起波長560nm,蛍光波長 580 nm)を分光蛍光プレートリーダー(FluorPlus,Tecan)により 定量した。 3)珪藻 Asteroplanus karianus 溶藻因子の性状解析 ここでは,あらかじめ調製・保存しておいた海水懸濁物を材料に用いた。海水試料は, 2014 年 9 月の広島湾表層海水,ならびに 2015 年 1 月有明海における A. karianus ブルーム 期間中の表層と底層の混合水である。これらに含まれる懸濁粒子を孔径 3 μm ヌクレポア フィルター上に捕集し,−80℃で凍結保存した。当該サンプルを試験直前に解凍し,RNA 抽出・cDNA 作製を実施した。cDNA 作製にはピコルナ系ウイルス RdRP の増幅プライマ ー(MplSC)ならびにランダムプライマー6mer を用いた。さらに,作製 cDNA(鋳型)と MplSC プライマーを用いた PCR により,懸濁物中に存在していた可能性のあるピコルナ 系ウイルスのポリメラーゼ遺伝子領域の増幅を試みた。 (3)結果及び考察 1)珪藻 Asteroplanus karianus の栄養塩利用に関わる遺伝子の分布 有明海北部では,冬季になると毎年のようにA. karianus が発生し,養殖ノリの栄養不良・ 色落ち被害を引き起こしている(松原ほか2014,Yamaguchi et al. 2014)。そのため,本種 の栄養塩摂取プロセスを明らかにし,それに基づいた栄養塩の変動予測技術を確立するこ とが重要である。今回,A. karianus による栄養塩摂取を理解するために,本種無菌クローン 株由来の mRNA を網羅解析したところ,多数の配列を得ることができ,とりわけ栄養塩利用に深 く関わるとされる硝酸塩輸送体およびリン酸塩輸送体の配列を見出すことができた(表 1)。これま での事業成果により,珪藻 Chaetoceros tenuissimus は窒素あるいはリン欠乏に陥ると,それぞれ 硝酸塩輸送体およびリン酸塩輸送体の転写量を増大させることが明らかにされている。したがっ て,それらの転写量に基づいて,窒素あるいはリン欠乏状態を推定することが可能と考えられる。 今回,その転写量の相対定量に欠かせないハウスキーピング遺伝子の候補も得られた。現段 階では,アクチンならびにTATA 結合タンパク質の配列がそれに該当する。当初の予定よりもアノ テーションが進んでいないが,次年度に行う解析によって,より多くのハウスキーピング遺伝子の 候補が得られるであろう。また今後,それらを用いた逆転写定量PCR により,A. karianus の窒素・ リン利用,それらに関わる生理状態が診断可能になるものと期待される。 興味深いことに,昨年度,A. karianus 無菌クローン株からはアルカリフォスファターゼ・フォスフ ォジエステラーゼの活性が検出されていないにもかかわらず,今回の解析ではそれらの遺伝子発 現が認められた。これらを総合すると,A. karianus は有機態リン分解に関わる酵素群 DNA を有し ているものの,それらの転写量あるいは翻訳は何らかの理由で制限されているようである。そのた め,有機態リン分解活性としては検出できないレベルにあるのではないかと考えられる。今後,得 られた各遺伝子のmRNA 量(転写)と活性(翻訳)との関係を精査する必要がある。 2)正リン酸塩を迅速に蛍光定量可能な系の確立 水中の正リン酸塩は,モリブデン酸と結合することで,モリブドリン酸と呼ばれる錯体 を形成する。この錯体を還元することにより正リン酸塩を比色定量するのが,いわゆる定 法“モリブデンブルー法”である。我々が着目したのは,モリブドリン酸の濃度に比例し て,試薬ローダミンB 由来の蛍光が減衰するという化学反応(本水ほか 2000)である。本 研究では,当該反応を 96 穴プレートで引き起こし,ローダミン B 由来の蛍光強度をプレ ートリーダーで高速測定することで,正リン酸塩の迅速なる蛍光定量系の基盤構築を果た
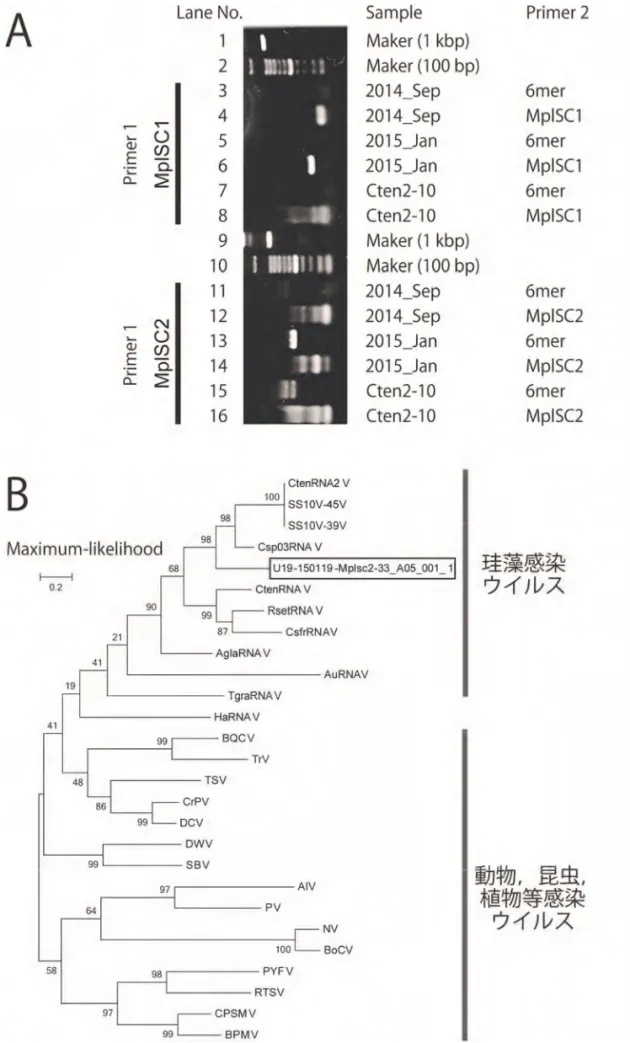
すことに成功した。 今回の試験において正リン酸塩濃度(Pi)の増大に伴う蛍光強度(Fluor)の減少が認め られ(図1),両者の関係は以下の対数関数(1)で表すことができた。 ܲ ൌ െͲǤͷͶ ή ܨ݈ݑݎ ǤͲ͵ (1) 本式から得られた理論値と実測値との間には,有意かつ高精度な相関関係(R ≥ 0.991,P < 0.01,N = 24)が認められたことから,蛍光値から正リン酸を回帰できると判断された。 現在,QuAAtro(BL TEC K.K.)に代表される栄養塩自動分析であれば,多検体にのぼる 正リン酸塩の定量を約1 時間程度で処理可能である。従来の手分析ベースの定量に比べる と,労力の軽減が大幅に改善されているのは周知のとおりである。その一方で,試料の用 量は 5~10 mL/sample であり,また,それらを凍結保存するのであれば,相当の保管スペ ースが必要となる。今回,ローダミンB の消光法と定量機器プレートリーダーを組み合わ せることで,100 近い多くの検体であっても 1 分以内に測定可能なことが示された。しか も,定量に要する試料は,一試料当たり200 μL 程度に満たないほどの微量で事足りる。こ のように,今回検討した正リン酸塩の定量系は,試料の用量削減,それに伴う保存スペー スの削減,さらに測定時間の大幅な短縮が図れる点で優れていると判断される。したがっ て,正リン酸塩の定量が関わる実験を効率よく推し進める上で,今回の手法は有用なツー ルになろうと思われる。 モリブデンと錯体を形成する化合物の一つにケイ酸塩が挙げられる。ケイ酸塩と正リン 酸塩が共存する場合,前者が後者の定量に及ぼす負の影響を考慮する必要がある。現段階 では予備的ではあるものの,本定量結果に及ぼすケイ酸塩の影響は無視できるほど小さい ことが明らかになりつつある。今後は,これら各種栄養塩ならびに塩分が蛍光値に及ぼす 影響を検討することで,海水中の正リン酸定量の正確性・迅速性について評価できると期 待される。 3)珪藻 Asteroplanus karianus 溶藻因子の性状解析 近年の成果により,A. karianus の栄養塩摂取プロセスの解析は進展しつつあるものの, 摂取停止ひいてはノリ養殖場の栄養塩回復・再生につながり得る本種の衰退プロセスにつ いてはほとんど理解されていない。昨年度に実施した培養試験によって,我々はA. karianus 推定溶藻因子を発見するに到った。しかしながら,本種ブルームに対する当該因子の関与 については不明であり,A. karianus の衰退現象とそれに伴う栄養塩摂取停止現象について は考察できなかった。 本年度,有明海において発生したA. karianus 冬季ブルーム期間中の懸濁物サンプルより, ピコルナ系ウイルス(ssRNA ウイルス)のポリメラーゼ様配列(約 500 bp,MplSC2 + 6mer, 2015_Jan)を検出することに成功した(図 2A)。その配列を分子系統解析に供した結果,当 該配列は,珪藻感染ウイルスの系統に含まれるものの,新規な配列であることが示唆され た(図2B)。 これまでに,我々はA. karianus 培養株の溶藻現象を見出し,その際行った透過電子顕微 鏡観察により,ウイルス粒子と推察される因子を検出することに成功している。今回,A. karianus ブルームの懸濁粒子から新規珪藻感染 RNA ウイルス配列を検出することができ, それを含めて総合的に考察すると,A. karianus 個体群減衰要因の一つに,ウイルス感染の 影響が可能性として推察される。よってA. karianus 冬季ブルームに伴う栄養塩の消費を予
測する上で,ウイルス感染による消費停止プロセスは無視すべきではないと考える。 本事業の遂行にあたり,貴重なサンプルの提供にご協力いただいた紫加田知幸博士・松 原賢博士に厚く御礼申し上げます。 4)引用文献 松原賢, 横尾一成, 川村嘉応. 有害珪藻 Asteroplanus karianus の有明海佐賀県海域における 出現動態と各種環境要因との関係. 日水誌, 80, 222–232, 2014. 本水昌二, 大島光子, 勝村直也. 海水中のリンの蛍光検出フローインジェクション法. J. Flow Injection Anal., 16, 101, 2000.
Yamaguchi H, Minamida M, Matsubara T, Okamura K. Novel blooms of the diatom Asteroplanus karianus deplete nutrients from Ariake Sea coastal waters. Mar. Ecol. Prog. Ser., 517, 51–60, 2014.
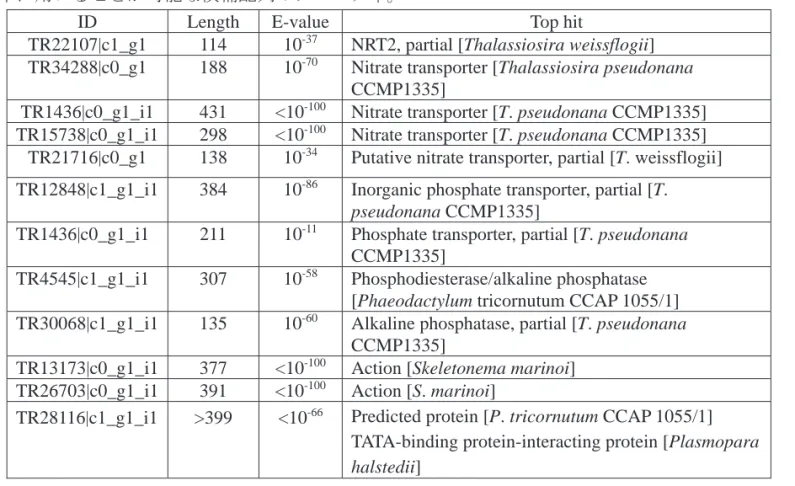
表 1 Asteroplanus karianus の栄養塩利用に関わる cDNA 配列のプロパティ。ならびに内部標 準に用いることが可能な候補配列のプロパティ。
ID Length E-value Top hit
TR22107|c1_g1 114 10-37 NRT2, partial [Thalassiosira weissflogii] TR34288|c0_g1 188 10-70 Nitrate transporter [Thalassiosira pseudonana
CCMP1335]
TR1436|c0_g1_i1 431 <10-100 Nitrate transporter [T. pseudonana CCMP1335] TR15738|c0_g1_i1 298 <10-100 Nitrate transporter [T. pseudonana CCMP1335]
TR21716|c0_g1 138 10-34 Putative nitrate transporter, partial [T. weissflogii] TR12848|c1_g1_i1 384 10-86 Inorganic phosphate transporter, partial [T.
pseudonana CCMP1335]
TR1436|c0_g1_i1 211 10-11 Phosphate transporter, partial [T. pseudonana CCMP1335]
TR4545|c1_g1_i1 307 10-58 Phosphodiesterase/alkaline phosphatase [Phaeodactylum tricornutum CCAP 1055/1] TR30068|c1_g1_i1 135 10-60 Alkaline phosphatase, partial [T. pseudonana
CCMP1335]
TR13173|c0_g1_i1 377 <10-100 Action [Skeletonema marinoi] TR26703|c0_g1_i1 391 <10-100 Action [S. marinoi]
TR28116|c1_g1_i1 >399 <10-66 Predicted protein [P. tricornutum CCAP 1055/1] TATA-binding protein-interacting protein [Plasmopara halstedii]
図 1 正リン酸濃度(Pi)とローダミン B 由来の蛍光値(Fluorescence)との関係.図中の 破線は想定される直線回帰の傾向を示し,実線は正リン酸塩の回帰曲線を示している.
図 2 2014 年 9 月広島湾海水ならびに 2015 年 1 月 Asteroplanus karianus ブルーム海水に含ま れる懸濁粒子より得られた PCR 産物の電気泳動像(A),2015 年 1 月サンプルの一部(囲み 指定)より得られた配列とその他のウイルス配列に基づいて作成した最尤系統樹(B)。鋳型 には懸濁粒子由来の RNA(cDNA)を用い,cDNA 合成および PCR のプライマーにはピコル ナ系 RNA ウイルス検出用 deg.プライマー(MplSC1 および MplSC2)・ランダムプライマー 6mer)を用いた。分子系統樹の分岐箇所にはブートストラップ値が示されている。
4)栄養塩類等の水質環境が低次生産生物に及ぼす影響解明
①海域の栄養塩環境が低次生産に及ぼす影響解明
ア.播磨灘における基礎生産簡易測定技術の開発
兵庫県立農林水産技術総合センター水産技術センター 宮原一隆,原田和弘 水産研究・教育機構 瀬戸内海区水産研究所 阿保勝之 1 全体計画 (1) 目的 近年,瀬戸内海海域では,貧栄養化が進行したことによる水産資源への影響が指摘され ている。同海域の低次生物生産に関しては,各種モニタリングによって現存量のデータは 整備されつつあるものの,生産速度や転換効率に関する知見は断片的であり,瀬戸内海各 海域においてどのような生物生産構造が成り立っているのかについては明らかになって いない。そこで.瀬戸内海東部の播磨灘において,栄養塩等の海域の水質環境調査を実施 するとともに動・植物プランクトンの低次生物生産に関する調査を行い,水質環境が海域 の生産力に及ぼす影響を把握するとともに,基礎生産速度等を簡易に測定する技術の開発 を目指す。 (2) 試験等の方法 瀬戸内海東部の播磨灘において定点(図1,計 6 定点)を設置し,年 5 回の現場観測調 査を実施する。水温,塩分,光条件等の鉛直プロファイルを獲得するとともに,採水調 査およびネット採集調査を実施し,低次生物生産量を算定する。また,光合成活性を測 定する機器(パルス変調型光合成測定装置)を活用し現場海域での水柱基礎生産量を推 定するとともに,従来法(疑似現場培養法)による測定結果と比較検討する。 2 平成 28 年度計画および結果 (1) 目的 全体計画と同じ。 (2) 試験等の方法 全体計画と同じ。 平成28 年 6 月 3 日,7 月 15 日,9 月 13 日,11 月 16 日,平成 29 年 1 月 16 日,3 月 9 日(予定)に調査を実施し,各分析を実施した(本報告では平成29 年 1 月調査までの 5 回分をとりまとめた)。なお,単年の調査結果のみでは,基礎生産や光合成活性の季節変 動や各環境要因との関連について十分には議論できないため,本報告書では,各調査期 における植物プランクトンの発生状況,パルス変調による光合成測定(以下,PAM 測定) の結果と明暗瓶法との比較を中心に記載した。 調査分析項目と分析方法: 1) 全点実施(Stn. H1,H2,H6,H7,H10,H29;図 1)・CTD 観測(水温・塩分・蛍光強度・DO),透明度
多項目式CTD(JFE アドバンテック RINKO Profiler),セッキ板を用いて測定した。 ・動物プランクトン採集(改良型ノルパックネット,100μm メッシュ) 濾水計を装着したネットを海底まで垂下し,おもり着底後から海面まで約 1m/秒の 速度で鉛直曳網した。採取したネットプランクトンサンプルは,中性ホルマリン(最 終濃度約10%)で固定し,課題 4-①-エの解析に供した。 ・栄養塩(DIN,DIP,D-Si),クロロフィル a(Chl.a)濃度,植物プランクトン組成 栄養塩濃度は,自動分析装置(QuAAtro,ビーエルテック株式会社)を用いた比色 法によって測定した。Chl.a 濃度は,アセトン抽出蛍光法を用いた。植物プランクトン 組成は,原則として表層(0.5m 深)の試水 1mL に含まれる全数を計数した。 ・PAM 測定 光合成測定装置(WATER-PAM CUVETTE-Version,WALZ 社)を用いた。測定用の 試水は表層から採水し,300μm メッシュのネットにより動物プランクトン等の捕食生 物を除去した。濾水を500mL 褐色瓶に入れて密封・遮光した後,船上に設置した大型 コンテナ内に海水を満たし,その中で保管することによって温度を一定に保った(海 水は適宜交換した)。 光合成の測定は,研究室に戻り次第実施した。測定条件は,試水量 1.2mL(専用キ ュベットを使用),ML レベル 12 以下,Gain レベルはデフォルト,PM-gain レベル 14 以下,PAR(光合成有効放射)の測定間隔は 30 秒とした。測定 PAR 値は,機器固有 の設定(0~2678 の 9 段階)を使用し,それぞれの PAR 条件で得られた rETR(=PAR ×ΦII,相対電子伝達速度,μmol electron mg/Chl.a/s)とのデータセットから,光合成 曲線:rETR=ETRmax×(1-exp(-(a×PAR)/ETRmax))にあてはめた場合の残差平方和が最小 となるように2 種類のパラメータ(ETRmax=最大電子伝達速度μmol electron mg/Chl.a/s, a=曲線の傾きを示す指標)を算出した。 なお,以下の特定点では,透明度の測定値から,光量子量が表層の 48%(以下,中 層 1),33%(中層 2)にそれぞれ減衰する層を船上で推定し,それぞれから採取した 中層水についてもPAM 測定を実施し,表層水と同様の方法で光合成曲線を推定した。 2) 特定点実施(Stn.H2 と H10) ・光量子量測定
光量子計(LI-COR 社球形光量子センサーまたは JFE アドバンテック DEFI-L)を用 いて,鉛直垂下により現場観測した。 ・明暗瓶法(酸素法) PAM 測定に供した海水(表層,中層 1,中層 2 の計 3 層分)と同じ試水を用いた。 それぞれの試水を明瓶と暗瓶(各 duplicate)にてインキュベータ内で 12 時間培養し た後,溶存酸素濃度の差から算出した。培養温度は,調査日の播磨灘北部(兵庫県明 石市二見町地先,水産技術センター定置観測)の水温とした。光条件は,蛍光灯を用 いて,表層海水は480μmol/m2/s,中層 1 はその 48%,中層 2 は 33%となるように黒色 遮光幕を使って減衰設定した。各 2 本の平均値の明暗瓶差から酸素発生量を算出し,