修 士 学 位 論 文
題 名
小笠原固有種オオシラタマカズラの 二型花柱性とその繁殖に関する研究
指 導 教 員 菅 原 敬 准 教 授
令 和 2年 1月 10日 提 出
首都大学東京大学院
理 学 研 究 科 生 命 科 学 専 攻 学修番号 18846436
氏 名 宮 川 彩 花
1 目次 要旨
Ⅰ. はじめに
Ⅱ. 材料と調査地
Ⅲ. 調査方法
①花粉サイズ測定
②柱頭上に付着する花粉の比較
③人工授粉実験
④野外での結果率調査
⑤蜜量の調査
⑥訪花昆虫の調査
⑦統計解析方法
Ⅳ. 結果
①二型花の花粉サイズ
②二型花の柱頭上花粉量と花粉由来
③二型花の人工授粉実験
④野外での二型花の結果率
⑤蜜量
⑥訪花昆虫
Ⅴ. 考察
1)二型花の柱頭上花粉 2)二型花の授粉実験 3)送粉昆虫
4)野外におけるオオシラタマカズラの繁殖の実態
Ⅵ. 結論と今後の課題
Ⅶ. 謝辞
Ⅷ. 参考文献
Ⅸ. 図表
2
A Study on Distyly and Reproduction of Psychotria boninensis Nakai (Rubiaceae), Endemic to the Bonin (Ogasawara) Islands
Distyly is a genetically controlled floral dimorphism in which a plant population is composed of two floral morphs: long (L)-styled and short (S)-styled morphs. These floral morphs usually differ in stigma and anther heights from each other and show self- and intramorphic incompatibility. Therefore, this floral dimorphism is regarded as a mechanism for promoting cross-pollination between the two morphs, mediated by insect pollinators.
Psychotria boninensis Nakai is a perennial liana endemic to the Bonin (Ogasawara) Islands, Japan. Its plants open small-cup shaped flowers less than 1cm long from May to June, and mature with white drupes from September to October. In a previous study it was reported that the flowers were morphologically distylous with L- and S-styled morphs, and show self- and intramorphic incompatibility. Moreover, their fruit set percentages were usually more or less 10% in open pollination, and the fruit set percentages of the S-styled flowers were usually lower than those of the L-styled flowers. I supposed that stigma clogging influenced the fruit set, especially of the S-styled flowers. In case of a cup-shaped flower such as Psychotria boninensis, self-pollens may inhibit pollination from the stigma by falling pollens (stigma clogging). In order to well understand reproductive nature of the distylous plants in field, I investigated the following three items for two morphs: 1) quantity of pollen grains attached on stigma, 2) artificial pollination experiments, 3) examination of the fruit set percentages of open pollination and pollination system.
I obtained the following results. 1) The intramorph pollen grains were dominant on both stigmas of L- and S-styled flowers. 2) The fruit set percentages of intermorph cross- pollination after intramorph cross-pollination were lower than those of intermorph cross- pollination, suggesting that stigma clogging has been caused by intramorph pollen grains on stigma of P. boninensis. 3) The fruit set percentages in open pollination were about 10%, indicating that the percentages were very low. Although the flower visitors effective for pollination were supposed to be flying insects, they were very rare in field and honey bees and small moths were occasionally found in natural population.
These results suggest that in flower dimorphism of P. boninensis, stigma clogging may be cause by attachment of intramorph pollen grains in natural population. But, lack of effective pollinator and pollinator shift from original insect fauna influence the fruit set percentages in the present Bonin Islands more strongly.
3
学位論文要旨(修士(理学))
宮川 彩花 小笠原固有種オオシラタマカズラの二型花柱性とその繁殖に関する研究
野外において複数の種が同所的に生息するとき、種間で繁殖干渉が生じて繁 殖成功に影響を及ぼすことがある。繁殖干渉とは「配偶過程において適応度の低 下をもたらすあらゆる種類の種間相互作用」を指し、植物の場合には、他種花粉 が柱頭を覆い隠してしまい同種花粉での受粉ができなくなるstigma cloggingな どが知られている(高倉 2010)。しかし、同種内においても、例えば二型花柱性 を示すような植物では、二型花が自家・同型花不和合性を示すことから、二型花
間でstigma cloggingのような現象が生じている可能性が考えられる。その為、
本 研 究 で は 小 笠 原 固 有 の 異 型 花 柱 性 オ オ シ ラ タ マ カ ズ ラ(Psychotria boninensis)を用いてstigma cloggingの可能性に着目しながら、本種の繁殖の 実態をより明らかにすることを目的として、以下のような調査を進めた。なお、
本種は先行研究より、短花柱花と長花柱花を持つ二型花柱性で自家・同型花不和 合性を示し、短花柱花の結果率が有意に低いことが報告されている。そこで、1)
二型花の花粉サイズの違いに着目して、柱頭上に付着する花粉量を二型花間で 比較する。2)二型花間の授粉実験、特に同型花の花粉受粉後に異型花の花粉を 受粉した場合に結果率が低下するか確認する。3)野外での結果率と送粉昆虫の 実態を解明する、などを行なって上記の課題について検討することを進めた。
調査の結果、1)二型花の花粉サイズは明瞭に異なるため、そのサイズに着目 して、野外で柱頭上に付着する二型花の花粉量を比較したところ、いずれの花粉 においても同型花の花粉が優占していることが確認された。2)柱頭に同型花の 花粉受粉後に異型花の花粉を付着させた花では、異型花授粉と比較して明瞭に 結果率が低下した。これは実際にstigma cloggingが起こっていることを示唆す る事例である。3)野外での二型花の結果率は、2018 年調査で 10%前後と低い 値であった。一方訪花昆虫は、セイヨウミツバチに加えてアリやカメムシ、トビ ムシ等の訪花が確認されたが、アリを除いて全体的に訪花頻度が低く有効なポ リネーターが欠如している状況であった。
以上の結果を総合的に考慮すると、オオシラタマカズラの二型花では、野外で 同型花の花粉の付着が頻繁に起こり、stigma cloggingのような現象が生じてい る可能性も示唆されるが、現在の小笠原諸島父島においては、ポリネーターの欠 如が結果率により強く影響している可能性が考えられる。
4
Ⅰ.はじめに
被子植物では様々な繁殖様式が見られるが、最も複雑な様式として二型花柱性が知
られている。二型花柱性 distyly とは、葯の位置が低く柱頭が高い長花柱花(L 型)
と、葯の位置が高く柱頭が低い短花柱花(S型)の二型花をそれぞれ別個体につける 繁殖様式である。この様式では、葯と柱頭の高さが二型花間相互で対応しており、一 般に自家・同型花不和合性を示す(Ganders 1979, Barrett 1992, 菊沢 1995)。従って 二型花柱性は他家受粉を促進する繁殖様式であり(西廣 2000, 鷲谷 2006)、種子を 形成するためには花粉媒介者による異型花間での花粉の移動が必要となる(Beach
and Bawa 1980, 鷲谷 2006)。なお、この二型性は遺伝的に制御されている。柱頭と
葯の相互の位置関係や花粉サイズなどの性質を支配する遺伝子は、スーパージーンモ デルで代表されるように互いに強く連鎖し、L型は劣性ホモ、S型はヘテロの遺伝子
型のである(Barrett 1992)。二型花は異型花間での交配を行なうため、L型とS型の 個体が1:1の分離比で生じる(西廣 2000, 鷲谷 2006)。
小笠原諸島固有のオオシラタマカズラ(Psychotria boninensis, アカネ科)の花は、
形態的に葯と柱頭の高さが相互に異なる二型性で、長花柱花(L型, Fig. 2A)と短花 柱花(S型, Fig. 2B)を持つことが報告されている(Kondo et al. 2007)。また、S型 の花粉サイズが L 型に比べて大きいことも報告されている。二型花のいずれでも花 粉は高い染色性を示したことから、L型もS型も機能的には両性であることが示唆さ
5
れている(Kondo et al. 2007)。さらに人工授粉実験より、自家・同型花間授粉では結 実しないことから自家・同型花不和合性を示すことが報告されている(Sugawara et
al. 2014)。この結果より、オオシラタマカズラの二型花は機能的に両性であるが、野
外において自然結果率が10%前後と低く、またS型の結果率が L型に比べて有意に 低いという現象が報告されている(Sugawara et al. 2014)。このS型花での結果率の 低さの原因として、S型からL型への片方向の送粉が優先されている可能性が挙げら れている(Sugawara et al. 2014)。一方で、本研究では、stigma cloggingが生じてい る可能性があるのではないかと考えた。
Stigma clogging とは、他種花粉や不和合性を示す自花粉など、受粉できない不適
合花粉が柱頭を覆い隠してしまい、本来受粉するべき、適合花粉での受粉ができなく なってしまう現象を指す(Holland and Chamberlain 2007)。過去の研究において、ポ リネーターが同じ花を複数回訪れることにより、不和合性を示す自花粉が柱頭に付着 し、stigma clogging が生じて結果率が低下することがキク科植物などで示唆されて いる(Ehlers 1999)。二型花柱性において、S型の花では、柱頭より葯が上に位置し ているため、上の葯から不適合な自花粉が柱頭へこぼれ落ち、柱頭を覆ってしまうこ とで、異型花からの花粉が受粉しにくくなっている可能性がある。しかし、これまで 二型花柱性の植物において、このような特性に着目した研究はあまり行なわれていな い。
本研究では、小笠原諸島の固有種であるオオシラタマカズラを用いて stigma
6
cloggingの可能性に着目しながら、本種の野外における繁殖の実態をより明らかにす
ることを目的とした。そのために、1)二型花の花粉サイズの違いに着目して、柱頭 上に付着する花粉量の二型花間での比較、2)二型花間の授粉実験、特に同型花の花 粉受粉後に異型花の花粉を受粉した場合に結果率が低下するか否かの確認、3)野外 での結果率と送粉昆虫の実態の解明、を行なった。
Ⅱ.材料と調査地
オオシラタマカズラ(Psychotria boninensis Nakai)は、アカネ科ボチョウジ属に 属する小笠原諸島固有のつる性木本植物である。父島、兄島、弟島、母島、向島など の山地帯照葉樹林の林床や林縁などに多く自生している。5~6 月頃、集散花序を形
成して 1cm に満たない小さな筒状の花を開き、9~10 月頃結実する。個々の花は小 さな筒状で、朝に開花し、夕方から夜には花筒が落ちてしまう1日花である。個々の 花序は、毎日数個の花を開く(豊田 2014)(Fig. 1)。
調査は小笠原諸島父島の巽道路終点付近にて行った。開花期と結実期に合わせて年
2回、2018年6月2日~12日、8月18日~26日、2019年6月8日~30日、10月 2日~14日、現地にて調査を行った(Fig. 2)。
調査対象とした個体は、道路南側では林床内から道路脇にかけて地面上に生育し
7
(Fig. 1C)、北側では地面から1-2m程度の斜面上に生育していた(Fig. 1D)。約10m 範囲にL型とS型が生育し、一部は混在していた。周囲には、ムニンヒメツバキ、オ オバシマムラサキ、ムニンアオガンピ、ウラジロエノキ、オオバナセンダングサ、メ ドハギ等の開花した個体が生育していた。
野外における L 型と S 型の位置関係と結果率を調査するために、調査地の模式図 を作成した。
Ⅲ.調査方法
①花粉サイズ測定
オオシラタマカズラの花粉は球状で3孔粒である(Kondo et al. 2007)。L型花粉 とS型花粉の範疇を決めるために、花粉サイズ(発芽孔を通過する、花粉外膜の直径)
を測定した(Fig. 2C)。2018年6月に70%エタノールで液浸標本とした花から葯を 取り出し、ラクトフェノールコットンブルー染色液を滴下し、葯から花粉だけを取り 出してプレパラートを作成し、半日以上染色した。その後、光学顕微鏡(BH-2;
OLYMPUS)を用いて、1 つのプレパラートにつき約 20 粒ずつランダムに選び、L
型5花135粒、S型4花118粒、花粉サイズを測定した。
8
②柱頭上に付着する花粉の比較
野外における花粉の付着状況を調査するため、柱頭上に付着する花粉を二型花間で 比較した。葯の花粉が付着しないよう気を付けながら、野外で開花した花から当日の 午後に柱頭をピンセットで採取し、ラクトフェノールコットンブルー染色液を滴下し て、柱頭を軽く押しつぶしながらプレパラートを作成した。その後、染色液が蒸発し ないようマニキュアで封入し、半日以上染色した。その後、光学顕微鏡(BH-2;
OLYMPUS)を用いて柱頭上の花粉サイズを測定し、事前に測定した花粉サイズの頻 度分布に基づいてL型花粉・S型花粉に分類した。2018年6月にL型6花、S型10 花、2019年6月にL型18花、S型14花採取した。
③人工授粉実験
Stigma cloggingの可能性を探るために、人工授粉実験を行った。2018年6月2日
~12 日、2019年6月 8日~30日に、次のような合計8パターンの組み合わせで交 配実験を行った。同型花間授粉(①L×L、②S×S):L型19花、S型23花を用いた、
異型花間授粉(③L×S、④S×L、): L型26花、S型31花を用いた、同型花間授粉 後に異型花間授粉(人為的stigma clogging)(⑤L×L×S、⑥S×S×L): L型43花、
S型25花を用いた、異型花間授粉後に同型花間授粉(⑦L×S×L、⑧S×L×S)L型 34花、S型23花を用いた。⑦、⑧の異型花間授粉後に同型花間授粉する交配実験は 2019年のみ行い、それ以外の交配実験は2018年、2019年ともに実験を行った。交
9
配実験を行う花は、事前に花の蕾を含む花序にネットをかけておき、翌日の午前中に ネットの中で開花した花を用いた。花序内で開花していない蕾は摘み取った。授粉処 理の際は、柱頭に花粉が付着しないよう気を付けながら花筒と葯を全て取り除き、柱 頭を剥き出しの状態にした。その後、ピンセットを用いて裂開している葯をつまみ花 粉を柱頭につけた。授粉実験を行った花は不織布(7cm×7cm)で覆い、近くに処理 内容と花数を記したタグ(1cm×3cm)を針金でつけた。その際処理を行った花の総 数も記録した。
④野外での結果率調査
自然状況下における結実の状況を調べるために、L花とS花それぞれ花序をランダ ムに選び、花数を数えて花数を記したタグをつけた。2018 年 6 月 2 日~12 日の間 に、L型19花序 870花、S型9花序342花を、2019年6月8日~30日の間に、L 型12 花序560 花、S型10 花序416 花を調査した。これらの花の結実、そして人工 授粉実験による結実は、それぞれ2018年8月18日~26日、2019年10月2日~14 日に現地へ行き、果実形成の有無を記録した。そして、果実を形成した花数÷調査し た花数を結果率として算出した。なお、自然結果を調査した花序の位置はGPSで記 録し、地図上にプロットした。
10
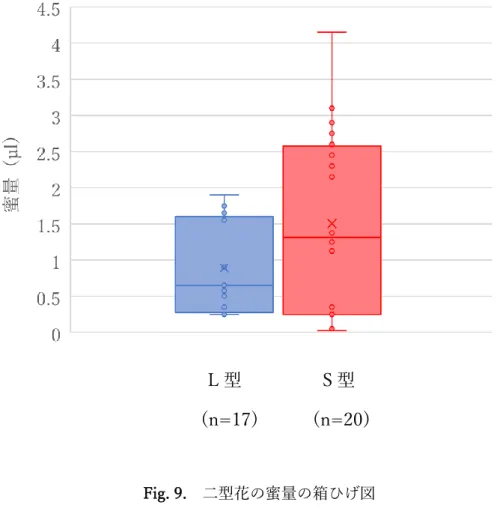
⑤蜜量の調査
蜜量の測定は2018年 6月2日~12日、2019 年6月8日~30日に行った。0.5µl マイクロチューブ(minicaps®, HIRSCHMANN)を用いて花の基部にある蜜の採取を 行った。それぞれL型17花、S型20花を調査した。
⑥訪花昆虫の調査
送粉昆虫を明らかにするために、本種の花に訪花する昆虫の調査を行った。デジタ ルカメラを用いたインターバル撮影による観察と、目視による訪花者の観察を行った。
1~2台のデジタルカメラを花序の前に設置し、昼頃から翌日まで約 24 時間、2分間 隔で連続的に撮影した。夜間も撮影できるようフラッシュを併用した。2019年 6月 8日~30日にL型9花序を計323時間、S型14花序を計206時間調査した。
⑦統計解析方法
全ての統計解析には統計解析ソフトの R ver.3.6.1 を利用した。花粉サイズ、蜜量 を2型花(長花柱花、短花柱花)間で比較する際にはWilcoxonの順位和検定を行っ た。結果率を2型花間、群間で比較する際には、Fisherの正確確率検定法を用いた。
11
Ⅳ.結果
①二型花の花粉サイズ
測定した二型花の花粉サイズでヒストグラムを作成したところ(Fig. 4A)、二型花
の花粉サイズは明瞭に異なり、L型花粉(平均:34.56µm)がS型花粉(平均:61.67µm)
より有意に小さかった(Fig. 4B, Table 1)(Wilcoxonの順位和検定, p<0.001)。また、
L型花粉(SD:1.97)はS型花粉(SD:4.11)と比べてバラつきが小さく、全て 40µm 以下であったため、本研究では、40µm 以下の花粉をL型花粉、40µm より大きい花 粉をS型花粉と定義した。
②二型花の柱頭上花粉量と花粉由来
二型花の柱頭上の花粉量を、L型とS型、それぞれ24花ずつ比較した。花粉の総 付着数は、L型675粒、S型204粒で、L型の方が多かった。付着花粉のサイズでヒ ストグラムを作成したところ、L型とS型いずれの柱頭においても、幅広い花粉サイ ズの柱頭への付着が見られた(Fig. 5)。
花粉の付着が見られた柱頭は、2018 年は L型 6 花(100%)、S 型 4 花(40%)、 2019年はL型14花(77.8%)、S型5花(35.7%)であった。合わせると、L型は20 花(83%)、S型は9花(38%)の柱頭に花粉の付着が見られ、S型の割合が有意に低 かった(prop.test, p<0.001)。先に述べた花粉サイズの柱頭に従って、個々の柱頭に
12
付着していた花粉を分類すると、Fig. 6,7のような結果となった。花粉の付着がある 柱頭では、L型、S型いずれにおいても同型花の花粉が優占していた。
③二型花の人工授粉実験
調査した年度ごとの交配実験の結果率と、それらを総合した結果率をTable 2に示 した。結果率調査の際に個体の枯死が見られたものや、花序が風で吹き飛ばされてい たものは、枯死数としてカウントし、実験した花数から除いて結果率を出した。
2018年の調査において、L型(7花)とS型(3花、内1花枯死)は同型花間の交
配実験では全く結実しなかった。一方、異型花間の交配実験では L型(18 花)を母 親とした時には88.9%で結実が得られ、S型(6花)を母親とした時には100%結実 した。二型花間の結果率に統計的に有意な差は認められなかった(Fisherの正確確立 検定, p=1)。また、同型花の花粉授粉後に異型花の花粉を授粉した交配実験では、L
型(13花、内3花枯死)を母親とした時には60%が結実し、S型(10花)を母親と した時には 40%で結実が得られた。二型花間の結果率に統計的に有意な差は認めら れなかった(Fisherの正確確立検定, p=0.656)。また、L型とS型いずれの花におい ても、各交配実験の結果率は独立であった(Fisherの正確確立検定, L : p<0.001, S :
p<0.05)。更に、異型花間の交配実験と、同型花の花粉授粉後に異型花の花粉を授粉
した交配実験で、S 型で有意に結果率が低下していた(Fisher の正確確立検定, L : p=0.147, S : p<0.05)。
13
2019年の調査では、同型花間の交配実験ではL型(12花、内3花枯死)は1花の
み、11.1%結実し、S型(20花)は全く結実しなかった。一方、異型花間の交配実験
ではL型(8花)を母親とした時には87.5%で結実が得られた。S型(25花)は全て 個体が枯死していたため、データを得ることができなかった。また、同型花の花粉授 粉後に異型花の花粉を授粉した交配実験では、L型(30花)、S型(15花)ともに全 て個体が枯死していたため、データを得ることができなかった。異型花の花粉授粉後
に同型花の花粉を授粉した交配実験では、L 型(34 花)を母親とした時には 73.5%
で結実が得られ、S 型(23 花、内 7 花枯死)を母親とした時には全く結実しなかっ た。二型花間では有意な差が認められた(Fisherの正確確立検定, p<0.001)。2019年 の調査では、植物体の枯死が多数確認されたため、台風による塩害や渇水が起こった 可能性が考えられる。
④野外での二型花の結果率
調査した年度ごとの野外における結果率を Table 3 に示した。2018 年の調査にお
いて、L型が 11.26%、S 型が7.60%と、10%前後の結果率が見られた。二型花間で
は、S型の結果率が低くなる傾向が認められた(Fisherの正確確立検定, p=0.059)。
2019年の調査では、L型は全く結実せず、S型は0.87%の結実が得られた。2019年 は人工授粉実験と同様に、台風や渇水の影響が考えられる。
また、群集内における二型花の花序の位置の違いによる結実状況を把握するため、
14
2018 年に自然結果率を調査した花序の GPS 情報とその結果率を地図上にプロット した(Fig. 8)。なお、オオシラタマカズラはつる性植物であるため、各個体を識別す
ることは困難である。そのため、L型とS型の花序が見られた範囲を生育範囲として 囲んで表した。
⑤蜜量
L型は0.25~1.9µl、S型は0.025~4.15µl の蜜量を有していた。平均値はL型0.89µl、
S型 1.51µl であったが、有意差は認められなかった(Wilcoxonの順位和検定,p=0.344)
(Fig. 9, Table 4)。
⑥訪花昆虫
今回、インターバル撮影中に一度しか訪花が確認されなかった訪花者は、集計には 含めなかった。また、花弁に昆虫の体表が触れたものを訪花としてカウントした。写
真1枚ごとに1回の訪花として集計した。
デジタルカメラを用いたインターバル撮影の結果、アリ(ハチ目)複数種、アヤト ビムシ科(トビムシ目)の一種、オオモンシロナガカメムシ(直翅目)、ヒバリモド キ科(直翅目)の一種(幼虫)、カネタタキ(直翅目)の一種(幼虫)、ノメイガ?(鱗 翅目)の一種が、インターバル撮影で2回以上訪花が確認された(Fig. 10)。いずれ の訪花者についても、写真からは花粉の付着は確認することができなかった。風によ
15
ってぶれた写真や、花が咲かず花序内に1つも花が見られない写真は、訪花昆虫の集 計には含めなかった。すると、時間ごとの写真の枚数にバラつきがあったため、訪花 回数を有効な写真の枚数で割ったものを訪花頻度とした。Fig. 11 は、各昆虫の訪花
頻度を1時間ごとに表したものである。アリ類は昼~夕方頃に特に多いものの、一日 中訪花が確認された。アヤトビムシは反対に、夜~早朝にかけて特に多く、一日中訪 花が確認された。オオモンシロナガカメムシは連続的に撮影されることが多く、長時 間蜜を吸っていると推測することができる。そのため、1時間当たりの訪花頻度はこ れよりもさらに低いと考えられる。特に夜間に多かったが、時折昼間にも訪花が確認 された。ヒバリモドキとカネタタキは、日付が代わるまでの夜間に主に訪花が確認さ れた。オオモンシロナガカメムシと同様に、連続的に撮影されることも多く、長時間 蜜を舐めていることが推測される。ノメイガは、16時台でのみ訪花が確認された。
また、目視の観察によってセイヨウミツバチ(ハチ目)の訪花を確認することがで きた。セイヨウミツバチは花粉を収集し、脚に花粉団子を形成する様子を確認するこ とができた(Fig. 12)。また、暖かくなる昼頃から目視で確認されることが多かった。
16
Ⅴ.考察
1)二型花の柱頭上花粉
野外のオオシラタマカズラでは、L型の柱頭により多くの花粉が付着しているもの の、柱頭上の花粉量は L 型と S 型いずれにおいても同型花の花粉が優占しているこ とが分かった。これは、柱頭に同型花の花粉が付着することで異型花の花粉が付着し にくくなり、stigma clogging が起こりうる状況であることを示唆している。また、
Fig. 7にあるように、L型とS型のいずれの柱頭でも8割近くが同型花の花粉付着で
ある。葯と柱頭の位置関係を考慮すると、葯より柱頭が下になるS型の花での同型花 の花粉付着がより高くなるように思われるが、実際はS型の花だけでなく、二型花の いずれでも stigma cloggingが生じている可能性があると考えられる。さらに、調査 した柱頭の中でも、花粉が付着している柱頭の数に L 型と S 型で有意差が認められ たことから、S型の結果率の低さは、花粉が付着していないことによる可能性も考え られる。
2)二型花の授粉実験
同型花間授粉ではほぼ結実せず、異型花間授粉ではL型、S型ともに9割近く結実 したことから、オオシラタマカズラは先行研究(Sugawara et al. 2014)で示されてい るように、同型花不和合性を持ち、異型花間で相互に交配することが確認された。ま た、同型花間授粉後に異型花の花粉を付着させると、S 型で有意に結果率が低下し、
17
L型でも低下の傾向が確認された。従って、同型花の花粉の付着によって結果率が低
下するstigma cloggingという現象が生じる可能性が示唆される。
3)送粉昆虫
オオシラタマカズラの自然結果率は、先行研究(Sugawara et al. 2014)の結果、そ
して2018年の本調査の結果においても10%前後であった。一方、南西諸島に産する 近縁種のシラタマカズラPsychotria serpensでは、40~50%の結果率が報告されてい る(Sugawara et al. 2013, Sugawara et al. 2016)。シラタマカズラに比べて、オオシラ タマカズラの自然結果率は極めて低い状況にあり、その要因として有効な送粉昆虫が 不足している可能性が考えられるが、これについて今回のデータから考えてみたい。
オオシラタマカズラの二型花の平均蜜量はL型で 0.9µl、S型で 1.5µl であるため、飛 翔性の小型ハナバチ類の誘因には十分な蜜量である(Willmer 2011)。今回オオシラ タマカズラでは、訪花者として、飛翔性のセイヨウミツバチや小型のガ類、そしてカ メムシ類、地上徘徊性のアリやトビムシ等が確認された。一方、シラタマカズラでは、
短舌の狩りバチやハナバチ類が有効な花粉媒介者であることが報告されている
(Sugawara et al. 2016)。花形態やサイズ、蜜量等の類似性を考慮すると、オオシラ タマカズラにおいても、本来の送粉者はハナバチ類や短舌の狩りバチ類である可能性 が高いが、現在の小笠原ではそれらの訪花が少なく、有効なポリネーターは欠如して いると考えられる。
18
4)野外におけるオオシラタマカズラの繁殖の実態
上で述べたように、オオシラタマカズラの二型花は野外において、同型花の花粉付 着量が多く、また授粉実験で同型花花粉を付着すると結果率が低下する傾向にあるこ
とから、stigma cloggingが生じる状況であることが示唆される。しかし、二型花では
いずれの柱頭も同型花の花粉をより多く付着することから、S型だけがその影響を受 けて結果率がより低くなってしまうとは考えにくい。ここには現在の送粉システムが 影響しているように思われる。
オオシラタマカズラと同じく小笠原固有種である二型花柱性オガサワラボチョウ
ジPsychotria homalospermaでは、オオシラタマカズラと同様に結果率が1~10%前
後と低く、年によってバラつきはあるものの、S型の結果率が極端に低い傾向がある
(Watanabe 2017)。オガサワラボチョウジでは、本来のポリネーターであるガのよ うな長舌昆虫から、セイヨウミツバチのような短舌昆虫へポリネーターシフトが起こ
り、上方に位置するS型の葯とL型の柱頭との間の送粉が優先されている(渡邊 2007,
Watanabe 2017)。オオシラタマカズラでは、シラタマカズラ同様に、短舌昆虫が有効
なポリネーターと推測されるが、実際の訪花は極めて少ない。わずかにセイヨウミツ
バチが有効に働いているように思われる。セイヨウミツバチは1880年に初めて養蜂 のために小笠原諸島に導入された移入種で(船越 1990)、現在では小笠原諸島の多く の植物にとって主要な送粉者となっている(Kato et al. 1999, Abe 2006)。現地で実際 に数回のオオシラタマカズラへの訪花は確認できたものの、周囲にはムニンヒメツバ
19
キも開花しており、セイヨウミツバチはそちらに強く誘引されている可能性もある。
オオシラタマカズラのポリネーターとしてセイヨウミツバチや小型のガ類がある程 度働いている可能性もあるが、それらの訪花頻度は極端に低く、また本来の花粉媒介
者でないことから、より下方に位置する S 型の柱頭に異型花の花粉が運ばれにくく なり、S型の結果率低下を招いているのではないかと考えている。また、地上徘徊性 の昆虫の訪花は自花粉、すなわち同型花花粉の付着を招いている可能性もあるのでは ないかと考えている。
20
Ⅵ.結論と今後の課題
1. 全体のまとめ
小笠原諸島のオオシラタマカズラの二型花では、野外で同型花の花粉の付着が頻繁
に起こることで、stigma cloggingが生じる可能性も示唆される。しかし、現在の父島 においては、従来の昆虫相からの変化による、ポリネーターシフトや、有効なポリネ ーターの欠如、それによる訪花頻度の低さといったものが、結果率により強く影響し ていると考えられる。
2. 今後の課題 1)人工授粉実験
2019年に異型花間授粉後に同型花の花粉を付着させる、人為的stigma cloggingと は順序を反対にした交配実験を行なったが、台風や渇水の影響ではっきりとした結果
を得ることができなかった。この実験から、人為的 stigma cloggingの結果と比較す ることで、結果率の低下(異型花間授粉の阻害)がどの段階で起こっているものであ るのか、推測することができる。
2)訪花者調査
2分間隔インターバルでは、撮り逃している訪花者が存在する可能性も考えられる。
そのため、ビデオカメラやより短い間隔でのインターバル撮影によって、より詳細に
21
訪花者を観察する。セイヨウミツバチでは、片方向への送粉(S⇒L)が起こりやすい のか、1回訪花の実験等で確認する。
3)複数地点での調査
オオシラタマカズラはつる性植物であるため、今回の調査地のみでは、実際の個体 数がかなり少ない可能性も考えられる。父島内や他の島でも人工授粉実験を行ない、
結果が一致するか確認すべきである。また、今回調査した野外での繁殖の実態は、別 地点では状況が異なっている可能性も高い。特に、小笠原諸島従来の昆虫相が未だ残 っているような属島では、本来のポリネーターが送粉を行ない、高い結果率を見せる 可能性も考えられる。属島をはじめとした複数の地点で、オオシラタマカズラの野外 における繁殖の実態、特にポリネーターに関して調査するべきであると考える。
22
Ⅶ.謝辞
本研究を進めるにあたり、きっかけを与えてくださり、調査や研究等で全面的にサ ポート、丁寧なご指導を頂きました、指導教員の菅原敬准教授に心から感謝致します。
また、多くの助言をしていただいた村上哲明教授、江口克之准教授、加藤英寿助教を はじめとした教員の方々に心から感謝致します。苅部治紀氏、岸本年郎氏、森英章氏 には、昆虫の同定や、小笠原諸島の昆虫について貴重な情報を頂きました。深く御礼 申し上げます。
牧野標本館の教員・学生の皆様をはじめ、首都大学東京生命科学専攻の教員・学生 の方々には多くの助言や知識・サポートを頂きました。他にも、小笠原諸島の調査・
研究をされている方々からも貴重な助言を頂きました。多くの方々の御協力の下、本 研究を進めてくることができました。ここに深く感謝致します。
23
Ⅷ.参考文献
Abe T., 2006. Threatened pollination systems in native flora of the Ogasawara (Bonin) Islands. Annals of Botany. 98(2): 317-334.
Barrett: S. C. H. 1992. Heterostylous genetic polymorphisms: Model systems for evolutionary analysis. In: Barrett S. C. H. (ed.), Evolution and Function of Heterostyly. Springer-Verlag, Berlin. 1-29
Beach, J. H. and Bawa, K. S. 1980. Role of pollinators in the evolution of dioecy from distyly. Evolution 34: 1138-1142.
Ehlers, B. K. 1999. Variation in fruit set within and among natural populations of the self-incompatible herb Centaurea scabiosa (Asteraceae). Nordic Journal of Botany. 19: 653-663
Ganders, F. R. 1979. The biology of heterostyly. New Zealand Journal of Botany 17:
607-635.
Holland J. N. and Chamberlain S. A. 2007. Ecological and evolutionary mechanisms for low seed : ovule rations: need for a pluralistic approach? Ecology 88(3): 706- 715.
船越真樹. 1990. 小笠原諸島におけるギンネム林の成立―移入と分布の拡大をめぐる 覚え書―その4. 小笠原研究年報14: 21-51.
24
Kato M., Shibata A., Yasui T. and Nagamasu H. 1999. Impact of introduced honeybees, Apis mellifera, upon native bee communities in the Bonin (Ogasawara) Islands.
Research on Population Ecology. 41: 217-228.
菊沢喜八郎. 1995. 植物の繁殖生態学 蒼樹書房 279.
Kondo Y., Nishide M., Watanabe K. and Sugawara T. 2007. Floral dimorphism in Psychotria boninensis Nakai (Rubiaceae) endemic to the Bonin (Ogasawara) Islands. The Journal of Japanese Botany 82: 251-258.
西廣淳. 2000. 花のかたち:二型花柱性における花の形態変異と繁殖成功. 『花生態 学の最前線』 種生物学会編 文一総合出版 125-144.
Sugawara T., Watanabe T. and Tabata M. 2013. Distyly in Psychotria serpens (Rubiaceae) in the Ryukyu Islands, Japan. Acta Phytotaxonomica et.
Geobotanica. 64: 127-136.
Sugawara T., Yumoto M., Tsuneki S. and Watanabe K. 2014. Incompatibility and reproductive output in distylous Psychotria boninensis (Rubiaceae), endemic to the Bonin (Ogasawara) Islands, Japan. The Journal of Japanese Botany 89: 22- 26.
Sugawara T., Watanabe K., Takaso T., Tabata M. and Shimizu A. 2016. Incompatibility and pollination of distylous Psychotria serpens (Rubiaceae) in the Ryukyu Islands, Japan. Acta Phytotaxonomica et Geobotanica. 67(1): 37-45.
25
豊田武司. 2014. 小笠原諸島固有植物ガイド. ウッズプレス 260-262.
鷲谷いづみ. 2006. サクラソウー植物の保全と進化を科学する. サクラソウの分子遺 伝生態学. 東京大学出版会 1-14.
渡邊謙太. 2007. 「小笠原諸島固有種オガサワラボチョウジ(アカネ科)の二型花柱 性と繁殖様式:ポリネーターシフトによる片方向への送粉」 修士論文
Watanabe K., Kato H., Kuraya E. and Sugawara T. 2017. Pollination and reproduction of Psychotria homalosperma, an endangered distylous tree endemic to the oceanic Bonin (Ogasawara) Islands, Japan. Plant Species Biology. 10.1111:
1442-1984.
Willmer P. 2011. Pollination and Floral Ecology. Princeton University Press, New Jersey. 202-205.
26
Ⅸ. 図表
Table 1. オオシラタマカズラの二型花の花粉サイズ
型 花数 花粉粒数 花粉サイズ(µm)
平均±SD p値
L型 5 135 34.56±1.97
p<0.001
S型 4 118 61.67±4.11
Wilcoxonの順位和検定
27
Table 2. オオシラタマカズラ二型花間の人工授粉実験による結果率
L型 S型
2018
花序数 (枯死数)
実験花数 (枯死数)
結果数
結果率 (%)
花序数 (枯死数)
実験花数 (枯死数)
結果数
結果率 (%)
p値
自家授粉 3 7 0 0.00 1(1) 2(1) 0 0.00 1 異型花授粉 4 18 16 88.89 2 6 6 100.00 1 同型花→
異型花授粉
3(1) 10(3) 6 60.00 3 10 4 40.00 0.656
異型花→
同型花授粉
No date No date
p<0.001 p<0.05
2019
自家授粉 1(1) 9(3) 1 11.11 2 20 0 0.00 0.310 異型花授粉 1 8 7 87.50 0(1) 0(25) No date
同型花→
異型花授粉
0(2) 0(30) No date 0(2) 0(15) No date
異型花→
同型花授粉
2 34 25 73.53 1(1) 16(7) 0 0.00 p<0.001
p<0.001 total
自家授粉 4(1) 16(3) 1 6.25 3(1) 22(1) 0 0.00 0.421 異型花授粉 5 26 23 88.46 2(1) 6(25) 6 100.00 1 同型花→
異型花授粉
3(3) 10(33) 6 60.00 3(2) 10(15) 4 40.00 0.656
異型花→
同型花授粉
2 34 25 73.53 1(1) 16(7) 0 0.00 p<0.001
p<0.001 p<0.001
Fisherの正確確立検定
28
Table 3. オオシラタマカズラの野外における二型花の自然結果率
L型 S型
調査年
花序数 (枯死数)
花数 (枯死数)
結果した 花数
結果率 (%)
花序数 (枯死数)
花数 (枯死数)
結果した 花数
結果率 (%)
p値
2018 19 870 98 11.26 9 342 26 7.60 0.059
2019 5(7) 368(192) 0 0.00 8(2) 343(73) 3 0.87 0.112
total 24(7) 1238(192) 98 7.92 17(2) 685(73) 29 4.23 p<0.01
Fisherの正確確立検定
29
Table 4. オオシラタマカズラ二型花の蜜量
型 花数 蜜量(µl)
平均±SD p値
L型 17 0.89±0.65
0.344
S型 20 1.51±1.29
Wilcoxonの順位和検定
30
Fig. 1. オオシラタマカズラ(Psychotria boninensis)の花と果実、生育状況。
A:長花柱花の花序. B:果実. C,D:生育地の状況.
E:カメラによる送粉昆虫の調査風景
A B
C D
E
31
Fig. 2. オオシラタマカズラの花と花粉.
A, B:オオシラタマカズラの長花柱花(A)と短花柱花(B).
C:コットンブルーで染色した花粉、及び花粉サイズの測定部位.
A B
花粉サイズ(直径)
C
32
Fig. 3. 小笠原諸島の父島と調査地点
1000m 5000ft
ⒸOpenStreetMap contributors
調査地点
33
Fig. 4. オオシラタマカズラの二型花の花粉サイズ
A. 花粉サイズの頻度分布図 B. 二型の花粉の箱ひげ図
L型花粉 S型花粉 (n=135) (n=118)
0 10 20 30 40 50 60 70
≦27.5 30 32.5 35 37.5 40 42.5 45 47.5 50 52.5 55 57.5 60 62.5 65 67.5 70 72.5 75
花粉数
花粉サイズ(µm)
L 型花粉 S型花粉
B.
A.
34
Fig. 5. オオシラタマカズラの二型花の柱頭上に付着した花粉数
0 50 100 150 200 250 300
柱頭上花粉数
花粉サイズ(µm)
L花(n=24)
S花(n=24)
35
L 型
S 型
Fig. 6. L型(上)とS型(下)の柱頭に付着した二型花の花粉数
L型は≦40µm、S型は>40µm
0 20 40 60 80 100 120 140 160 180 200
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
付着花粉数
柱頭No.
0 50 100 150 200
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
付着花粉数
柱頭No.
S型花粉 L型花粉
L型花粉 S型花粉
36
L 型
S 型
Fig. 7. 二型花の柱頭上に付着するL型/S型花粉の比率
(上)L型柱頭.(下)S型柱頭.
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
1 (193)
2 (113)
3 (110)
4 (96)
5 (36)
6 (26)
7 (23)
8 (20)
9 (17)
10 (13)
11 (7)
12 (6)
13 (3)
14 (3)
15 (2)
16 (2)
17 (2)
18 (1)
19 (1)
20 (1)
21 (0)
22 (0)
23 (0)
24 (0)
花粉比率
柱頭No.
(付着花粉数)
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
1 (92)
2 (38)
3 (20)
4 (18)
5 (12)
6 (11)
7 (6)
8 (4)
9 (3)
10 (0)
11 (0)
12 (0)
13 (0)
14 (0)
15 (0)
16 (0)
17 (0)
18 (0)
19 (0)
20 (0)
21 (0)
22 (0)
23 (0)
24 (0)
花粉比率
柱頭No.
(付着花粉数)
S型花粉 L型花粉
L型花粉 S型花粉
37
Fig. 8. オオシラタマカズラ巽集団における二型花の花序ごとの結果率
L型とS型が混在して生育 S型が単体で生育
L型が単体で生育 道路
駐車場
38
Fig. 9. 二型花の蜜量の箱ひげ図
L型 S型 (n=17) (n=20)
39
Fig. 10. オオシラタマカズラに訪花した昆虫
(A)アリ(ハチ目). (B)アヤトビムシ科(トビムシ目)の一種.
(C)オオモンシロナガカメムシ(直翅目). (D)ヒバリモドキ科(直翅目)の一種(幼虫).
(E)カネタタキ(直翅目)の一種(幼虫). (F)ノメイガ?(鱗翅目)の一種
A B
C D
E F
40
Fig. 11. オオシラタマカズラに訪花した昆虫の時間ごとの訪花頻度
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
0:00 1:00 2:00 3:00 4:00 5:00 6:00 7:00 8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00 19:00 20:00 21:00 22:00 23:00
訪花頻度 (1時間当たりの訪花数/有効写真枚数)
各昆虫の訪花時間
アリ(ハチ目) ノメイガ?(直翅目)
カネタタキ(直翅目) ヒバリモドキ科(直翅目)
オオモンシロナガカメムシ(直翅目) アヤトビムシ科(トビムシ目)
41
Fig. 12. 目視にて確認されたセイヨウミツバチの訪花