運動における分岐鎖アミノ酸の生理機能
Physiological Functions of Branched-Chain Amino
Acids in Exercise
名古屋大学大学院生命農学研究科
応用分子生命科学専攻
応用生命化学講座
栄養生化学研究分野
徐
旻珺
2018 年 3 月
目次
略語 1
第一章 緒言
1.1. 分岐鎖アミノ酸とは 4 1.2. BCAA 代謝とその調節 5 1.3. 筋特異的 BCAA 代謝亢進動物モデル 7第二章 筋特異的
BCAA 分解亢進が運動持久力と運動中の代謝変動に与える影響
2.1. 序論 21 2.2. 材料及び方法 22 2.3. 結果 32 2.4. 考察 35第三章 筋特異的
BCAA 分解亢進マウスの運動適応性に対する BCAA 投与の影響
3.1. 序論 54 3.2. 材料及び方法 55 3.3. 結果 57 3.4. 考察 59結論
68
参考文献
69
謝辞
75
報文
1
略語
βact: neo: β-actin promoter-neomycin-resistant gene Ala: alanine
Asp: asparate
ATP: adenosine triphosphate BCA: bicinchoninic acid
BCAA: branched-chain amino acids BCAT: branched-chain aminotransferase BCKA: branched-chain α-keto acids
BCKDC: branched-chain α-keto acid dehydrogenase complex BCKDH: branched-chain α-keto acid dehydrogenase BDK: branched-chain α-keto acid dehydrogenase kinase BDK-KO:global BDK knockout
BDK-mKO: muscle specific BDK knockout
BDP: branched-chain α-keto acid dehydrogenase phosphatase
BPB: bromophenol blue
BSA: bovine serum albumin CAR: carnitine
CE-TOFMS: capillary electrophoresis time-of-flight mass spectrometry CoA: coenzyme A
Cre : Cre recombinase CK: creatine kinase
DHAP: dihydroxyacetone phosphate
DTNB: 5, 5’-dithio-bis-[2-nitrobenzoic acid] DTT: dithiothreitol
EDTA: ethylenediaminetetraacetic acid EGTA: ethylene glycol tetraacetic acid
EuMMCR: european mouse mutant cell repository En2SA : splicing acceptor site
ES 細胞: embryonic stem cells FLP: flippase
FRT: flippase recognition target F1,6BP: fructose 1,6-bisphosphate βgal: β-galactsidas
2 G6P: glucose 6-phosphate
GTP: guanosine triphosphate
HEPES: 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid HMT: human metabolome technologies
IRES: internal ribosomal entry site IB-CAR: isobutyryl-carnitine IB-CoA: isobutyryl-coenzyme A IV-CoA: isovaleryl-coenzyme αKG: α-ketoglutarate KIC: α-ketoisocaproate KIV: α-ketoisovalerate KMV: α-keto-β-methylvalerate lacZ: β-galactsidase gene LDH: lactate dehydrogenase LPL: lipoprotein lipase αMB-CAR: α-methylbutyryl-carnitine βMB-CAR: β-methylbutyryl-carnitine αMB-CoA: α-methylbutyryl-CoA βMB-CoA: β-methylbutyryl-CoA MuRF1: muscle RING-finger protein-1
NAD: oxidized nicotinamide adenine dinucleotide NADH: reduced nicotinamide adenine dinucleotide Neo: neomycin-resistant gene
NFE: nitrogen free extract OA: oxaloacetate
pA: polyadenylation signal sequence PCA: perchloric acid
PCR: polymerase chain reaction PEP: phosphoenolpyruvate 2PG: 2-phosphoglycerate PMP: pyridoxamine 5'-phosphate PMSF: phenylmetylsulfonyl fluoride PVDF: polyvinylidene fluoride Pyr: pyruvate
SDS: sodium dodecyl sulfate
3 TCA:tricarboxylic acid
TNB: 2-nitro-5-mercapto-benzoic acid TPP: thiamine pyrophosphate
Tris: tris(hydroxymethyl)aminomethane
4
第 一 章
緒 言
1.1 分 岐 鎖 ア ミ ノ 酸
タ ン パ ク 質 を 構 成 す る 20 種 類 の ア ミ ノ 酸 の う ち 、側 鎖 に 分 枝 構 造 を 持 つ 3 つ の ア ミ ノ 酸 ( ロ イ シ ン 、 イ ソ ロ イ シ ン 、 バ リ ン ) は 分 岐 鎖 ア ミ ノ 酸
(branched-chain amino acids: BCAA) と 総 称 さ れ て い る ( Fig. 1.1) 。 BCAA は ヒ ト の 必 須 ア ミ ノ 酸 で あ る た め 、 ヒ ト は 一 般 的 に そ れ ら を 食 物 か ら 摂 取 し て い る 。BCAA は 食 物 タ ン パ ク 質 の 必 須 ア ミ ノ 酸 の 約 50%、 筋 タ ン パ ク 質 中 の 必 須 ア ミ ノ 酸 の 約 35%を 占 め て お り [1]、ヒ ト は 多 く の BCAA を 摂 取 し 、ま た 体 内 に 保 有 し て い る 。 体 内 に お け る ア ミ ノ 酸 の 多 く は タ ン パ ク 質 と し て 存 在 し て い る が 、 一 部 は 遊 離 型 と し て も 存 在 す る 。 こ の 遊 離 ア ミ ノ 酸 は 、 ま と め て ア ミ ノ 酸 プ ー ル と 呼 ば れ て お り 、 食 事 タ ン パ ク 質 の 消 化 吸 収 お よ び 体 タ ン パ ク 質 の 分 解 に よ っ て 供 給 さ れ る 。 遊 離 ア ミ ノ 酸 の 機 能 は タ ン パ ク 質 合 成 の 基 質 と し て 考 え ら れ て き た が 、 近 年 の 研 究 に よ っ て 様 々 な 生 理 機 能 を 有 す る こ と が 報 告 さ れ て い る 。 特 に 遊 離 型 と し て 存 在 し て い る BCAA は 、 タ ン パ ク 質 合 成 に お け る 基 質 と な る だ け で な く 、 タ ン パ ク 質 の 合 成 促 進 及 び 分 解 抑 制 の 働 き を 持 つ[2,3]。 他 に も 、 グ ル コ ー ス 代 謝 の 調 節 、 運 動 時 に お け る グ リ コ ー ゲ ン 消 費 抑 制 作 用 を 持 つ 可 能 性 が 報 告 さ れ て い る[4,5]。 一 方 、 タ ン パ ク 質 合 成 に 必 要 な 多 く の ア ミ ノ 酸 が 肝 臓 で 分 解 さ れ る の に 対 し て 、6 種 類 の ア ミ ノ 酸 が 筋 肉 に お い て 分 解 さ れ る 。 こ れ ら の ア ミ ノ 酸 の な か で 、 ア ラ ニ ン 、 ア ス パ ラ ギ ン 酸 と グ ル タ ミ ン 酸 は 、 ア ミ ノ 基 転 移 反 応 に よ っ て 、TCA サ イ ク ル の 前 駆 体 や 中 間 体 に 変 換 さ れ 、さ ら に そ の 逆 反 応 で TCA サ イ ク ル の 中 間 体 な ど が ア ミ ノ 基 を 受 け 取 っ て そ れ ら の ア ミ ノ 酸 を 合 成 す る 。
5
よ っ て 、 こ れ ら の ア ミ ノ 酸 は 筋 肉 か ら の ア ミ ノ 基 の 搬 出 に 重 要 な 役 割 を 担 っ て い る と 考 え ら れ る[6,7]。 一 方 、 BCAA は 、 か な り 多 く の 反 応 ス テ ッ プ を 経 て acetyl-CoA や succinyl-CoA へ 代 謝 さ れ 、 最 終 的 に TCA サ イ ク ル に 入 る 。 よ っ て 、BCAA は TCA サ イ ク ル の 基 質 で あ る acetyl-CoA や succinyl-CoA の 一 つ 供 給 源 で あ る た め 、 ヒ ト で も ラ ッ ト で も 運 動 に よ り そ の 分 解 が 促 進 さ れ [8,9]、 筋 肉 の エ ネ ル ギ ー 代 謝 に 影 響 す る と 考 え ら れ る 。 そ こ で 、 ま ず 体 内 で の BCAA 分 解 の 調 節 機 構 を 説 明 し 、 次 い で こ の 調 節 機 構 に 基 づ い て 、 BCAA 代 謝 亢 進 動 物 モ デ ル の 生 理 的 特 徴 に つ い て 解 説 す る 。
1.2 BCAA 代 謝 と そ の 調 節 機 構
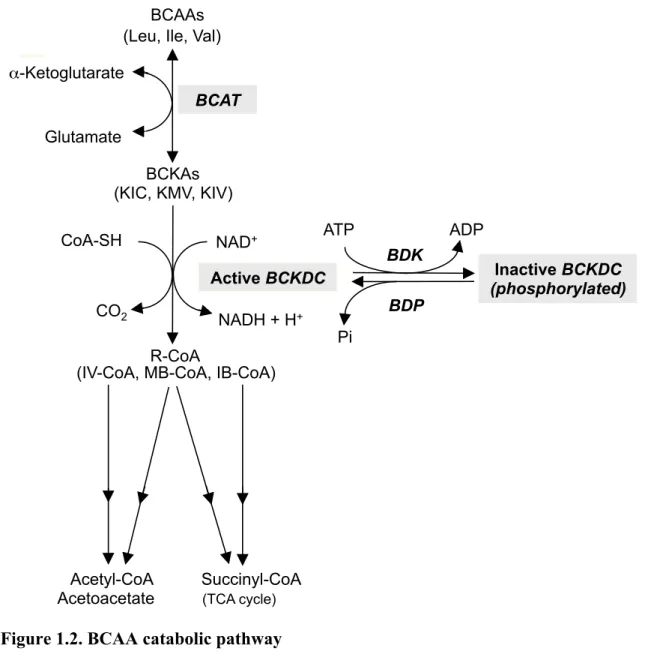
BCAA 分 解 経 路( Fig. 1.2)の ほ と ん ど は ミ ト コ ン ド リ ア 内 に 局 在 し て い る 。BCAA 分 解 の 最 初 の 2 つ の ス テ ッ プ は 3 つ の BCAA に 共 通 で あ る [10]。 第 1 ス テ ッ プ は 分 岐 鎖 ア ミ ノ 酸 ア ミ ノ 基 転 移 酵 素 ( branched-chain amino acid aminotransferase: BCAT) に よ っ て 触 媒 さ れ る 反 応 で あ り 、 BCAA が ア ミ ノ 基 転 移 を 受 け て 分 岐 鎖 α‐ ケ ト 酸 ( branched-chain α-keto acids: BCKA) と な る 。 BCAT に は 、 ミ ト コ ン ド リ ア 型 ( BCATm) と 細 胞 質 型 ( BCATc) の ア イ ソ ザ イ ム が 存 在 し て い る 。BCATm は 肝 臓 以 外 の ほ ぼ 全 身 の 臓 器 細 胞 に 発 現 し て い る の に 対 し て[11]、 BCATc は 脳 ・ 神 経 な ど の ご く 一 部 の 組 織 で の み 発 現 し て い る[12]。そ の た め 、脳・神 経 な ど の ご く 一 部 の 組 織 で は 、細 胞 質 で も BCAA の ア ミ ノ 基 転 移 反 応 が 可 能 と な っ て い る 。
第 2 ス テ ッ プ は 分 岐 鎖 α‐ ケ ト 酸 脱 水 素 酵 素 複 合 体 ( branched-chain α-keto acid dehydrogenase (BCKDH) complex: BCKDC) に よ っ て 触 媒 さ れ る 酸 化 的 脱 炭 酸 反 応 で あ り 、BCKA は そ れ ぞ れ に 対 応 す る CoA 化 合 物 へ と 変 換 さ れ る 。
6
第 1 ス テ ッ プ が 可 逆 的 で あ る の に 対 し て 第 2 ス テ ッ プ が 不 可 逆 的 で あ る こ と か ら 、後 者 が BCAA 代 謝 に お け る 律 速 段 階 と さ れ て い る [1]。そ の た め 、BCKDC の 調 節 は 遊 離 BCAA 濃 度 の 恒 常 性 に お い て 非 常 に 重 要 で あ る 。
BCKDC は 、E1 成 分( E1α + E1β: branched-chain α-keto acid decarboxylase)、 E2 成 分( dihydrolipoyl transacylase)、E3 成 分( dihydrolipoamide dehydrogenase) か ら 構 成 さ れ る 。E1 は BCKA の 脱 炭 酸 反 応 を 触 媒 す る 。 E2 は 活 性 部 位 に リ ポ 酸 を 持 ち 、 脱 炭 酸 を 受 け た 残 り の 炭 化 水 素 基 を CoA-SH と 反 応 さ せ て CoA 化 合 物 を 生 成 す る 。E3 は E2 の リ ポ 酸 を NAD+依 存 的 に 再 度 酸 化 さ せ 、 全 体 の 酵 素 活 性 を 維 持 し て い る 。こ れ ら の 成 分 の う ち 、E1 と E2 が 強 固 に 結 合 し て い る が 、E3 の 結 合 は 弱 い [13, 14]。 BCKDC は 特 異 的 キ ナ ー ゼ で あ る BCKDH キ ナ ー ゼ ( BCDKH kinase: BDK) に よ り リ ン 酸 化 さ れ る と 不 活 性 化 さ れ る[15-17]。 こ の と き E1α サ ブ ユ ニ ッ ト の Ser293 と Ser303 が リ ン 酸 化 さ れ る が 、 実 際 に は Ser293 の リ ン 酸 化 の み が そ の 不 活 性 化 と 対 応 し て い る[18, 19]。BDK に は BCKDC 結 合 型 と 遊 離 型 が 存 在 す る[20]。 BCKDC 結 合 型 BDK の み が 活 性 を 持 ち 、 結 合 型 BDK の 量 が BCKDH の 活 性 に 影 響 す る と さ れ て い る [20]。 し か し 、 結 合 型 BDK と 遊 離 型 BDK の 変 換 機 構 は わ か っ て お ら ず 、BDK と BCKDC の 相 互 作 用 に つ い て も 未 知 な 部 分 が 多 い 。BDK 活 性 の 阻 害 剤 は い く つ か 報 告 さ れ て い る [21]。 そ の な か で も ロ イ シ ン の 代 謝 中 間 体 で あ る α-ketoisocaproate( KIC) は BDK を 阻 害 し 、BCKDH を 活 性 化 す る [22, 23]。 BCKDC の E1 成 分 の 補 酵 素 で あ る TPP も ま た BDK 活 性 を 阻 害 す る [1]。 し か し 、 こ れ ら の 詳 し い 阻 害 様 式 は 明 ら か に な っ て い な い 。 一 方 、BCKDC は BCKDH ホ ス フ ァ タ ー ゼ( BCKDH phosphatase: BDP)に よ る 脱 リ ン 酸 化 で 活 性 化 さ れ る 。BDK の ク ロ ー ニ ン グ が 1992 年 に 完 了 し て い
7 た の に 対 し 、BDP の ク ロ ー ニ ン グ に 関 す る 報 告 は 2007 年 と 最 近 で あ る た め 、 BDP の 特 性 に 関 し て は ま だ 不 明 点 が 多 い 。現 在 ま で の 研 究 に よ っ て 、BDP は ミ ト コ ン ド リ ア 内 のMn2 +依 存 型protein phosphatase 2C( PP2Cm)で あ る [24, 25]。 し か し 、 そ の BCKDC に 対 す る 比 活 性 が か な り 低 い こ と [25]、 精 製 さ れ た BCKDC に BDP の サ ブ ユ ニ ッ ト が 検 出 さ れ な い こ と か ら [26]、BCKDC に 対 す る 結 合 は BDK よ り も 弱 い と さ れ る 。 よ っ て 、 BCKDC の 活 性 調 節 に は BDK が よ り 強 く 影 響 す る と 考 え ら れ る 。 骨 格 筋 に お い て 、 安 静 状 態 で は BCKDH が ほ と ん ど リ ン 酸 化 さ れ て お り 、 そ の 活 性 は 極 め て 低 い が 、 こ の BCAA 分 解 の 抑 制 は 筋 タ ン パ ク 質 合 成 の 基 質 と し て の BCAA を 確 保 す る た め と 考 え ら れ て い る [11,27]。 一 方 、 ラ ッ ト に ト レ ッ ド ミ ル 走 行 運 動 を 2 時 間 さ せ る こ と に よ り 骨 格 筋 の BCKDC の 約 80%が 脱 リ ン 酸 化 さ れ 、BCAA 代 謝 が 亢 進 す る と 報 告 さ れ て い る [8]。 ま た 、 ヒ ト に お い て も 同 様 な 結 果 が 認 め ら れ る[9]。 タ ン パ ク 質 が 不 足 す る 場 合 で は 、 ラ ッ ト の 肝 臓 の BCKDC の 活 性 は 低 い が 、 運 動 負 荷 に よ り 肝 臓 BCKDC の 活 性 は 上 昇 し た[28]。こ れ ら の 結 果 か ら 、運 動 が BCKDC の 活 性 に 強 く 作 用 す る と 考 え ら れ る 。
1.3 筋 特 異 的 BCAA 代 謝 亢 進 動 物 モ デ ル
1.3.1 背 景 BCAA の 生 理 機 能 を 解 明 す る た め に は 、 BCAA が 不 足 し た 実 験 動 物 の 作 製 が 有 用 な 実 験 方 法 で あ る 。 し か し 、 食 餌 中 の BCAA を 欠 乏 さ せ て も 体 タ ン パ ク 質 に は 豊 富 に BCAA が 含 ま れ る た め 、組 織 中 の BCAA 濃 度 を 著 し く 低 下 す る こ と は 困 難 で あ る 。 ま た 、 上 述 し た 通 り 、BDK が BCKDC の 活 性 を 強 く 制 御 し て い8 る の で 、Joshi ら は 全 身 組 織 の BDK ノ ッ ク ア ウ ト マ ウ ス を 作 製 し た と こ ろ 、 組 織 及 び 血 液 中 の BCAA 濃 度 が 減 少 し て 、 そ の マ ウ ス で は 脳 機 能 の 異 常 が 認 め ら れ た[29]。 当 研 究 室 で も 、 全 身 組 織 BDK-ノ ッ ク ア ウ ト ( BDK-KO) マ ウ ス を 作 製 し 、BCKDC 活 性 を 上 昇 す る こ と に よ り BCAA 分 解 を 促 進 し た と こ ろ 、 こ の マ ウ ス で も 組 織 及 び 血 中 の BCAA 濃 度 が 減 少 し 、 成 長 不 全 や 尾 懸 垂 し た 際 に 後 肢 を 抱 え 込 む 脳 機 能 異 常 の 特 徴 を 示 し た[30]。 さ ら に 、 こ の マ ウ ス で は 、 糖 負 荷 試 験 に よ る耐糖能試験において耐糖能が低下することが明らかになった(未発表デ ータ)。 そ の た め 、BDK-KO マ ウ ス は 運 動 負 荷 実 験 に は 適 し て い な い と 考 え ら れ た 。 そ こ で 、 本 研 究 で は 筋 組 織 特 異 的 に BDK を 欠 損 し た マ ウ ス ( BDK-mKO マ ウ ス ) を 作 製 し ( 第 一 章 、[30]の 所 見 を 基 に し た 内 容 ) 、 慢 性 的 に 筋 組 織 で の み BCAA 分 解 を 亢 進 さ せ BCAA を 減 少 さ せ た 状 態 に お け る 運 動 負 荷 の 影 響 に つ い て 検 討 し 、 こ の 内 容 を 本 論 文 の 第 二 章 ( 文 献[31]の 所 見 を 基 礎 と し た 研 究 ) に 記 載 し た 。 さ ら に 、BDK-mKO マ ウ ス に BCAA を 慢 性 的 に 摂 取 さ せ 、 BCAA 不 足 を 改 善 し た BDK-mKO マ ウ ス に 対 す る 運 動 負 荷 の 影 響 に つ い て も 検 討 し 、 そ の 内 容 を 第 三 章 ( 文 献[32]の 所 見 を 基 礎 と し た 研 究 ) に 記 載 し た 。 以 下 の BDK-mKO マ ウ ス に 関 す る 記 載 は 、文 献 [30]の 内 容 に 基 づ い た も の で あ る 。 1.3.2 BDK-mKO マ ウ ス の 作 製 方 法 [30] マ ウ ス の 筋 肉( 骨 格 筋 、心 筋 )で 特 異 的 に BDK を 欠 損 さ せ る た め に 、Cre–loxP シ ス テ ム を 利 用 し た ノ ッ ク ア ウ ト マ ウ ス 作 製 方 法 を 用 い た (Fig.1.3) 。 ま ず 、 BDK の キ ナ ー ゼ 活 性 領 域 で あ る エ ク ソ ン 9 か ら 12 を loxP 配 列 で 挟 み 込 ん だ タ ー ゲ ッ テ ィ ン グ ベ ク タ ー を EuMMCR よ り 入 手 し た 。こ の タ ー ゲ ッ テ ィ ン グ ベ ク
9
タ ー に は 、Neomycin 耐 性 遺 伝 子 ( Neo) 及 び β-galactsidase 遺 伝 子 ( lacZ) が 挿 入 さ れ て い た 。Neo は 薬 剤 耐 性 を 利 用 し た 細 胞 を 選 択 す る た め に 、 lacZ は 遺 伝 子 発 現 を 解 析 す る た め に 組 み 込 ま れ た 。 そ れ ら の 遺 伝 子 は 、 酵 母 の 組 み 換 え 酵 素 で あ る フ リ ッ パ ー ゼ(FLP)の 認 識 配 列 で あ る flippase recognition target( FRT) で 挟 み 込 み こ ん だ 。 こ の ベ ク タ ー を PCR に よ っ て 確 認 し た 後 、 C57BL/6N マ ウ ス 由 来 の ES 細 胞 へ の ト ラ ン ス フ ェ ク シ ョ ン 、薬 剤 耐 性 ク ロ ー ン の 単 離 を 行 い 、 ク ロ ー ン ES 細 胞 を 得 た 。こ れ ら の 細 胞 か ら ゲ ノ ム DNA を 抽 出 し 、PCR に よ り 正 し く 相 同 組 換 え が 起 こ っ て い る か を 確 認 し た 。 得 ら れ た ク ロ ー ン ES 細 胞 に お け る 核 型 解 析 を 実 施 し 、80%以 上 で 正 常 な 核 型 を 確 認 で き た ク ロ ー ン を 、ICR 系 統 マ ウ ス 由 来 の 4 細 胞 期 胚 に イ ン ジ ェ ク シ ョ ン し 、キ メ ラ マ ウ ス を 作 製 し た 。 こ の キ メ ラ マ ウ ス と C57BL/6 系 統 マ ウ ス を 交 配 し て 繁 殖 し た と こ ろ 、PCR に よ る 相 同 組 換 え の 有 無 を 確 認 す る こ と で 片 方 の BDK の 対 立 遺 伝 子 が loxP で 挟 み 込 ま れ た 組 換 え マ ウ ス(BDKZ n f l o x / +マ ウ ス )を 得 る こ と が で き た 。こ の 段 階 に お い て は 、ま だ BDK ゲ ノ ム DNA 領 域 に 、ES 細 胞 を 単 離 す る た め の 薬 剤 耐 性 遺 伝 子 の Neo が 存 在 し て い る た め 、Neo を 取 り 除 く 必 要 が あ る 。そ の た め 、BDKZ n f l o x / + マ ウ ス と 体 の 全 組 織 で FLP を 発 現 す る ト ラ ン ス ジ ェ ニ ッ ク マ ウ ス を 交 配 さ せ る こ と に よ っ て 、FRT で 挟 ま れ た Neo や lacZ を 含 む 遺 伝 子 領 域 を 取 り 除 い た BDKf l o x / f l o xマ ウ ス を 作 製 し た 。こ の マ ウ ス(BDKf l o x / f l o xマ ウ ス )を 、筋 肉 特 異 的 に 発 現 す る ク レ ア チ ン キ ナ ー ゼ(CK)の プ ロ モ ー タ ー を 持 っ た Cre 遺 伝 子 を 導 入 し た CK-Cre ト ラ ン ス ジ ェ ニ ッ ク マ ウ ス と 交 配 し 、 Cre 転 写 遺 伝 子 を 持 つ BDKf l o x / + ; Cre–マ ウ ス 及 び 、 BDKf l o x / + ; Cre+マ ウ ス を 作 製 し た 。 作 製 し た こ れ ら の マ ウ ス を 交 配 さ せ 、BDKf l o x / f l o x ; Cre+マ ウ ス 、 す な わ ち 筋 肉 特 異 的 BDK 欠 損 マ ウ ス (BDK-mKO マ ウ ス ) を 作 製 し た 。 1.3.3 BDK-mKO マ ウ ス の 生 理 特 徴 [30]
10 ( 1 ) 体 重 と 組 織 重 量
3 週 齢 か ら 10 週 齢 ま で の BDK-mKO マ ウ ス( BDKf l o x / f l o x ; Cre+マ ウ ス )の 体 重 は 、Control マ ウ ス ( BDKf l o x / f l o x ; Cre–マ ウ ス ) の そ れ と ほ ぼ 同 じ で あ っ た (Fig. 1.4) 。 10 週 齢 で 解 剖 し た マ ウ ス の 各 組 織 の 重 量 も 、 両 マ ウ ス で 差 は な か っ た (Table 1.1) 。
( 2 ) 各 組 織 で の BDK タ ン パ ク 質 の 発 現
BDK-mKO マ ウ ス( BDKf l o x / f l o x ; Cre+マ ウ ス )と Control マ ウ ス( BDKf l o x / f l o x ; Cre– マ ウ ス ) の 肝 臓 、 骨 格 筋 、 腎 臓 、 心 臓 、 脳 を 用 い て 、Western blot 法 に よ り 、 BDK の 有 無 を 分 析 し た 。 結 果 は 、 Fig. 1.5 で 示 す よ う に 、 BDK タ ン パ ク 質 発 現 量 は 、Control マ ウ ス の 骨 格 筋 と 心 筋 で 明 確 に 検 出 さ れ た の に 対 し て 、BDK-mKO マ ウ ス で 著 し く 低 下 し て い た こ と が 観 察 さ れ た 。 よ っ て 、BDK-mKO マ ウ ス が 筋 肉 特 異 的 に BDK を 欠 損 す る こ と が 確 認 さ れ た 。 ( 3 ) 血 漿 と 筋 肉 で の ア ミ ノ 酸 分 析 血 漿 ロ イ シ ン 、イ ソ ロ イ シ ン 、バ リ ン 濃 度 が BDK-mKO マ ウ ス で 、Control マ ウ ス と 比 較 し て 、 有 意 に 低 下 し た 。BCAA 以 外 の ア ミ ノ 酸 濃 度 は 、 BDK-mKO マ ウ ス と Control マ ウ ス で 変 わ ら な か っ た ( Table 1.2) 。 心 筋 で 測 定 さ れ た ロ イ シ ン 、 イ ソ ロ イ シ ン 、 バ リ ン 濃 度 は 、BDK-mKO マ ウ ス で Control マ ウ ス の 34%、48%と 45%ま で 低 下 し た 。ア ラ ニ ン 以 外 の ほ か の ア ミ ノ 酸 濃 度 は 両 マ ウ ス 間 で 差 が 見 ら れ な か っ た 。心 筋 の ア ラ ニ ン 濃 度 が Control マ ウ ス と 比 較 し て BDK-mKO マ ウ ス で 有 意 に 上 昇 し た ( Table 1.3) 。 骨 格 筋 の ロ イ シ ン 、 イ ソ ロ イ シ ン 、 バ リ ン の 濃 度 が 、BDK-mKO マ ウ ス で 、 Control マ ウ ス と 比 較 し て 50% 以 下 に 低 下 し た( ロ イ シ ン:39%、イ ソ ロ イ シ ン: 21%、 バ リ ン : 50%) ( Table 1.4) 。 さ ら に 、 BDK を ノ ッ ク ア ウ ト す る こ と で
11 BCAA 代 謝 亢 進 に よ っ て 、ア ミ ノ 基 転 移 反 応 に 関 わ る ア ミ ノ 酸 で あ る ア ラ ニ ン 、 ア ス パ ラ ギ ン 酸 、グ ル タ ミ ン 酸 、グ ル タ ミ ン の 濃 度 がBDK-mKO マ ウ ス で Control マ ウ ス と 比 較 し て 有 意 に 上 昇 し た 。 そ れ ら 以 外 の ア ミ ノ 酸 濃 度 が 、BDK-mKO マ ウ ス と Control マ ウ ス で 変 わ ら な か っ た ( Table 1.4) 。 こ れ ら の 結 果 よ り 、 BDK-mKO マ ウ ス で BCAA 代 謝 が 亢 進 し た こ と が 推 察 さ れ る 。
12
Figure 1.1. L-Branched-chain amino acids
13
Figure 1.2. BCAA catabolic pathway
BCAT:
branched-chain
aminotransferase
MB-CoA:
α-methylbutyryl-CoA
BCKA:
branched-chain α-keto acids
IV-CoA:
isovaleryl-CoA
BCKDC:
BCKA dehydrogenase
IB-CoA:
isobutyryl-CoA
BDK:
BCKDH kinase
BDP:
BCKDH phosphatase
KIC:
α-ketoisocaproate
KMV:
α-keto-β-methylvalerate
KIV:
α-ketoisovalerate
BCAAs (Leu, Ile, Val)BCKAs (KIC, KMV, KIV) BCAT α-Ketoglutarate Glutamate R-CoA
(IV-CoA, MB-CoA, IB-CoA)
CoA-SH NAD+ NADH + H+ CO2 Active BCKDC BDK BDP ATP ADP Pi Inactive BCKDC (phosphorylated) Acetyl-CoA Succinyl-CoA
14
Figure 1.3. Conditional BDK gene knockout using the Cre–loxP system [30]
En2SA:
Splicing acceptor site
FLP:
Flippase
IRES:
Internal ribosomal entry site
lacZ:
β-Galactsidase gene
Neo:
Neomycin-resistant gene
βgal:
ß-Galactsidase gene
βact:neo:
β-Actin promoter-neomycin-resistant gene
pA:
Polyadenylation signal sequence
15
Figure 1.4. Growth curves from 8 weeks old to 13 weeks old of control and
BDK-mKO mice [30]
16
Figure 1.5. Western blotting of BDK in control and BDK-mKO mice [30]
Liver
Skeletal
muscle
Kidney
Heart
Brain
BDK-mKO
Control
BDK
17
Table 1.1. Tissue weights of the control and BDK-mKO mice [30]
Tissue Control mBDK-KO (g/100 g BW) Liver 4.68 ± 0.11 4.59 ± 0.11 Brain 1.61 ± 0.03 1.61 ± 0.05 Kidney 1.35 ± 0.06 1.34 ± 0.04 Skeletal muscle 1.08 ± 0.02 1.07 ± 0.02 Adipose 0.90 ± 0.06 1.01 ± 0.06 Testis 0.68 ± 0.03 0.66 ± 0.02 Pancreas 0.57 ± 0.04 0.67 ± 0.06 Heart 0.50 ± 0.02 0.50 ± 0.05 Spleen 0.37 ± 0.05 0.32 ± 0.02
Values are means ± SEM (Control: n=12, BDK-mKO: n=10). Skeletal muscle is a mixture of soleus, gastrocnemius, and plantaris muscles of the one leg. Epididymal adipose tissues from both sides of the body were collected from the mice and combined.
18
Table 1.2. Plasma concentrations of amino acids in control and BDK-mKO mice
[30]
Amino acid Control BDK-mKO %
(µM) Isoleucine 76 ± 7 40 ± 5** 52 Leucine 108 ± 9 67 ± 5** 62 Valine 168 ± 10 113 ± 5** 68 Alanine 348 ± 16 394 ± 20 113 Arginine 73 ± 7 69 ± 4 95 Aspartate Citrulline 8 ± 1 114 ± 7 7 ± 2 109 ± 9 97 95 Glutamate 20 ± 3 23 ± 3 113 Glutamine 502 ± 22 534 ± 20 106 Glycine 319 ± 15 335 ± 11 105 Histidine 38 ± 6 46 ± 8 121 Lysine 194 ± 15 200 ± 10 103 Methionine 115 ± 7 112 ± 6 98 Ornithine 30 ± 3 33 ± 2 110 Phenylalanine 60 ± 3 58 ± 3 97 Serine 109 ± 5 110 ± 7 101 Taurine 441 ± 14 493 ± 36 112 Threonine 113 ± 6 111 ± 5 98 Tyrosine 78 ± 5 84 ± 7 108
19
Table 1.3. Concentrations of amino acids in heart muscle of control and
BDK-mKO mice [30]
Amino acid Control BDK-mKO % (nmol/g tissue) Isoleucine 70 ± 3 32 ± 2** 45 Leucine 95 ± 5 46 ± 2** 48 Valine 132 ± 7 45 ± 4** 34 Alanine 2776 ± 127 3235 ± 134* 117 Arginine 180 ± 3 178 ± 6 99 Asparagine 229 ± 22 231 ± 20 101 Aspartate 1756 ± 167 1923 ± 286 110 Citrulline 377 ± 23 450 ± 40 119 Glutamate 3484 ± 351 3106 ± 362 89 Glutamine 5873 ± 168 6510 ± 292 111 Histidine 248 ± 45 298 ± 82 120 Methionine 193 ± 10 178 ± 10 92 Serine 287 ± 19 261 ± 14 91 Taurine 38832 ± 1692 34506 ± 855 89 Threonine 272 ± 9 258 ± 8 95 Tyrosine 108 ± 19 107 ± 8 99
20
Table 1.4. Concentrations of amino acids in skeletal muscle of control and
BDK-mKO mice [30]
Amino acid Control BDK-mKO %
(nmol/g tissue) Isoleucine Leucine Valine Alanine 64 ± 5 73 ± 4 158 ± 9 2011 ± 107 13 ± 5** 28 ± 2** 79 ± 5** 2699 ± 249* 21 39 50 134 Arginine 134 ± 21 137 ± 16 102 Aspartate 801 ± 29 937 ± 39* 117 Carnosine 2572 ± 240 2101 ± 90 82 Citrulline 210 ± 22 280 ± 44 133 Glutamate 763 ± 35 910 ± 60** 119 Glutamine 1671 ± 76 2141 ± 184* 128 Glycine 3185 ± 150 3167 ± 183 99 Histidine 68 ± 16 91 ± 20 133 Lysine 445 ± 50 429 ± 41 96 Ornithine 21 ± 3 22 ± 4 105 Phenylalanine 52 ± 3 40 ± 7 77 Serine 311 ± 17 336 ± 20 108 Taurine 35363 ± 1040 33053 ± 2034 93 Threonine 236 ± 12 254 ± 15 108 Tyrosine 60 ± 8 60 ± 16 100
Skeletal muscle is a mixture of soleus, gastrocnemius, and plantaris muscles. Values are means ± SEM (Control: n=12, BDK-mKO: n=10). *P < 0.05、**P < 0.01
21
第二章 筋特異的
BCAA 分解亢進が運動持久力と運動中の代謝
変動に与える影響
2.1 序論
運 動 に よ り 、 ヒ ト の エ ネ ル ギ ー 代 謝 は 著 し く 亢 進 す る 。 こ の エ ネ ル ギ ー 代 謝 は 酸 素 消 費 量 に 反 映 さ れ る[33]。 健 康 な 成 人 で は 、 運 動 時 の 酸 素 消 費 量 は 安 静 時 ( お よ そ 250 ml/分 ) よ り 約 10 倍 増 加 す る 。 さ ら に 、 よ く ト レ ー ニ ン グ さ れ た ア ス リ ー ト で は 、運 動 時 の 酸 素 消 費 量 が 20 倍 に も 増 加 す る こ と が 知 ら れ て い る 。 エ ネ ル ギ ー 代 謝 の 基 質 と し て は 、 糖 質 と 脂 質 が 中 心 的 に 利 用 さ れ る が 、 亢 進 し た エ ネ ル ギ ー 代 謝 で は そ の 生 理 状 態 に 応 じ て 全 エ ネ ル ギ ー の 3~ 18%が タ ン パ ク 質 や ア ミ ノ 酸 か ら 供 給 さ れ る よ う で あ る [34]。し た が っ て , ア ミ ノ 酸 も 重 要 な エ ネ ル ギ ー 源 で あ る 。 特 に 筋 肉 は 最 も 重 要 な 運 動 器 官 で あ る と 同 時 に 、 ア ミ ノ 酸 の 貯 蔵 庫 と し て 、 運 動 す る 時 に BCAA が 効 率 的 に 利 用 さ れ る と 、 糖 質 の 利 用 が 抑 制 さ れ る こ と に よ っ て 乳 酸 の 産 生 も 抑 制 さ れ 、 運 動 持 久 力 の 維 持 に つ な が る 可 能 性 が あ る[35]。 さ ら に 、 運 動 は 骨 格 筋 で の BCKDC の 脱 リ ン 酸 化 を 促 進 し て 活 性 化 し 、BCAA の 酸 化 分 解 を 高 め る こ と が わ か っ て い る[7,28,36]。 ま た 、 ト レ ー ニ ン グ は 、 ミ ト コ ン ド リ ア を 増 加 す る こ と に よ っ て 、BCKDC の 酵 素 量 も 増 加 し て 、BCAA を 酸 化 す る 能 力 を 高 め る こ と も 明 ら か に さ れ て い る[37]。 一 方 、BCAA 代 謝 系 の 第 一 ス テ ッ プ の 酵 素 で あ る BCAA ア ミ ノ 基 転 移 酵 素 を 欠 損 し た マ ウ ス で は , 運 動 持 久 力 が 著 し く 低 下 す る こ と が 報 告 さ れ て い る [38]。 こ の マ ウ ス で は 、 BCAA を 分 解 で き な い た め に 正 常 マ ウ ス と 比 べ て 筋 中 の BCAA 濃 度 は 10 倍 以 上 に 上 昇 す る が 、 逆 に ア ラ ニ ン と グ ル タ ミ ン 濃 度 は 50%以 下 に 低 下 し た 。 よ っ て 、 運 動 中 の 効 率 的 な BCAA 代 謝 が 運 動 持 久 力22 な ど の パ フ ォ ー マ ン ス を 発 揮 す る た め に 重 要 と 考 え ら れ る 。 し か し 、 こ の 報 告 は BCAA 量 が 増 加 し た と き の 報 告 で あ り 、 BCAA 量 が 慢 性 的 に 減 少 し た 状 態 に お け る 運 動 時 の 影 響 に つ い て は 報 告 が な い 。 そ の た め 、 本 研 究 で は 筋 特 異 的 に BDK を 欠 損 し た マ ウ ス を 使 用 し 、 慢 性 的 BCAA 減 少 状 態 の 筋 機 能 に 対 す る 運 動 負 荷 の 影 響 に つ い て 調 べ た[31]。
2.2 材 料 及 び 方 法
2.2.1 実 験 動 物 と そ の 飼 育 法 第 一 章 で 述 べ た 方 法 を 用 い て 、BDKf l o x / f l o x ; Cre+マ ウ ス を 作 製 し て 、 筋 特 異 的 BCAA 分 解 亢 進 動 物 モ デ ル と し た 。一 方 、Control マ ウ ス と し て 、BDKf l o x / f l o x ; Cre–マ ウ ス も 作 製 し た 。8 週 齢 の 雄 性 BDK-mKO マ ウ ス ( BDKf l o x / f l o x ; Cre+) と Control マ ウ ス (BDKf l o x / f l o x ; Cre–) を 個 別 の ケ ー ジ で 飼 育 し 、 食 餌 、 水 と も に 自 由 摂 取 と し た 。 飼 育 室 は 室 温 が 23±1℃ 、 明 暗 サ イ ク ル は 12 時 間 ( 明 期 : 午 前 8 時 か ら 午 後 8 時 、 暗 期 : 午 後 8 時 か ら 午 前 8 時 ) と な る よ う に 設 定 し た 。 食 餌 は D12450B( Research Diets Inc., New Brunswick, NJ, USA) を 用 い た 。 そ れ ぞ れ の 餌 の 組 成 を Table 2.1 に 示 し た 。 2.2.2 実 験 プ ロ ト コ ー ル ( 実 験 1 ) 運 動 持 久 力 の 測 定 マウスの走運動持久力を、2 週間の運動トレーニングの前後で測定した。上記の方法で 飼育した運動トレーニングしていない12 週齢のマウスを用い、トレッドミル(Natsume, Tokyo, Japan)を用いて走運動持久力を測定した(測定法は以下の 2.2.4 を参照)。その 後2 日間安静にして、2 週間のトレーニングを負荷した(トレーニング方法は以下の 2.2.3
23 を参照)。トレーニング負荷最終日から2 日間安静にしたのち、同様な方法で再度運動 持久力の測定を行った。 ( 実 験 2 ) メ タ ボ ロ ー ム 解 析 の た め の マ ウ ス の 調 製 と サ ン プ ル の 採 取 マウスは12 週齢からトレッドミルを用いて 2 週間の走運動トレーニングを負荷した。 トレーニングの最終日から2 日間休ませた後、実験最終日に、8 時間絶食の条件の下で、 32 分間の急性運動(持久能力の測定方法 2.2.4 と同様)をさせた。運動直後に、頸部脱臼 により屠殺した。屠殺後、速やかに骨格筋、下大静脈から血液、心臓、肝臓、精巣上体 脂肪組織、腎臓、脾臓を採取した。採取した組織はフリーズクランプによる急速冷凍の 後、分析まで-80℃で保存した。骨格筋サンプルでは、右後肢からはヒラメ筋と足底筋+ 腓腹筋採を分けて採取し(ヒラメ筋は酵素活性測定用;足底筋+腓腹筋はメタボローム 解析用)、左後肢からはこれらをまとめて採取した(グリコーゲン定量用)。採取した 血液は、800 µl をヘパリン(Mochida Pharma, Tokyo)入り血漿サンプル用チューブに入れ、 200 µl をヘパリンなし血清サンプル用チューブに入れた。それらを 15,000×g、14℃で 15 分間の遠心分離を2 回行い、それぞれの上清を血漿サンプルと血清サンプルとした。 2.2.3 ト レ ー ニ ン グ 方 法 トレッドミルによる走運動トレーニングを2 週間実施した。1 週目のトレーニング内容 は、勾配10%、走行速度 15 m/分、60 分/日、5 日/週とした。それに次ぐ 2 週目のトレー ニング内容は、勾配10%、走行速度 18 m/分、60 分/日、5 日/週とした。 2.2.4 走 運 動 持 久 力 の 測 定 方 法 トレッドミルを用いて走運動持久力を測定した。勾配10%で、速度 15 m/分から開始し、 4 分毎に 1 m/分ずつ速度を上げ、マウスが疲労困憊になるまでの走行時間を測定した。5 秒間走行不可能な状態が5 回続いた時を疲労困憊と定義した。走行時間を走行距離に変 換してマウスの走運動持久力を評価した。
24 2.2.5 メ タ ボ ロ ー ム 解 析 生体内には核酸やタンパク質のほか、糖、有機酸、アミノ酸など数多くの物質が存在 し、これらの濃度は代謝活動により変動する。今日では細胞の働きを包括的に理解する ために、DNA 配列の網羅的解析(ゲノム解析)やタンパク質の網羅的解析(プロテオミ クス解析)に加え、代謝物質の網羅的解析(メタボローム解析)も行われるようになっ た。本研究ではメタボロームの網羅的解析をヒューマン・メタボローム・テクノロジー ズ株式会社(HMT)に依頼して実施した。 [測定サンプル調製] 粉末状のマウスの骨格筋に50%(v/v)アセトニトリル水溶液 (内部標準物質濃度 ; 20 µM(カチオン測定用)、5 µM(アニオン測定用))を加え冷却下にて破砕した。その後 遠心分離し限外濾過し、ろ液を乾固させ、50 µl の MlilliQ 水に溶解して測定に用いた。 2.2.6 血 漿 ア ミ ノ 酸 濃 度 測 定 [サンプル調製] 血漿に3%スルホサリチル酸溶液を等量加え、5 分ごとにボルテックスで撹拌して、 15 分後に遠心(12,000×g、15 分、4℃)して上清を得た。その上清 50 µl にサンプル希 釈液50 µl 加えた後、濾過したものを分析サンプルとした。 [測定方法] 試料の所定量を、陽イオン交換樹脂を充填したカラムに導入し、種々の緩衝液を流す ことにより、各々のアミノ酸成分に分離され、カラムより溶出した。溶出液にニンヒド リン試薬を加え、加熱することによりアミノ酸と反応した。それを測定し、検出された 信号をデータ処理部に送り、アミノ酸の定性と定量を行った。 [器具]
25 [試薬の調製] 緩衝液組成表 1st 2nd 3rd 4th 5th 再生液 Li 濃度(N) 0.105 0.25 0.8 1.54 1.54 0.4 pH 2.98 3.3 3.64 2.92 3.8 クエン酸リチウム 6.9g 9.9g 35.3g 69.4g 69.4g 塩化リチウム 1.3g 6.2g 17.3g 33.9g 33.9g クエン酸 8.8g 2.1g 塩酸 4.0ml 6.0ml エタノール 40.0ml Bridge-35(20%) 3.1ml 3.1ml 3.1ml 3.1ml 3.1ml 3.1ml チオジグリコール 2.5ml 2.5ml 2.5ml 2.5ml n-カプリル酸 0.1ml 0.1ml 0.1ml 0.1ml 水酸化リチウム 16.8g 全量 1000ml 1000ml 1000ml 1000ml 1000ml 1000ml 標準試料: 下記の(1)~(5)の試料を各 50 µl 秤量し、サンプル希釈液で全量を 1.25 ml にした。 (1) 和光純薬(株)製標準アミノ酸混合液 ANⅡ型 (2) 和光純薬(株)製標準アミノ酸混合液 B 型 (3) アスパラギン酸溶液(1.25 µmol/ml) (4) グルタミン溶液(2.5 µmol/ml) (5) トリプトファン溶液(2.5 µmol/ml) 標準試料作製後、分注し使用まで-20℃で保存した。 サンプル希釈液:1st 緩衝液に濃塩酸を加え、pH 2.2 に調整した。 3%スルホサリチル酸溶液:10 ml の蒸留水に 300 mg のスルホサリチル酸を溶解し作製 した。 2.2.7 タ ン パ ク 質 濃 度 測 定 [サンプル調製]
26 採取した後肢筋(腓腹筋、ヒラメ筋、足底筋)を、液体窒素によって冷却したステンレス 製乳鉢と乳棒を用いて粉末にした。粉末にした心筋を約30 mg 秤量し、Extraction buffer を400 µl 加えた後、氷上でホモジナイズした。また、粉末にしていないヒラメ筋に Extraction buffer を 200 µl 加え、氷上でテフロンホモジナイザーを用いてホモジナイズし た。その後遠心分離(15,000g、15 分、4℃)し、上清をタンパク質抽出試料として用い た。これらの試料のタンパク質濃度を測定した後に、ウェスタンブロット、クエン酸酵 素合成酵素活性測定、シトクロムc オキシダーゼ活性測定に用いた。 [測定方法]
タンパク質の定量は、BCA 法に従った [41,42]。BCA Protein Assay Kit を用いて行い、 562 nm の吸光波長を分光光度計で測定した。検量線の作製には、標準タンパク質として ウシ血清アルブミン(bovine serum albumin: BSA)を用いた。
[器具] テフロンホモジナイザー(HOM; アズワン株式会社、大阪) SPECTRA MAX 190(モレキュラーデバイスジャパン株式会社、東京) 卓上微量冷却遠心機 CT15RE(日立工機株式会社、東京) UV1700(島津製作所、京都) [キット、試薬の調製]
BCA Protein Assay Kit(サーモフィッシャーサイエンティフィック株式会社、神奈川) BSA(Bovine serum albumin)(SIGMA Chemical Company, St. Louis, USA)
(1)
Extraction buffer:
50 mM HEPES(pH 7.4 with KOH)、1%(w/v)Triton X-100、
1 mM EDTA、1 mM EGTA、2 mM Na
3VO
4、
100 mM NaF、
50 mM Na
2P
2O
7、
1 mM PMSF、5 µg/ml leupeptin、5 µg/ml
aprotinin、0.1 mg/ml trypsin inhibitor
27 [サンプル調製] タンパク質抽出サンプルとExtraction buffer を用いて、5.0 µg/µl のタンパク質抽出液を 200 µl 調製した。その後、2×Treatment buffer を 200 µl 加え(2.5 µg/µl のタンパク質)、 95℃で 5 分間加熱することでウェスタンブロット用サンプルを調製した。 [測定方法] ポリアクリルアミドゲルの各スロットにウェスタンブロット用サンプルをアプライし (各組織のタンパク量:心臓、腎臓、膵臓が25 µg、脳が 30 µg、骨格筋、脾臓、肝臓、 脾臓、精巣が50 µg)、SDS-PAGE (poly-acrylamide gel electrophoresis)を Stacking gel まで20 mA、separating gel から 30 mA で泳動した。
Polyvinylidene difluoride(PVDF)膜をメタノールに短時間浸たした後、Transfer buffer に浸した。ろ紙3 枚を Transfer buffer に浸した後、3 枚重ねてセミドライブロッティング 装置の上に置き、その上にPVDF 膜、次いで Transfer buffer に軽く浸たしたゲルを重ねた。 最後にTransfer buffer に浸したろ紙 3 枚を置いて気泡を抜き、8V の電圧で 1 時間、転写 した。転写後のPVDF 膜を Blocking buffer(3%スキムミルク(w/v))に浸し、室温で 2 時間振動した。
その後、PVDF 膜を、一次抗体 BDK-pAB 、BDKDC を 1000 倍希釈した Blocking buffer 溶液中で、4℃で 1 晩振動した。翌日、PVDF 膜を蒸留水で洗浄後、TBST で 20 分間(5 分間×4)洗浄した。洗浄した PVDF 膜を、二次抗体(anti rabbit)を 2000 倍希釈した Blocking buffer 中で 2 時間振動した。PVDF 膜を蒸留水で洗い流した後、TBST で 10 分間(5 分間 ×2 回)、次いで TBS で 15 分間(5 分間×3 回)洗浄した。
目標タンパク質のバンドの検出はECL 検出キットを用いて行った。PVDF 膜上に ECL 液を均等に散布し、撮影を行った。
ELC 検出を行った PVDF 膜を Stripping buffer 100 ml に浸し、容器をウォーターバスで 68 ℃で 45 分間振とうした。PVDF 膜を蒸留水で洗浄後、TBST で 20 分間(5 分間×4 回)
28
洗浄した。その後は、一次抗体E2-pAB、二次抗体 anti rabbit を用いて、上記の抗原抗体 反応、バンドの検出と同様の操作を行った。
[器具]
セミドライブロッティング装置(The Trans-Blot Semi-Dry Electrophoretic Transfer (Bio-Rad, Hercules CA))
撮影装置(The AE-6962 Light Capture system, the AE-6905H Image Saver, the AE-6955 Cool Saver(ATTO, Tokyo)) [キット、試薬の調製]
(1)
2×Treatment
buffer :
4%(w/v)SDS、125 mM Tris-HCl(pH 6.8)、20%(w/v)
グリセロール、10%(v/v)2-メルカプトエタノール、
0.05%(w/v)BPB
実験に使用するまで-40℃で保存した。(2)
Tank buffer:
25 mM Tris、192 mM グリシン、0.1%(w/v)SDS
(3)
Transfer buffer :
25 mM Tris、192 mM グリシン
(4)
TBS :
20 mM Tris-HCl(pH 7.5)、0.5 M NaCl
(5)
TBST :
20 mM Tris-HCl(pH 7.5)、0.5 M NaCl、0.05%(w/v)Tween
20
(6)
Blocking
buffer :
5%(w/v)スキムミルク、1×TBST
(7)
Stripping
Buffer:
62.5 mM Tris-HCl(pH 6.7)、2 %(w/v)SDS、 100 mM
2-メルカプトエタノール
実験に使用するまで室温で保存した。29 2.2.9 グ リ コ ー ゲ ン 定 量 [サンプル調製] 採取した骨格筋 (足底筋、腓腹筋、ヒラメ筋)、肝臓を液体窒素によって冷却したステ ンレス製乳鉢と乳棒を用いて粉末状にした。骨格筋を約30 mg、肝臓を約 50 mg それぞれ 秤量し、30% KOH (硫酸ナトリウム飽和溶液) を 1000 µl 加え、90℃の湯浴で 30 分間加熱 した。加熱の途中と直後に激しく撹拌し、液体の温度が下がるまで氷上で冷却した。100% エタノールを1200 µl 加えよく混合させた後、そのまま氷上で 30 分間静置した。静置後 に撹拌、遠心分離 (7500×rpm、30 分、4℃) し、上清をアスピレーターによって除去し デシケーター中で風乾させた。筋グリコーゲンにおいてはMilliQ 水 3 ml を、肝臓グリコ ーゲンにおいてはMilliQ 水 5 ml を加えて溶解し、各臓器のグリコーゲンサンプルを得た。 [測定方法] 筋グリコーゲン量は、フェノール硫酸法を用いて測定した[43]。筋グリコーゲンの定量 には原液のサンプル、肝臓グリコーゲンの定量には4 倍希釈したサンプルを用いた。そ れぞれサンプル200 µl に 5%(w/v)フェノール 200 µl を加えた後、速やかに濃硫酸 1000 µl を静かに加え十分に混合させた。室温で 10 分間静置した後、25~30℃の水浴で 15 分 間振盪させて反応を止め、波長490 nm の吸光度を測定した。 2.2.10 ク エ ン 酸 合 成 酵 素 活 性 測 定 [測定方法] クエン酸合成酵素測定方法は、Shepherd らの方法に従った [44]。試料中のクエン酸合 成酵素が、acetyl-CoA とオキサロ酢酸からクエン酸と CoA-SH を生成する反応を触媒す る。このとき生成したCoA-SH をエルマン試薬(5,5'-dithio-bis-[2-nitrobenzoic acid]: DTNB) と反応させると、412 nm にピークの吸光波長をもつメルカプチドイオンが生成される。 このメルカプチドイオンの生成速度を求め、クエン酸合成酵素活性を測定した。
30
30℃でインキュベートした Assay cocktail を 955 µl に 20 mM acetyl-CoA を 15 µl、タン パク質抽出試料を10 µl 加え、10 分間 30℃でインキュベートし、吸光波長 412 nm の吸光 度変化を2 分間記録したものをベースラインとした。その後、20 µl の 25 mM オキサロ酢 酸を加え反応させ、吸光度の変化から、一分間に生成された2-ニトロ-5-メルカプト安息 香酸(TNB)の量を算出した。1 µmol/分を 1 U とし、クエン酸合成酵素タンパク質 1 mg で1 分間に生成する TNB の量を U/mg protein で表した。 [器具] 分光光度計 UV-160A(島津製作所、京都) 吸光度RECORDER U-135(島津製作所、京都) [試薬調製]
(1)
1 M Tris-HCl(pH 8.1):
Tris(hydroxymethyl)aminomethane、MilliQ 水
6 N HCl で pH 8.1 に調節し、4℃で保存した。
(2)
20 mM Acetyl-CoA:
Acetyl Coenzyme A Trilithium Salt、 1 mM HCl
(3)
25 mM Oxaloacetic acid:
Oxaloacetic acid、 0.1 M Tris-HCl
(4)
1 mM DTNB:
DTNB、1 M Tris-HCl(pH 8.1)
(5)
Assay Cocktail :
1 mM DTNB、 MilliQ 水
(1 mM DTNB 1 ml に対し、 MilliQ 水を 8.55
ml の割合で混合した。)
2.2.11 シ ト ク ロ ム c オ キ シ ダ ー ゼ ( cytochrome c oxidase; COX) 活 性 測 定
[測定方法] シトクロムc オキシダーゼの活性は、Wharton らの方法に従った [45]。試料中のシト クロムc オキシダーゼの触媒反応によって、還元型のシトクロム c は酸化型に変換され る。還元型のシトクロムc は 550 nm の特異的な吸光波長のピークをもつが、酸化型は持 たない。この吸光波長におけるピークの違いを利用して、550 nm の吸光度の減少速度か ら還元型シトクロムc の消費速度を算出し。シトクロム c オキシダーゼ活性を求めた。
31 30℃でインキュベートした 880 µl の MilliQ 水に、0.1 M リン酸緩衝液を 100 µl、還元 型シトクロムc を 10 µl 加え、30℃で 5 分間インキュベートした後、吸光波長 550 nm に おける吸光度変化を2 分間測定し、これをベースラインとした。その後、タンパク質抽 出試料を10 µl 加え反応させ、吸光度の変化から、一分間に減少する還元型シトクロム c の量を算出した。1 µmol/分を 1 U とし、タンパク質 1 mg が 1 分間に減少させる還元型シ トクロムc の量を U/mg protein で表した。 [器具] 分光光度計 UV-160A(島津製作所、京都) 吸光度RECORDER U-135(島津製作所、京都) Zeba スピン脱塩カラム(サーモフィッシャーサイエンティフィック株式会社、神奈川) [試薬調製]
(1)
0.1 M Phosphate buffer:
(
pH 7.0、4℃で保存し
た)
0.2 M NaH
2PO
4・
2H
2O, 0.2 M Na
2HPO
4・
12H
2O
(2) 還元型シトクロム c 溶
液
:
30mg/ml シトクロム c, 0.01 M リン酸緩衝
液
30 mg シトクロム c を 0.01 M リン酸緩衝液 1
mL に溶解し、さらにアスコルビン酸を数 mg
加え混合した。次いで、過剰なアスコルビン
酸を
Zeba スピン脱塩カラムを用いて除去し
た。
2.2.12 統 計 処 理 結果は全て平均値±標準誤差(means ± S.E.)の形式で表示した。検出限界以下の物質 濃度は、同群内で検出されたサンプルの最低濃度の半値として統計を行った[46]。全ての データの統計処理は、Stat View 5.0 (Abacus Concepts, Inc.Berkeley, CA) を用いて、2 群間32
の比較はStudent t-test により、3 群以上の比較は 2 way ANOVA を行ったあと Tukey-Kramer による有意差検定を行った。なお、有意水準は P < 0.05 とした。 2.3 結 果 2.3.1 運 動 持 久 力 ( 実 験 1 ) 2 週間トレーニング前の走持久力及びトレーニング後の走持久力を Fig. 2.1 に示した。 トレーニング前の走持久力はBDK-mKO マウスと Control マウスで変わらなかった。2 週 間のトレーニングによって、走持久力はBDK-mKO マウスと Control マウスともにトレー ニング前より約2 倍増加した。しかし、トレーニング後の走持久力では Control マウスよ りBDK-mKO で有意に低値であった。この結果より、BDK-mKO マウスでは、運動トレ ーニングに対する運動持久力の適応が抑えられる可能性が示唆された。 2.3.2 マ ウ ス の 体 重 と 組 織 重 量 ( 実 験 2 ) 実験最終日のControl マウスおよび BDK-mKO マウスの体重には、4 群間で差はなかっ た(Table 2.2)。実験最終日に採取した臓器のうち、骨格筋、心臓、肝臓、精巣上体脂肪 の体重100g 当たりの重量は、いずれの群間でも差を認めなかった(Table 2.2)。 2.3.3 マ ウ ス の 各 組 織 に お け る BDK 量 ( 実 験 2 ) 実験2 で用いた BDK-mKO マウスの筋特異的 BDK 欠損を確認するために、ウェスタン ブロット法により、体内の主要な組織である脳、心臓、骨格筋、肝臓、腎臓、膵臓、脾 臓と精巣におけるBDK のタンパク質量を測定した(Fig. 2.2)。その結果、BDK-mKO マ ウスにおいて、Control マウスと比較して、骨格筋及び心筋でのみ BDK 量の著しい低下 が確認された。 2.3.4 各 群 の マ ウ ス の 骨 格 筋 に お け る BCKDC 量 ( 実 験 2 )
33
各群のマウスの骨格筋BCKDC の E1α、E1β と E2 をウェスタンブロットにより検出し た結果をFig. 2.3 に示した。BCKDC の E1α、E1β と E2 のタンパク質量には各群間で差は なかった。BDK-mKO マウスで E1α は脱リン化しているため、そのバンドの位置は Control マウスと比較してわずかに低分子側に検出された。
2.3.5 血 漿 BCAA 濃 度 ( 実 験 2 )
血漿ロイシン、イソロイシンとバリンの濃度は、急性運動と関係なく、BDK-mKO で 明らかに低値であった(Fig. 2.4(A-C))。急性運動によって血漿 BCAA 濃度が Control マウスで低下する傾向にあったのに対して、BDK-mKO マウスでは有意に上昇した。 2.3.6 BCAA 以 外 の 血 漿 ア ミ ノ 酸 濃 度 ( 実 験 2 ) 安静状態のマウスでは、血漿アスパラギン酸濃度がControl マウスよりも BDK-mKO マ ウスで高い傾向にあったが、他の血漿アミノ酸濃度では両マウス間で差はなかった(Fig. 2.5)。血漿 BCAA 濃度で観察されたように、Control マウスの多くの血漿アミノ酸濃度 が急性運動により低下する傾向にあったのに対して、BDK-mKO マウスでは逆に上昇す る傾向にあった(Fig. 2.5)。そのため、血漿アラニン、アルギニン、アスパラギン酸、 グルタミン酸、グリシン、ヒスチジン、トレオニンの濃度が急性運動により、Control マ ウスに比べて、BDK-mKO マウスで有意に高値であった(Fig. 2.5)。血漿リシンも同様 な傾向にあった。 2.3.7 筋 肉 の エ ネ ル ギ ー 代 謝 に 対 す る BDK-mKO と 急 性 運 動 の 影 響 ( 実 験 2 の メ タ ボ ロ ー ム 解 析 ) ( 1 )BCAA お よ び そ の 代 謝 系 中 間 体 ( Fig. 2.6) 筋肉BCAA レベルは、両マウスの安静群および急性運動群ともに、BDK-mKO マウス で著しい低値であった(A-C)。筋肉の BCKA レベルは、BDK-mKO マウスで検出限界 以下まで低下した(E)。一方、acyl-CoA の代謝産物の一つである acyl-carnitine レベルは、 安静群および急性運動群ともにBDK-mKO マウスで有意な高値であり、Control マウスの
34
isobutyryl- carnitine のみ急性運動で上昇した(F, G)。これらの結果より、BDK-mKO マ ウスでは、BCAA 分解により生成した BCKA の多くは速やかに acyl-carnitine へ代謝され たと考えられる。BCAT の補酵素であるpyridoxamine 5'-phosphate(PMP)レベルが、安 静群および急性運動群ともにControl マウスよりも BDK-mKO マウスで低値傾向にあった (D)。
( 2 ) 解 糖 系 代 謝 中 間 体 (Fig. 2.7)
解糖系の成分では、glucose 6-phosphate(G6P)、fructose 1,6-bisphosphate(F1,6BP)、 dihydroxyacetone phosphate(DHAP)、2-phosphoglycerate(2PG)、phosphoenolpyruvate
(PEP)、lactate が検出された。G6P レベルは群間で差が見られなかった(A)。解糖
系の律速階段と見られている第三ステップの代謝産物であるF1,6BP レベルは、安静群お よび急性運動群ともにControl マウスよりも BDK-mKO マウスで低値を示した(B)。安 静群のDHAP(C)、2PG(D)、PEP(E)レベルは、いずれも Control マウスよりも BDK-mKO で低値傾向にあり、Control マウスでのみ急性運動により低下する傾向を示した。Lactate レベルでは各群間で差はなかった(F)。
( 3 )Acetyl-CoA お よ び TCA サ イ ク ル 代 謝 中 間 体 ( Fig. 2.8)
Acetyl-CoA レベルは、安静群および急性運動群ともに Control マウスよりも BDK-mKO マウスで低値を示し、Control マウスでのみ急性運動により低下した(A)。安静群の citrate (B)と isocitrate(C)レベルは、Control マウスよりも BDK-mKO マウスで低値を示し、 急性運動によりControl マウスでは低下傾向、BDK-mKO マウスでは上昇傾向を示した。 Succinate(D)、fumarate(E)、malate(F)レベルは各群間で差はなかった。以上のこ とから、筋特異的BDK ノックアウトは、acetyl-CoA と TCA サイクル前半の代謝中間体 を減少することが判明した。
35
TCA サイクルや BCAA 代謝で生じる NADH レベルは、安静群および急性運動群とも にControl マウスよりも BDK-mKO マウスで低値傾向を示し、Control マウスでのみ急性 運動により低下する傾向を示した(A)。しかし、NAD+(B)、ATP(C)、GTP(D)、 phosphocreatine(E)レベルは各群間で差はなかった。 ( 4 ) 骨 格 筋 に お け る BCAA 以 外 の ア ミ ノ 酸 レ ベ ル ( Fig. 2.10) 骨格筋のBCAA 以外のアミノ酸レベルを Fig. 2.10 に示した。アラニン、アスパラギン 酸、ヒスチジン、リシン、トレオニンの濃度が、安静群および急性運動群ともにControl マウスよりもBDK-mKO マウスで高値傾向を示したが、いずれも急性運動の影響はほと んどなかった。トリプトファンレベルは、Control マウスと BDK-mKO マウスで差はなか ったが、いずれのマウスでも急性運動により上昇した。 2.3.8 グ リ コ ー ゲ ン 量 ( 実 験 2 ) 肝臓グリコーゲン量と骨格筋グリコーゲン量をFig. 2.11(A, B)に示した。肝臓グリコ ーゲン量は、いずれの群間でも差が見られなかった(A)。骨格筋グリコーゲン量は、安 静群および急性運動群のいずれにおいてもControl マウスよりも BDK-mKO マウスで低値 を示し、両マウスで急性運動により有意に減少した(B)。 2.3.9 ミ ト コ ン ド リ ア エ ネ ル ギ ー 代 謝 系 の 酵 素 活 性 ( 実 験 2 ) 骨格筋のクエン酸合成酵素活性とシトクロムcオキシダーゼの活性をFig. 2.12(A, B) に示した。いずれの酵素活性も、Control マウスよりも BDK-mKO マウスで低値を示し、 いずれのマウスでも急性運動により上昇傾向を示した。 2.4 考 察 これまでの報告より、慢性的なBCAA 濃度減少状態では、運動に負の影響を与えると 推察されるが[47-50]、一方で BCAA の代謝亢進状態では運動時により多くのエネルギー
36 供給を可能にするとも考えられる[51]。このように BDK-mKO マウスでは運動において 2 面性が考えられる。そこで本実験では、まずBDK-mKO マウスの走運動持久力を測定し た。その結果、トレーニングしていないBDK-mKO マウスと Control マウスの間では運動 持久力に差が見られなかった。このことによりBCAA 代謝亢進の結果引き起こされる慢 性的BCAA 濃度減少は非トレーニング状態マウスの運動持久力に影響を与えないことが 示唆された。次に2 週間のトレーニングを負荷したマウスで運動持久力を測定したとこ ろ、トレーニング前にくらべ両群とも2 倍以上に運動持久力の上昇がみられたが、Control マウスに比べてBDK-mKO マウスの運動持久力が有意に低値となった。これらの結果よ り、BDK-mKO マウスでは、トレーニングにより運動持久力は上昇するが、その上昇が 一部抑制される可能性が示唆された。 BDK-mKO マウスでこのような現象が見られた原因を解明するために、運動時の骨格 筋のメタボローム解析、および運動持久力に影響を与える筋グリコーゲン量や、エネル ギー生産に関わる骨格筋のクエン酸合成酵素活性およびシトクロムc オキシダーゼ活性 を測定した。
血漿BCAA 濃度は、Control マウスに比べて BDK-mKO マウスで有意に低値であった。 急性運動を負荷すると、Control マウスでは減少傾向に、BDK-mKO マウスでは上昇傾向 にあった。これについては、Control マウスは運動により BCKDH 活性が上昇し BCAA 代 謝が亢進し、BCAA も一つのエネルギー源として運動中に消費されたと推測される。そ れに対して、BDK-mKO マウスでは BCKDH 活性が常に最大に近い値であるため運動に よるタンパク質分解が生じBCAA の流量が上昇したためと推察される。 メタボローム解析におけるBCAA 代謝中間体の解析において、骨格筋の BCAA レベル はBDK-mKO マウスで有意に低い値であり急性運動による変化はなかった。BCAA 代謝 の最初の反応であるアミノ基転移反応により生じるBCKA のうち KIC と KMV は BDK-mKO マウスではすべてのマウスで測定限界を下回っていた。このことにより、
37
BDK-mKO マウスでは生じた BCKA が速やかに代謝されたと推察される。BCAA は完全 代謝されるとacetyl-CoA および succinyl-CoA となり TCA サイクルに入るが、BDK-mKO マウスではacetyl-CoA-濃度が有意に低い値を示した。一方、BCKDC 反応により生じる branched-chain acyl-CoA は carnitine と反応し branched-chain acyl-carnitine を生成すること ができる。BDK-mKO マウスではこの branched-chain acyl-carnitine が有意な高値を示した。 このことから、BDK-mKO マウスでは BCAA の代謝は亢進しているが、TCA サイクルの 基質とはならずbranched-chain acyl-carnitine の形で流出しておりエネルギー産出への寄与 が低い可能性が示唆された(Fig. 2.13)。
糖代謝中間体については、F1,6BP, DHAP, 2PG, PEP レベルが BDK-mKO マウスで減少 する傾向が見られた。さらに、各エネルギー源による生じるTCA サイクルの基質である acetyl-CoA と TCA サイクルの前半の代謝中間体が BDK-mKO マウスで減少した。これら の現象と関連して、BDK-mKO マウスでは、BCAA 分解経路の第一ステップの反応と関 連するグルタミン酸、アラニンとアスパラギン酸が有意に上昇したことから、BCAA 代 謝の第一ステップのアミノ基転移反応が運動中にも亢進された。そのため、このアミノ 基転移反応で必要なオキサロ酢酸とビルビン酸が消費され、acetyl-CoA と TCA サイクル の前半の代謝中間体がBDK-mKO マウスで低下した原因と推測される(Fig. 2.13)。これ らの運動にかかわるエネルギー代謝基質の不足が、BDK-mKO マウスの運動適応力の低 下を引き起こす可能性が考えられる。一方、TCA サイクル後半の代謝物であるコハク酸、 フマル酸とリンゴ酸レベルは、BDK-mKO マウスでも減少しなかったのは、正確な理由 は不明であるが、一部はイソロイシンおよびバリンの分解から生成されるsuccinyl-CoA の供給による可能性が考えられる。BDK-mKO マウスの骨格筋では、これらのエネルギ ー代謝の乱れはあったもののATP などの高エネルギー化合物の減少は認められなかった ので、さほど深刻なエネルギー代謝の乱れとは考えられない。いずれにしても詳細なメ カニズムの解明には今後さらなる検討が必要である。
38 ミトコンドリア内の酵素である、クエン酸合成酵素活性、シトクロムc オキシダーゼ 活性は、Control マウスに比べて BDK-mKO マウスで低い傾向にあった。以前の研究から 中高齢マウスにBCAA を摂取させると、筋ミトコンドリアの合成と機能を促進すること が報告された[52]。また、トレーニングがミトコンドリア合成を増大する報告もある[53]。 今回の研究より、BCAA の不足する状態では、トレーニングによる筋ミトコンドリア機 能の上昇が抑制される可能性が示唆された。 筋グリコーゲン量は、Control マウスに比べて BDK-mKO マウスで有意に低い値であっ た。ことからBDK-mKO マウスでは運動トレーニングによるグリコーゲン量の増加が抑 制されている可能性が示唆された。正常状態では、運動トレーニングにより骨格筋での 糖輸送体GLUT-4 の増加により、筋細胞へのグルコース取り込みも上昇することが知ら れている。さらに、運動後のグリコーゲン合成酵素活性はインスリンによって安静状態 より活性化され、グリコーゲンを効率よく合成することができるので、筋グリコーゲン 回復を増大することが明らかにされている[54-56]。一方、インスリン抵抗性を呈する疾 病である肝硬変においては血中BCAA 濃度が有意に低下するが、その肝硬変モデル動物 にBCAA を与えると、GLUT4 の筋細胞膜への移動を促すことで耐糖能異常が改善される 報告がある [57]。これらのことから、BDK-mKO マウスでは、BCAA 不足により筋への グルコース取り込みを低下させることによって、グリコーゲンの回復に負の影響を与え ると推測される。また急性運動を負荷することにより筋グリコーゲン量が両マウスとも 同程度減少していたことから、運動による筋グリコーゲンの消費にはBDK-mKO の影響 は少ないと判断される。 以上より、トレーニングしたBDK-mKO マウスで運動持久力が Control マウスに比べて 有意に低くなった原因としては、筋グリコーゲン量の減少、ミトコンドリア酵素活性の 低下によるエネルギー産生能の低下、BCAA 代謝物がエネルギー生産とは異なる運命を たどることなどの可能性が示唆された。しかし、正確な原因については不明である。今
39
後この原因についてさらに究明するために、BDK-mKO マウスに BCAA を投与して、 BCAA 不足を解消する研究が必要と考えられる。
40
Table 2.1. The ingredients of the diet
Energy source g% kcal%
Protein 19.2 20
Carbohydrate 67.3 70
Fat 4.3 10
Total energy (kcal/g) 3.85 100
Ingredient g kcal Casein, 30 Mesh 200 800 L-Cystine 3 12 Corn starch 315 1260 Maltodextrin 10 35 140 Sucrose 350 1400 Cellulose, BW200 50 0 Soybean oil 25 225 Lard 20 180 Mineral mix S10026 10 0 Dicalcium phosphate 13 0 Calcium carbonate 5.5 0
Potassium citrate, 1 H2O 16.5 0
Vitamin mix V-10001 10 40
Choline bitartrate 2 0
41
Table 2.2. Body and tissue weights of control and BDK-mKO mice with and
without the running exercise bout
Values are means ± SE. Skeletal muscle is a mixture of soleus, gastrocnemius, and plantaris muscles of the left hind-limb. Both sides of epididymal adipose tissues were collected from mice and
42
Figure 2.1. Exercise performance of control and BDK-mKO mice
Running distance to exhaustion before and after 2 weeks of training.43
Figure 2.2. Western blotting of BDK and E2 component of the BCKDC in control
and BDK-mKO mice.
Tissue extracts were applied on SDS-PAGE, followed by transfer of proteins to PVDF membranes and immunostaining of BDK and E2 component, as described in Materials and Methods. Protein amounts per lane applied on the SDS-PAGE were 25 µg for heart, kidney, and pancreas; 30 µg for brain; and 50 µg for skeletal muscle, liver, spleen, and testis.
44
Figure 2.3. Typical Western blots of the BCKDC extracted from skeletal muscle of
control and BDK-mKO mice.
Age-matched untrained mice (both control and BDK-mKO mice) were prepared for this experiment. Therefore, trained and untrained mice (both control and BDK-mKO mice) were used in the Western blotting. The tissue extracts (gastrocnemius and plantaris muscles) were applied on SDS-PAGE, followed by transfer of proteins to PVDF membranes and immunostaining of BCKDC, as described in Materials and Methods. Protein amounts per lane applied on the SDS-PAGE were 25 µg.
45
Figure 2.4. Plasma BCAA concentrations of control and BDK-mKO mice with and
without the running exercise bout
# Significant difference between control and BDK-mKO mice. * Significant difference in the same group of mice with and without the exercise bout.
46
Figure 2.5. Plasma BCAA concentrations of control and BDK-mKO mice with and
without the running exercise bout
Ala, alanine; Arg, arginine; Asn, asparagine; Asp, aspartate; Gln, glutamine; Glu, glutamate; Gly, glycine; His, histidine; Lys, Lysine; Met, methionine; Phe, Phenylalanine; Ser, serine; Thr, threonine; Trp, tryptophan; Tyr, tyrosine.
# Significant difference between control and BDK-mKO mice. * Significant difference in the same group of mice with and without the exercise bout.
47
Figure 2.6. BCAAs and their metabolites
Changes in the metabolite levels in the skeletal muscle of control and BDK-mKO mice with and without the exercise bout are shown.
# Significant difference between control and BDK-mKO mice. * Significant difference in the same group of mice with and without the exercise bout.
48
Figure 2.7. Metabolites in the glycolytic pathway
Changes in metabolite levels in skeletal muscle of BDK-mKO mice and control mice with and without the exercise bout are shown.
49
Figure 2.8. Acetyl-CoA and metabolites in the TCA cycle
Changes in metabolite levels in skeletal muscle of BDK-mKO mice and control mice with and without the exercise bout are shown.
# Significant difference between control and BDK-mKO mice. * Significant difference in the same group of mice with and without exercise bout. a
50
51
Figure 2.10. Muscle levels of amino acids except for BCAAs in control and
BDK-mKO mice with and without the exercise bout.
Ala, alanine; Arg, arginine; Asn, asparagine; Asp, aspartate; Gln, glutamine; Glu, glutamate; Gly, glycine; His, histidine; Lys, Lysine; Met, methionine; Phe, Phenylalanine; Ser, serine; Thr, threonine; Trp, tryptophan; Tyr, tyrosine.
# Significant difference between control and BDK-mKO mice. * Significant difference in the same group of mice with and without the exercise bout.
52
Figure 2.11. Glycogen contents in skeletal muscle and liver of control and
BDK-mKO mice with and without the exercise bout
# Significant difference between control and BDK-mKO mice. * Significant difference in the same group of mice with and without the exercise bout.
53
Figure 2.12. Citrate synthase and cytochrome c oxidase activities
# Significant difference between control and BDK-mKO mice.54

![Figure 1.3. Conditional BDK gene knockout using the Cre–loxP system [30]](https://thumb-ap.123doks.com/thumbv2/123deta/6871883.745940/16.892.130.744.160.731/figure-conditional-bdk-gene-knockout-using-cre-loxp.webp)
![Figure 1.4. Growth curves from 8 weeks old to 13 weeks old of control and BDK-mKO mice [30]](https://thumb-ap.123doks.com/thumbv2/123deta/6871883.745940/17.892.134.633.164.484/figure-growth-curves-weeks-weeks-control-bdk-mice.webp)
![Figure 1.5. Western blotting of BDK in control and BDK-mKO mice [30]](https://thumb-ap.123doks.com/thumbv2/123deta/6871883.745940/18.892.135.470.194.611/figure-western-blotting-bdk-control-bdk-mko-mice.webp)
![Table 1.1. Tissue weights of the control and BDK-mKO mice [30]](https://thumb-ap.123doks.com/thumbv2/123deta/6871883.745940/19.892.129.687.232.530/table-tissue-weights-control-bdk-mko-mice.webp)
![Table 1.2. Plasma concentrations of amino acids in control and BDK-mKO mice [30]](https://thumb-ap.123doks.com/thumbv2/123deta/6871883.745940/20.892.129.779.257.853/table-plasma-concentrations-amino-acids-control-bdk-mice.webp)
![Table 1.3. Concentrations of amino acids in heart muscle of control and BDK-mKO mice [30]](https://thumb-ap.123doks.com/thumbv2/123deta/6871883.745940/21.892.132.682.270.782/table-concentrations-amino-acids-heart-muscle-control-bdk.webp)
![Table 1.4. Concentrations of amino acids in skeletal muscle of control and BDK-mKO mice [30]](https://thumb-ap.123doks.com/thumbv2/123deta/6871883.745940/22.892.132.737.268.871/table-concentrations-amino-acids-skeletal-muscle-control-bdk.webp)

