至った(岡本, 1970a; Ma � t a L , 1989) . 農業的に必須とは, 施用 によって生長量及び穀実収量が顕著に増加することは明確であるが, 効 果の基盤となる生理作用に対するケイ酸の直接的な役割は不明であるこ とを意味する. これを, Arnon and Stout (1939) の必須元素の基準にて
らすと, ケイ酸は, 必須元素が植物の生活環の成就に 関与すること, 及 び欠乏症に元素特異性があることといった基準は満たすが, 必須元素が 植物の生理作用中重要な反応及び関連化合物分子の不可欠な構成元素で
あるという基準を満たしていない.
このように, 植物の生育とケイ酸の関係について明確な結論が出され ていないのが現状であるが, これには従来の研究の方向性にも問題があ ったように思われる. これまでケイ酸の効果として最も広く認知されて
いるも のは, 前述したように葉身の直立化, 倒伏防止及び病虫害抵抗性 の増加等, 換言すれば, 多量に蓄積, 重合したケイ酸による組織の物理 的な強化作用であり, 他の必須元素にみられるような生化学的 ・ 代謝生 理的作用で は な い. これはケイ酸の肥効が着目された当時の測定機器 ・ 手法の問題も考えられるが, ケイ酸の葉身内の存在形態、及び局在部位が 化学的に不活性で, 代謝生理的機能 に対する効果が期待されなかったこ とにも起因する(R a v e n, 1 9 8 3; S a n g s t e r a n d H 0 d s 0 n, 1986;高橋, 19 87) . 近年, 一部のケイ藻のDNA複製(Darley an d Volcani, 1969) 及び鳥類の骨格の石灰化にとってケイ酸が必須であることが明らかとな っており(Carlisle, 1972), 高等植物においても, 必須性を示唆する 代謝生理的な 作用を有することが期待される. 従来の研究の方向性を変 え, 生理的機能, 例えば光合成 ・ 呼吸といった植物の生長に直媛関与し た諸反応を新手法で検討すれば, このような新たな生理機能を見いだす
ことができると考えられる.
ケイ酸の効果は生殖生長期に顕著となる(Lewin an d Reimann, 1969;
Takahashi an d Miyake, 1977; Elawa d an d Green jr, 1979; Ma e t a 1.
1989) . 生殖生長期における水稲体の健全性は, 栄養生長期に発現する
生理的な諸反応に加え, 開花 ・ 稔実生理に関連した諸反応系が関与する.
さらに 収量の決定期には, 栄養生長期に蓄積された同化産物の穂への再 転流を左右する窒素 ・ 炭水化物代謝に関与した反応系がこれに加わる.
したがって, 水稲の収量向上に関与する律速因子の限定 ・ 解析は難しく,
ケイ酸のように細胞内の挙動がいまだ確定されていない元素の機能を,
生殖生長期における作用も含め解明することは困難を極める. 一方, 栄
A斗A
養生長期の生長量に関与する反応系は全反応系に対する光合成反応の寄 与度が高く, 比較的単純化が可能である. また, 出稼以前, 栄養生長期 に同化された乾物重と収量との間には密接な関係があり, 栄養生長期の 貯蔵炭水化物の増加は収量の増加に大きく貢献する(翁ら, 1982) . こ れらのことから, 本研究では栄養生長期あるいは出穂直後の水稲を対象 に, この時期の生長量及び個葉の生理的諸反応に及ぼすケイ酸の影響を 検討することにした.
栄養生長期の生長量は, 乾物生産的には, 総光合成量と総呼吸量との 収支として与えられる純生産量の積算量として捉えられ, 個体葉群の積 算C 0 2固定量がこれを大きく左右する. 自然条件下で種々の環境変化に さらされる水稲体の個体葉群全体の積算C 0 2固定量は着生葉身個々の最 大光合成能力, 葉群の空間配置及び着生葉身の形状, さらに環境変動に 対応した光合成効率及びその能力の維持能力に大きく左右されると考え られる. このような観点から, 第2章ではまず生長 ・ 乾物生産に対する ケイ酸の効果を, 効果の品種間差及び遮光処理の影響を比較する中で検 討し, 生長に直接関与すると考えられる個体全 体及び主稗着生葉身の光 合成速度を測定し, 下位葉の老化抑制及び草型に及ぼすケイ酸の影響の 比較から生長 ・ 乾物生産に対する施用効果の要因を解析した.
第3章では, 個葉の ガス交換速度に対するケイ酸の影響を検討するこ とを目的に, 個葉の光合成反応を, 光化学反応系, 炭酸ガス拡散系及び 炭酸固定系に大別し, それぞれに対するケイ酸の影響を検討した. あわ せて, 急激な 環境変化に対する個葉の光合成効率向上に関与した孔辺細 胞の環境応答に及ぼすケイ酸の影響を 光(青色光) , 湿度及びC 0 2濃度 の変化に対する拡散伝導度の変化から検討した.
第4章では, 個葉光合成能力の維持に及ぼすケイ酸の影響を種々の環 境ストレスに対する耐性の比較から明らかにすることを目的に, 細胞強 度の指標である細胞膜安定性を電解質の漏出度から評価し, 漏出度を左
右する諸 要因に及ぼすケイ酸の影 響を 検 討した.
第5章では, 光合成速度の内生的な維持能力(老化抑制)に対するケ
イ酸の作用について, ケイ酸の作用形態及び作用器官の限定を目的に,
ケイ酸添加日数の異なる個体及び切除葉身に, 人為的に老化を誘導し,
老化の進行度とケイ酸の重合度との関 連性を検討した.
以上の結果を基に, 第6章の総合考察では, 水稲の乾物生産を左右す る要因をC 0 2固定能とその維持能力とに大別し, 個体レベルから細胞レ
ベルにわたるケイ酸の生理作用を総合的に考察した.
にU
空手事 2 生主 画変σ〉景2三謹E
1 . はじめに
車乞�勿 産主Lこ五三三Lますーク{イ
化成肥料を多投する多肥集約型農法は, 比較的容易に収量増加を実現 する反面, 有機物還元投与の低下による減収を招く . これは一つに水稲 による土壌からの可給態ケイ酸の収奪が原因である(馬場, 1990;高橋 ら , 1990). 高度に集約化が進んだわが国の水稲裁培においてはケイ酸
質肥料の大量投与が不可欠と なっている(農林水産省肥料機械諜, 1993) 零細農が主体で土地生産性の増加が急務である国々では, いずれもこの ような収量低迷の可能性を有して おり, 効率的な施肥法を考慮した基礎
的な情報の需要が今後増大するものと 考えられる.
水稲に対するケイ酸 施用効果については, わが国が先鞭を取るかたち で研究が行われ多くの情報集積がある(Takahashi e t al!_ 1990) . しか し, 国内の土壌肥料学的な問題を念頭においたものが多く , ほとんどが
日本型品種を供試したものである. また, 乾物重に対する明確な効果を 報告したものの中には乾物重にケイ酸重を含め評価したものも 多い. 水 稲体内に蓄積したケイ酸は水稲乾物重の数%を占め ることを考慮すると,
これまでわが国で報告されているケイ酸施用による乾物重増加は, 単に ケイ酸重の増加であった可能性も考えられる. したがって, 上述した問 題点、に答えるべく ケイ酸施用効果を明確化するためには, 広い地域から 品種を選抜し厳密な水耕栽培を行った上で, 有機物の増加に関する施用 効果を検討する必要がある.
水稲の生長 ・ 乾物 生産に対するケイ酸の施用効果として, こ れまで多 くの報告がなされ, おおむね正の効果があるとされる(Elawad and Gre en jr, 1979) . 施用効果の生理的要因としてこれまで, 1)葉身直立化 による群落光合成の増大(岩田 ・ 馬場, 1961; Yoshida e t al!_ , 1969),
2 )耐病害虫性の向上(秋本 , 1929; 石塚 ・ 早川, 1951; Tanak a and P ark, 1966; Jones and Handreck, 1967), 及び3 )過剰蒸散の抑制 Yoshida e t a1!_ , 1959・ 1962c; 吉田, 1965)等が報告されている.
特に, 葉身の直立効果は, 耐肥性向上のため に不可欠な要因であったこ とから, 窒素肥料との複合試験で 数多くの検討が行われた. この効果は 個体の光合成速度, ひいては生長 ・ 乾物生産量を左右する重要な要因の 一つであり詳細に検討する必要がある. しかし, 従来の報告では群落構 造及び吸光係数等を用いて効果を数量化したものが少なく , 要因解析が 充分行われていないのが現状である(岩田 ・ 馬場, 1961; Yoshida et a Lι, 1969)
ところで, ある元素の施用効果の発現は, 植物のおかれた環境条件に より大きく変動する. 劣悪な環境条件下で植物を栽培し, 施用試験を行 うことは処理区間差をより明確にし, 元素の生理機能を把握するための 有効な手段である. また, その環境下で発現 する植物体の反応を数値化 し一般化を試みることは, 栽培上有効な情報を得る重要な手段である.
これまで, 乾燥(吉田, 1965), 高低温(岡本, 1969a, b) , 弱光(岡本,
1970b; 東江ら, 1992), 塩(Ahmad � t a1!_ , 1992) , 放射線(高橋,
1966)及び有害物質(高橋, 1987)等の下で行われてきたケイ酸施用効
果の検討もこのような発想、を基に行われたと考えられる. 上述したよう に, ケイ酸欠如個体では葉身の下垂が多く, 相互遮弊の影響をうけやす い. さらに, 第3章で検討するように, 近年, 重合蓄積したケイ酸が光 合成の光化学反応系に関与することが示唆されていること から, ケイ酸 の影響は光環境との関連で顕著になると考えられる.
以上のことをふまえ本章では以下の実験を行った. まず第1実験では,
生長 ・ 乾物生産に対するケイ酸施用効果を明らかにし, あわせて効果の 著し い品種の選定を行うことを目的に, 世界各地で栽培されている水稲 1 0品種を同ーの水耕液で栽培し, 草丈, 分げつ数等の生長形質並びに各
。。
器官の有機物乾物重の比較を行った. さらに, 乾物重増加の要因解析と して, 選定した品種の個体構造, 吸光係数及び主稗着生葉身の光合成速 度を測定した. 第2実験では, 不良環境条件下でのケイ酸施用効果を明 らかにすることを目的に, 第1 実験で施用効果が異なった3品種につい て, 遮光条件下における生長 ・ 乾物生産の比較及び生長解析法による乾 物生産向上の要因解析を行い, さらに関連要因として主得着生葉の光合 成速度及びクロロフィル含量を測定した.
2. 材料と方法
第1実験 生長 ・ 乾物生産及び光合成速度に対するケイ酸施用効果の品 種間差
1 )供試材料と栽培条件
品種間差の検討には, 水稲Q工ヱ主主sativ a 種の, 日本型3品種, インド 型4品種, 日印交雑型2品種およびジャワ型1品種の計10品種(第1表) を供試した. 個体全体の光合成速度の測定にはコシヒカリを供試した.
発芽時における籾から匪乳へのケイ酸の転流を防ぐため, 催芽処理に
先立って種子の籾を取り除いた. その種子をべンレ ートT1000倍液で30 分間消毒した後, プラスチック製シャ ーレ内のろ紙上に併置し, 30 oc一 定に保った恒温器中で催芽させた. 播種目は, バングラデシュ産の3品 種で1991年6月28日, 他の品種で同年6月14日, 個体全体の光合成速度を 測定した コシヒカリで同年8月 21日とした. 幼苗を九州大学農学部実験園 場内のビニールハウスに移し約1週間蒸留水で栽培した後, 自作した培 養槽(4802)(第1図)に移植しYosh i daら(1976)の培養液で水耕栽 培した. 水耕液のpHは5.0"-' 5.5に調節し, 12"-'14日に1回液を更新した.
移植1週間後, ケイ酸処理を開始した.
2 )ケイ酸処理
ケイ酸処理には, 水ガラス(ケイ酸ナトリウム)を用い, ケイ酸( S i
Table 1. List of rice cul tivars (Oryza sativa L.) lJsed i n
this experiment.
Types Cultivars Habitats
Japonica Nipponbare Japan
Koshihikari Japan Nishihomare Japan
Japonica-indica Suweon 258 Republic of Korea 阿ilyang 28 Republic of Korea
lndica S47-2 阿adagascar
Chinigura (aman) Bangladesh Kalizira (aman) Bangladesh Nizrshail (aman) Bangladesh
Javanica AF70 Kenya
Fig. 1. Water culturing system used in this study,
showing Si02 treated (rlght) and untreated p 1 an t s (1 e f t ) .
-lO-!
O2) 100ppm処理区(+ S i区)および無処理区(- S i区)の2区を設 けた.
3 )生長形質の調査
ケイ酸処理開始時から 1週間毎に分げつ数, 草丈を調査した . ケイ酸 処理後35日と45日にサンプリングし葉面積を測定した. その後サンプル を800Cで乾燥させ, 器官別の乾物重を測定した.
4 )ケイ酸含量の測定
乾燥後, サンプルの反復を 1つにし各処理区のケイ酸含量を器官ごと に測定した. 測定には重量法を用いた(吉田 ・ 阿部, 1975). また, 水
耕液のケイ酸含量の測定には比色法(吉田 ・ 岡部, 1975)を用いた.
5 )光合成速度の品種間差
11--13 葉齢に達した植物体の最上位展開葉における光合成 ・ 蒸散速度 を, 携帯型光合成 ・ 蒸散速度測定装置(A D C, S P B -H 3 )を用いて測定した.
晴天時には自然光下で, 曇天時にはメタルハライドランプ(東芝, D400) で補光し測定した. 測定時の光条件は飽和光強度(1500μ molm-2s-1 )以 上とした.
6 )個体の光合成速度
ケイ酸処理後32日目から37日目, 1 2 葉齢に達した植物体の, 地上部全
体の光一光合成反応を, 環境条件を制御した条件下で通気式同化箱法に より測定した. この時の光強度は0'"'-' 1202.5μ molm-2s-1, 葉温は30:t O.
020C, 湿度は54. 1:t O. 73%であった . 測定には各処理区2個体供試した . 7 )個体構造及び吸光係数
個体の光合成速度測定後, 同化箱にセットした状態で地際から5cm毎の 照度を, 照度計(TOPCON MI- 3)を用いて各層5 --- 10ヶ所測定し, 各層毎 の平均相対照度と層別葉面積から, 以下に示したBear-Lambertの式(M 0
nsi and Saeki, 1953)を用いて吸光係数を算出した.
1 = 1 ø exp ( K F )
( 1 : 同化箱内の光強度, 1 ø : 植物体直上の光強度, F : 積算葉面積) 照度測定後, チャ ンパー底部から高さ5cm毎にサンプリングし800Cで乾 燥させ, 乾燥重を測定した. 乾燥重を基に個体構造を決定した.
8 )主稗着生葉の光合成速度
個体の光合成速度を測定した個体について, 主稗に着生する第1葉(
最上位展開葉)から下位第4葉までの光合成速度を, 携帯型光合成 ・ 蒸 散速度測定装置(A D C, S P B -H 3 )を用い測定した. 測定は, 環境条件を一
定にするため室内で, メタルハライドランプ(東芝, D-400)を光源とし 行った. 測定条件は光強度1505.8:t10.8μ molm-2s-1, 葉温29. 7:t 2. 3"C,
湿度55. 8:t 9.6%であった.
9 )クロロフィル含量
バングラデシュの3品種はケイ酸処理開始20日目, それ以外の品種で は27日目, 主稗に着生する最上位展開葉から下位第5葉までの葉緑素含 量を測定した. 測定には葉緑素計(ミノルタ, SPAD502)を用いた. クロ ロフィル含量(mgdm-2)はSPAD値を次式に代入し求めた. Y = -0.56+0.
1 3 X (Y: クロロフィル含量, X: SPAD値, r=0.82・・・).
第2実験 異なる光環境下で生育させた水稲の生長 ・ 乾物生産に及ぼす ケイ酸の影響
1 )供試材料と栽培条件
{共試材料には, 日本型の日本晴とコシヒカリ, 日印交雑型の水原258号
の3品種を用いた. 水稲種子の籾を取り除き, 玄米を殺菌処理後, プラ スチック製のシャーレ上に併置した. 矯種目は日本晴とコシヒカリで19 89年8月19日, 水原258号で同年8月31日であった. 発芽直後, 蒸留水で5
日間生育させた後, 42 Q容の水耕ポットに移植し, 第1実 験と同様な培 養液で水耕栽培した. 水耕液の更新間隔は6� 7日, 水耕液のpHは常時5.
0"-' 5.5に調節した.
円〆ム1i
2 )処理方法
2 - 1 )ケイ酸処理
水耕装置に移植5日後, 3葉齢期にケイ酸処理を開始した. 処理には,
水ガラス(ケイ酸ナトリウム)を用い, ケイ酸無処理区( 0 p pm, - S i 区)及び処理区(60 p pm, + S i区)の2区を設けた.
2 - 2 )遮光処理
播種後約1ヶ月間自然、光で栽培した後, 遮光処理を開始した. 光条件 は無遮光区と遮光区(遮光率40 % )の2段階とした.
3 )調査方法
ケイ酸処理開始時から5�7日毎に, 生育調査を行った. 日本晴, コ シヒカリでケイ酸処理後32日目と44日目, 水原258号で38日目と46日目に 各処理区4個体についてサンプリング調査した. 調査後, 植物体を器官 別に分け, 80 ocで乾燥後乾物重を測定した.
4 )ケイ酸含量の測定
乾燥後, サンプルの反復を1つにし, 各処理区のケイ酸含量を器官ご とに測定した. サンプル及び水耕液のケイ酸含量の測定法は第1実験と
同様である.
5 )主稗着生葉身の光合成速度
遮光処理後21日目(日本晴, コシヒカリでケイ酸処理後 40日目, 水原 258号で 42日目), 主稗に着生する第l葉から第5葉までの光合成速度を,
携帯型光合成 ・ 蒸散速度測定装置 (A D C, S P B -H )を用いて測定 した. 測 定は, 晴天時には太陽光下で, 曇天時にはメタルハライドランプ(東芝,
D400)で補光 し行った. 測定時の光条件は, 無遮光区で2000μ molm-2s- 1, 遮光区で1200"-'1500μ molm-2s-1とした.
6 )主稗着生葉身のクロ ロフィル含量
光合成速度測定後, 被測定葉の葉身約2cm2を切取り, 葉面積を測定し た後, 99 %エタノール(10m 1 )でクロ ロ フィルを溶出させ, 波長665及び
649nmにおける吸光度を測定した(W i n t e r m a n s a n d d e M 0 t S, 196 5) .
3. 結 果
第1実験 生長 ・ 乾物生産及び光合成速度に対する ケイ酸施用効果の品 種間差
1 )各器官のケイ酸含量
第2表に供試全品種系統のケイ酸処理後35日目の器官別ケイ酸含量を
示した. + S i区の葉, 茎および根のケイ酸含量は平均値でそれぞれ7.
59, 5.62および0.78%であった. ケイ酸含量は, 密陽28号で愚大であっ た. - S i区では器官別の蓄積量は0.05---0.8%であり, 蓄積の器官特異
性および品種間差は認められなかった.
2 )生長形質及び乾物生産
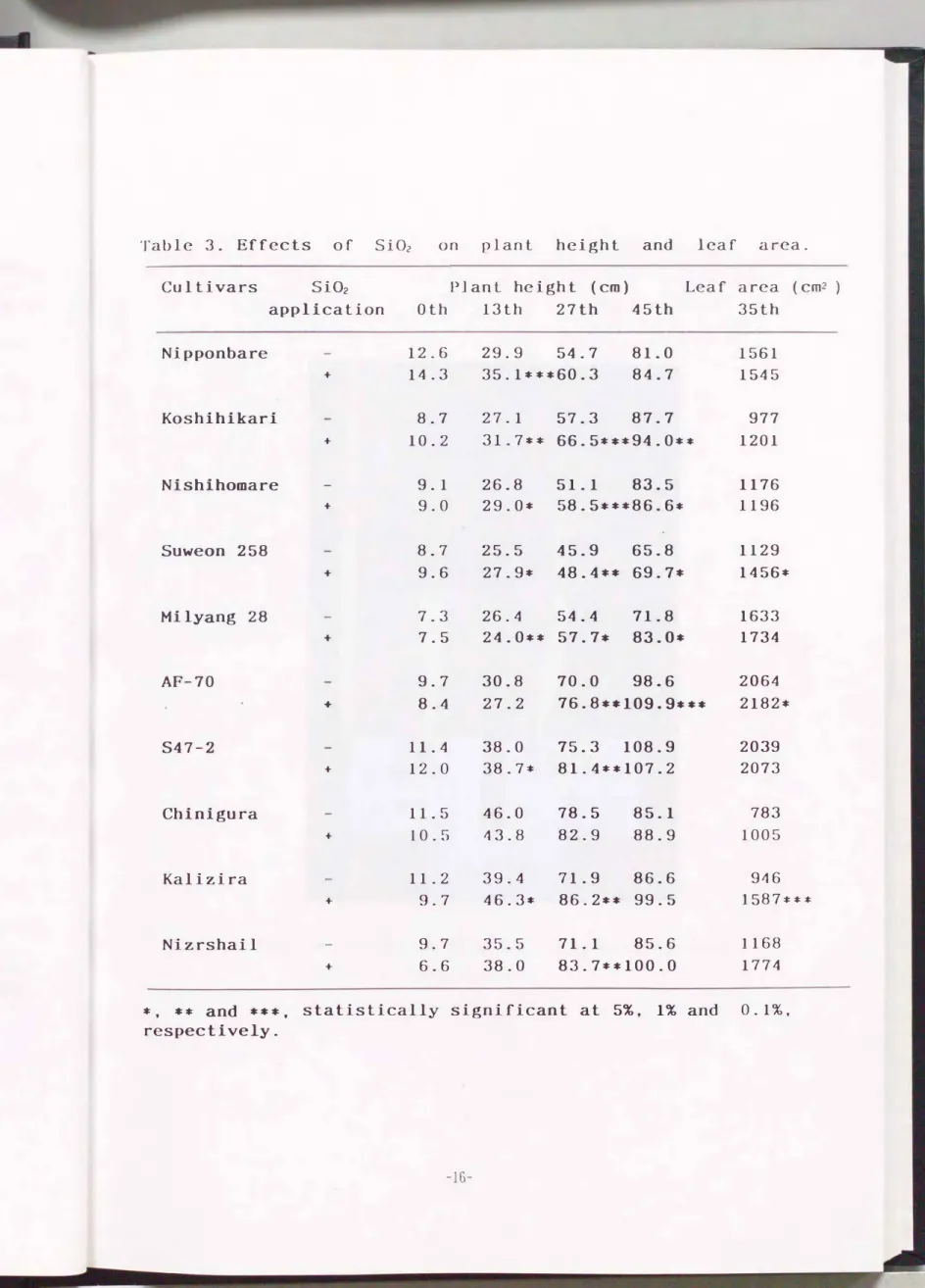
第3表にケイ酸処理後の草丈, 分げつ数および葉面積の経時的変化を 示した. ケイ酸施用の効果は草丈に顕著に現れ, + S i区は- S i区に 比べ処理後27日目で平均5%�15%増加した. 効果には品種間差があり,
①生育初期より効果がみられるもの(日本晴, コシヒカリ), ②処理後 20日前後からみられるもの(ニシホマレ, 密陽28号, AF-70)及び③効果 がみられないもの(S47-2, Chinigura)に大別された. 葉面積は水原25 8号, AF-70, およびKaliziraで効果が認められたが, 分げつ数にはどの 品種にも有意な効果はみられなかっ た.
第2図に最も生育差の顕著だったコシヒカリの生育状況を示した. 分 げつ数及び草丈に両区で明確な差異が認められた.
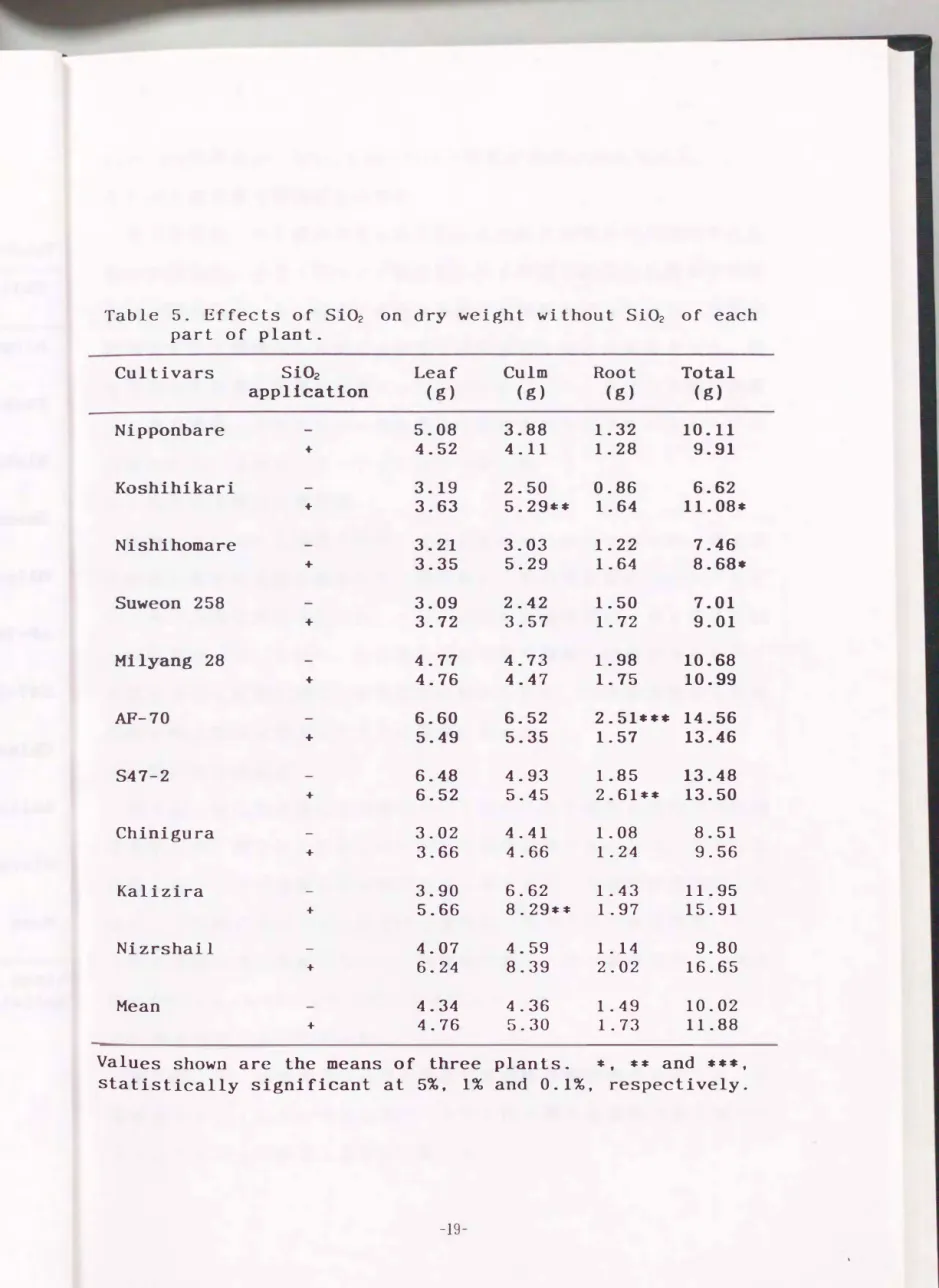
有機物蓄積量に対するケイ酸の効果をより厳密に検討するため, 乾物 重にケイ酸を含む場合(第4表)と含まない場合(第5表)とで比較し た. ケイ酸を含む場合, コシヒカリ, ニシホマレ, 水原258号, S47-2お よびKaliziraの5品種に施用効果がみられた. ケイ酸を除いた場合, 品 種では日本型のコシヒカリ及びニシホマレに, 器官別にはコシヒカリと
-1 4-
Table 2. SiOξ content in each part of plant.
Cul ti vars Si02 application
Leaf
(% )
Culm
(% )
Root
(%)
Total
(% )
Nipponbare
+
0.32 8.32
Koshihikari 0.05
9.81
+
Nishihomare 0.32
7.20
+
5uweon 258 0.32
7.62
+
Milyang 28 0.27
9.58
+
AF-70 0.37
8.38
+
547-2 0.41
7.40
+
Chinigura 0.26
5.46
+
Kalizira 0.35
7.25
+
Nizrshail 0.80
4.86
+
Mean 0.35
7.59
+
0.33 6.51
0.34 3.55
0.29 4.16
0.55 5.29
0.44 9.49
0.27 1.59
0.52 1.05
0.29 3.95
0.41 3.99
0.39 4.66
0.38 5.62
0.36 0.66
0.05 0.58
0.23 0.62
0.37 0.32
0.24 1.59
0.29 1.05
0.12 0.22
0.28 0.77
0.31 1.02
0.29 1.00
0.25 0.78
0.33 6.47
0.16 5.42
0.29 4.82
0.42 5.92
0.34 8.65
0.32 8.30
0.40 4.84
0.28 4.04
0.37 4.95
0.56 4.33
0.35 5.78 Si02 content was measured on 35 days after treatment.
Table 3. Effects of SiOz on plant height and leaf area.
Cultivars Si02 Plant height (cm) Leaf area (cm2) application Oth 13th 27th 45th 35th
Nipponbare 12.6 29.9 54.7 81.0 1561
+ 14.3 35.1本本*60.3 84.7 1545
Koshihikari 8.7 27.1 57.3 87.7 977
+ 10.2 31.7** 66.5***94.0*本 1201
Nishihomare 9.1 26.8 51.1 83.5 1176
+ 9.0 29.0本 58.5傘**86.6本 1196
Suweon 258 8.7 25.5 45.9 65.8 1129
+ 9.6 27.9本 48.4本* 69.7本 1456本
Milyang 28 7.3 26.4 54.4 71.8 1633
+ 7.5 24.0** 57.7* 83.0傘 1734
AF-70 9.7 30.8 70.0 98.6 2064
+ 8.4 27.2 76.8**109.9*** 2182本
S47-2 11.4 38.0 75.3 108.9 2039
+ 12.0 38.7本 81.4本*107.2 2073
Chinigura 11.5 46.0 78.5 85.1 783
+ 10.5 43.8 82.9 88.9 1005
Kalizira 11.2 39.4 71.9 86.6 94 6
+ 9.7 46.3本 86.2** 99.5 1587字本本
Nizrshail 9.7 35.5 71.1 85.6 1168
+ 6.6 38.0 83.7本*100.。 1774
本 . 傘本 and 傘傘本, statistically significant at 5%, 1% and 0.1%.
respectively.
-16-
Fig. 2. Effect of Si02 on the growth of rice plant (cv. Koshihikari)
Table 4. Effects of Si02 on dry weight of each part of planl.
Cultivars Si02 application
Leaf
(g)
Culm
(g)
Root (g)
Total (g)
Nipponbare
+
5.10 4.44
Koshihikari 3.20
4.60
+
Nishihomare 3.21
3.61本
+
Suweon 258 3.09
3.72
+
Milyang 28 4.77
4.76
+
AF-70 6.60
5.49
+
547-2 6.48
6.52
+
Chlnlgura 3.02
3.66
+
Kalizira 3.90
5.66
+
Nizrshail 4.07
6.24
+
Mean 4.34
4.76
+
3.89 3.97
3.14 5.55*
3.04 3.62*
2.42 3.57
4.73 4.47
6.52 5.35
1.42 1.10
1.15 1.65*
1.22 1.64
1.50 1.72
1.98 1.75
10.58 9.51
7.48 11.80*
7.46 8.68*
7.01 9.01
10.68 10.99
にUにucd89
凋官、υ
* ++ -' 唱止唱i 55 'i句,
勺ゐ唱i
4.93 1.85 13.48 5.45 2.61** 13.50
4.41 1.08 8.51 4.66 1.24 9.56
6.62 1.43 11.95 8.29字本 1.97 15.91
4.59 1.14 9.80 8.39 2.02 16.65
4.36 1.49 10.02 5.30 1.73 11.88
Values shown are the means of three plants. 傘 , 本本 and 字本牟,
statistically significant at 5%, 1% and 0.1%, respectively.
-18-
Table 5. Effects of Si02 on dry weight without Si02 of each part of plant.
Cultivars Si02 Leaf Culm Root Total appllcatlon (g) (g) (g) (g)
Nipponbare 5.08 3.88 1.32 10.11
+ 4.52 4.11 1.28 9.91
Koshihikari 3.19 2.50 0.86 6.62
+ 3.63 5.29*本 1.64 11.08*
Nishihomare 3.21 3.03 1.22 7.46
+ 3.35 5.29 1.64 8.68・
Suweon 258 3.09 2.42 1.50 7.01
+ 3.72 3.57 1.72 9.01
Milyang 28 4.77 4.73 1.98 10.68
+ 4.76 4.47 1.75 10.99
AF-70 6.60 6.52 2.51*** 14.56
+ 5.49 5.35 1.57 13.46
S47-2 6.48 4.93 1.85 13.48
+ 6.52 5.45 2.61本* 13.50
Chinigura 3.02 4.41 1.08 8.51
+ 3.66 4.66 1.24 9.56
Kalizira 3.90 6.62 1.43 11.95
+ 5.66 8.29本字 1.97 15.91
Nizrshail 4.07 4.59 1.14 9.80
+ 6.24 8.39 2.02 16.65
Mean 4.34 4.36 1.49 10.02
+ 4.76 5.30 1.73 11.88
Values shown are the means of three plants. 司�, 本本 and 傘$司1:,
statistically significant at 5%, 1% and 0.1%, respectively.
Kaliziraで得重に, S47-2とKaliziraで根重に効果が認められた.
3 )ケイ酸含量と乾物重との関係
第3図には, ケイ酸蓄慢量と各器官の乾物重との関係を品種間で比転 検討するためt + S i 区のケイ酸含量とケイ酸重を除去した器 官別 乾物 重との関係を示した. 図中の直線は各軸の平均値を示している. 器官別
乾物重と ケイ 酸含量 との間には有意な相関関係は認められなかった. 個 体全体の乾物重に効果が顕著だったコシヒカリ, ニシホマレの総乾物重 は. 第3象限. すなわちケイ酸含量及び乾物重ともに小さいグループに 分類された. 得重のパタ ー ンがこ れと一致した.
4 )光合成速度の品種間差
乾物生産 に お ける施用効果を, 光合成速度から検討 するため, 巌上 位 展開業の光合成速度を測定した(第6表) . 光合成速度にはバングラデ シュの3品種に効果がみられ, + S i区の光合成速度は- S i区より11
""4096高かった. し かし, 生長量及び乾物重で顕著な効果がみられた日 本型品種には処理区間で大きな差異は認められず, 光合成速度から乾物 生産の向 上効果を説明するこ と は 困難であった.
5 )個体光合成速度
第4 図には乾物生産に直後関 与 すると考 えられる 個体全体の光合成速 度を示した. 測定には乾物生産に顕著な施用効果のあったコシヒカリを
供試した. 光合成速度は葉面積当たりに換算した. 光強度の増加にとも ない, 光合成速度は + S i区で高く推移し, + S i区では直線型, - S i区では飽和型の曲線になった. 強光域で両区の差が顕著になり, 光強 度が1200μmolm-2s-1下では約27%の差となった.
6 )個体構造および吸光係数
第5 図には, 個体光合成速度 を 測定 した個体の個体情造を 示 した. 総 葉面積は+ S i区でわずかに高く, + S i区の積算葉面積の最大値が
S i区のものより群落上層部に位置した.
-20-
18 3
‘9 e6
...s e7
... � ・・3
2 OL
ぷZ喝圃.
OL ω て2
52ト
‘・4・
8
t出 e6
e2 e5
• 10
.7
:g 16 �・9
z
・214ト
匂
む12
�
E 川
8 e4旬81 0
」同
8
」瞳
7
3ムqv•
8
4
1 27 • e 10
.. 5
2
• e6
e 1
• 9
,・崎、む£ 1
も6ト
ω 主 て3と5ト
色刷伺
コ4ト 9
9
• 9
• 10
• 5
�
e8 7... 三
7ト
5←
(凶)2“一ω主h』℃E-2ハ》
3 .4
•
4
7
よ
6
e8
3 4
ム
8
e1
"e
.ム
6
..L
4
..L
3 2
0 9 10
Si02 content
(%) 5 8
10
Si02 content
(%)
weight of dry
and content Si02
between Relationship
Fig. 3.
Nipponbare; 2 rice cul ti vars. 1,
ten ln plant of
part each
ト1ilyang 28・
5,
258・
Suweon 4 ,
Nishihomare;
3,
Koshihikari;
Kalizira; 10,
9 Chinigura;
8,
S47-2;
AF70; 7,
6,
average the
indicate figure
the ln lines The
Nizrshail.
values.
Table 6. Photosynthetic rate of the youngest fully expanded leaf.
Cul ti vars Si02 application
Photosynthetic rate (Ilmo1m- 2 s- 1 )
Nipponbare 24 .81
+ 25.97
Koshihikari 25.97
+ 24 .92
Nishihomare 26.40
+ 23.78
Suweon 258 25.52
+ 25.24
Milyang 28 26.30
+ 23.79
AF-70 26.50
+ 26.65
S47-2 24.84
+ 23.09
Chinigura 1 4 .96
+ 20.29
Kalizira 15.74
+ 21.90
Nizrshail 16.79
+ 19.15
Measurements were made at saturated light intensity (above 1516 μmol・m-2・s- 1) t 35:t 1.6 C leaf temperature. 42.8:t10_8 % air humidity.
内/ω勺,ム
12
ふJω
伺 8
い /ー、、
u...-.
.� I
� ω ωp、J
,4C J 1 E 4
ロ rl
み0〉ms、 、2do E ' a,E
O 4
。
-4
o 500 1000 1500
Light intensity
(pr叫
Fig. 4. Response of top photosynthetic rate to light intensity in +Si (.) and -Si (0) plants.
intensity_
(% ) 40 60 light Relative
100 80 0 20
50 40
20 10 30
40
20 10 30
(日υ)
匂ロロ0・H凶
むk,O【何回
け伊耳切判ω国
0
1.5 1.0 0.5 。 0.5 1.0 1.5
weight area Dry
leaf Cumulated
plant- 1 ) (dm2 ) (g
structure plant
of Comparison Fig. 5.
1n +Si intensity
light rela ti ve
and
Open plants.
(lower) and -Si
(upper)
line solid
and bars,
dotted and
and dry area leaf
cumulated indicates
intensity,
light relative
-2 4- and
respectively.
weight,
第6図には植物体をいれない同化箱内の相対光強度を示した . 光源、 か ら離れるにつれ相対光強度は低下したがケイ酸処理区間で大きな差はな かった.
第7図は第5図に示した個体の積算葉面積の変化にともなう吸光係数 を示した. 吸光係数は+ S i区でO. 04低かった.
1 )主得着生葉身の光合成速度
第8図は, 個体光合成速度を左右する要因の一つである主稗着生葉身 の光合成速度を示した. 光合成速度は葉位の低下にともない低下した.
低下の度合は- S i区で大きく, 第4葉位の光合成速度は霞上位展開業 の約11% (+Si区)及び約12% (- S i区)であった.
8 )主稗着生葉身のクロロフィル含量
第9図には, 全供試品種の震上位展開葉から下位5葉身までのクロロ フィル含量を示した. 葉位が下がり老化が進行するにつれクロロフィル 含量は低下した. 低下 度は- S i区で大きかった. 下位葉身の低下度は 1 0品種中, コシヒカリで愚も顕著であった.
第2実験 異なる光環境下で生育させた水稲の生長 ・ 乾物生産に及ぼす ケイ酸の彫響
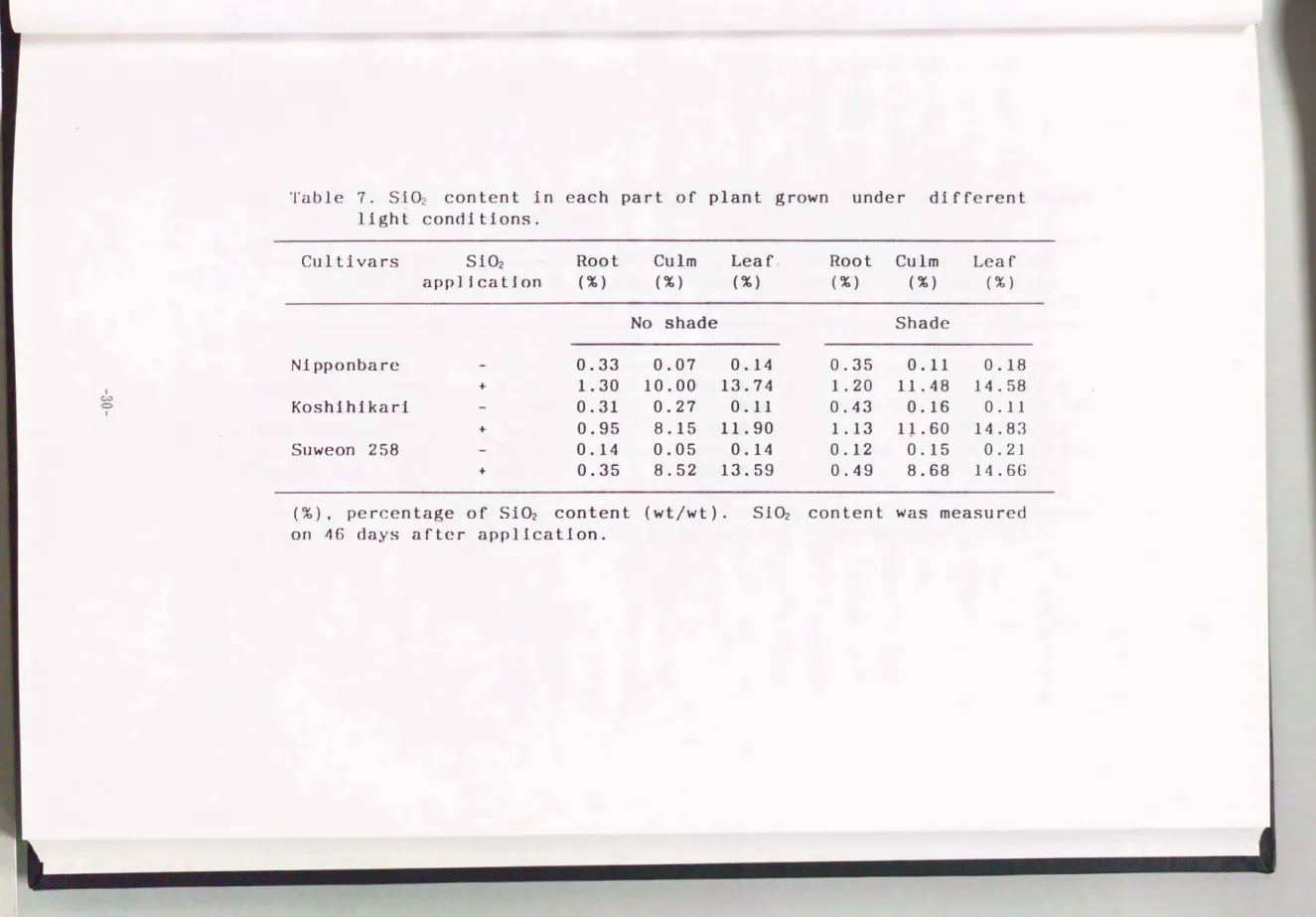
1 )各器官のケイ酸含量
ケイ厳処理後46日目の各器官ケイ酸含量を第7表に示した. ケイ酸含 量は実験lの場合と同様, いずれの品種でも葉, 茎, 根の順に多かった.
また, 含量は遮光処理によって変動し, 特に コシヒカリ稗中のケイ酸含 量が顕著に増加した.
2 )生長形質および乾物生産
第8表には, ケイ酸処理後46日目の草丈, 分げつ及び葉面積を示した.
無遮光区では, 日本晴の草丈と葉面積, コシヒカリの草丈および水原25 8号の葉面積にケイ酸の効果がみられ, + S i区の値は, 14.5%, 25.9%,
50
40
30
20 10
(Eυ)
匂ロ20・M凶
ωk〆。門出国
パVZ凶判む出
100 (%)
60 80
intensity
40
light
20
Relative
。
。
the ìntensity 111
light Relative
Fig. 6.
used ln chamber
assimilation empty
Fig. 4 symbols.
shown ln the for
-26- as 4 measurement
Fig.
See 5.
the and
Cumulated leaf
area(dm2)
。 1 2 3 4 5
0 5
0
5
0
ハU 守J
《J
勺4
1i可i
(収)kCJhmcωμロ刊μぷ∞刊.Hω〉JhμωHω出
-Si 0.40
Fig. 7. Relationship between cumulated leaf area and relative light intensity in +Si (・) and -Si ( 0 ) plants. Numerals are light extinction coefficient.
15
.w ω
い伺
(.)r-ア 1 0
OM I .w ω ..c: ω守、4
.w E ロ〉、 r-f Om 江E o 5
且, Cふο J
4
、』ノ
。
1 2 3 4
Leaf pos工t工on
Fig. 8. Photosynthetic rate in each leaf on the main culm in +Si (・) and
-Si ( 0 ) plants.
-28-
Ja仰nic.a Japoniω-indka lodica Javanica
6
「
4十
日略、
2
E
aE aB6
H
t E a
。 J
4 2
一て
4 -d ω
Eち
b
。
=
=
.. Kωhihikari
41ぃ�
2 3 4 56 42��て
2t
Ni目hih。田直也I
2 3 4 5 6
4
2
\,
2 3 4 5
L組f position from the youngest fully e玄関nded leaf
Fig. 9. Comparison of chlorophyll content in each leaf on the main culm between +Si (・) and -Si (0)
plants of ten rice cultivars.
Table 7. S102 content in each part of plant grown under different light conditions.
Cultivars Si02 Root Culm Leaf Root Culm Leaf appli.catjon (% ) (% ) (% ) (% ) (% ) (% )
No shade Shade
N1pponbare 0.33 0.07 0.14 0.35 0.11 0.18
+ 1.30 10.00 13.74 1.20 11.48 14.58
CC心D Kosh1h1kar1 0.31 0.27 0.11 0.43 0.16 0.11
+ 0.95 8.15 11.90 1.13 11.60 14.83
Sllweon 258 0.14 0.05 0.14 0.12 O. 15 0.21
+ 0.35 8.52 13.59 0.49 8.68 14.66
(%) , percentage of Si02 content (wtjwt). S102 content was measured on 46 days after application.