ウキクサ葉状体からの新規植物生育促進細菌の分離と性質
9
0
0
全文
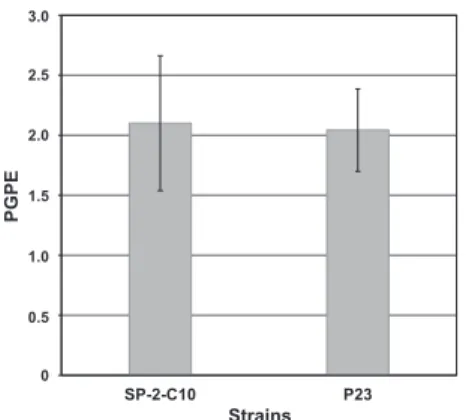
図


関連したドキュメント
As Riemann and Klein knew and as was proved rigorously by Weyl, there exist many non-constant meromorphic functions on every abstract connected Rie- mann surface and the compact
This paper presents an investigation into the mechanics of this specific problem and develops an analytical approach that accounts for the effects of geometrical and material data on
Although the Sine β and Airy β characterizations in law (in terms of a family of coupled diffusions) look very similar, the analysis of the limiting marginal statistics of the number
While conducting an experiment regarding fetal move- ments as a result of Pulsed Wave Doppler (PWD) ultrasound, [8] we encountered the severe artifacts in the acquired image2.
Moreover, in fashioning his theory of semisimple groups, Weyl drew on a host of ideas from such historically disparate areas as Frobe- nius’ theory of finite group characters,
廃棄物の再生利用の促進︑処理施設の整備等の総合的施策を推進することにより︑廃棄物としての要最終処分械の減少等を図るととも
職場環境の維持。特に有機溶剤規則の順守がポイント第2⇒第3
職場環境の維持。特に有機溶剤規則の順守がポイント第2⇒第3
