理 学 療法 学 第
33
巻 第2
号76
〜
81
頁 (2006
年 )研 究 報 告
尾
部
懸
垂
前
の
運
動
が
廃
用
性萎
縮
筋
の
ユ
ビ
キ チ
ン ー
プ
ロ
テ ア
ソ
ー ム
系
に及
ぼ
す 抑 制
効
果
*藤
野 英 己
1
−
3
)#上
月
久 治
3
)4
)武
田
功
D
田
崎 洋 光
1
)近 藤 浩 代
1
>石 田 寅 夫
2
)梶 谷 文 彦
3
)5
) 要旨
廃 用 性
筋
萎 縮の予 防 法 と し て プレコ ンディ ショ ニ ング 運動
の 制御 機構
につ い て,
タ ンパ ク質 分 解
に作
用 す るユ ビキ チンー
プロ テア ソー
ム系
, 及びタン パ ク質新
生・
修 復
に 関与 す
る熱
ス トレ ス タ ンパ ク質 (
HSP
)
につ いて検
討 し た。
ラッ トを 用いて,1
)
2
週 間の尾 部 懸 垂 群 (
HS
),
2
)
HS
直 前
に トレッ ドミ ル走 行 を行
い2
週 間の尾 部 懸 垂 を し た 群 (ExHS
),
及 び3
) 対 照 群 に 分 類 し た。
ヒ ラ メ筋
を 摘 出 して,
筋
原線 維
タ ン パ ク質
量 を計
測 し, リアル タ イム定
量PCR
法
でユ ビキ チン リガー
ゼE3
, 及び72k
−
Da
HSP
(
HSP72
)
の mRNA を 測定
し た。
筋
原線 維
タンパ ク質
量 は,
対 照群
と 比較
し てHS
とExHS
で は有 意
な低
ドを示
し た が,
ExHS
ではHS
に 比 較 して有 意 に 高 値 を示 し た。一
方,
ユ ビキ チ ン リガー
ゼE3
mRNA
は,
対 照 群 と比 較 してHS
で有
意 に増 加
し,
ExHS
で は増 加
を 示 さ な かっ た。
ま た,HSP72
mRNA は, 対 照 群 と比 較 してHS
で有
意 に減
少 し,
ExHS
で は 減 少 が抑
制 さ れ た。
プ レコ ンデ ィショ ニ ング 運動
はユ ビキ チ ンリ ガー
ゼE3
の活
性化
を 抑 制 し,
HSP
の低 下 を 減 衰 さ せ た。
これ らの結 果 か ら プレコ ンデ ィショ ニ ング 運 動 は 尾 部懸
乖中
の筋
タンパ ク質分 解 系
の活性 化
を抑 制 す
る制 御 機 構
が働 く
こと が示 唆
さ れ た。
キー
ワー
ドユ ビ キ チンリ ガ
ー
ゼ,
熱
ス ト レス タンパ ク質
,
mRNA緒
言骨 格 筋
の廃用 性 萎 縮
は収 縮
タ ンパ ク質
の合
成 と分 解
の バ ラン スが崩 壊 し
,
タ ンパ ク質
の分解 速 度 が 合 成 速 度 を
*lnhibltory
Effects
ofExercise
・
preconditioningbeforc
Hindljmb
Suspension
onUbiquitin
−
proteasomePathway
Actlvity
in
M
し1sc}e
Atrophy
l
〕 鈴 鹿 医 療 科 学 大 学 保 健 衛 生 学 部(〒
510
−
0293
≡重県 鈴 鹿iij岸 岡町]1001
−
1
)
Hidemi
Fuji
[10,
RPT
,
PhD
,
王saoTakeda
,
RPT
,
PhD
,
Hiromitsu
Tasaki
,
RPT
、
MS
,
Hir
【}yoKondo
,
MS
:FacuLL
}・
o!HealLh
Sciences
,
Suzuka
University
ofMedical
Sciellc
:e2
) 同 東 洋医学研究 所
Hidemi
Fujin
〔},
RPT
,
PhD
,
Torao
Ishidu
,
PhD
:Institute
〔エf
Traditio
”alMedicine
,
Suzuka
University
(,f
Medica
ユScience
3
)岡 山大 学 大 学 院 医 歯 蘂 学 総 合 研 究 科システム循 環 牛 理 学
Hidemi
Fujino
,
RPT
,
PhD
,
Hisaharu
Kohzuki
,
MD
,
PhD
,
Fumihiko
Kajiya
,
MD
,
PhD
;Departnlent
ofCardiovascutar
Physiology
,
Okayama
University
Craduate
Schoot
ofMedicine
,
Dentistry
andPharmaCeUtiCat
SCiellCeS
d
)岡 山 学 院 大 学 人 間 生活 学部
Hisaharu
Kohzuki
,
MD
.
PllD
:Department
ofFood
andNutrition
.
Okayama
Gakuin
University
5
)川崎 医 科大学 医用 工学・
川 崎 医療短 期 大学
F
しlmihik (,Kajiya
,
MD
,
PhD
:DeparLmenL
ofMedical
Engineering
,
Kawasaki
Medical
Scho
【)L
,
andKawasaki
College
〔)f
Allied
Health
Pr
〔,fessi
(,1hS #E
−
mail :fujino
@.
suzuka.
u.
ac.
jp
〔受 イ
・
J
日2DO5
年10
月27
H
/受理 日2DO6
奪2
月2
目 }超
え る た めに発
生す
ること が知
ら れてい る1−
3〕。
ラ ッ ト の尾 部 懸
垂による不 荷
重直 後
にはタ ン パ ク質
の合 成低
下 が 主因
で筋
タン パ ク質
の減
少 が 起こり
1〕2)’
1),
数時
間 後 にはタ ンパ ク質 分 解 速 度
が増 加
して,
相 対 的
に タン パ ク質
の分 解
が優 位
と なる と考
え ら れている 1)2
)。
ラ ッ トヒ ラ メ筋
の タ ンパ ク質 損
失経
過 を観 察
した研
究で は,
尾部
懸 垂
5
時 間後
に ヒラメ筋
の筋
タ ンパ ク質
が減 少
し,
2
週
間 ま
で筋
タ ンパ ク質
の分 解
が促
さ れ て い るD2
)。 ま た,
先 行研 究
に お いても
2
週間
の尾 部懸
垂で筋 原 線 維
タン パ ク質 量
が40
% 減 少 す
る ことを確 認 し
て い る51/ 6)。 これ ら筋
原線 維
タ ンパ ク質 量
の減 少
は タ ンパ ク質
の分 解系
が活性 化
され た こ とで生 じ た現 象
であ
ると考
えら れ る上一
3)。
筋
タン パク質
の分 解系
と して働 く経 路
とし
て,
リソ ソー
ム系
,
カ ル パ イン系
,
及びユ ビキ チンー
プロ テアソー
ム系
が あ り, ユ ビキ チ ンー
プロテアソー
ム系
に よ る経 路
は廃
用性 筋 萎 縮
に お け る タン パ ク質 分解 系
の促 進
に強 く関
与 し てい る と 考 え ら れ てい る3)。
ユ ビキ チ ンー
プロ テ ア ソー
ム系
は3
つ の 酵素
であるユ ビキ チン活 性 化 酵 素
, ユ ビキ チン結 合酵 素
,
ユ ビキ チン連 結 酵 素 (
ユ ビ キ チンリ ガー
ゼE3
) が 関 与 す る 漣 の経 路 で作
用 す る3)7)。
Japanese Physical Therapy Association
NII-Electronic Library Service
Japanese Physioal Therapy Assooiation
尾 部懸重前の運 動が廃用性 萎 縮 筋のユ ビキ チン
ー
プロテ ア ソー
ム系に及ぼす 抑 制 効 果77
HSP )
はタ ン パ ク質
の新
生,
修
正等
に 関 与 す る 4〕8)。
HSP
は 変 性 過 程の タン パ ク質
を修
止し, リボ ソー
ム で 合 成 さ れ た新
生 ポ リペプ チド鎖
の伸 張 を補 助 す
る分
子 シ ャ ペ ロ ン と して の機 能
をも
つ 8〕。
特
に70k
−Da
HSP
ファ ミリー
に属 す
るHSP72
は細 胞 質 内
に存 在
し,様
々 なス トレ スに よっ て誘 導 され
るHSP
であ
る8)。
HSP72
は一・
過性
の 運動
6)9)10),
電 気 刺 激
11)や 温 熱負
荷 12丿に よ り増 加
し,
筋
タ ンパ ク質 分 解
の抑 制
12 )や筋
肥 大 ]3
) に関
与 す
る ことが 報 告
さ れ ている。
先 行
研 究 で は 尾部 懸
垂前
の プレ コ ン デ ィ シ ョ ニ ン グ運 動 でHSP72
が 誘導
さ れ,
そ れに伴 う尾 部 懸 垂 中
の筋
原線 維
タンパ ク質
量の減 少
を抑 制
でき
る ことを報 告
し た6)。
し か し,
プ レコ ン デ ィシ ョ ニ ン グ運動
の筋
萎 縮 予 防 に 対 す る 制 御 機構
につ い て は明確
に でき
なかっ た。
そ
こ で,
本研 究
で は 尾 部懸 乖 前
に行 う
プレコ ン デ ィシ ョ ニ ン グ運動
が 廃 用 性 筋 萎 縮 を予
防す
る制御機 構
を検 証
す
る た め に タ ンパ ク質
分解 系
に働 く
ユ ビキ チ ンー
プロ テ アソー
ム系
のユ ビ キ チン リ ガー
ゼE3
mRNA,
及
び分 子
シ ャペ ロ ン機
能 を もつHSP72
mRNA の変 化
につ いて検
討
した。
方
法
ユ.
実
験 動 物Wistar
雄 性
ラッ ト15
匹 (8
週齢
,
体 市
ユ86
±8g
)を
使
用 し た.
こ れ らラ ッ ト を 次の3
群
に区 分
し た。
1
)
Morey
法 1’
t〕15)に より
2
週 間
の尾 部 懸 垂 を 行
っ た群
(
HS ,
n=
5)
,
2)
尾部 懸
垂 を行 う前
に25
分 問
の トレッ ドミル走 行 (プレコ ンデ ィシ ョ ニ ン グ運 動
,
20m
〆s
, ト り 坂20
°
)
を して,
2
週 間
の尾 部 懸 垂 を 行
っ た群
(
ExHS ,
n≡
5
)
,
及
び3
) 対 照 群 (
Control
,
n
≡
5
) と
し た。
ラ ッ ト は 室 温22
±1
℃の環 境
.
ドで個
別 ケー
ジに て飼育
さ れ,
食 餌
,
及
び飲 水
は自山
と した
。本 研 究 は
,
凵本
生 理学 会
の「
生理 学 領 域
に おけ
る動 物
実 験 に 関 する基本 指 針 」
,
及
び鈴 鹿 医療 科 学
大学 動 物
実 験指 針
に従
っ て行
い,
鈴 鹿 医 療 科 学 大 学 動 物 実 験
承 認(
承 認番
号
;第
20
号
) を得
ている。
2
.
ヒラメ筋
の総
RNA
抽 出
とcDNA
合 成
]6)17)HS ,
ExHS
は2
週 間
の尾 部 懸 垂 を終 了
し た後
,
ま た,
Control
は2
週間
の白 由 飼 育 後
にペ ン トバ ル ビ ター
ル(
50mg
,
ikg,
i
.
p
.
)
で麻 酔 し
て,一
側
ヒラメ筋
を摘
出 し た。
摘 出 し たヒラ メ筋
の中 央 部
2
mg程 度
を 切 り 出 し,
1ml
の
TRIzol
Reagent
(
lnvirtogen
) を 加 え
た16マ イ ク ロチュ
ー
ブ(
エ ッ ペ ンド
ル フ) 内
で均 質 化
(
N556
,
コ スモ バ イ オ ) を行
い,
200
μ♂クロ ロホル ム を加
え,
15
秒
聞
の振 盪 を行
った
。微 量 高 速 冷 却
遠心 機 (
MX −
301
,
ト ミー
精
工)
で遠 心 (
4
℃,
15
分
,
15
,
000rp
,
m.
) 後
,
上清 液
500
μ
に イソ プ ロ パ ノー
ル500
μt
加
えて,
遠 心
(
4
℃,
15
分
,
15,
000r .
p
.
m.
)処
理 し, 総RNA
を沈 殿
さ せ た。
上清 液 を取 り除 き
,
エ タ ノー
ル ]ml を加
えて沈
殿
を洗 浄
し, エ タ ノー
ルを 蒸 発 さ せ た。
RNase
Free
水
を加 え
て,
20
μ」の総
RNA
を抽
出・
精 製
し た。
分 光 光 度
計
(
UV −
1700
,
SHIMADZU
)
で 吸 光度
(
OD
)
260
/280nm
の 比 を 求 め,
精 製 度
の確 認
を行
い,
OD
260
/280nm
比 が1,
7
以.
ヒで あっ た場 合
は良 好 な精 製
が行
われ た も
のと判 断
し た。
次
に ラン ダム プラ イマー
に よるRNA
逆 転 写 酵 素
(
High
Capacity
cDNAArchive
Kit
,
Applied
Biosystems
)
を使 用
して,
逆 転
写 を行
い,
cDNA を合成
し た。
3.
リア ルタ イム定
量Polymerase
chain rcaction(
PCR
)
法
に よ るmRNA の測定
本
研
究 に お い て はTaqman
(
R
)
Gene
Expression
Assays
(
Applied
Biosystems
)を使 川 し
て,
ユ ビ キ チ ン リガ
ー
ゼE3
mRNA(
RnOl535853
−
ml,
Applied
Biosystems
)
,
及
び72
−
kDa
HSP
(
HSP72
)
mRNA(
RnOl487057
−
ml,
Applied
Biosystems
) を 測 定
し た。
ま た,
内 在 性
の コ ン トロー
ル と して リ ボゾー
ム18S
mRNA(
Hs99999901
−
sl,
Applied
Biosystems
) を 測 定 し,
検
量線 法
で各
mRNA量 を計 測
した
。.
各 遺 伝
子の塩
基
配列
は表
1
の通 り
であ
り,
Taqman
(
R
)Gene
Expression
Assays
では各塩 基 配 列
の特 定 部
分のみ を増
幅
して い る。
PCR
に はリ ア ル タイ
ムPCR
シス テ ム(
7500Fast
Real
−
time
PCR
,
Applied
Biosystems
) を使用
し,
TaqMan Fast
Universal
Master
Mix
試薬
(
Applied
BiosysteIns
)
で,
45
サ イ クル の反応
を計
測 し た。
PCR
の条件
はFaSt
モー
ドで全 量
20Pt
の反 応 系 を 用 い て,
1
サイ
ク ル95
℃
,
3
秒
, 及 び60
℃
,30
秒
を繰 り
返 し た。 ユ ビキ
チンリガー
ゼE3
mRNA
,
及 びHSP72
mRNA は各
サ ン プ ル の リボ ゾー
ム18S
mRNA
で補 正 し,
Control
を キ
ャ リブ レー
タ(
Control
を1
と して正 焼化 )
と し た。表
1
リ ア ル タイム定 量
PCR
に使
用 し た mRNA,
及びTaqman
(R
)Gene
Expression
Assays
mRNA
Taqman
(R
)Gene
Expression
Assays
塩 基 配 列 ユ ビキ チン リ ガー
ゼE3
72k
−
Da
HSP
リ ボゾー
ム18S
RnO1535853
mlRnOl487057
mlHs99999901
sl5
’
−
GCCATACTTGTTTTAACCGCCTGGA
−
3
’
5
’
−
CTATGAAAAGGGATGGATCTTTACA
−
3
’
5
’
・
ATTGGAGGGCAAGTCTGGTGCCAGC
−
3
,各
mRNA は 塩基 配 列の一
部 を増 幅してい る.
Taqman
(R
)で使用 さ れて い るプライマー
などの詳 細は非公 開 と なっ てい る,
N工 工一
Eleotronio Library78
理 学療法
学第
33
巻第
2
号4
.
筋 原線 維
タンパク 質の 抽 出⊥8 )と定 量化
RNA
抽 出
した 反対
側後
肢 か らヒラメ筋
を摘 出
し,
液
体 窒 素
で冷 却
し た イソ ペ ンタン内
で瞬 間凍 結 し
,−
80
℃ に冷 凍 保 存
し た。
摘 出 筋の 中 央 部20mg
程 度 を切 り出
し,
20
倍 量 (
v /w)
の抽 出 溶 液 (
シ ョ糖
250mM
,塩 化
カ リ ウ ム100mM
,
イ ミ ダ ゾー
ル20
mM,
EDTA
5
mM
) を
加
えて,
超 音 波
ホモ ジ ナ イ ザー
(
Sonifier
S−
150D
,
BRANSON
)
で均 質 化
し た。
均 質化
し た抽 出溶 液 を
4
℃ で遠心 分 離 (
3
,
000r
.
p
.
m,
101nin
)
し,一
ヒ清 液 を取 り除
き
, ペ レッ トを 回収
し た。
次 に2
種 類の溶 液(
塩 化
カ リ ウム175mM ,0.
5
%
ト リ トンX
−
100
,
及
び塩
化カ リウ ム150mM ,
イ
ミ ダ ゾー
ル20
mM)
を 用 い て,
膜 結 合
ATPase
を取 り除
き,
筋 原 線 維
タンパ ク 質 を 回 収 し た。
Bradford
法 (
Protein
assay,
Bio
−
Rad
)
に よ り, 分光 光
度 計
(
UV −
1700
,
SHIMADZU
)
で595
nm のOD
を測 定
し た。
OD
から筋 原 線 維
タ ンパ ク質
量 を 算 出 し,
ヒ ラ メ筋
lg
重 量 当 た り
の筋 原 線 維
タンパ ク質
量(
mg 〆g
)
と し て表 示
した
。
5
.
統 計 処
理得
ら れ た測 定 値 は平 均
±標 準 誤 差
で表示
し た。
検定
はKruskat
−WaUis
検 定
, 及び事 後 検 定
と してDunn
検
定 を使
用 し た。
ま た,有 意 水 準
5
%
により有 意 差
を判 定
し た。
結
果
2
週間
の尾部 懸 垂
に より
HS
,及
びExHS
の ヒラメ筋
湿
重 量 は そ れ ぞ れ53
士2mg ,
54
±3mg
とな り
,
Control
の91
±2mg
に 比べ て有 意
に減 少
を示
した。
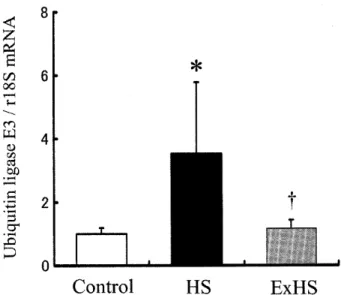
ユ ビ キ チン リ ガ
ー
ゼE3
mRNA(
a
.
u
.
)
はControl
に比
較
して,
HS
で は3
.
5
±2
.
3
で有 意
に増 加 を 示 し
た が,
ExHS
で は1
,
2
±0
,
3
と変 化
し な か っ た(
図1
)
。一
方
,
HSP72
mRNA(
a.
u.
)
はControl
に 比較
し て,
HS
は0
,
3
±0
.
1
と有
意に低
下 し た が,
ExHS
で は0,
8
±0.
2
と変
化 を 示 さ な
かっ た(
図
2
)
。
筋
原線 維
タ ン パ ク質 量
はContro1
がll3
±3mg
/g
で あ るの に 比較
し,HS
で は70
±13mgfg
と有
意に減
少 し た が,
ExHS
で は107
±7mglg
と筋 原 線 維
タン パ ク質
量の減
少 を減 衰
させ た(
図
3
)
。考
察
本研 究
で は廃 用 性 筋 萎 縮
の予 防 対 策
と し て 尾 部懸 垂 前
の 運動 を 行
っ た。 その結 果
,
2
週間
の実
験期
間内
でラ ッ トヒ ラメ筋
の筋 原 線 維
タ ン パ ク質
量の損 失
を減
衰 で き た。
この制 御 機 構
とし
て,
タン パク質分 解 系
で あ るユ ビ キ チン リガー
ゼE3
の活 性 化
を抑 制
してい ること が検 証
で き た。
ま た, タンパ ク質
の分 子
シ ャペ ロ ン作 用
を もつHSP
の低
下を減 衰 す
る こと ができ
た。 こ れ らの結 果
か ら尾 部 懸垂 前
の運 動
は尾 部 懸 垂 中
の筋
タ ンパ ク質
分解
系
8
6
4
2
0
<
耄
の ゜。霍
註
瞭器
b。 =驀
哨
9
哨
^5
*
†
Controi
HS
ExHS
図
1
ユ ビ キ チンリ ガ
ー
ゼ (Ubiquitin
ligase
)E3
mRNA の変
化萎縮筋 (
HS
)で はユ ビ キ チンリ ガー
ゼE3
mRNA が増加し,
筋タンパク質
分解系
の活 性 化が促 進 されて いる.
プレコ ンディ ショ ニ ング筋
(ExHS
>
で はユ ビキチンリ ガー
ゼE3
mRNA が 抑 制さ れ て,
筋タンパ ク質分解 系の活 性 化が抑 制 されて いる.
ユ ビ キ チン リ ガー
ゼE3
mRNA は各サンプル の リボ ゾー
ム18S
mRNA で補正 し,
正常 筋 (
Control
) をキャ リブ レー
タ と して 表 示し てある.
*,
Control
に対 する有 意 差 (p
<0
.
05
);†,
HS
に対 するExHS
の有 意差 (p
<0
.
05
).
<7
[
舷
鬟
の QQ一
』 \N
卜 贏 Qり2
1
0
*
COBtrol
†
HS
ExHS
図2
72k
−
Da
熱ス トレ スタン パ ク質 (
HSP72
)mRNA の変化萎 縮 筋 (
HS
)ではHSP72
mRNA が 減 少 し,
筋タン パク質 合 成系が低 下して いる,
プ レコ ンディショ ニ ング運 動 筋 (ExHS
) ではHSP72
mRNA の減少が抑制
さ れ て,
筋タン パク質合 成 系の 低下 が減 衰 して い る.
HSP72
mRNA は各サンプル の リボ ゾー
ム18S
mRNA で補正し,
正 常 筋 (Control
)を キャ リ ブ レー
タ と し て表示 し てある.
*,
CQntrol
に対 す る 有 意 差 (p
<O
.
05
>
;†,
HS
に対す
るExHS
の 有 意差 (p
<O
.
05
).
の活 性 化 を抑 制
でき
,
廃 用 性 萎
縮の進 行
を 予防
でき
るこ とを示 唆
し ている と考
え ら れる。
筋 萎 縮
は主
に筋
タンパ ク質分 解 系
の促
進 で 生 じる筋 線
維
の縮小
19)20)や筋 細 胞 核 断 裂
の結 果
で 生 じ る筋 線 維 数
の減 少
21−
23)に よっ ても
た ら さ れ る。
尾 部 懸 垂 に よ る廃
用性 萎 縮 筋
では尾 部 懸 垂 直 後
か らHSP70
の低
下 を伴
っ て タンパ ク質
の合 成
が抑 制
さ れ る こ と が報
告
さ れ ていJapanese Physical Therapy Association
NII-Electronic Library Service
Japanese Physioal Therapy Assooiation
尾部 懸垂前の運 動 が 廃 用 性 萎 縮 筋のユ ビキ チン
ー
プロ テ ア ソー
ム系に及ぼす 抑 制 効 果79
( 。{
8
鬢
。の \b
。ヨ
)蕩
蒼
脅
郎辱
蒻
鼠
芝
i50
lOO
50
0Control
HS
ExHS
図
3
筋原線 維タン パ ク質 (),lyofibrillar
protein
)量の変 化正常 筋 (
Control
)に 比較して,
萎 縮 筋 (HS
)の筋 原 線 維タ ン パ ク質量 (mg.
〆g
)は有 意に低下 し た,
HS
前に プレコ ンデ ィシ ョニ ング 運動 を行っ た群 (ExHS
)
で はControl
に比 較 して,
筋 原 線 維タ ン パ ク質量の低下 が み ら れ る が,
HS
に比較 して高 値 を 示した.
*,
ContrQl
に 対 す る 有 意 差 (p
く0
.
05
);†,
HS
に対 するExIIS
の有 意 差 (p
〈O
.
05
).
る。ま
た,Thomason
ら 1)2
)は尾 部 懸 垂
の数 時 間後
から
タ ンパ ク質 分 解 系
の活 性
化 が お こり
,
タン パ ク質 合 成
より分 解
の速
度 が 増 加 して,
筋 原 線維
タン パ ク質
の減 少
が 生 じ る こ とを 明 ら か に してい る。2
週 問
の尾 部 懸 垂
で は筋 線 維 数
の減 少
21−
23)も観 察
さ れ る が,
筋
タ ンパ ク質 分
解系
の促 進
が 主 体 に な る3)。
筋
タン パ ク質
の分 解
に関与
す
るも
の とし て,
リソ ソー
ム系
,
カル パイ
ン系
,
ユ ビ キ チンー
プロテア ソー
ム系
が知
ら れて い る。
リソ ソー
ム に は多
くのプロテ アー
ゼ が存 在
し,
数種 類
の カ テ プ シ ンが筋
原線 維
タンパ ク質
の分解
に関 与 し
ている と考 え
ら れ て い る3 )。
ま た,
カル シ ウ ム依 存 性
の プロテアー
ゼであ
る カ ル パ イン はZ
線の タン パ ク質
を分 離 す
ると考 え
ら れ る 3)24−
26 )。
これ ら はユ ビキ チ ン活 性 化
・
結 合
・
連 結 酵
素
に よ りユ ビキ チ ン化
さ れ,
ユ ビ キ チ ンー
プロテアソー
ム系
で 処 理 分解
さ れる3
)7)25
)。
Bodine
ら27)は萎
縮筋
で ユ ビキ チンー
プロ テ ア ソー
ム系 が 活 性 化
さ れ てい るこ とを 報 告
してい る。
ま た,Jones
ら28〕 は ヒ トの研 究
で2
週 間の不 動 化 に よる筋 萎 縮
でE3
リ ガー
ゼを含
めた タン パ ク質
分解 系
の酵 素 活 性
が 上昇 し
て い ること を報 告
し て い る。
本 研 究 で もユ ビ キ チン連 結 酵 素
であ
るユ ビ キ チ ン リガー
ゼE3
mRNA の測 定
を行
い,
萎 縮 筋
で は有
意 な増
加
が認
め ら れ た。タンパク
質
の新
生,
修
止等
の分 子
シ ャペ ロ ン作
用 を も つHSP72
が萎
縮筋
で減 少 す
る こと が多
く報告
さ れてい るn)6
)12)29
)。
Ku
らa〕は18
時 間
の尾 部 懸 垂
でヒラ メ筋
のHSP70
が減 少
してい る ことを確 認
し,
後 肢 懸 垂 直 後
か ら タンパ ク質
合 成が低
下す
る こ とを
示 し た。
Naito
ら12) は8H
問
の尾 部 懸 垂
でHSP72
が著
明 に減 少
し てい る こ と を報 告
している。ま
た,
Goto
ら
29 )や先 行
研究
f}にお い て も 尾部 懸
垂に よ る萎 縮 筋
で はHSP72
の減
少 が確 認
さ れてい る。本
研究
に おいても
HSP72
mRNA の低
ドが観 察
さ れており
,
萎 縮 筋
で は タンパ ク質 合 成
・
保護機 能
も低
下 して い る ことが示 唆
さ れ るc これ らの結 果
か ら,
タン パ ク質 分 解 系
の増 加
, お よ び タンパ ク質 合
成・
保 護
系
の低 下
に よる相 乗 効 果
で筋 萎 縮
が生
じ る もの と考
え ら れる。
そ
こ で,
廃 用 性 筋 萎 縮
を 予 防す
る方 法
と して,
尾 部 懸
垂 前
の プレコ ンディ ショ ニ ング 運 動 を 試み た。
運 動
はHSP72
を誘 導 す
る こ とが知
ら れて おり
6) 9’
, 10〕,
HSP72
が 筋 萎 縮 を 抑 制
す る こ と が示
さ れ て い る 6)12)。Febbraio
ら
30)はヒ ト を 用 い た研
究で運 動
がHSP72
mRNA を 増 加 させ るこ と を報 告
してい る。ま
た,
Naito
ら
12)は尾 部 懸 垂 前
に1
時 間
の温 熱 負 荷
を行 う
こ と でHSP72
が増 加
し,
8
日 間の尾部 懸
垂で の筋 萎 縮 を抑 制
す る 可能 性
を示
唆 してい る。HSP72
が 筋 萎 縮
の制 御
に 深く
関 わっ てい る と考
え,
プレコ ン デ ィショ ニ ング運動
で尾 部 懸 垂 す
る前
にHSP72
を誘 導
して筋 萎 縮
の抑 制
が で き ないか と考
え た。
そ
の結 果
,
筋 原 線 維
タ ンパ ク質 量
の低
下 を減 衰
さ せ るこ と が可 能
であ
る こ とを先 行 研 究
6〕 で報告
し た。
し か し,
先行研 究
で は萎 縮
予防
の制 御 機 構
ま で確 認す
るこ と ができな か
っ た。本研 究
で は プレコ ン ディショニ ング運 動
の予 防 効 果
の制御 機 構 と
して,
タン パ ク質
分解
系
のユ ビ キチ ンとタ ンパ ク質合 成
・
保 護 系のHSP72
に注
目 して検 証 を 行
った
。 その結
果, ユ ビキ チ ンE3
リ ガー
ゼ mRNA は萎 縮 筋
より
プレコ ン デ ィショ ニ ン グ 運動
を行
っ た骨 格 筋
で低
ドし,
タンパ ク 質 分 解の ユ ビキ チンー
プロテ ア ソー
ム系
の活 性 化
が抑
え ら れてい る こ と が検 証
でき
た。ま
た,
タ ンパ ク質 合 成
・
保 護
系
のHSP72
mRNA は萎 縮 筋
で低
ドしてい た が,
プレコ ンデ ィショニ ング運 動
で はHSP72
mRNA
の低
下 が抑
え ら れ てい ること が検 証
でき
た。HSP72
は分 子
シ ャ ペ ロ ン機
能
を もつ こと か ら変性 過 程
の タ ンパ ク質
と 結 合・
リブt
一
ルデ ィングし
,
機 能 型
タンパ ク質
へ修 復
を サ ポー
トS)す
るこ とで変性
タ ンパク質
の蓄 積 を抑 制
した と考
え ら れ る。
これ らの
結 果 か
ら,
2
週
間の尾部 懸 垂 前
のプ レコン デ ィショ ニ ング運動
は筋
タン パ ク質 分 解
の抑
制 と筋
タン パ ク質
合 成の低
下 を減 衰
させ る こ とが検
証で き た。
し か し,
本 研 究
で得
られ た 結 果
は2
週 間の 限 ら れ た 期間
での予 防
効 果
であ り
,
2
週 間 以
F.
の尾 部 懸 垂
の場 合
の変 化
な どに つ いても
,
今後
,
継 時
的 な 変 化 を 観 察 してい く 必 要 も あ る と考 え ら
れ る。 さ ら に 運動
強 度 やユ ビ キ チ ンー
プロテ ア ソー
ム系
やHSP72
な ど を 制 御
す る他
の負 荷 方 法
につ いても研 究
を進
め てい く必 要 性 が あ る と考 え ら れる。
謝 辞
:本 研 究
は文 部 科 学 省 科
学 研究
費
基 盤研 究 (
課 題
番 号
16300188
,
17300187
)
,
萌 芽 研 究(
課
題番 号
N工 工一
Eleotronio Library
so
-\utik\
17650171),
X4orfi
(t/'vevease15700384,
16700432),
thKtL-CXtuLk.
!
wt
1)
Thomason
DB,
Biggs
RB,
etal:Protein
metabelism andbeta-myosin
heavy-cbain
mRNAin
unweightcd soleusmuscle.
Am
J
Physiol
Regutatory
Tntegrative
Comp
Physiol
257:
30e-305,
1989.
2)
Thomason,
DB,
Booth
F"/:
iltrophy ofthe
soleuscle
by
hindlimb
unweighting,J
Appl
Physiol
68:
1-l2,
1990.
3)
Jackman
RW,
Kandarian
SC:
The
molecularbasis
ofskeletal mvscle atrophy.
Am
J
Physiol
Celt
Physiol
287:
C834-C843,
20e4.
4)
Ku
Z,
Yang
J.
et al./Decreased
polysomal
HSP-70
rnaystow
polypeptide
etengationduring
skeletal musclephy,
Am
J
Physiel
268:
C1369-C1374,
1995,
5)
Fujino
H,
Kohzuki
H,
et al:Regrcssien
of capi!tarywork
in
atrophied soteus muscleinduced
by
hindllmb
unweighting.
J
Appl
Physiol
98:
1407-1413,
2005.
6)
eewn-ptE,
l-.Hza.ifi
・
fik
:
7v
Feti-Bma.eut/]oVL.u
;ifI
tx
g =lf
p'meutth{zaM・[Cffifma
l・:Mt:';-'i
Mblk.
ge"
fii・kt}E
32(7):
400-405,
2005.
7)
Taillandier
D,
Combaret
L,
etal,:The
role ofproteasome-dependent
proLeolysis
in
the
remodelling ofskeletal muscle,
Proc
Nutr
Sec
63(2):
357-361,
2004.
8)
Beckmann
RP,
Mizzen
LE,
et al./Interaction
ofHSP
70
with newly synthesized
proteins/
implieutions
for
protein
folding
and assembly.Science
248{4957}:
850-854,
1990.
9)
Skidmore
R,
Gutierrez
JA,
et al.:HSP70
induction
in.or
exercise andhcat
stressin
rats:rele efinternal
perature,
Am
J
Physiol
268(1
Pt
2}:
R92-97,
I995,
10)
Milne
KJ,
Neble
EGi
Exercisc-induced
elevation orHSP70
is
intensity
depe]denl.
J
i'lpptPhysiol
93:
561-568,
2002.
11)
Neufer
PD,
Ordway
GA,
et at.:Continuous
contractilcactivit>,
induces
fiber
type
specific expression ofHSP70
in
skeletal musclc.Am
J
Physiol
Cell
Physiol
271:
1828-1837,
1996.
I2)
Naito
H,
Powers
SK,
et al:Heat
stress attenttatestalmuscle atrophy
in
hindlimb-unweighted
rats.J
Appl
Physiol
88:
359-363,
2000,
]3)
Uehara
K.
Goto
K,
et al:Heat-stress
enhances arive potentialin
rat soleus muscle.Jpn
J
Physiol
54:
263-271,
2004.
1・D
Morey
ER/
Spuceflight
andbone
turnover/
corretationwith a new rat model of weightlessness.
Bioscience
29/
168-172,
1979.
15)
Morey
ER,
Sabelman
EE,
et al:A
new rat modelutating some aspects of space
flight.
Physiologist
22:
S23-S24,
1979.
eg33tsca2e
16)
Chomczynski
P,
Sacchi
N:
Singie-step
method ofRNA
isolation
by
acidguanidinium
form
extraction.Anal
Biochcm
162:
156-159,
1987,
17J
I'Iildebrandt
AL,
Neufcr
PD:
Exercise
attenuaiesthe
ing-induced
transcriptional
activation of metaboliegenes
18)
19)
20)
20
22)
23)
24)
25)
26)
27)
28)
29)
30)
in
skeletal muscle.Am
J
Physiol
Endocrinol
Metab
278/
1078-1086,
2000.
Tsika
RW,
Herrick
RE,
et al.:Interaction
ef compen-satory overload undhindlimb
suspension on myosiniso-form
expression.J
Appl
Physiol
62:
2180-2186.
1987,
Desplanches
D,
Mayet
MII,
et al./Structural
andfunc-tional
responses toprolon.cred
hindlimb
suspensionin
ratmuscle.
J
Appl
Physiol
63:
558-563,
1987.
Templeton
GH,
Sweeney
HL,
et al,/Changes
in
fiber
composition of soleus muscle
during
rathindtimb
sus-pension.JAppl
Physiol
65:
1191-1195,
1988,
Siu
PM,
Pistilli
EE,
et al.:Apoptotic
respenses tohindlimb
suspensionin
gastrocnemius
musc]esfrom
yeung adult and aged rats,
ilm
J
Physiol
Regul
lntegr
Comp
Physiel
289:
RIO15-RI026,
2005.
Siu
PM,
PistMi
EE,
et al.:Aging
infiuences
cellular and molecu!ar responses of apoptosisto
skeletal muscle unloading.Arn
J
Physiol
Cell
Physiol
288:
C338-C349,
2005.I.eeuwenburgh
C,
Gurley
CM,
et al,:Age-related
differ-ences
in
apoptosis withdisuse
atrophyin
soteus muscle,Am
J
?hysiol
Regul
Intcgr
Comp
Physiol
288:
R1288-R1296,
2005.
Goll
DE,
Thompson
VF,
et al,:The
calpain system.Physiol
Rev
83/
731-SOI,
20e3.
Taillandier
D,
Aurousseau
E,
etal,/Coerdinate
activation oftvsosornal,
CaLi'-actix,ated
undATP-ubiquitin-depen-dent
proteinases
in
the
unweighted rat soleus muscle.Biochem
J
316:
65-72,
1996.
Witliams
AB,
Decourten-Myers
GM,
etal:Sepsis
stimu-lutes
release of myofi]amentsin
sketetal rnuscleby
a cal-cium-dependent mechanism.FASEB
J
13/
1435-14.t3,
1999,Bodine
SC,
Latres
E,
et al./ldentification
ol ubiquitin tig-ases requiredtor
skeletat musc]e atrephy.Science
294:
1704-1708,
2eOl.
Jones
SW.
Hill
RJ,
et al./Disuse
atrophy and exercise rehabilitatienin
humans
profoundly
affectsthe
expres-sion ofgenes
associated wiihthe
regulation of sketetal muscle mass.FASEB
J
18:
le25-1027,
2004.
Goto
K,
IIonda
M,
etal.:Heat
stressfacilitates
thc recos,-er}, of atrophied so]eus musclein
rat.Jpn
J
Physiol
54:
285-293,
2004.
Febbraio
MA,
Koukoulas
I:
HSP72
gene expressionpro-gressively
increascs
in
human
skeletal muscteduring
protonged,
exhaustive exercise,J
Appt
Pbysiol
89/
1055-1060,
2000.
Japanese Physical Therapy Association
NII-Electronic Library Service
JapanesePhysicalTherapy Association
EffB,fi.M.gBt'
a)
me
ru
DstesM・me
ifnrm
a)
Ltl
}f
)-f
p
fiL
7
v
-
Ak
l・:Ucg-l'
pm
fiU
rt
ve
<Abstract>
81
Inhibitory
Effects
ofExercise-preconditioning
before
Hindlimb
Suspension
on
Ubiquitin-proteasome
Pathway
Activity
in
Muscle
Atrophy
Hidemi
FUJINO,
RPT,
PhD
Fkieugty
ofIIbalth
Scienee,
Su2uka
U}ziversity
ofMedieal
Sciences
Institute
of7>'aditional
Mledicine,
Suxuka
Uhiversity
ofMbdicaJ
Sciences
Dqpartment
ofCardiovascular
Physiology,
Okayama
University
Graduate
School
ofMledicine,
Dentistr:y
andPharmaceutical
Sciences
Hisaharu
KOHZUKI,
MD,
PhD
Dqpartment
ofCardiovascular
Physiolqgy,
Okayama
U}ziversity
Graduate
School
ofMedicine,
Dentistr)r
andPharmaceutical
Sciences
Department
ofFbod
andNutrition,
Ohayama
Gahuin
Uhiversity
Isao
TAKEDA,
RPT,
PhD,
Hiromitsu
TASAKI,
RPT,
MS,
Hiroyo
KONDO,
MS
thcult3,
ofIIbalth
Science,
Suxuha
U}ziversity
ofMledical
Sciences
Torao
ISHIDA,
PhD
Institute
ofZ-aditional
Mbdieine,
Suzuka
University
ofMbdical
Sciences
Fumihiko
KAJIYA,
MD,
PhD
Dqpartment
ofCardiovascular
Pdysiolqgy,
Ok[ayama
Uhiversity
Graduate
School
ofMedicine,
Denttstr),
andPharmaeeutical
Scienees
Dqpartment
of
Mbdical
Engineering
Kdwasahi
Mbdical
School,
andKdwasaki
Collage
ofAlliedHealth
Proflessions