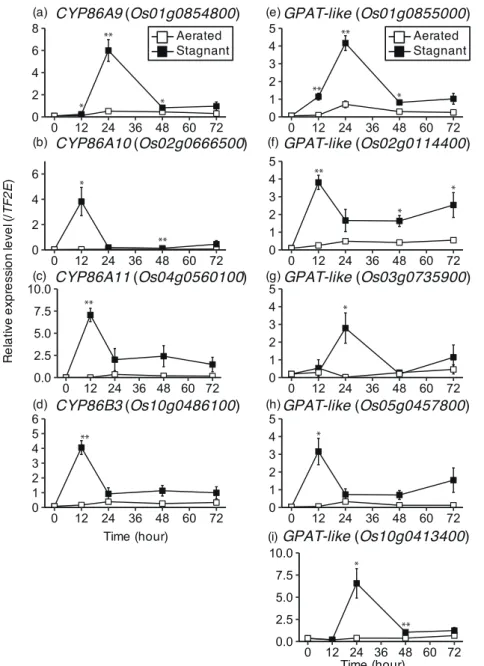
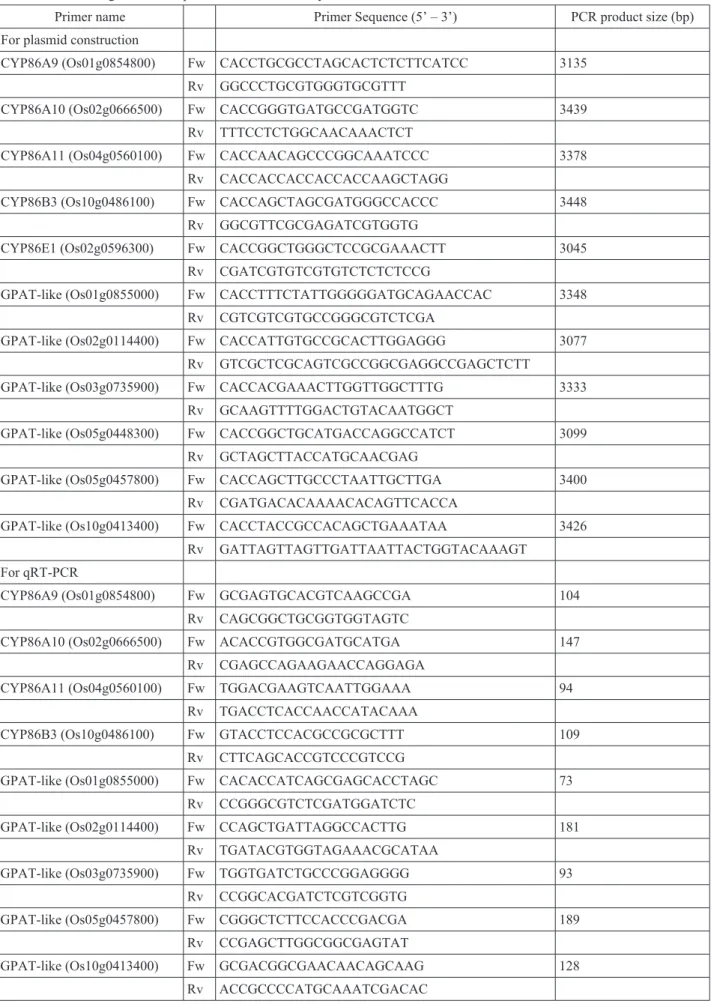
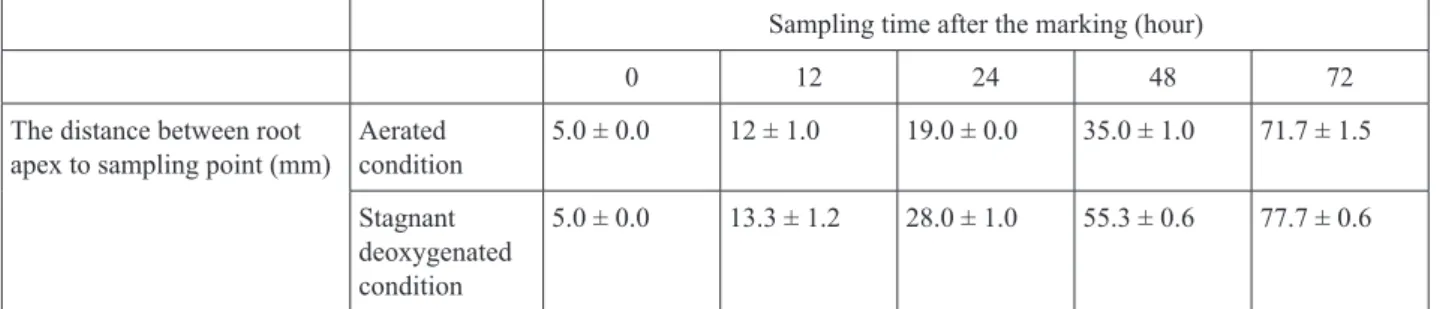
Expression analysis of genes for cytochrome P450 CYP86 and glycerol-3-phosphate acyltransferase related to suberin biosynthesis in rice roots under stagnant deoxygenated conditions
19
0
0
全文
図



+3
関連したドキュメント
Effect of gemcitabine on the expression of apoptosis-related genes in human pancreatic cancer cells.
reported that gemcitabine-mediated apoptosis is caspase- dependent in pancreatic cancers; Jones et al [14] showed that gemcitabine-induced apoptosis is achieved through the
To determine whether expression of HPV genes had any influence upon HIF-1α activation or levels in normoxia and hypoxia, we first examined whether HIF-1α levels were induced
Relative expression levels of RaSTM (open) and RaYAB2 (shaded) in phylloclade primordia (PP), shoot apices and bud scales subtending them (S), floral buds (F), scale leaves (L)
Thalidomide (50 mg/kg) was administered 22 h and 2 h before endotoxin injection (1 mg/kg). Blood samples were collected at designated intervals to measure concentra- tions of NOx
Expression of cereolysine AB genes in Bacillus anthracis vaccine strain ensures protection against experimental hemolytic anthrax infection. Vaccine,
熱力学計算によれば、この地下水中において安定なのは FeSe 2 (cr)で、Se 濃度はこの固相の 溶解度である 10 -9 ~10 -8 mol dm
These results are motivated by the bounds for real subspaces recently found by Bachoc, Bannai, Coulangeon and Nebe, and the bounds generalize those of Delsarte, Goethals and Seidel
[Mag3] , Painlev´ e-type differential equations for the recurrence coefficients of semi- classical orthogonal polynomials, J. Zaslavsky , Asymptotic expansions of ratios of
