Title
離島 壱岐・対馬に生息するオバボタル(Lucidina biplagiata)の地理的分布及
び遺伝子解析
Author(s)
藤森 憲臣
Citation
福岡工業大学環境科学研究所所報 第11巻 P91-P98
Issue Date
2017-10
URI
http://hdl.handle.net/11478/788Right
Type
Departmental Bulletin Paper
Textversion publisher
福岡工業大学 機関リポジトリ
FITREPO
離島 壱岐・対馬に生息するオバボタル(Lucidina biplagiata)の地理的分布及び遺伝子解析
藤森 憲臣(福岡工業大学総合研究機構環境科学研究所) 藤井 直紀(日本離島の螢探検隊) 濵 茂久(壱岐・対馬離島の螢探検隊) 平田 秀彦(中部大学応用生物学部) 大場 裕一(中部大学応用生物学部)The diurnal firefly, Lucidina biplagiata, inhabiting in two islands of Genkai Bay,
Kyushu region of Japan, and its molecular
analysis
Abstract--- We investigated the distribution of the diurnal firefly, Lucidina biplagiata, in two islands of Genkai channel in Kyusyu region of Japan from May to June in 2016.
Molecular analyses revealed that the populations in Ikinoshima Islands and Tsushima Islands are grouped in a clade of the populations in Japan Mainlands.
Keywords:Lucidina biplagiata, Molecular analysis, Ikinoshima Islands, Tsushima Islands はじめに オバボタル Lucidina biplagiata(以下、本種)は、日 本に広く分布するホタル科マドボタル亜科 (Lampyrinae)の一種である。 タイプ標本の産地は Japon で、保管はロシア・モス クワ大学博物館でされている。 本種の分布は、千島列島及び日本(北海道、本 州、四国、九州、屋久島)、韓国([4]Kawashima, 2003)と広く分布が確認されている。 離島では、屋久島(南限)、対馬(北西限)、佐渡、 粟島、などからも記録がある。ただし、壱岐からは その記録がなかった([2]浦川,1994)。 島はその周囲を海に囲まれ、生息分布する移動 性の低い生物群では、独立進化する物理場環境 がそこに創生されやすい。 本種は全国各地に普通に見られる。標高の高い ところから低地に至る地域に分布する([4]大場, 2004)。 著者は、特に富士山では標高 1750m 地点及び南 アルプスでは 2000m 地点(しらびそ峠)の高地にて、 本種の生息分布を確認している。 本種の生息域は偏在している。林縁の草地に多 く、森林登山道や林道沿いの草地上にも見られる。 尾瀬ヶ原や釧路湿原、白神山地のブナ林にも生息 する([4]大場,2004)。著者らは、九州島内でも本種 のモニタリングをしており、生息域が偏在する事実 はこれまでの現地調査の知見とも合致する。 本調査では、玄界灘に位置する離島群より 2 島 を選定し、筆者らはヒメボタルの確認及び記録 ([12]藤森ら,2015)し、筆者らは本種の確認及び記 録([14]藤森ら,2016)した。2016 年度は、2 島にお ける本種の生息分布の確認を主目的とした。 オバボタルの外部形態による系統的位置づけ オバボタル Lucidina 属は現在、世界に 21 種と日 本には 4 種(オバボタル、オオオバボタル、コクロオ バボタル、ナツミオバボタル)が分布する。 本種は体長 10-11mm、頭部は黒色、前胸背は 黒色の中に赤紋、上翅は黒色である。本種に近縁 のオオオバボタル Lucidina accensa は体長 12- 14mm 程度で一回り大きく、頭部は黒色、前胸背は 黒色の中に大きな赤紋、上翅は黒色である([12]藤 森ら,2016)。
また、韓国に分布する Lucidina kotbandia は本 種に酷似している([7]Park et al,2005)。オバボタル とオオオバボタルの 2 種があるとされていたが、そ の後、韓国のオオオバボタルとされていたものはオ バボタルであるとされた(Medvedev and Ryvkin, 1992)。ただし、Kim et al.(1994)では 2 種の存在が 認められている。 過去長らく外部形態による分類から、朝鮮半島 の韓国産 Lucidina 属 L. kotbandia 及び日本産 Lucidina 属 L. biplagiata は、同種として扱われてい た。しかし、新種記載 L. kotbandia 報告([7]Park et al,2005)により、L. kotbandia 及び L. biplagiata は 別種として扱われることとなった。 調査地概要(壱岐・対馬) 長崎県管轄の離島で玄界灘の中央に浮かぶ主 な 2 島を「壱岐・対馬」と呼び、この 2 島は福岡・博 多港-韓国・釜山港の直線上に位置している。福 岡・博多港から、壱岐島は約 60km、対馬は約 120km の距離にある([13]藤森ら,2016)。(Fig.1) ・壱岐について 九州北方の玄界灘にある長崎県管轄の島で、 南北に 17 ㎞、東西に 14 ㎞、面積は 133.8km2、人 口は 26325 人(2017)である。 主島は「壱岐島」で、周囲には 23 の属島(有人 4、 無人 19)を所管する。 ・対馬について 九州北方の玄界灘にある長崎県管轄の島で、 面積は 708.5km2、人口は 32000 人である。主島は 「対馬島」で周囲には 100 以上の属島を所管する。 東海岸の一部及び下島の西海岸の一部を除く 全域でリアス式海岸が発達し、海岸線の総延長は 約 915km となる。 島の分水界は東に偏っていて、主要 6 河川のう ち 5 河川が西向きに流れる。各河川の下流部には 谷底平野があるものの、耕作に適した平地は非常 に少ない。 島の面積のうち、照葉樹を主とした森林(89%)で 占める。また、原始林及びソテツなどで天然記念物 に指定されており、自然豊かである。 動植物とも大陸系種及び対馬固有種、日本本土 系種が混在する、独特の生物相を形成している。 韓国から近くて行きやすい海外として、近年、韓 国人旅行者が対馬島内に非常に多くなっている。 材料及び方法 壱岐・対馬におけるホタル科昆虫の記録では、 対馬には固有種としてツシマヒメボタル及びアキマ ドボタルが生息し、市民にも重要種として広く認知 されている。また、出現情報などをメディアが特集 することで容易に耳に届く。
Fig.1 Map of Genkai Channel (Ikinoshima Islands and Tsushima Islands).
Red squares show the locations of current insect survey.
それに対して、壱岐では壱岐動物目録(1994)及 び壱岐高校公開のホタルマップ以外はホタル情報 が皆無に等しい([11]藤森ら,2016)。 2014 年まで、ゲンジ及びヘイケ、オオマドボタル 3 種の既知情報があった([2]浦川ら,1994)。筆者 らは、壱岐島にて本種の確認及び記録([14]藤森ら, 2016)した。2016 年度は、2 島における本種の生息 分布の確認を主目的とした。 (1)地理的分布 現地には当日夕方までに渡航・入島し、日中に 調査ルートを踏査した。その後、夜行性のホタル科 昆虫を日没直後より 26 時程度まで、本種を含む陸 生ホタル科昆虫の幼虫類の発光に関しては日没 後 21 時、23 時、25 時の 3 回に分け、ルートセンサ ス法(歩速:2km/h)により目視調査(見つけ捕りを含 む)を行った(Fig.2)。 また、本種(成虫)については翌朝 7-13 時まで の間に昼行性ホタル科昆虫(成虫)を対象種とし、 同様にルートセンサス法により目視調査を行った。 2016 年 5-6 月の定点調査では、捕獲し生体を 99.5%エタノール液浸し 4℃で保存した。 (2)遺伝子解析
DNA 抽出には、QIAamp DNA mini kit(キアゲ ン社)を用いてプロトコル通りの方法で抽出した。 ミトコンドリア DNA の COI 遺伝子を PCR により増 幅し、Dye terminator 法にて塩基配列を決定した。 DNA ポリメラーゼは,SpeedSTAR HS(タカラバイオ 社)を用いた。
得られた PCR 産物は、BigDye Terminator kit v 3.1 及び ABI Prism 3130(アプライドバイオシステム
ズ社)を用いたシークエンス法によりすべて両側か ら解析した。決定された塩基配列は、どの配列から も 1 塩基以上異なった場合 1 つのハプロタイプとし て分類し、MEGA3.1:Molecular Evolutionary Genetics Analysis software(Kumar et al.,2005)を用 いて分子系統解析を行った。 系統樹の構築には、近隣結合法(NJ 法)を用い 系統樹の各分岐の信頼性は 10000 回繰り返しによ るブートストラップテストによって評価を行った。推 測した。 結果及び考察 (1)地理的分布 対馬では、聞き取り情報([3]浦田ら,1999)を更 新して、本種個体の目撃及び捕獲した。 本種の生息分布では、壱岐・対馬の両島にて確 認でき、また両島内全域に広く分布していることを 明らかにした。ただし、各地点における捕獲個体数 (Table1,2)は極めて少ないため、生息に関わる絶 対個体数は低い可能性が推察される。 好適な生息環境では群生することがある本種の 個体数が、残念ながら本調査ではまとまった数で 確認できなかった。 その理由は 2 点考えられ、1)物理場環境が不十 分であること、または 2)繁殖好適時期を外している ことが挙げられる。 1)物理場環境が不十分 この点に関しては、筆者のこれまでの知見より 物理場環境を確認し総合的に判断した。
Fig.2 Survey method (Looking method)
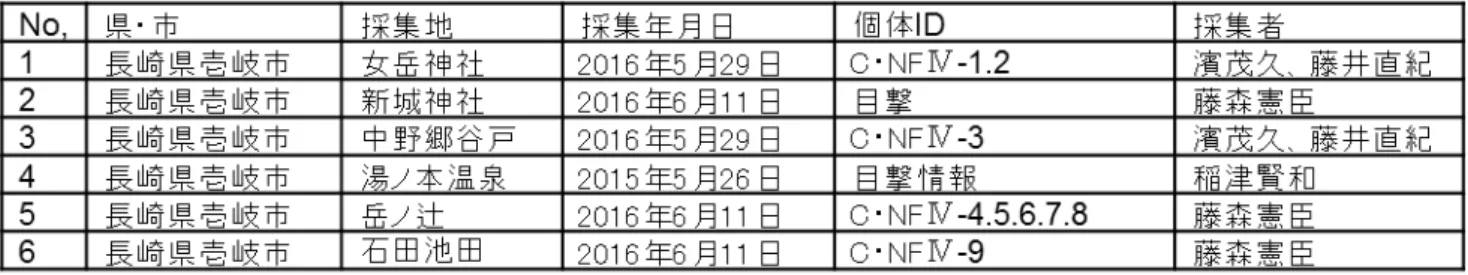
基本的に、環境依存度が高くない種であり「昼間 でも薄暗条件及び一定の湿度がある場所、林内の 薄暗条件下([4]大場,2004 )」がこれまでの調査研 究から明らかである。 現地調査の結果、壱岐・対馬の両島より本種生 息に好適環境が認められた(Fig.3)。 2)繁殖好適時期を外している 本種(成虫個体)の出現期は、一般に 5 月下旬- 8 月(低地:5 月下旬-6 月下旬、高地:6 月下旬-8 月)である([4]大場,2004)。 ただし、筆者は、4 月下旬-10 月上旬(低地-高 地を含む)の既知時期より広い期間で、これまでに 本種(成虫個体)を確認してきた経験がある。 今回、既知時期(低地:5 月下旬-6 月下旬)に調 査期を設定して現地実施してきたが、この結果を 受けて今後は可能な限り 4 月の中・下旬からの調 査を開始するべきである。 以下に、文献による既知情報「大場信義:ホタル 点滅の不思議([4]大場,2004)」を示し、新規情報を 含め整理する。 <オバボタル Lucidina biplagiata> 【形態】 全形:体長 約 10-11mm 【分布】 北海道(千島、樺太を含む)及び本州、四国、九州 (屋久島まで) 【出現期、活動習性】 出現期:4 月下旬-10 月上旬 (低地:5 月下旬-6 月下旬、高地:6 月下旬-8 月) 活動習性:昼行性(定位行動は、朝-夕方に確認) 午前中が活発であり、直射が当たる日中は活動が 低下する。晴天日よりも曇天日によく活動する。 【種同定の根拠】 壱岐から、オバボタル成虫を 9 個体捕獲(女岳神 社にて 2 個体及び中野郷谷戸にて 1 個体、岳ノ辻 にて 5 個体、石田池田にて 1 個体)した。捕獲個体 は全て♂個体である。(Table1) 島嶼個体の特徴は、本州及び九州産と比較する と体幅がひと回り(平均、0.3-0.5mm 程度)細い。 前胸背板の赤斑は、イ型及びロ型の 2 型分類 ([1]神田,1981)にならうと共に分布していた。その 中でも、赤斑が△に示されるイ型がほとんどであっ た([14]藤森ら,2016)。
Table 2. Collection and observation of L. biplagiata in Tsushima Islands
Fig.4 L. biplagiata (male) in Ikinoshima
対馬から、オバボタル成虫を 8 個体捕獲(舟志の 森にて 2 個体及び目保呂ダムにて 1 個体、ナムロ 水源の森にて 1 個体、ゆくみ水源の森にて 2 個体、 太祝詞神社にて 1 個体)した。捕獲個体は全て ♂個体である。(Table2) 島嶼個体の特徴は、本州及び九州産と比較する と体幅がひと回り(平均、0.3-0.5mm 程度)細い。 【形態差異のある Lucidina 属の目撃情報】 2016 年度に引き続き、2017 年度も 5 月中旬に壱 岐・対馬にて、本種成虫のモニタリング調査を実施 した。その際、対馬下島・加志地区、太祝詞神社境 内にて本種酷似した個体を目撃した。このことは、 仮説としている韓国に分布する L. kotbandia が対 馬においても生息している可能性を否定できない。 (2)形態比較と遺伝子解析 壱岐における調査地の数ヶ所から本種と考えら れる個体を採集して報告した([14]藤森ら,2016)。 今回はさらに、対馬における調査地の数ヶ所から 本種と考えられる個体を採集したことを報告する。 ただし、韓国産オバボタル属が別種 L. kotbandia として記載されている([7]Park et al.,2005)ことで、 壱岐・対馬の本種個体が本当にオバボタル L. biplagiata と言えるかどうかが問題である。 そのため、形態比較(Fig.5)と遺伝子解析により、 壱岐・対馬の本種個体が L. biplagiata なのか L. kotbandia なのかを考察する必要がある。 既に、本種とは別種として記載されている韓国 産 L. kotbandia は、その種の遺伝子解析がなされ Genebank 登録により情報開示されている。 そこで、中部大学応用生物学部で解析が進めら れている全国各地のサンプルデータとして本種及 び L. accensa、L. kotbandia の遺伝子と比較して、こ れまでに明らかになっている情報を系統樹(Fig.6) として評価した。
Fig.5 Different shape in the pronotum of
firefly (L. biplagiata)
解析には、壱岐、対馬産の本種の他、愛知、熊 本県産の本種と愛知県産の L. accensa(Fig.7)、 GenBank に登録されている L. kotbandia の COI 部 分配列 608 塩基を用いた。
また外群には、同じマドボタル亜科に属する比 較 的 近 縁 な 埼 玉 県 産 Pyropyga sp. ( GenBank accession number, AB608774 ) 及 び カ ナ ダ 産
Lucidota atra(KR486033)を加えた。 マルチプルアラインメント並びに近隣結合法によ る 系 統 解 析 に は ClustalW ( デ フ ォ ル ト 設 定 、 http://clustalw.ddbj.nig.ac.jp)を用いた。 解析の結果、壱岐、対馬産の本種は愛知県、熊 本県産の本種と単系統群を作り、近縁種であった。 L. accensa 及び L. kotbandia はその外側に位置す ることがわかった。 このことは、壱岐、対馬に産する個体は本州、九 州産と同じ本種であり、L. accensa や L. kotbandia とは明らかに異なることを示している。 分 子 系 統 解 析 に 用 い た 対 象 サ ン プ ル で 、 L. accensa についても文献による既知情報「大場信 義:ホタル点滅の不思議([4]大場,2004)」を記す。 <オオオバボタル Lucidina accensa> 【形態】 全形:体長 約 12mm 【分布】 北海道(千島、樺太を含む)及び本州、四国、九州 (屋久島まで) 【出現期、活動習性】 出現期:6 月-8 月の間に記録がある。 活動習性:昼行性(定位行動は、朝-夕方に確認) 午前中が活発であり、直射が当たる日中は活動が 低下する。晴天日よりも曇天日によく活動する。 まとめ (1) 地理的分布 本種の分布に関して、以下の 4 項目を確認した。 ・壱岐・対馬の両島各定点にて、オバボタル成虫を 目視確認及び捕獲した。 ・生息環境として、昼間は薄暗条件及び一定の湿 度がある場所、林内の薄暗条件下である。水分量 が補完され、下草の繁茂環境を好んでいる。 ・生息密度は低い。 オバボタル成虫が確認される生息地は局所的で あることが多いが、生息適地の場合には群生する 特徴もある。今後、生息地のモニタリング調査を実 施することで明らかにしていく必要がある。 ・活動習性は、成虫において昼行性(朝-夕方)で 晴天時に比較して、曇天時にはより活動が活発化 ([4]大場,2004)とされている。 本調査でも、7-13 時までの間で雨天前後の湿 度が高い条件時に本種の飛翔が確認された。多 発生、多飛翔している期間は、環境条件さえ整って いれば、日没前の 18 時半まで活動が確認された。 また、本種成虫の夜間活動は十分な調査法(ス ウィーピング法(Fig.8)その他を実施できておらず、 目視だけでは個体が確認できない可能性が高い。
Fig.7 L. accensa (male) in Kyusyu(FUKUOKA)
そのため、一般的に活動していないとされるが、日 没直前まで観察される事例があるため、夜間活動 の可能性も否定できない。 (2) 形態比較と遺伝子解析 本種の形態及び遺伝子解析に関して、以下の 4 項目を確認した。 ・壱岐、対馬産の本種は、愛知県、熊本県産の本 種と単系統群になり近縁種と推察される。
・L. accensa と L. kotbandia は、本種 L. biplagiata のクレードの外側に位置した。
・壱岐、対馬に産する個体は本州、九州産と同種 (本種 L. biplagiata)である。
・L. accensa や L. kotbandia は、本種 L. biplagiata とは明らかに異なることを示している。 今後の予定 発光しないホタル科昆虫の成虫も調査を実施し てその生態を解明する必要があり、夜間の調査法 を再度検討していくべきである。 また壱岐・対馬の他に、日本産 Lucidina 属(L. biplagiata 及び L. assensa、コクロオバボタル L. okadai ) を 広 く 遺 伝 子 解 析 し 、 さ ら に 韓 国 産 L. kotbandia の解析情報を取得し、比較する予定であ る。 L. kotbandia は、前胸背板の紅紋が日本産 L. biplagiata に 比 べ て 非 常 に 小 さ い と さ れ る が ([7]Park et al.,2005)、日本本土及び離島にも紅紋 が小さい個体はこれまでにも観られる。 また、♂個体交尾器の先端がフック状になるも のが L. kobtandia であるとされる([7]Park et al., 2005)。しかし、この点も日本産個体を広く調べて みる必要がある。
ただし、遺伝子解析の結果でみると、韓国産 L.
kotbandia は日本産 2 種(L. biplagiata、L. assensa)
とはかなり離れていることは確認されている。 対馬には、本種の外部形態と異なる Lucidina 属 個体の目撃もあることから、より詳細な生態調査及 び遺伝子解析を必要とする。 謝辞 本調査を遂行するにあたり、まず本報告の基と なる貴重な目撃情報の提供について稲津賢和氏、 そして壱岐における調査協力者(特に、永村春義氏 及び植村圭司氏)に深く感謝申し上げる。 離島における調査の場合は連日連夜の不規則 なスケジュールになる上に、島間渡航を含んだ天 候等による変則的な日程での調査実施になること が多々ある。 その場合、各島にてお世話になる宿泊施設運営 者(壱岐島:民宿明るい農村,対馬:ホテル美津和 館,原旅館、ペンションひのきの森,万松閣,喫茶 ひまつぶし[食事])の方々のご厚意及びご理解、ご 協力なしには達成なし得なかった。 その他、日頃よりサンプル管理をはじめ、遺伝子 解析において全面的にご協力いただいている中部 大学応用生物学部・発光生物学研究室所属の各 位に深く感謝申し上げる。また、本研究は日本離 島の螢探検隊、藤森文臣氏からの助成(2016)を 受けて行われているもので、ここに謝意を表す。 引用及び参考文献 [1] 神田左京: ホタル; サイエンティスト社 97-110 (1981) [2] 浦川虎郷ら: 壱岐島動物目録; 壱岐「島の科 学」研究 64(1994) [3] 浦田明夫,國分英俊: 対馬の自然-対馬の自 然と生きものたち-; 杉屋書店,対馬の自然誌 Ⅱ (1999)
[4] Kawashima, I., H. Suzuki and M. Sato. A Check-list of Japanese Fireflies (Coleoptera, Lampyridae and Rhagophthalmidae). Jap. J. Sys. Ent., 9(2): 241-261. (2003) [5] 大場信義: ホタルの点滅の不思議-地球の 軌道-; 横須賀市自然・人文博物館,特別解 説書 7 158-160, 172 (2004) [6] 大場信義: オバボタルとオオオバボタルの形 態と色彩斑紋パターン; 横須賀市博研報(自 然),48,117-130 (2001)
[7] Park et al: Lucidina kotbandia a New Species (Coleoptera: Lampyridae) from Korea; 2005) [8] 小俣軍平ら: 日本産ホタル 10 種の生態研究; 板当沢ホタル調査団・船舶印刷, 120-125(2006) [9] 旅する長崎学 12;長崎文献社,53 (2009) [10] 藤森憲臣ら:伊勢・三河湾の離島に生息 するホタル科昆虫;名古屋産業大学論集 (環境情報ビジネス学会)」 25 号,17-22 (2014) [11] 藤森憲臣ら:伊勢・三河湾の離島に生息 するヒメボタル(L. parvula)の遺伝子解析 (予報);名古屋産業大学論集(環境情報 ビジネス学会)」 25 号,67-69 (2015) [12] 藤森憲臣ら:離島 壱岐島に生息するヒメボタ ル(L. parvula)の新産地記録;名古屋産業大 学論集(環境情報ビジネス学会)」 26 号, 43-46 (2015) [13] 藤森憲臣ら:壱岐・対馬の離島に生息するホ タル科昆虫について;福岡工業大学環境科学 研究所報・環境研究発表 2016,77-81(2016) [14] 藤森憲臣ら:離島 壱岐島に生息するオバボ タル(L. biplagiata)の新産地記録;名古屋産業 大学論集(環境情報ビジネス学会)」 28 号, 25-28 (2016) (平成 29 年 9 月 11 日受付)