ラット膵フラグメントからのアミラーゼ放出にたいする細胞外2価陽イオンの影響
11
0
0
全文
(2) ^iSjt&WA^a^ (^2SBC) ^39^ Igl^ PSW63^W!. Journal of Hokkaido University of Education (Section II 0 Vol. 39, No. 1 October, 1988. Influence of Extracellular Divalent Cations on Amylase Release from Rat Pancreatic Fragments. Naoto ASADA Nutritional Physiology Laboratory, Asahikawa College, Hokkaido University of Education, Asahikawa 070. 7 -y YWv 7 y^VY^h^T ^ 7- -^?i^. ^i-^^^?^2W^^v^m m B ^ ^. 1iW^^.^±^W\9-K^^ikm^^. ABSTRAST The influence of extracellular divalent cations was examined in pancreatic lobule preparations of rats. Carbachol (1.07/^M) stimulated amylase release from the pancreatic lobules, and the rate of amylase release increased to 10-times the basal amylase release. Mn2+ solely stimulated amylase release, and the response to Mn2+ was transient. CCh-stimulated amylase release decreased in the presence of 5 mM Mn2+, and only 1 or 2.5 mM Mn2+induced a biphasic. increase in amylase release, a rapid transient phase and a delayed slowly rising phase. With 10 mM Mn2+, the rapid and transient phase of amylase release was not observed. Co2+ also. stimulated amylase release, but it was without any effect on CCh-stimulated amylase release. Amylase output in response to Co2+ increased with increasing Coz+ concentration. Both Mn2+. and Co2+probably increase the intracellular Ca2+concentration that activates the enzyme release from pancreatic acinar cells. Sr2+(2.5mM) was without effect on basal- and stimulated-. amylase release in the absence of Ca2+, but with small amounts of Ca2+, Sr2+(5 mM) inhibited CCh-stimulated amylase release transiently. Sr2+ does not substitute for Ca2+ in enzyme release from the pancreas.. INTRODUCTION Pancreatic acinar cells release zymogens in response to food ingestion by episodical exocytosis. The release of zymogens is physiologically controlled by the gut hormone. (7).
(3) 8. Naoto. ASADA. cholecystokinin-pancreozymin (CCK) and the neurotransmitter acetylcholine (ACh). It is commonly believed that the effect of these secretagogues is exerted by an increase in the cytoplasmic concentration of free, diffusable Ca2+. The primary role of the Ca2+ is to act as a 'second messenger' between the stimulation and the secretary process (stimulus-secretion coupling). (Douglas &Rubin, 1961). The effects of divalent cations in the exocrine pancreas hava been thoroughly investigated. Kanno & Nishimura (1976) ,Petersen & Ueda (1976), and Argent et al. (1982) reported that Mn2+ alone stimulate enzyme release and that secretagogue-stimulaed enzyme release is inhibited in. the presence of Mn2+. The stimulatory effect of Mn2+ was also observed in the parotid glands (Petersen et al. 1977). Co2+ also stimulates amylase release from the pancreas (Argent et al.. 1982). In adrenal medullary cells, it was reported that Sr2+can substitute for the Ca2+ that mediates ACh-stimulated catecholamine release (Douglas & Rubin, 1964), and both Mg2+ (over 5 mM) and Co2+ (1-2 mM) completely inhibits ACh-stimulated catecholamine release (Kidokoro et al. 1979). Using pancreatic lobule preparation, this study examines effects of Mn2+, Co2+, Sr2+, and. Mg2+ on amylase release. We confirmed the stimulatory effect of Mn2+ (Argent et al. 1982 ; Kanno & Nishimura, 1976 ; Petersen & Ueda, 1976 ; Petersen et al. 1977)and Co2+(Argent st al. 1982) in the absence of secretagogue. But our results indicate that high concentrations of Mn2+ (10 mM) blocks an immediate increase in amylase release as odserved by Argent et al. (1982). Sr2+ was without any effect on amylase release in response to CCh in the absence of Ca2+, could. not be substituted for Ca2+in the exocrine pancreas, and Mg2+ (5 mM) was also without any effect on CCh-stimulafed amylase release.. MATERIALS AND METHODS Pancreatic Lobules —— Sprague-Dawlay strain rats (200-350g) were fasted but allowed water for 20 h before each experiment. Rats were exsanguinated via the carotid under ether anesthesia, then t'he pancreas was quickly excised and immersed in ice-cold saline (154mM NaCl). The pancreatic lobules were prepared according to the procedure described by Scheele and Palade (1975). Samples of three or four lobules were preincubated for 30 min in 3 ml of oxygenated media at 37°C, then the lobules were rinsed and resuspended in fresh medium (5 ml) and incubated for 90 or 120 min. Every 10 min, 0.3 ml of the medium was collected, and the flasks with the media were gassed every 20 min throughout the incubation period. Solutions and Drugs — The standard incubation medium was a Krebs-Henseleit bicarbon-. ate buffer (KHB) of : 118 mM NaCl ; 4.7 mM KC1 ; Z.SmMCaClz ; 1.18mM MgClz ; 25 mM NaHCOa ; 1.18mM NaHzPO^ ; and 5.5 mM glucose. The medium was equilibrated with 95% Oz/5% COz. In some experiments the standard KHB was modified in a number of ways. In the Ca2+-free solution, CaClz was removed and 0.5 mM EGTA (ethylene glycol bis (yS-aminoetyl ether) -N, N, N', N'-tetraacetic acid) was added, CaClz was removed for the Ca2+-deficient. (8).
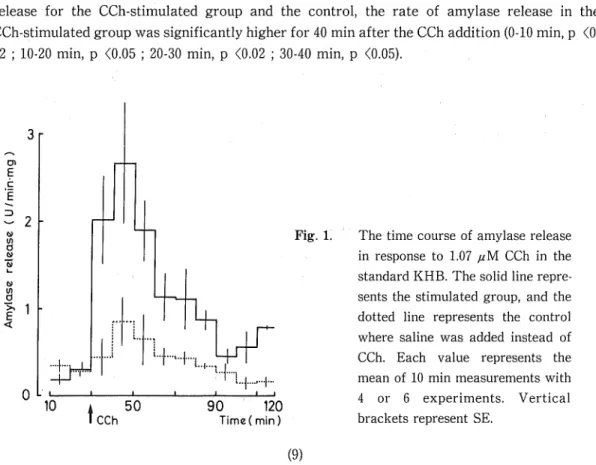
(4) Divalent Cations and Exocrine Pancreas 9. solution, NaHzP04 was replaced by NaCl in the NaH2P04-deficient solution. Carbachol (CCh) was dissolved in saline and 100 //I of CCh solution was added to the incubation media. Estimation of Amylase Release — Amylase activity in the collected media was assayed by modified Bernfeld (1955) method described by Kanno (1975). One unit of amylase activity was defined as the amount of enzyme which produced 1 mg of maltose during a 5 min incubation at 37 °C by the modified method. The wet weight of lobules was measured after the experiment, and the rate of amylase release to the medium was expressed as units per min per mg of tissue. (U/min/mg). Statistics — Results are expressed as the mean ± SE of several experiments, and are analyzed by Student's / -test.. RESULTS Time course of amylase release in a standard medium — The addition of CCh (1.07;uM) markedly increased the rate of amylase release. Until 30min after the CCh addition, the rate of CCh-stimulated amylase release was about 6-to 10-times that at the resting (before the CCh addition). The amylase release reached a maximum for 10-20 min after the CCh addition, and then gradually decreased (Fig. 1). Comparing the 10 min measurements of the rate of amylase release for the CCh-stimulated group and the control, the rate of amylase release in the. CCh-stimulated group was significantly higher for 40 min after the CCh addition (0-10 min, p <0. 02 ; 10-20 min, p <0.05 ; 20-30 min, p <0.02 ; 30-40 min, p <0.05).. 3r 01. E. c. 6. 32 N in a N u. w. 0. >*. <. Fig. 1. The time course of amylase release in response to 1.07 /<M CCh in the standard KHB. The solid line represents the stimulated group, and the. 1h. dotted line represents the control where saline was added instead of CCh. Each value represents the mean of 10 min measurements with. 10. CCh. 50. 4 or 6 experiments. Vertical. 90 120. brackets represent SE.. Time( min). (9).
(5) 10. Naoto ASADA. Effect of Mn2+ and Mgz+ — The pancreatic lobules were incubated in the NaHzP04deficient media, where NaH2P04 was replaced by NaCl to prevent precipitation with high concentrations of MnClz. Here MnCl2 or MgClzwas added (5 mM) 10 min before the CCh addition (0. 2^M) (Fig. 2A). The amylase release was stimulated solely by Mn2+, but CChstimulated amylase release was smaller than that of the controls. On the other hand, Mg2+ had no effect on amylase release alone, and the CCh-stimulated amylase release in the presence of. 5 mM Mg2+ was also similar to that of the control. The effect of Mnz+alone on amylase release was tested, and 1.0 and 2.5 mM Mn2+stimulated the amylase release (Fig. 2B). This Mn2+-. stimulated amylase release was biphasic, with a rapidly increasing transient phase and a delayed slowly rising phase. The rate of amylase release during 10 min after the Mn2+addition (rapidly increasing transient phase) was about 5 times that at resting. The rate of amylase release in the delayed phase was about 3 times that at resting at the end of the experiment. With 10 mM Mn2+, amylase release was little stimulated during the first 10 min, but from 10 min after the Mn2+ addition, the rate of amylase release gradually increased through the experimental period.. B. 3r 01. E c. E. 6> Ifl. d N. -L >>. !. D^-J--J. <. 0L. 10. Fig. 2.. ii-L-i I !. -J—. L-L--. 11. 50 CCh. 90. Time (min). 0. 10. 'fn. 50. 90. Time(min). Time course of amylase release stimulated by 0.2 /^M CCh in the presence of Mn2+ or Mg2+ (A), stimulated by various concentrations of Mn (B). A : Pancreatic lobules were incubated in NaHzP04 -deficient KHB, and MnClz (5 mM) (—) or MgClz (5 mM) (-----) was added at M. The thin line repersents the time course of amylase release in the control in which saline was added instead of metal ions. Each value represents the mean from 5 experiments. The vertical bars represent 1 SE. B : Pancreatic lobules were incubated in the NaHzPOi -deficient KHB, and MnCla ( 1 mM, ----- ,2.5 mM, ••••••; 10 mM, — ) was added at Mn. Each value. represents the mean from 4 experiments. Vertical bars represent 1 SE.. (10).
(6) 11. Divalent Cations and Exocrine Pancreas. B. 3r. 01. E c. I2. =>. w^. &). 8N 6). L.. &> 1 1/> 0. 10 f ^ 50 M "CCh. 90. Timiz(min). 0. ^H.-..i. 10. -}-^. L4-. f 50. Co. 90. Time (min). Fig. 3. Time course of amylase release stimulated by 0.2 jnM CCh in the presence of Co2+ (A) and by various concentrations of Coz+ (B). A : Pancreatic lobules were incubated in the Ca24- -free KHB, and CaClz (2.5 mM) + CoCla (2.5 mM) (—) or CaClz + saline (-----) was added at M. Each value represents the mean from 6 or 7 experi-. ments. Vertical brackets represent SE. B : Pancreatic lobules were incubated in the NaH^PO, -deficient KHB, and Cod, ( 1 mM, -----; 2.5 mM, ••••••; 10 mM, —). wad added at Co. Each value represents the mean from 4 experiments. Vertical brackets represent SE.. Effect of Co2+ — Co2+ (2. 5mM) stimulated amylase release from the pancreatic lobules in the Ca2+-free media without CCh (Fig. 3A). The increase iti amylase release was significant (p <0, 01). But CCh (0. 2//M)-stimulated amylase release was not affected by the presence of Co2+. The total CCh-stimulated amylase output in the standard KHB (2. 5 mM Ca2+) was 58. 69 ±2.95 U/40 min/mg. The total amylase output in the Ca2+-free KHB with 2.5 mM Co2+ and without Co2+ was 46.37 ±5.58 U/40 min/mg and 47.90 ±5.96 U/40 min/mg. To investigate the effect of Co2+ alone, 1.0, 2.5, and 10 mM Co24- was tested in the NaH2P04-deficient KHB (Fig.. 3B). The amylase release was stimulated with all three concentrations of Co2+. The total amylase output with 1,0 mM Co24- was 26.65 ±2.82 U/20 min/mg, with 2.5 mM 38.1 ±3.93 U/ 20 min/mg and with 10 mM Co2+ it was 40.78 ±2.74 U/20 min/mg. The increase in amylase output stimulated by 2.5 mM or 10 mM Co2+ was significantly. (11).
(7) 12. Naoto ASADA. higher than with ImM Co24- (p <0.05, p <0.01). The effect of 10 mM Co2+ on amylase release in the Ca2+-free media was also tested (Fig. 4). Basal amylase release was relatively low in the Ca2+-free KHB, and the Co2+-stimulated amylase release was more transient in the Ca2+-free. KHB. The decrease in amylase release 10-20 min after the Co2+ addition in the absence of Ca2+ was statistically significant (p <0.01).. 3r en. E. c. Fig. 4. Time course of amylase release. E. stimulated by 10 mM Co2+ in the. 3 2. presence (—) or absence of Ca2+ (-----). Pancreatic lobules. were incubated in the standard KHB or in the Ca2+ -free KHB,. N U) a. t. 4-;. <. =-K 0L. and CoClz (10 mM) was added at Co. Each value represents the. ih-. mean from 6 or 7 experiments.. p-l--!. Vertical brackets represent SE.. 10. t 50. Co. 90. Time (min). Effect of Sr2+ — The pancreatic lobules were incubated in the Ca2+-free media, with no CaCl^and 0.5 mM EGTA, and SrClz was added (2.5mM) 10 min before the CCh addition (0.2//M), The CCh-stimulated amylase release in the presence of Sr2+ was not different from that in the absence of Sr2+ (Fig, 5A). But the amylase release in response to CCh (0.2 /<M) in the presence of Srz+ was much smaller than that in the presence of Ca2+ (standard KHB). From this experiment, Sr2+ appeared to inhibit the stimulated amylase release. Next the pancreatic lobules were stimulated by CCh (0.2 //M) in the Ca2+-deficient KHB (on CaClz and no EGTA), and 20 min later Sr2+ (5mM) or Ca2+ (2. 5mM) was added (Fig. 5B). The stimulated amylase release was strongly inhibited by the addition of Sr2+, while the amylase release was maintained by the additional Ca2+. This inhibition by Sr2+ was significant (p <0. 01).. (12).
(8) 13. Divalent Cations and Exocrine Pancreas. B en. E. c 2. E 6). 8N. £1 &>. 3 OL. 10. Fig. 5.. t f^,50. M. 'CCh. 90. Time (min). 10. 50 ) 90. ICCh M Time (min). Time course of amylase release stimulated by 0.2 /<M CCh in the presence of Srz+ added before (A) or after (B), the CCh addition. Effect of Srz+ on amylase release in response to 0.2 ;uM CCh. A : Pancreatic lobules were incubated in the Ca2+ -free KHB, and SrCl, (2.5 mM) (—) or saline (-----) was added at M. The thin line represents the time course of amylase release, in the standard KHB. Each value represents the mean from 6 or 7 experiments. Vertical brackets represent SE. B : Pancreatic lobules were incubated in the Caz+ -deficient KHB, and CaClz (2.5 mM) (—) or SrClz (5 mM) (••••••) was added at M. Each value represents the mean from 4 experiments. Vertical brackets represent SE.. DISCUSSION Amylase release stimulated by CCh from pancreatic fragments — We used the pancreatic lobule preparation to investigate the amylase release from pancreatic acinar cells. In this preparation one fragment is a separated lobule which was removed by excision with fine scissors (Scheele and Palade, 1975). This pancreatic lobule releases zymogens in response to secretagogues e. g, CCK and carbachol (synthetic acetylcholine) that are added to the incubation media. Amylase release from the pancreatic lobules increased with more than 0. 03 /<M, and reached a maximum at 3.3 /<M. When the lobules were incubated for 40min with 3.3 /^M CCh, about 30% of the initial amount of amylase was released (data not shown). In the present experiment, we collected an aliquot of the incubation media every 10 min and followed the time course of amylase release. Submaximal concentration of CCh (1.07 //M) induced rapid increases in amylase release (Fig. 1), but the rate of amylase release reached maximum during 10-20 min after the CCh addition. In the isolated perfused pancreas of rats, a more sensitive preparation,. (13).
(9) 14. Naoto. ASADA. pancreatic juice and amylase release in response to ACh (0.1 /^M) were maximal in the first 5 min of continuous stimulation (Habara, 1980 ; Kanno et al. 1983). CCh-stimulated amylase release from the pancreatie lobules appeared to be biphasic, a relatively high response phase in the first 30 min followed by a relatively low response phase. In this experiment CCh was added to the incubation media only once, the CCh concentration probably decreased gradually and the true concentration of CCh at a particular time was unknown. Why the time course of amylase release from the pancreatic lobules was biphasic is unclear.. Effect of Mnt+ —— Mnz+ (1 or 2.5 mM) stimulated amylase release biphasically without CCh, first a rapidly increasing, transient phase and next a delayed, slowly rising phase. Stimulation by Mn2+ alone was previously reported for amylase release from the pancreas. (Kanno & Nishimura, 1976 ; Petersen & Ueda, 1976 ; Argent et al. 1982) and from the parotid (Petersen et al. 1977) and the biphasic effect of Mn2+ was observed by Argent et al. (1982) in the pancreas and by Petersen et al. (1976) in the parotid. The stimulation by Mn2+ is not due to released ACh from vagal nerve terminals in the lobules (see Kanno & Nishimura, 1976 ; Argent et al. 1982) but due to lysis of acinar cells (see Argent et al. 1982). The present results confirmed these results, but with 10 mM Mnz+ we did not find a rapid, transient phase as reported for the biphasic effect on amylase release by Argent et al. (1982). In the parotid (Petersen et al. 1977) the stimulation with 10 mM Mn2+ was no more effective than that with 5 mM Mn2+. The source of the Ca2+ that mediates Mn2+-stimulated amylase release was unclear in our results, because experiments in Ca2+-deprived media were not performed. The source of Ca2+ in Mn2+-stimulated amylase release is obscure and conflicting results have been reported :. Stimulation by Mn2+ in the pancreatic fragment did not occur in Caz+-free media (with 1 mM EGTA) (Argent et al. 1982) ; the only rapid phase of amylase release from the parotid fragments disappeared in Ca2+-free media (with 2 mM EGTA) (Petersen et al. 1976) ; the delayed phase in isolated perfused pancreas was observed in Ca2+-deficient (no EGTA) (Kanno & Nishimura, 1976) ; or the stimulatory effect in the segments of pancreas was only observed in Ca2+-and Mg2+-free media (Petersen & Ueda, 1976). When Mnz+ was added 10 min before the CCh addition, CCh-stimulated rapid amylase release was not observed. This confirms previous observations by Heisler et al. (1972) and Kanno & Nishimura (1976) but is contrary to those of Petersen & Ueda (1976). The inhibitory effect of Mn2+ on secretagogue-stimulated amylase release has been explained by a simple. competitive interaction with Ca2+ (Kanno & Nishimura, 1976) or by action st some distal site within the secretary, process, e. g. fusion of the bilayer membrane (Argent et al. 1982). Our. results, provided no further information of the inhibitory effect. Further experiments are required to clarify the effect of Mn2+ alone and with a secretagogue. Effect of Co2+ and Mg2+ — It has been reported that Co2+ (Kidokoro et al. 1979) or Mg2+ (Douglas & Rubin, 1964) inhibits catecholamine release from the adrenal medulla and that these ions block the voltage-dependent Ca2+ channels. But in the exocrine pancreas, Co2+ and Mg2+. (14).
(10) Divalent Cations and Exocrine Pancreas 15 were without any effect on CCh-stimulated amylase release on these experiments. Pancreatic. acinar cells hyperpolarized in response to CCK in the isolated perfused pancreas (Kanno, 1972 ; Kanno ; 1985), therefore the acinar cells are not considered to have voltage-dependent Ca2+ channels. The present result, that Co2+ stimulated amylase release alone in a dose-dependent manner. and that the stimulation by Co2+ partially depends on extracellular Caz+ confirmed previous observations made by Argent et al. (1982). A similar conclusion was reached by Mitchell et al. (1980) for the cockroach salivary gland. In our results, Co2+ -stimulated amylase release during the first 10 min was not affected by Ca2+ deprivation, but it might be due to ACh release from vagal nerve terminals that were partly stimulated by Co2+.. Effect of 5r2+ — Sr2+ has been reported to substitute for Ca2+ in the adrenal medulla (Douglas & Rubin, 1964). But Sr2+ failed to substitute for Caz+ in the exocrine pancreas, and had no effect on CCh-stimulated amylase release in the absence of Ca2+ in the present experiments.. When the CCh stimulation was followed by Sr2+ addition, Sr2+ completely inhibited the CCh-stimulated amylase release that could be maintained by Ca2+ addition. The same inhibition was observed in the absence of Ca2+ (data not shown). The inhibitory mechanism of Sr2+ on CCh-stimulated amylase release is not clear from the present results, and further experiments are required.. I would like to thank Miss Yukiko Igarashi, Miss Toshiko Ooi, Mrs. Rie Seo (Ohno), and Miss Kumiko Saruta for their valuable assistance in these experiments.. REFERENCES Argent, B. E., Case, R. M. & Hirst, F. C. (1982) The effects of manganese, cobalt and calcium on amylase secretion and calcium homeostasis in rat pancreas. /. Physiol. 323, 353-375.. Bernfeld, P. (1955) Amylase a and /?. In : Methods in Enzymology vol. 1, Ed. by Colowick, S. P. and Kaplan, N. 0. pp. 149-158. Academic Press, New York.. Douglas, W. W. & Rubin, R. P. (1961) The role of calcium in the secretary response of the adrenal medulla to acetylcholine. /. Physiol. 159, 40-57. Douglas, W. W. & Rubin, R. P. (1964) The effects of alkaline earths and other divalent cations on adrenal medullary secretion. /. Physiol. 175, 231-234.. Habara, Y. (1980) Influences of ionic environments on ACh-induced secretary responses in isolated perfused pancreas of rats. Jpn. J. Pkysiol. 30, 561-574.. Heisler, S., Fast, D. & Tenenhous, A. (1972) Role of Ca2+ and cyclic AMP in protein secretion from rat exocrine pancreas. Biochim. Biophys. Acta 279, 561-572.. Kanno, T. (1972) Calcium-dependent amylase release and electrophysiological measurements in cells of the pancreas. /. Physiol. 226, 353-371.. Kanno, T. (1975) The electrogenic sodium pump in the hyperpolarization and secretary effects. (15).
(11) 16. Naoto. ASADA. of pancreozymin in the pancreatic acinar cell. /. Physiol. 245, 599-616.. Kanno, T. (1985) Electrophysiology and stimulus-secretion coupling of the pancreatic acinar cell. In The electropkysiology of the secretary cell, Ed. by Poisner, A. M. & Trifaro, J. M. pp. 3-24. Elsevier, Amsterdam/New York/Oxford. Kanno, T. & Nishimura, 0. (1976) Stimulus-secretion coupling in pancreatic acinar cells :. inhibitory effects of calcium removal and manganese addition on pancreozymin-induced amylase release. /, Physiol. 257, 309-324.. Kanno, T., Saito, A. & Ikei, N. (1983) Dose-dependent effects of acetylcholine stimulating respiratory chain and secretion of isolated perfused rat pancreas. Biomedical Research 4, 175-186.. Kidokoro, Y., Ritchie, A. K. & Hagiwara, S. (1979) Effect of tetrodotoxin on adrenaline secretion in the perfused rat adrenal medalla. Nature, Land. 279, 63-65. Mitchell, M. R., Ginsborg, B. L. & House, C. R. (1980) A novel effect of cobalt treatment on calcium-dependent responses of the cockroach salivary gland. Experimentia 36, 98-99. • Petersen, 0. H. & Ueda, N. (1976) Pancreatic acinar cells: the role of calcium in stimulussecretion coupling. /. Physiol. 254, 583-606. Petersen, 0. H. & Ueda, N. , Hall, R. A. & Gray, T. A. (1977) The role of calcium in parotid amylase secretion evoked by excitation of cholinergic, a- and /?-adrenergic receptors.. Pfliigers Arch. 372, 231-237. Scheele, G. E. & Palade, G. E. (1975) Studies on the guinea pig pancreas. /. Biol. Chem. 250, 2660-2670.. (16).
(12)
図


関連したドキュメント
In particular, we consider a reverse Lee decomposition for the deformation gra- dient and we choose an appropriate state space in which one of the variables, characterizing the
We use these to show that a segmentation approach to the EIT inverse problem has a unique solution in a suitable space using a fixed point
For instance, Racke & Zheng [21] show the existence and uniqueness of a global solution to the Cahn-Hilliard equation with dynamic boundary conditions, and later Pruss, Racke
Inside this class, we identify a new subclass of Liouvillian integrable systems, under suitable conditions such Liouvillian integrable systems can have at most one limit cycle, and
We present sufficient conditions for the existence of solutions to Neu- mann and periodic boundary-value problems for some class of quasilinear ordinary differential equations.. We
In this paper, we extend this method to the homogenization in domains with holes, introducing the unfolding operator for functions defined on periodically perforated do- mains as
inter-universal Teichm¨ uller theory, punctured elliptic curve, number field, mono-complex, ´ etale theta function, 6-torsion points, height, explicit esti- mate, effective
“Breuil-M´ezard conjecture and modularity lifting for potentially semistable deformations after
