Title Sphingosine 1-phosphate (S1P) suppresses the collagen-induced activation of human platelets via S1P4 receptor( 本文(Fulltext) )
Author(s) 大沼, 隆史
Report No.(Doctoral
Degree) 博士(医学) 甲第1060号
Issue Date 2018-03-25
Type 博士論文
Version ETD
URL http://hdl.handle.net/20.500.12099/75197
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
1 S
Sphingosine 1-phosphate (S1P) suppresses the collagen-induced activation of human platelets 1
via S1P4 receptor 2
3
Takashi Onumaa,b, Kumiko Tanabea, Yuko Kitoa,b, Masanori Tsujimotob,c, Kodai Uematsub,c, 4
Yukiko Enomotoc, Rie Matsushima-Nishiwakib, Tomoaki Doid, Kiyoshi Nagasea, Shigeru 5
Akamatsue, Haruhiko Tokudaf, Shinji Ogurad, Toru Iwamac, Osamu Kozawab and Hiroki Iidaa 6
7
aDepartment of Anesthesiology and Pain Medicine, Gifu University Graduate School of Medicine, 8
Gifu, Japan 9
bDepartment of Pharmacology, Gifu University Graduate School of Medicine, Gifu, Japan 10
cDepartment of Neurosurgery, Gifu University Graduate School of Medicine, Gifu, Japan 11
dDepartment of Emergency and Disaster Medicine, Gifu University Graduate School of Medicine, 12
Gifu, Japan 13
eDepartment of Anesthesiology and Critical Care Medicine, Chuno Kosei Hospital, Seki, Japan 14
fDepartment of Clinical Laboratory and Biobank, National Center for Geriatrics and 15
Gerontology, Obu, Japan 16
17 18
Corresponding author: 1-1 Yanagido, Gifu 501-1194, Japan. Tel: +81 58 230 6214; fax: +81 58 19
230 6215 20
E-mail: [email protected] (O. Kozawa) 21
22
Key words: Sphingosine 1-phosphate, collagen, platelet, HSP27, phosphorylation, receptor 23
24 25 26
2 A
Abstract 1
Sphingosine 1-phosphate (S1P) is as an extracellular factor that acts as a potent lipid 2
mediator by binding to specific receptors, S1P receptors (S1PRs). However, the precise role of 3
S1P in human platelets that express S1PRs has not yet been fully clarified. We previously 4
reported that heat shock protein 27 (HSP27) is released from human platelets accompanied by 5
its phosphorylation stimulated by collagen. In the present study, we investigated the effect of 6
S1P on the collagen-induced platelet activation. S1P pretreatment markedly attenuated the 7
collagen-induced aggregation. Co-stimulation with S1P and collagen suppressed 8
collagen-induced platelet activation, but the effect was weaker than that of S1P-pretreatment.
9
The collagen-stimulated secretion of platelet-derived growth factor (PDGF)-AB and the soluble 10
CD40 ligand (sCD40L) release were significantly reduced by S1P. In addition, S1P suppressed 11
the collagen-induced release of HSP27 as well as the phosphorylation of HSP27. S1P 12
significantly suppressed the collagen-induced phosphorylation of p38 mitogen-activated protein 13
kinase. S1P increased the levels of GTP-bound Gαi and GTP-bound Gα13 coupled to S1PPR1 14
and/or S1PR4. CYM50260, a selective S1PR4 agonist, but not SEW2871, a selective S1PR1 15
agonist, suppressed the collagen-stimulated platelet aggregation, PDGF-AB secretion and 16
sCD40L release. In addition, CYM50260 reduced the release of phosphorylated-HSP27 by 17
collagen as well as the phosphorylation of HSP27. The selective S1PR4 antagonist CYM50358, 18
which failed to affect collagen-induced HSP27 phosphorylation, reversed the S1P-induced 19
attenuation of HSP27 phosphorylation by collagen. These results strongly suggest that S1P 20
inhibits the collagen-induced human platelet activation through S1PR4 but not S1PR1.
21 22 23 24 25 26
3 IIntroduction
1
Sphingosine 1-phosphate (S1P), which is formed from the breakdown of sphingomyelin, is 2
a potent bioactive lipid [1]. Accumulating evidence indicates that S1P acts not only 3
intracellularly but also extracellularly through specific receptors, called S1P receptors, that are 4
classified as G-protein coupled receptors (GPCRs) [1,2]. S1P has been implicated in a variety of 5
fundamental biological cellular behaviors, including migration, adhesion, survival and 6
proliferation, and physiological processes such as embryogenesis, heart development and 7
immunity [1-3]. S1P also plays essential roles in the hematopoietic systems [4]. S1P in 8
plasma sourced from red blood cells and endothelial cells participates in lymphocyte egress from 9
lymphoid organs [5,6]. Human platelets as well as megakaryocytes, which are precursors of 10
platelets, express several types of S1P receptors (i.e. S1PR1, 2, 4) [4,7]. S1PR1 in particular is 11
essential for human platelet formation from megakaryocytes and their release into circulation 12
from the bone marrow space [2,4,7]. However, although S1PR4 is proposed to be involved in 13
the acceleration of platelet generation, the exact role of S1PR4 in platelet formation remains to 14
be elucidated [4]. Human platelets abundantly contain S1P, which is released into the 15
extracellular space along with platelet activation [4]. Regarding the effects of S1P on human 16
platelets, it has been reported that S1P itself induces platelet shape change without affecting 17
the secretion of serotonin [8] or platelet aggregation [9]. In addition, S1P reportedly 18
downregulates platelet aggregation by thrombin receptor-activating peptide or noradrenaline 19
[10]. However, it has been shown using whole blood that endogenous S1P amplifies the platelet 20
aggregation induced by ADP or protease-activated receptor 4-peptide through S1PR1 [11]. The 21
effects of S1P on human platelet aggregation therefore remain controversial, and the exact roles 22
of S1P and S1PRs in human platelet functions have not yet been fully clarified.
23
Human platelets play a pivotal role in hemostasis. Adhesion of human platelets to the 24
injured vessel wall is mediated by adhesive receptors, such as glycoprotein (GP) Ib/IX/V, which 25
interacts with von Willebrand factor, mediating the rolling and tethering of platelets [12].
26
4
Collagen is generally recognized as one of the most important stimuli for human platelet 1
activation via GPVI and integrin α2β1 on the plasma membrane of platelets. Platelet 2
activation through GPVI leads to the stabilization of platelet adhesion, resulting in thrombus 3
formation [13]. Accompanied by platelet aggregation, the activated platelets secrete granule 4
contents including platelet-derived growth factor-AB (PDGF-AB) and release inflammatory 5
mediators such as soluble CD40 ligand (sCD40L). These secreted and generated mediators 6
trigger a positive feedback mechanism, which potentiates further platelet activation [12-14].
7
PDGF-AB is a potent mitogenic growth factor, which acts mainly in connective tissue, such as 8
vascular smooth muscle cells, and promotes atherosclerosis [15]. However, sCD40L, which is 9
released by activated platelets, induces inflammatory responses in endothelium [16]; this event 10
is an important component in the pathogenesis of atherosclerosis [12].
11
Heat shock proteins (HSPs) are induced by various stresses, including environmental, 12
metabolic and pathophysiological stress [17]. HSPs act intracellularly as molecular chaperones 13
and facilitate the refolding of nonnative or stress-accumulated misfolded proteins, preventing 14
their aggregation [17]. HSPs are currently classified into seven major groups, including HSPB 15
(small HSP) [17]. One HSPB, HSP27 (HSPB1), is ubiquitously expressed in its aggregated 16
form in a variety type of cells, including human platelets. HSP27 is well established to be 17
phosphorylated at three serine residues: Ser-15, Ser-78 and Ser-82 [18]. With this 18
phosphorylation, HSP27 transforms from an aggregated complex into a dissociated form, with 19
subsequent modulation of its chaperone activity [19,20]. In addition to its chaperone activity, 20
accumulating evidence suggests that HSP27 has various biological activities, with involvement 21
in membrane stability, actin polymerization, proinflammatory gene expression and apoptosis, 22
among other functions [20]. In addition, it has recently been shown that HSP27 extracellularly 23
functions as a regulator of both anti-inflammatory factors and pro-inflammatory factors in 24
macrophages [21].
25
Regarding HSP27 in human platelets, we previously demonstrated that ADP or 26
5
collagen-induced HSP27 phosphorylation via p44/p42 mitogen-activated protein (MAP) kinase is 1
accompanied with the granule secretion of PDGF-AB and the release of sCD40L [22-24].
2
Additionally, we have shown that antithrombin III or (-)-epigallocatechin gallate suppress the 3
ADP-induced phosphorylation of HSP27 via p44/p42 MAP kinase, leading to a decrease in 4
platelet aggregation along with PDGF-AB secretion and sCD40L release [25,26]. In our recent 5
study [27], we reported that HSP27 is released from collagen-stimulated human platelets into 6
plasma following its phosphorylation in diabetic patients.
7
In the present study, we investigated the effect of S1P on collagen-induced human platelet 8
activation. We found that S1P inhibits collagen-induced aggregation and the phosphorylation 9
of HSP27 via S1PR4 but not S1PR1 in human platelets, resulting in the suppression of 10
PDGF-AB secretion and of the release of sCD40L and phosphorylated-HSP27.
11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
6 M
Materials and methods 1
Materials
2S1P was purchased from Sigma-Aldrich (St. Louis, MO, USA). SEW2871, CYM50260 and 3
CYM50358 were obtained from Tocris Bioscience (Bristol, UK). Collagen was purchased from 4
Takeda Austria GmbH (Linz, Austria). Anti-phospho HSP27 (Ser-78) antibodies were obtained 5
from Stressgen Biotechnologies (Vitoria, BC, Canada). Anti-phospho p38 MAP kinase 6
antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). Anti-GAPDH 7
antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
8
PDGF-AB enzyme-linked immunosorbent assay (ELISA) kit and sCD40L ELISA kit were 9
obtained from R&D Systems, Inc. (Minneapolis, MN). A phosphorylated-HSP27 (Ser-78) 10
enzyme-linked immunosorbent assay (ELISA) kit was purchased from Enzo Life Sciences, Inc.
11
(Farmingdale, NY, USA). Gαi activation assay kit and Gα13 activation assay kit were 12
purchased from NewEast Biosciences (King of Prussia, PA, USA). Other materials and 13
chemicals were obtained from commercial sources. S1P, SEW2871, CYM50260 and CYM50358 14
were dissolved in dimethyl sulfoxide. The maximum concentration of dimethyl sulfoxide was 15
0.1%, which did not affect platelet aggregation, the protein detection using Western blotting or 16
ELISA for PDGF-AB, sCD40L and phosphorylated-HSP27.
17 18
Preparation of platelets
19Human blood was drawn from healthy volunteers, and sodium citrate (14 μM) was added 20
immediately as an anti-coagulant. Platelet-rich plasma (PRP) was obtained by centrifuging at 21
155 ×
g
for 12 min at room temperature. Platelet-poor plasma (PPP) was obtained from the 22residual blood by centrifuging at 1,400 ×
g
for 5 min. This study was approved by the Ethics 23Committee of Gifu University Graduate School of Medicine, Gifu, Japan. Written informed 24
consent was obtained from all participants.
25 26
7
Platelet aggregation
1
Platelet aggregation was measured using an aggregometer (PA-200 Kowa Co. Ltd., Tokyo, 2
Japan) with laser-scattering, which measures the size of platelet aggregates by particle counting 3
(small: 9-25 μm, medium: 25-50 μm and large: 50-70 μm). Briefly, PRP was pre-incubated for 1 4
min at 37°C with a stirring speed at 800 rpm. When indicated, PRP was pretreated with 5
various doses of S1P, SEW2871 or CYM50260 for 15 min. After the pretreatment, PRP was 6
stimulated by collagen, and platelet aggregation was monitored for 4 min. The dose of collagen 7
achieving a % transmittance of 80%-100% was adjusted individually. The percentage of 8
transmittance of isolated PRP was recorded as 0%, and that of the appropriate PPP (blank) was 9
recorded as 100%. When indicated, PRP was co-stimulated by 0.5 μg/ml of collagen with 30 μM 10
of S1P or vehicle.
11 12
Protein preparation after stimulation
13After stimulation by collagen, platelet aggregation was terminated by adding an ice-cold 14
EDTA (10 mM). The mixture was collected and centrifuged at 10,000 ×
g
at 4°C for 2 min.15
The supernatant was collected for ELISA and stored at –80°C. The pellet was washed twice 16
with phosphate-buffered saline and then lysed by boiling in a lysis buffer containing 62.5 mM 17
Tris-HCl, pH 6.8, 2% sodium dodecyl sulfate (SDS), 50 mM dithiothreitol and 10% glycerol for a 18
Western blot analysis.
19 20
Western blotting
21A Western blot analysis was performed as described previously [27]. Briefly, 22
SDS-polyacrylamide gel electrophoresis (PAGE) was performed via the method described by 23
Laemmli in a 12.5% polyacrylamide gel [28]. The proteins in the gel were transferred onto a 24
polyvinylidene difluoride (PVDF) membrane, which was then blocked with 5% fat-free dry milk 25
in Tris-buffered saline with 0.1% Tween 20 (TBS-T; 20 mM Tris, pH 7.6, 137 mM NaCl, 0.1%
26
8
Tween 20) for 2 h before incubation with the indicated primary antibodies. Peroxidase-labeled 1
anti-rabbit IgG antibodies were used as secondary antibodies. The primary and secondary 2
antibodies were diluted to optimal concentrations with 5% fat-free dry milk in TBS-T. The 3
peroxidase activity on the PVDF membranes was visualized on X-ray film using an ECL 4
Western blotting detection system (GE Healthcare, Buckinghamshire, UK) as described in the 5
manufacturer’s protocol. A densitometric analysis was performed using a scanner and an 6
imaging software program (Image J version 1.47; National Institutes of Health, Bethesda, MD, 7
USA). The phosphorylated levels were calculated as follows: the background-subtracted 8
intensity of each signal was normalized to the respective intensity of GAPDH and plotted as the 9
fold increase compared with control cells.
10 11
ELISA for PDGF-AB, sCD40L and phosphorylated-HSP27
12The levels of PDGF-AB, sCD40L and phosphorylated-HSP27 (Ser-78) in the supernatant of 13
the conditioned mixture after platelet aggregation were determined using ELISA kits for 14
PDGF-AB, sCD40L and phosphorylated-HSP27 (Ser-78) respectively, in accordance with the 15
manufacturer’s instructions.
16 17
Measurement of Gαi or Gα13 activity
18PRP was stimulated by 30 μM of S1P at 37°C for 30 sec, and the reaction was terminated 19
by the addition of ice-cold EDTA solution. The mixture was then centrifuged at 10,000 ×
g
at 204°C for 2 min, and the pellet was washed twice with ice-cold Tris-buffered saline. Using 21
activation assay kits for Gαi and Gα13 (NewEast Biosciences), the levels of GTP-bound Gαi or 22
GTP-bound Gα13 were determined in accordance with the manufacturer’s instructions.
23 24
Statistical analyses
25The data were analyzed by the Mann-Whitney U test. A probability of less than 5% was 26
9
considered to be statistically significant. The data are presented as the mean ± standard error 1
of the mean (SEM).
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
10 R
Results 1
Effect of S1P on collagen-induced human platelet aggregation
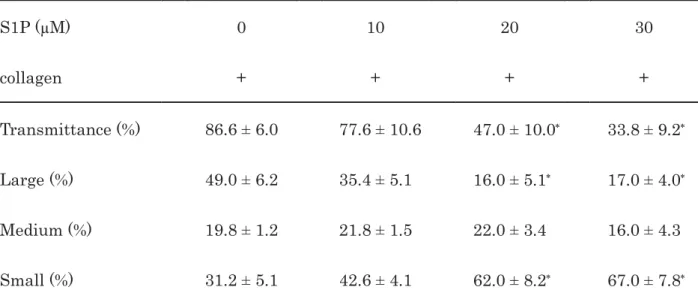
2First, we examined the effect of S1P on the human platelet aggregation induced by 3
collagen. The representative pattern of S1P-effect on the collagen-stimulated platelet 4
aggregation analyzed by an aggregometer with the laser scattering system is shown in Fig. 1.
5
S1P, which by itself had no effect on human platelet aggregation (data not shown), markedly 6
suppressed the collagen-induced platelet aggregation in a dose-dependent manner at doses of 7
10-30 μM. Regarding the size ratios of the platelet aggregates, S1P dose-dependently 8
decreased the prevalence of large aggregates (50-70 μm) but markedly increased that of small 9
aggregates (9-25 μm) (Table 1).
10
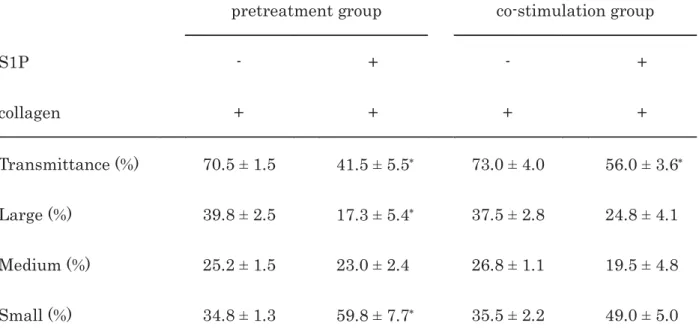
We examined the effect of co-stimulation with S1P and collagen on the platelet aggregation 11
and compared the outcome with that of S1P-pretreatment. Both co-stimulation with S1P and 12
collagen as well as pretreatment significantly suppressed the collagen-stimulated platelet 13
aggregation in terms of transmittance (Fig. 2). According to an analysis of the size of the 14
platelet aggregates, co-stimulation decreased the ratio of large aggregates but increased that of 15
small aggregates. In contrast, S1P-pretreatment markedly decreased the ratio of large 16
aggregates (50-70 μm) and increased that of small aggregates (9-25 μm) (Table 2).
17 18
Effects of S1P on collagen-induced secretion of PDGF-AB and release of sCD40L or
19phosphorylated-HSP27 from human platelets
20Collagen is recognized to stimulate the secretion of PDGF-AB as well as the release of 21
sCD40L [29,30]. In our recent study [27], we showed that collagen induces the phosphorylation 22
of HSP27 in human platelets, and subsequently, the release of phosphorylated-HSP27 into 23
plasma. We next examined the effects of S1P on the collagen-stimulated secretion of PDGF-AB 24
and release of sCD40L and phosphorylated-HSP27. S1P significantly reduced the 25
collagen-stimulated secretion of PDGF-AB in a dose-dependent manner at doses of 10-30 μM, 26
11
and 30 μM of S1P caused an approximately 80% reduction in the collagen effect (Fig. 3A). The 1
release of sCD40L induced by collagen was dose-dependently suppressed by S1P at doses of 2
10-30 μM (Fig. 3B). The maximum effect of S1P was observed at 30 μM, causing an 3
approximately 90% reduction in the collagen effect. In addition, S1P (30 μM) markedly 4
attenuated the collagen-stimulated release of phosphorylated-HSP27 (Ser-78) (Fig. 3C).
5 6
Effect of S1P on the collagen-induced phosphorylation of HSP27 and p38 MAP kinase in human
7platelets
8We previously reported that the collagen-induced phosphorylation of HSP27 in human 9
platelets of healthy volunteers was accompanied by the collagen-induced PDGF-AB secretion or 10
sCD40L release [22], and that the collagen-induced phosphorylation of HSP27 led to the 11
subsequent release of phosphorylated-HSP27 (Ser-78) from platelets of diabetic patients into 12
plasma [27].
13
We therefore examined the effect of S1P on the phosphorylation of HSP27 (Ser-78) 14
stimulated by collagen in human platelets. The collagen-induced phosphorylation levels of 15
HSP27 (Ser-78) in human platelets were significantly suppressed by 30 μM of S1P, which alone 16
had little effect on the phosphorylation of HSP27 (Ser-78) (Fig. 4A). Regarding the mechanism 17
underlying the collagen-induced phosphorylation of HSP27 in human platelets, it has been 18
reported that p38 MAP kinase is involved [31]. In our previous study[32], we showed that the 19
collagen-activated p38 MAP kinase levels reflect platelet hyperaggregability in type II diabetes 20
mellitus patients.
21
We therefore investigated the effect of S1P on the collagen-induced phosphorylation of p38 22
MAP kinase in the same samples showing an S1P effect on the HSP27 phosphorylation induced 23
by collagen. S1P, which alone had little effect on the phosphorylation of p38 MAP kinase, 24
significantly suppressed the collagen-induced phosphorylation of p38 MAP kinase as well as 25
HSP27 phosphorylation (Fig. 4B).
26
12 1
Effect of S1P on Gi or G13 activity in human platelets
2S1PR1 is known to be coupled to Gi, and S1PR4 is known to be coupled to Gi and G12/13 3
[33]. Activated G protein-coupled receptor (GPCR) has been reported to act as a guanine 4
nucleotide exchange factor for the α-subunit of heterotrimeric G protein, resulting in the release 5
of GDP and the binding of GTP for the activation of G protein [34]. We therefore examined 6
whether or not S1P increases the levels of GTP-bound Gαi or GTP-bound Gα13 in human 7
platelets. S1P markedly increased the levels of GTP-bound Gαi in human platelets (Fig. 5A).
8
Similarly, the levels of GTP-bound Gα13 were also up-regulated by S1P (Fig. 5B).
9 10
Effects of SEW2871 or CYM50260 on the collagen-induced platelet aggregation, the PDGF-AB
11secretion, the sCD40L release, the phosphorylated-HSP27 release or the phosphorylation of
12HSP27
13Regarding S1P receptors in human platelets, human platelets are known to express three 14
receptor subtypes: S1PR1, S1PR2 and S1PR4 [4,7]. Among these, S1PR1 and S1PR4 are 15
considered to be functionally important in human platelets [4,7]. In order to clarify which 16
subtypes of S1P receptors is involved in the inhibition by S1P of the collagen-induced platelet 17
aggregation, PDGF-AB secretion, sCD40L release, phosphorylation of HSP27 (Ser-78) and the 18
release, we examined the effects of selective agonists for S1PR1 or S1PR4 on these events.
19
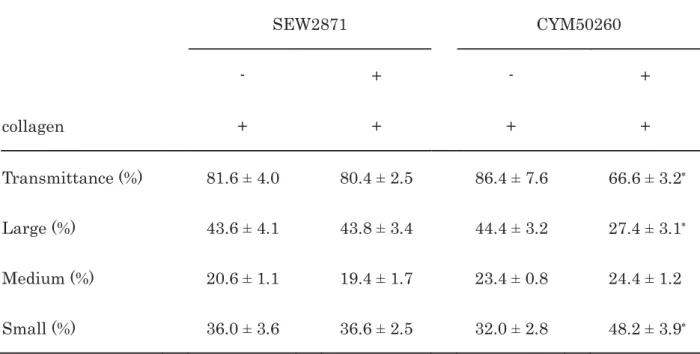
SEW2871 at doses <10 μM and CYM50260 at doses <25 μM are reported to selectively 20
activate S1PR1 and S1PR4, respectively [35,36]. SEW2871 (10 μM) failed to suppress the 21
collagen-induced platelet aggregation (Fig. 6A). Regarding the size of platelet aggregates, 10 22
μM of SEW2871 hardly affected the ratios of the aggregated particle size (Table 3). However, 23
25 μM of CYM50260 markedly inhibited the collagen-induced platelet aggregation (Fig. 6B).
24
Regarding the ratios of aggregated particle size, CYM50260 (25 μM) decreased the ratio of large 25
aggregates (50-70 μm) but markedly increased that of the small aggregates (9-25 μm) (Table 3).
26
13
We further examined the effects of SEW2871 and CYM50260 on the PDGF-AB secretion, 1
the sCD40L release and the phosphorylated-HSP27 (Ser-78) release from human platelets 2
stimulated by collagen. SEW2871 (10 μM) had little effect on the collagen-induced PDGF-AB 3
secretion whereas CYM50260 (25 μM) significantly inhibited the PDGF-AB secretion (Fig. 7A).
4
Regarding the collagen-induced sCD40L release, CYM50260 (25 μM) but not SEW2871 (10 μM) 5
significantly inhibited the release (Fig. 7B). In addition, CYM50260 (10 μM) significantly 6
reduced the phosphorylated-HSP27 (Ser-78) release from collagen-stimulated human platelets 7
(Fig. 7C).
8
We also examined the effects of SEW2871 or CYM50260 on the collagen-induced 9
phosphorylation of HSP27 (Ser-78) in human platelets. SEW2871 (10 μM) hardly affected the 10
phosphorylation levels of HSP27 (Ser-78) stimulated by collagen (Fig. 8A). In contrast, 11
CYM50260 (25 μM) significantly suppressed the collagen-induced phosphorylation levels of 12
HSP27 (Ser-78) (Fig. 8B). It therefore seems likely that S1PR4—but not S1PR1—might 13
mediate the inhibitory effects of S1P on the collagen-stimulated human platelet activation.
14 15
Effect of CYM50358 on the suppression by S1P of collagen-induced HSP27 phosphorylation
16To clarify whether or not the suppressive effects of S1P on collagen-stimulated human 17
platelets are mediated through S1PR4, we examined the effect of CYM50358, a selective S1PR4 18
antagonist [37], on the suppression by S1P of HSP27 (Ser-78) phosphorylation. CYM50358 (10 19
μM), which had no effect on the collagen-induced HSP27 phosphorylation (Fig. 9A), markedly 20
reversed the suppressive effect of S1P on the collagen-induced phosphorylation of HSP27 21
(Ser-78), almost to the levels of collagen alone (Fig. 9B).
22 23 24 25 26
14 D
Discussion 1
In the present study, we investigated the effect of S1P on the collagen-induced activation of 2
human platelets. We first demonstrated that S1P markedly attenuated the human platelet 3
aggregation stimulated by collagen. Regarding the size of the platelet aggregates, S1P 4
decreased the ratio of large aggregates (50-70 μm) but increased the ratio of small aggregates 5
(9-25 μm), suggesting that S1P down-regulates the formation of aggregate particles in human 6
platelets. We also examined the effect of co-stimulation with S1P and found that 7
S1P-pretreatment more potently down-regulates the collagen-induced platelet aggregation than 8
co-stimulation. These findings suggest that S1P prior to collagen addition rather than 9
co-stimulation exerts significant effects on the collagen-stimulated platelet activation.
10
Therefore, in the present study, we performed subsequent experiments with S1P pretreatment 11
prior to collagen stimulation but not co-stimulation.
12
Activated human platelets are well known to induce PDGF-AB secretion and sCD40L 13
release [29,30]. We showed that S1P inhibited the collagen-induced the secretion of PDGF-AB 14
and the release of sCD40L. Therefore, S1P likely suppresses the human platelet activation 15
induced by collagen, leading to a reduction in the platelet aggregation, granule secretion and 16
sCD40L release. In our previous study [22], we reported that HSP27 phosphorylation in 17
human platelets was accompanied by collagen-induced PDGF-AB secretion and sCD40L release.
18
We therefore next examined the effect of S1P on the collagen-induced phosphorylation of HSP27.
19
S1P significantly reduced the phosphorylation levels of HSP27 (Ser-78) induced by collagen. To 20
our knowledge, this is the first report clearly showing the attenuation by S1P of 21
collagen-induced phosphorylation of HSP27 in human platelets.
22
We recently showed in patients with type II diabetes mellitus that HSP27 is released from 23
human platelets into plasma, accompanied by its phosphorylation induced by collagen [27].
24
Therefore, we additionally examined the effect of S1P on the release of phosphorylated-HSP27 25
from the human platelets stimulated by collagen and showed that S1P significantly attenuated 26
15
the release of phosphorylated-HSP27 (Ser-78) from the platelets. Regarding the mechanism of 1
the phosphorylation of HSP27 in human platelets, we found that S1P suppressed the 2
collagen-induced phosphorylation of p38 MAP kinase as well as HSP27 phosphorylation.
3
Given these findings, S1P likely negatively regulates the collagen-induced p38 MAP kinase 4
activation in human platelets, at least in part, leading to the suppression of the HSP27 5
phosphorylation and the secretion of PDGF-AB and sCD40L.
6
S1PR1, S1PR2 and S1PR4 are recognized as receptors for S1P expressed on human 7
platelets [4,11]. Among these receptors, S1PR1 and S1PR4 have been reported to function in 8
the process of platelet formation from megakaryocytes [4]. S1PR1 is coupled to Gi, and S1PR4 9
is coupled to Gi and G12/13[33]. In the present study, we found that S1P increased the levels 10
of GTP-bound Gαi and GTP-bound Gα13 in human platelets. Therefore, it is probable that S1P 11
actually activates Gi and G12/13 through S1PR4 and/or S1PR1 in human platelets, resulting in 12
the inhibition of collagen-mediated platelet aggregation. We next showed that not the selective 13
S1PR1 agonist SEW2871 [35] but the selective S1PR4 agonist CYM50260 [36] markedly 14
attenuated the collagen-induced platelet aggregation. In addition, CYM50260 suppressed the 15
phospholyration levels of HSP27 (Ser-78) induced by collagen in human platelets, but SEW2871 16
showed no effect on the HSP27 phosphorylation. These results suggest that the inhibitory 17
effects of S1P on the collagen-induced platelet aggregation and HSP27 phosphorylation in 18
human platelet are mediated through S1PR4 but not S1PR1. We also found that CYM50260 19
reduced the PDGF-AB secretion as well as the release of sCD40L and phosphorylated-HSP27 20
(Ser-78) from collagen-activated human platelets. These results suggest that S1PR4 but not 21
S1PR1 mediates the S1P signaling in human platelets and reduces the collagen-induced platelet 22
activation. Furthermore, we demonstrated that the selective S1PR4 antagonist CYM50358 23
[33] markedly reversed the suppression by S1P of collagen-induced HSP27 (Ser-78) 24
phosphorylation, almost to the level of collagen stimulation alone, suggesting that the inhibitory 25
effect of S1P on the phosphorylation of HSP27 is truly mediated through S1PR4 in human 26
16
platelets. Taken together, our findings suggest that S1PR4 mediates the signaling of S1P as a 1
receptor for S1P in human platelets, leading to the suppression of the collagen-stimulated 2
platelet aggregation and phosphorylation of HSP27, subsequently resulting in the inhibition of 3
both the secretion of PDGF-AB and the release of sCD40L and HSP27 from the activated 4
platelets into plasma.
5
PDGF-AB secreted from α-granule in activated human platelets is firmly established as a 6
potent mitogenic growth factor that mainly acts on connective tissue, such as vascular smooth 7
muscle cells, and promotes atherosclerosis [15]. In addition, sCD40L released from activated 8
human platelets is known to induce inflammatory responses in the endothelium [16], and high 9
plasma levels of sCD40L are associated with an increased risk of vascular events, including 10
acute coronary syndrome[38,39]. However, regarding the extracellular effects of HSP27, it has 11
been reported that HSP27 stimulates the activation of nuclear factor κB in macrophages, 12
leading to the secretion of both pro-inflammatory factors like IL-1β and anti-inflammatory 13
factors like IL-10 [21]. In addition, HSP27 released from ischemic myocardium reportedly 14
induces a pro-inflammatory response in human coronary vascular endothelium cells [40].
15
Based on our findings, by using selective S1PR4 agonists or selective S1PR4 antagonists, the 16
modulation of the S1P signaling in human platelets might be useful as a therapeutic tool for 17
atherosclerosis and diseases caused by accelerated platelet aggregation, such as thrombosis.
18
G13 signaling induces platelet shape changes and granule secretion [14]. Regarding the 19
effects of S1P on human platelets, it has been reported that S1P induces platelet shape changes 20
or aggregation in washed platelets [8]. In addition, using whole blood, Urtz et al. showed that 21
S1P triggers platelet aggregation via S1PR1 [11]. They also demonstrated that the 22
pharmacological S1PR1 inhibition does not affect the collagen-induced platelet aggregation, 23
whereas the S1PR1 inhibition suppresses ADP- and the protease-activated receptor 4-peptide 24
(PAR4-P)-induced aggregation, indicating that the endogenous S1P plays a role in the platelet 25
aggregation induced by not collagen but ADP or PAR4-P [11]. Furthermore, using PRP, it was 26
17
previously reported that S1P alone does not induce platelet aggregation, whereas the 1
S1P-pretreatment inhibits platelet aggregation induced by thrombin receptor-activating peptide 2
or noradrenaline [10]. Using PRP, we showed in the present study that S1P-pretreatment via 3
S1PR4 suppressed the collagen-induced platelet activation. In addition, we demonstrated that 4
the selective S1PR4 antagonist CYM50358 had little effect on the collagen-induced HSP27 5
phosphorylation in platelets, suggesting that the inhibition of S1PR4 signaling does not 6
potentiate the collagen-induced phosphorylation of HSP27 in the absence of exogenous S1P in 7
human platelets. The S1P contained in human platelets is released into the extracellular space 8
with its activation [4]. It therefore seems unlikely that the endogenous S1P affects the 9
collagen-induced platelet activation through S1PR4, under our experimental conditions. It 10
therefore seems likely that the discrepancies of the effects of S1P on human platelet aggregation 11
are due to the differences among the experimental conditions and the platelet sample 12
preparation adopted whole blood, washed platelets and PRP. In addition, regarding exogenous 13
S1P, co-stimulation with S1P and collagen suppressed collagen-induced platelet activation, but 14
the effect was weaker than that achieved with S1P-pretreatment. In the case of 15
S1P-pretreatment, it is probable that the effect of S1P alone is exerted faster than that of 16
collagen stimulation. Therefore, it seems to follow that the inhibitory effect with 17
S1P-pretreatment would be greater than that with co-stimulation. Further investigations are 18
required to clarify the details underlying the discordance in the effects of S1P on human platelet 19
activation.
20
In conclusion, our results strongly suggest that S1P suppresses collagen-induced 21
aggregation and the phosphorylation of HSP27 via S1PR4 but not S1PR1 in human platelets, 22
resulting in the suppression of PDGF-AB secretion and of the release of sCD40L and 23
phosphorylated-HSP27.
24 25 26
18 A
Acknowledgments 1
We thank Yumiko Kurokawa for her skillful technical assistance. This work was 2
supported in part by a Grants-in-Aid for Scientific Research (26462335) from the Ministry of 3
Education, Science, Sports and Culture of Japan, and Research Founding for Longevity Science 4
(25-4) from National Center for Geriatrics and Gerontology (NCGG), Japan.
5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
19 R
References 1
1. Spiegel S, Milstien S. Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat Rev 2
Mol Cell Biol 2003;4:397-407.
3
2. Proia RL, Hla T. Emerging biology of sphingosine-1-phosphate: its role in pathogenesis 4
and therapy. J Clin Invest 2015;125:1379-87.
5
3. Brinkmann V, Billich A, Baumruker T, Heining P, Schmouder R, Francis G, et al.
6
Fingolimod (FTY720): discovery and development of an oral drug to treat multiple 7
sclerosis. Nat Rev Drug Discov 2010;9:883-97.
8
4. Hla T, Galvani S, Rafii S, Nachman R. S1P and the birth of platelets. J Exp Med 9
2012;209:2137-40.
10
5. Thuy AV, Reimann CM, Hemdan NY, Graler MH. Sphingosine 1-phosphate in blood:
11
function, metabolism, and fate. Cell Physiol Biochem 2014;34:158-71.
12
6. Mahajan-Thakur S, Bohm A, Jedlitschky G, Schror K, Rauch BH.
13
Sphingosine-1-phosphate and its receptors: a mutual link between blood coagulation and 14
inflammation. Mediators Inflamm 2015;2015:831059.
15
7. Zhang L, Orban M, Lorenz M, Barocke V, Braun D, Urtz N, et al. A novel role of 16
sphingosine 1-phosphate receptor S1pr1 in mouse thrombopoiesis. J Exp Med 17
2012;209:2165-81.
18
8. Yatomi Y, Ruan F, Hakomori S, Igarashi Y. Sphingosine-1-phosphate: a 19
platelet-activating sphingolipid released from agonist-stimulated human platelets. Blood 20
1995;86:193-202.
21
9. Ulrych T, Bohm A, Polzin A, Daum G, Nusing RM, Geisslinger G, et al. Release of 22
sphingosine-1-phosphate from human platelets is dependent on thromboxane formation.
23
J Thromb Haemost 2011;9:790-8.
24
10. Nugent D, Xu Y. Sphingosine-1-phosphate: characterization of its inhibition of platelet 25
aggregation. Platelets 2000;11:226-32.
26
20
11. Urtz N, Gaertner F, von Bruehl ML, Chandraratne S, Rahimi F, Zhang L, et al.
1
Sphingosine 1-phosphate produced by sphingosine kinase 2 intrinsically controls platelet 2
aggregation in vitro and in vivo. Circ Res 2015;117:376-87.
3
12. Davi G, Patrono C. Platelet activation and atherothrombosis. N Engl J Med 4
2007;357:2482-94.
5
13. Furie B, Furie BC. Mechanisms of thrombus formation. N Engl J Med 2008;359:938-49.
6
14. Li Z, Delaney MK, O'Brien KA, Du X. Signaling during platelet adhesion and activation.
7
Arterioscler Thromb Vasc Biol 2010;30:2341-9.
8
15. Heldin CH, Westermark B. Mechanism of action and in vivo role of platelet-derived 9
growth factor. Physiol Rev 1999;79:1283-316.
10
16. Henn V, Slupsky JR, Grafe M, Anagnostopoulos I, Forster R, Muller-Berghaus G, et al.
11
CD40 ligand on activated platelets triggers an inflammatory reaction of endothelial cells.
12
Nature 1998;391:591-4.
13
17. Bakthisaran R, Tangirala R, Rao Ch M. Small heat shock proteins: Role in cellular 14
functions and pathology. Biochim Biophys Acta 2015;1854:291-319.
15
18. Landry J, Lambert H, Zhou M, Lavoie JN, Hickey E, Weber LA, et al. Human HSP27 is 16
phosphorylated at serines 78 and 82 by heat shock and mitogen-activated kinases that 17
recognize the same amino acid motif as S6 kinase II. J Biol Chem 1992;267:794-803.
18
19. Kato K, Hasegawa K, Goto S, Inaguma Y. Dissociation as a result of phosphorylation of 19
an aggregated form of the small stress protein, hsp27. J Biol Chem 1994;269:11274-8.
20
20. Kostenko S, Moens U. Heat shock protein 27 phosphorylation: kinases, phosphatases, 21
functions and pathology. Cell Mol Life Sci 2009;66:3289-307.
22
21. Salari S, Seibert T, Chen YX, Hu T, Shi C, Zhao X, et al. Extracellular HSP27 acts as a 23
signaling molecule to activate NF-kappaB in macrophages. Cell Stress Chaperones 24
2013;18:53-63.
25
22. Kato H, Adachi S, Doi T, Matsushima-Nishiwaki R, Minamitani C, Akamatsu S, et al.
26
21
Mechanism of collagen-induced release of 5-HT, PDGF-AB and sCD40L from human 1
platelets: role of HSP27 phosphorylation via p44/p42 MAPK. Thromb Res 2
2010;126:39-43.
3
23. Kato H, Takai S, Matsushima-Nishiwaki R, Adachi S, Minamitani C, Otsuka T, et al.
4
HSP27 phosphorylation is correlated with ADP-induced platelet granule secretion. Arch 5
Biochem Biophys 2008;475:80-6.
6
24. Kageyama Y, Doi T, Akamatsu S, Kuroyanagi G, Kondo A, Mizutani J, et al. Rac 7
regulates collagen-induced HSP27 phosphorylation via p44/p42 MAP kinase in human 8
platelets. Int J Mol Med 2013;32:813-8.
9
25. Doi T, Adachi S, Takai S, Matsushima-Nishiwaki R, Kato H, Enomoto Y, et al.
10
Antithrombin III suppresses ADP-induced platelet granule secretion: inhibition of 11
HSP27 phosphorylation. Arch Biochem Biophys 2009;489:62-7.
12
26. Iida Y, Doi T, Matsushima-Nishiwaki R, Tokuda H, Ogura S, Kozawa O, et al.
13
(-)-Epigallocatechin gallate selectively inhibits adenosine diphosphatestimulated human 14
platelet activation: suppression of heat shock protein 27 phosphorylation via p38 15
mitogenactivated protein kinase. Mol Med Rep 2014;10:1383-8.
16
27. Tokuda H, Kuroyanagi G, Tsujimoto M, Enomoto Y, Matsushima-Nishiwaki R, Onuma T, 17
et al. Release of Phosphorylated HSP27 (HSPB1) from Platelets Is Accompanied with the 18
Acceleration of Aggregation in Diabetic Patients. PLoS One 2015;10:e0128977.
19
28. Laemmli UK. Cleavage of structural proteins during the assembly of the head of 20
bacteriophage T4. Nature 1970;227:680-5.
21
29. Rendu F, Brohard-Bohn B. The platelet release reaction: granules' constituents, secretion 22
and functions. Platelets 2001;12:261-73.
23
30. Hermann A, Rauch BH, Braun M, Schror K, Weber AA. Platelet CD40 ligand 24
(CD40L)--subcellular localization, regulation of expression, and inhibition by clopidogrel.
25
Platelets 2001;12:74-82.
26
22
31. Saklatvala J, Rawlinson L, Waller RJ, Sarsfield S, Lee JC, Morton LF, et al. Role for p38 1
mitogen-activated protein kinase in platelet aggregation caused by collagen or a 2
thromboxane analogue. J Biol Chem 1996;271:6586-9.
3
32. Hanai Y, Adachi S, Yasuda I, Takai S, Matsushima-Nishiwaki R, Kato H, et al.
4
Collagen-induced p38 MAP kinase activation is a biomarker of platelet 5
hyper-aggregation in patients with diabetes mellitus. Life Sci 2009;85:386-94.
6
33. Brinkmann V. Sphingosine 1-phosphate receptors in health and disease: mechanistic 7
insights from gene deletion studies and reverse pharmacology. Pharmacol Ther 8
2007;115:84-105.
9
34. Ritter SL, Hall RA. Fine-tuning of GPCR activity by receptor-interacting proteins. Nat 10
Rev Mol Cell Biol 2009;10:819-30.
11
35. Sanna MG, Liao J, Jo E, Alfonso C, Ahn MY, Peterson MS, et al. Sphingosine 12
1-phosphate (S1P) receptor subtypes S1P1 and S1P3, respectively, regulate lymphocyte 13
recirculation and heart rate. J Biol Chem 2004;279:13839-48.
14
36. Guerrero M, Urbano M, Zhao J, Crisp M, Chase P, Hodder P, et al. Discovery, design and 15
synthesis of novel potent and selective sphingosine-1-phosphate 4 receptor (S1P(4)-R) 16
agonists. Bioorg Med Chem Lett 2012;22:537-42.
17
37. Guerrero M, Urbano M, Velaparthi S, Zhao J, Schaeffer MT, Brown S, et al. Discovery, 18
design and synthesis of the first reported potent and selective sphingosine-1-phosphate 4 19
(S1P4) receptor antagonists. Bioorg Med Chem Lett 2011;21:3632-6.
20
38. Schonbeck U, Varo N, Libby P, Buring J, Ridker PM. Soluble CD40L and cardiovascular 21
risk in women. Circulation 2001;104:2266-8.
22
39. Heeschen C, Dimmeler S, Hamm CW, van den Brand MJ, Boersma E, Zeiher AM, et al.
23
Soluble CD40 ligand in acute coronary syndromes. N Engl J Med 2003;348:1104-11.
24
40. Jin C, Cleveland JC, Ao L, Li J, Zeng Q, Fullerton DA, et al. Human myocardium 25
releases heat shock protein 27 (HSP27) after global ischemia: the proinflammatory effect 26
23
of extracellular HSP27 through toll-like receptor (TLR)-2 and TLR4. Mol Med 1
2014;20:280-9.
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
24 T
Table 1. Effect of S1P on the collagen-induced platelet aggregation 1
2
S1P (μM) 0 10 20 30
collagen + + + +
Transmittance (%) 86.6 ± 6.0 77.6 ± 10.6 47.0 ± 10.0 33.8 ± 9.2 Large (%) 49.0 ± 6.2 35.4 ± 5.1 16.0 ± 5.1 17.0 ± 4.0 Medium (%) 19.8 ± 1.2 21.8 ± 1.5 22.0 ± 3.4 16.0 ± 4.3 Small (%) 31.2 ± 5.1 42.6 ± 4.1 62.0 ± 8.2 67.0 ± 7.8
PRP was pretreated with various doses of S1P at 37°C for 15 min and then stimulated by 1.0 3
μg/ml of collagen for 5 min. The reaction was terminated by the addition of ice-cold EDTA 4
solution. The results obtained from the aggregometer for the transmittance and the ratio of 5
large aggregates, medium aggregates and small aggregates are summarized. Each value 6
represents the mean ± SEM of five healthy donors.
p
<0.05 compared to the value of collagen 7alone.
8 9 10 11 12 13 14 15 16 17 18 19
25 T
Table 2. The comparison of the effect of S1P between the pretreatment and the co-stimulation on 1
the collagen-induced platelet aggregation 2
3
pretreatment group co-stimulation group
S1P - + - +
collagen + + + +
Transmittance (%) 70.5 ± 1.5 41.5 ± 5.5 73.0 ± 4.0 56.0 ± 3.6 Large (%) 39.8 ± 2.5 17.3 ± 5.4 37.5 ± 2.8 24.8 ± 4.1 Medium (%) 25.2 ± 1.5 23.0 ± 2.4 26.8 ± 1.1 19.5 ± 4.8 Small (%) 34.8 ± 1.3 59.8 ± 7.7 35.5 ± 2.2 49.0 ± 5.0
PRP was pretreated with 30 μM of S1P or vehicle at 37°C for 15 min and then stimulated by 4
collagen for 5 min (pretreatment group) or co-stimulated by collagen with 30 μM of S1P or 5
vehicle for 5 min (co-stimulation group). The reaction was terminated by the addition of 6
ice-cold EDTA solution. The results obtained from the aggregometer for the transmittance and 7
the ratio of large aggregates, medium aggregates and small aggregates are summarized. Each 8
value represents the mean ± SEM of four healthy donors.
p
<0.05 compared to the value of 9collagen alone in the same group.
10 11 12 13 14 15 16 17 18
26 T
Table 3. Effects of SEW2871 or CYM50260 on the collagen-induced platelet aggregation 1
2
SEW2871 CYM50260
- + - +
collagen + + + +
Transmittance (%) 81.6 ± 4.0 80.4 ± 2.5 86.4 ± 7.6 66.6 ± 3.2 Large (%) 43.6 ± 4.1 43.8 ± 3.4 44.4 ± 3.2 27.4 ± 3.1 Medium (%) 20.6 ± 1.1 19.4 ± 1.7 23.4 ± 0.8 24.4 ± 1.2
Small (%) 36.0 ± 3.6 36.6 ± 2.5 32.0 ± 2.8 48.2 ± 3.9 PRP was pretreated with 10 μM of SEW2871, 25 μM of CYM50260 or vehicle at 37°C for 15 min 3
and then stimulated by collagen for 5 min. The reaction was terminated by the addition of 4
ice-cold EDTA solution. The results obtained from the aggregometer for the transmittance and 5
the ratio of large aggregates, medium aggregates and small aggregates are summarized. Each 6
value represents the mean ± SEM of five healthy donors.
p
<0.05 compared to the value of 7collagen alone in the same group.
8 9 10 11 12 13 14 15 16 17 18
27 F
Figure Legends 1
2
Figure 1. Effect of S1P on the collagen-induced platelet aggregation. PRP was pretreated with 3
various doses of S1P at 37°C for 15 min and then stimulated by 1.0 μg/ml of collagen for 5 min.
4
The reaction was terminated by the addition of ice-cold EDTA solution. The black line 5
indicates the percentage of transmittance of each sample (isolated platelets recorded as 0%, and 6
platelet-poor plasma recorded as 100%). The blue line indicates small aggregates (9-25 μm);
7
green line, medium aggregates (25-50 μm); red line, large aggregates (50-70 μm). The lower 8
panel presents the distribution (%) of aggregated particle size as measured by laser-scattering.
9
The representative results obtained from five healthy donors are shown.
10 11
Figure 2. Effects of S1P on the collagen-induced platelet aggregation: comparison between the 12
pretreatment and the co-stimulation. (A) PRP was pretreated with 30 μM of S1P or vehicle at 13
37°C for 15 min and then stimulated by collagen for 5 min. (B) PRP was co-stimulated by 14
collagen with 30 μM of S1P or vehicle for 5 min. The reaction was terminated by the addition of 15
ice-cold EDTA solution. The blue line indicates small aggregates (9-25 μm); green line, medium 16
aggregates (25-50 μm); red line, large aggregates (50-70 μm). The lower panel presents the 17
distribution (%) of aggregated particle size as measured by laser-scattering. The 18
representative results obtained from four healthy donors are shown.
19 20
Figure 3. Effects of S1P on the collagen-induced secretion of PDGF-AB, the release of sCD40L or 21
the release of phosphorylated-HSP27. PRP was pretreated with various doses of S1P (A, B) or 22
30 μM of S1P (C) at 37°C for 15 min and then stimulated by collagen or vehicle for 15 min. The 23
reaction was terminated by the addition of ice-cold EDTA solution. The mixture was 24
centrifuged at 10,000 ×
g
at 4°C for 2 min, and the supernatant was then subjected to ELISA for 25PDGF-AB (A), sCD40L (B) and phosphorylated-HSP27 (Ser-78) (C), respectively. The net 26
28
increase in the levels of collagen alone is represented as 100%. The results from five healthy 1
donors are shown. Each value of PDGF-AB (A), sCD40L (B) and phosphorylated-HSP27 2
(Ser-78) (C) represents the mean ± SEM.
p
<0.05 compared to the value of collagen alone.3 4
F
Figure 4. Effects of S1P on the phosphorylation of HSP27 or p38 MAP kinase induced by 5
collagen. PRP was pretreated with 30 μM of S1P or vehicle at 37°C for 15 min and then 6
stimulated by collagen for 90 sec. The reaction was terminated by the addition of ice-cold 7
EDTA solution. The extracts of platelets were then subjected to SDS-PAGE with a subsequent 8
Western blot analysis using antibodies against (A) phospho-specific HSP27 (Ser-78) or GAPDH, 9
or (B) phospho-specific p38 MAP kinase or GAPDH. The histogram shows a quantitative 10
representation of the collagen-induced levels obtained from a densitometric analysis. The 11
phosphorylation is expressed as the fold increase compared to the basal levels, presented as lane 12
1. Each value represents the mean ± SEM of three independent experiments.
p
<0.05 13compared to the control value.
p
<0.05 compared to the value of collagen alone.14 15
Figure 5. Effects of S1P on the activation of Gi and G13 in human platelets. PRP was 16
stimulated by 30 μM of S1P at 37°C for 30 sec, and then the reaction was terminated by the 17
addition of ice-cold EDTA solution. (A) Using an activation assay kit for Gαi, the protein 18
extracts of platelets were harvested, and GTP-bound Gαi (GTP-Gαi) or total Gαi was detected by 19
a Western blot analysis performed in accordance with the manufacturer’s instructions. (B) 20
Using an activation assay kit for Gα13, the protein extracts of platelets were harvested, and 21
GTP-bound Gα13 (GTP-Gα13) or total Gα13 was detected by a Western blot analysis performed 22
in accordance with the manufacturer’s instructions 23
24
Figure 6. Effects of SEW2871 or CYM50260 on the collagen-induced platelet aggregation. PRP 25
was pretreated with 10 μM of SEW2871 (A), 25 μM of CYM50260 (B) or vehicle at 37°C for 15 26
29
min and then stimulated by collagen for 5 min. The reaction was terminated by the addition of 1
ice-cold EDTA solution. The black line indicates the percentage of transmittance of each 2
sample (isolated platelets recorded as 0%, and platelet-poor plasma recorded as 100%). The 3
blue line indicates small aggregates (9-25 μm); green line, medium aggregates (25-50 μm); red 4
line, large aggregates (50-70 μm). The lower panel presents the distribution (%) of aggregated 5
particle size as measured by laser-scattering. The representative results obtained from five 6
healthy donors are shown.
7 8
F
Figure 7. Effects of SEW2871 or CYM50260 on the collagen-induced secretion of PDGF-AB, the 9
release of sCD40L or the release of phosphorylated-HSP27. PRP was pretreated with 10 μM of 10
SEW2871, 25 μM of CYM50260 or vehicle at 37°C for 15 min and then stimulated by collagen for 11
15 min. The reaction was terminated by the addition of ice-cold EDTA solution. The mixture 12
was centrifuged at 10,000 ×
g
at 4°C for 2 min, and the supernatant was then subjected to 13ELISA for PDGF-AB (A), sCD40L (B) and phosphorylated-HSP27 (Ser-78) (C). The net 14
increase in the levels of collagen alone is represented as 100%. The results from five (A, B) or 15
three (C) healthy donors are shown. Each value of PDGF-AB (A), sCD40L (B) and 16
phosphorylated-HSP27 (Ser-78) (C) represents the mean ± SEM.
p
<0.05 compared to the 17value of collagen alone. N.S.: no significant difference between the indicated pairs.
18 19
Figure 8. Effects of SEW2871 or CYM50260 on the collagen-induced phosphorylation of HSP27.
20
PRP was pretreated with 10 μM of SEW2871 (A), 25 μM of CYM50260 (B) or vehicle at 37°C for 21
15 min and then stimulated by collagen for 90 sec. The reaction was terminated by the 22
addition of ice-cold EDTA solution. The extracts of platelets were then subjected to SDS-PAGE 23
with a subsequent Western blot analysis using antibodies against phospho-specific HSP27 24
(Ser-78) or GAPDH. The histogram shows the quantitative representation of the 25
collagen-induced levels obtained from a densitometric analysis. The phosphorylation is 26
30
expressed as the fold increase compared to the basal levels, presented as lane 1. Each value 1
represents the mean ± SEM of five (A) or three (B) independent experiments. N.S.: no 2
significant difference between the indicated pairs.
p
<0.05 compared to the control value.3 4
F
Figure 9. Effects of CYM50358 on the collagen-induced HSP27 phosphorylation or the 5
suppression by S1P of collagen-induced HSP27phosphorylation. (A) PRP was pretreated with 6
10 μM of CYM50358 or vehicle at 37°C for 15 min and then stimulated by collagen for 90 sec.
7
(B) PRP was first pretreated with 10 μM of CYM50358 or vehicle at 37°C for 1 min, and then 8
pretreated with 30 μM of S1P or vehicle for 15 min. PRP was then stimulated by collagen at 9
37°C for 90 sec. The reaction was terminated by the addition of ice-cold EDTA solution. The 10
extracts of platelets were then subjected to SDS-PAGE with a subsequent Western blot analysis 11
using antibodies against phospho-specific HSP27 (Ser-78) or GAPDH. The histogram shows a 12
quantitative representation of the collagen-induced levels obtained from a densitometric 13
analysis. The phosphorylation is expressed as the fold increase compared to the basal levels, 14
presented as lane 1. Each value represents the mean ± SEM of four (A) or six (B) independent 15
experiments. N.S.: no significant difference between the indicated pairs.
p
<0.05 compared to 16the control value.
p
<0.05 compared to the value of collagen with S1P.17