Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬科学) 報 告 番 号 甲第1645号 学 位 記 番 号 第328号 氏 名 山城 貴弘 授 与 年 月 日 平成 30 年 3 月 26 日 学位論文の題名 ヒト PCFT の葉酸輸送機能に対する myricetin の持続性阻害作用の分子機 構 論文審査担当者 主査: 林 秀敏 副査: 湯浅 博昭, 頭金 正博, 岩尾 岳洋
名古屋市立大学学位論文
ヒト
PCFT の葉酸輸送機能に対する
myricetin の持続性阻害作用の分子機構
平成
29 年度(2018 年 3 月)
名古屋市立大学大学院薬学研究科
薬物動態制御学分野
山城 貴弘
名古屋市立大学学位論文
ヒト
PCFT の葉酸輸送機能に対する
myricetin の持続性阻害作用の分子機構
名古屋市立大学大学院薬学研究科
薬物動態制御学分野
山城 貴弘
Molecular Mechanism of Sustained Inhibitory Effect of Myricetin on Folate Transport
Function of Human PCFT
Department of Biopharmaceutics
Graduate School of Pharmaceutical Sciences, Nagoya City University
一.本論文は、2018 年 3 月、名古屋市立大学大学院薬学研究科において審査されたもので ある。 主査 林 秀敏 教授 副査 湯浅 博昭 教授 頭金 正博 教授 岩尾 岳洋 准教授 二.本論文は、学術雑誌に収載された次の論文を基礎とするものである。 (基礎となる報文)
1. Takahiro Yamashiro, Kinya Ohta, Katsuhisa Inoue, Mai Furumiya, Yayoi Hayashi, Hiroaki Yuasa Kinetic and time-dependent features of sustained inhibitory effect of myricetin on folate transport by proton-coupled folate transporter.
Drug Metabolism and Pharmacokinetics, 30, 341-346, 2015.
2. Takahiro Yamashiro, Tomoya Yasujima, Kinya Ohta, Katsuhisa Inoue, Hiroaki Yuasa
Specific inhibitory effects of myricetin on human proton-coupled folate transporter: comparison with its effects on rat proton-coupled folate transporter and human riboflavin transporter 3. Drug Metabolism and Pharmacokinetics, 32, 311-314, 2017.
3. Takahiro Yamashiro, Tomoya Yasujima, Kinya Ohta, Katsuhisa Inoue, Hiroaki Yuasa
Identification of the amino acid residue responsible for the flavonoid sensitivity of human proton-coupled folate transporter.
Manuscript in preparation
(参考論文)
1. Mai Furumiya, Katsuhisa Inoue, Chihiro Nishijima, Takahiro Yamashiro, Erina Inaoka, Kinya Ohta, Yayoi Hayashi, Hiroaki Yuasa
Noncompetitive inhibition of proton-coupled folate transporter by myricetin. Drug Metabolism and Pharmacokinetics, 29, 312-316, 2014.
2. Mai Furumiya, Takahiro Yamashiro, Katsuhisa Inoue, Chihiro Nishijima, Kinya Ohta, Yayoi Hayashi, Hiroaki Yuasa
Sustained inhibition of proton-coupled folate transporter by myricetin. Drug Metabolism and Pharmacokinetics. 30, 154-159, 2015.
三.本論文の基礎となる研究は、湯浅 博昭 教授の指導の下に名古屋市立大学大学院薬学 研究科において行われた。
本論文では、以下の略号を用いた。
DMEM Dulbecco's modified Eagle's medium FBS fetal bovine serum
HEK293 human embryonic kidney 293 cell line IBMX 3-isobutyl-1-methylxanthine
MDBK Madin-Darby bovine kidney cell line MDCKII Madin Darby canine kidney II cell line OATP organic anion transporting polypeptide PBS phosphate buffered saline
PCFT proton-coupled folate transporter PCR polymerase chain reaction PMA phorbol 12-myristate 13-acetate SDS sodium dodecyl sulfate
S.E. standard error SLC solute carrier
目次
序論 1 本論 2 第1 章 PCFT に対する myricetin の持続性阻害作用の解析 1 myricetin の前処理時間依存的な阻害作用 2 2 myricetin 前処理による阻害作用の持続性 3 3 myricetin の濃度依存的な阻害作用 4 4 速度論的解析 55 PCFT による folate 取り込みに及ぼす protein kinase modulator の影響 7
6 まとめ 9 第2 章 myricetin の持続性阻害作用のヒト PCFT 特異性 1 myricetin の即時性阻害作用 10 2 myricetin の持続性阻害作用 12 3 まとめ 14 第3 章 PCFT の myricetin 感受性に関わるアミノ酸残基の同定 1 hPCFT-rPCFT 間でのキメラ体を用いた解析 16 2 部位特異的変異導入によるhPCFT の myricetin 感受性アミノ酸残基の同定 18 3 hPCFT の G158 の各種アミノ酸による置換の影響 21 4 各動物種におけるPCFT の myricetin 感受性種差 24 5 まとめ 26 結論 27 実験の部 I 実験材料 1 試薬 28 2 培養細胞 28 II 実験方法
1 各種トランスポーターのクローニング 28 1-1 human proton coupled folate transporter(hPCFT) 28 1-2 rat proton coupled folate transporter(rPCFT) 29 1-3 human riboflavin transporter 3(hRFVT3) 29
1-4 hPCFT-rPCFT 間のキメラ体 29
1-5 hPCFT 及び rPCFT の変異体 30
1-6 african green monkey proton coupled folate transporter(agmPCFT) 32 1-7 bovine proton coupled folate transporter(bPCFT) 33 1-8 mouse proton coupled folate transporter(mPCFT) 33
2 細胞の培養方法 33 3 遺伝子導入 34 3-1 安定発現系の作製:MDCKII 細胞 34 3-2 一過性発現系の作製:HEK293 細胞 34 4 取り込み実験 34 III データ解析 1 取り込みデータ処理 35 2 統計処理 36 謝辞 37 参考文献 38
1
序論
葉酸(folate, vitamin B9)は水溶性ビタミンの一種であり、核酸合成やアミノ酸代謝にお いて 1 炭素転移反応の補酵素として働き、生体の恒常性の維持に重要な役割を果たしてい る。また、folate は哺乳類の生体内では合成されないため、その供給は食物中の folate に依 存している。他の水溶性ビタミンと同様に、その効率的な摂取のため、腸管吸収にはトラン スポーターが関与すると考えられてきたが、近年、H+依存的にfolate を輸送するトランスポーターであるproton-coupled folate transporter(PCFT/SLC46A1)が同定され1)、PCFT が腸管
でのfolate 吸収を担うことやその機能特性などが明らかとなってきている2,3)。 一方で、アルコール飲料の連用摂取が葉酸欠乏につながる危険因子として知られており 4-6)、巨赤芽球性貧血や神経障害などの疾患を惹起するとされている7,8)。また、近年、フラボ ノイドや植物由来成分がトランスポーターや代謝酵素に対して影響を及ぼす例が、関心を 集めてきている9-17)。アルコール飲料中に含まれているフラボノイド類の葉酸吸収不良への 関与も疑われるところであり、また、folate と同様に PCFT を介して吸収される葉酸拮抗薬 の腸管吸収へのフラボノイド類の影響も予想される 18,19)。しかし、情報は依然として乏し く、その詳細なメカニズムも明らかとなっていない。葉酸欠乏への対処や葉酸拮抗薬の使用 における最適化調節のために、その分子機構の解明が望まれるところである。 このような状況のもと、その種の問題を引き起こし得るフラボノイドの一つとみられる myricetin について、ヒト PCFT(hPCFT)に対する即時性及び持続性の阻害作用が見出され た 20,21)。当研究室での最近の研究において、folate を含む試験液中に添加された myricetin (100 µM)が作用することによる即時性の阻害効果は、非競合阻害の様式を示し、hPCFT に よるfolate の輸送を約 50%低下させた。一方、細胞を myricetin で前処理することにより惹 起され、その除去後も持続する持続性の阻害効果については、100 µM の myricetin での 60 min 間の前処理により、hPCFT による folate 輸送の最大輸送速度(Vmax)が約90%低下した。
また、この時、ミカエリス定数(Km)の低下も生じていることが見出された。さらに、この 阻害作用はhPCFT の発現量及び細胞膜局在の低下によるものではないこと、myricetin 類縁 の一部のフラボノイド類も同様の作用を示すこと等が明らかとなってきているが、その阻 害機構等に関して、依然として不明な点が多いままである。 hPCFT に対する myricetin 類の持続性阻害効果は、作用が強く、また持続性である点から、 folate 類の腸管吸収への影響が特に大きいとみられる。そこで、本研究では、その特性の把 握を進展させ、また阻害機構の解明を図るため、阻害フラボノイドとしてmyricetin を用い、 阻害の惹起及び持続の過程の評価を含めた輸送機構論的解析に取り組んだ。さらに、その中 で、分子的及び機能的にhPCFT と極めて類似しているラット PCFT(rPCFT)が myricetin に よる阻害を受けないことが見出されたことを受け、両者の差異を利用した比較解析により、 hPCFT の myricetin 感受性に関わるアミノ酸残基の同定に取り組んだ。
4 3 myricetin の濃度依存的な阻害作用 myricetin での前処理による阻害作用の濃度依存性について検討した。その結果、阻害作用 は濃度依存的であり、20 µM から阻害作用が認められた(Fig. 3)。また、100 µM では、阻 害率が80%を超える強い阻害がみられた。実際の腸管腔内での myricetin の濃度は不明であ るが、ワイン中に含まれるmyricetin は 10-100 µM 程度と報告されている24,25)。本研究で用 いたmyricetin の濃度は、その範囲内に有り、腸管腔内で葉酸吸収不良を引き起こす要因の 一つであり得ると考えられる。 Myricetin concentration (M) 1 10 100 U ptak e rate (% o f co nt ro l) 0 20 40 60 80 100 120 * * * 0
Fig. 3. Effect of Myricetin on the Uptake of Folate by hPCFT Stably Expressed in MDCKII Cells: Dependence on Myricetin Concentration
Mean ± S.E. (n = 4); specific uptake of [3H]folate (5 µM) for 2 min at 37°C and pH 5.5;
pretreatment in buffer for 60 min at pH 5.5 and 37°C in the presence of myricetin or in its absence (control); control, 83.0 pmol/min/mg protein; * p < 0.05.
5 4 速度論的解析 速度論的観点からの阻害メカニズム解析を試みた。Michaelis-Menten 型の担体輸送を仮定 したモデルを用いた当てはめ計算を行った結果、myricetin の濃度上昇に伴い最大輸送速度 (Vmax)が低下し、阻害作用の要因となっていることが明らかとなった。しかし一方で、ミ カエリス定数(Km)の低下がみられ、folate に対する PCFT の親和性が上昇していることが 示唆された。また、このため Kmを大きく下回る低基質濃度での輸送活性を表す Vmax/Kmに 大きな変化はみられなかった(Fig. 4, Table 1)。これらの結果は、GFP 付加体を用いた先の 研究の結果とほぼ一致していた21)。本研究では、非修飾のPCFT に関して、これらの特徴を 確認すると同時に、myricetin 濃度に依存したパラメーター変化が生じていることを見出す ことができた。なお先の研究では、western blot 解析による GFP の検出及び GFP からの蛍光 の観察により、PCFT の発現量及び細胞膜局在には、myricetin 前処理による変化はみられて いない。よって本研究においても、PCFT の細胞膜局在への影響はなく、可逆性のメカニズ ムによるPCFT 分子の修飾により、最大輸送速度の低下と同時に親和性の上昇を伴う機能変 化を生じているものと推察された。
7
5 PCFT による folate 取り込みに及ぼす protein kinase modulator の影響
速度論的解析により PCFT の分子修飾の可能性が考えられたことやフラボノイド類が
様々なprotein kinase に対して影響を与える例が報告されていることから26)、myricetin によ
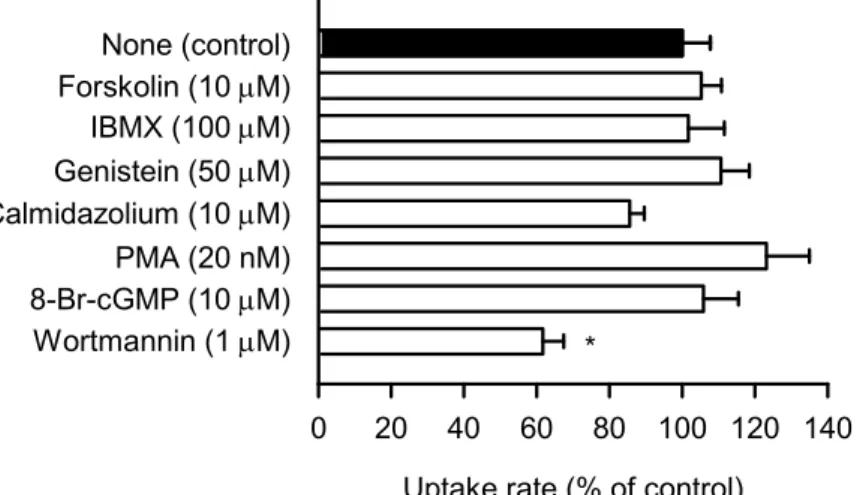
る阻害作用のメカニズムを探る試みとして、protein kinase modulator による前処理の影響を 検討した。その結果、PI3K(phosphatidylinositol 3-kinase)の阻害剤である wortmannin(1 µM) で、myricetin と比較するとやや弱いながらも、40%程度の阻害が認められた(Fig. 5)。
Uptake rate (% of control)
0 20 40 60 80 100 120 140 None (control) Forskolin (10 M) IBMX (100 M) Genistein (50 M) Calmidazolium (10 M) PMA (20 nM) 8-Br-cGMP (10 M) Wortmannin (1 M) *
Fig. 5. Effect of Protein Kinase Modulators on the Uptake of Folate by hPCFT Stably Expressed in MDCKII Cells
Mean ± S.E. (n = 4); specific uptake of [3H]folate (5 µM) for 2 min at 37°C and pH 5.5;
pretreatment in buffer for 60 min at pH 5.5 and 37°C in the presence of reagent or in its absence (control); control, 44.4 pmol/min/mg protein; * p < 0.05.
myricetin での前処理による PCFT 阻害の効果は、Vmaxの低下によって現れるものである。 一方で、Kmも同程度に低下するため、Vmax/Kmへの影響はほとんどない。このため、Vmax相 当の輸送速度でfolate が取り込まれる高 folate 濃度(5 µM 程度以上)で阻害作用が顕著と なる一方で、Vmax/Kmを輸送係数として、folate 濃度に比例した速度で取り込まれる線形領域 の低folate 濃度(Kmを大きく下回る濃度域)での阻害作用はほとんどない。ここで見出さ れたwortmannin の効果は、myricetin での前処理の阻害作用を検出し易い、標準的な試験条 件として用いている高 folate 濃度(5 µM)でのものである。したがって、wortmannin は myricetin と同様に Vmaxの低下を引き起こしているものと考えられる。 さらに、wortmannin の阻害作用の特徴を明らかにするため、wortmannin 濃度の影響を検 討すると共に、低folate 濃度(5 nM)での folate 取り込みへの wortmannin の影響も検討する ことにした。その結果、myricetin の場合と同様に、低 folate 濃度では有意な阻害作用は認め られなかったことから(Table 2)、Vmaxの低下と同時に Kmも低下していると考えられる。し
8 かし、wortmannin 濃度を上昇させても、高 folate 濃度(5 µM)での阻害率は、40%程度のま まで変化がなく、myricetin の場合にみられた 80%を超える阻害を惹起することはできなか った。したがって、wortmannin による阻害は、myricetin による阻害と共通する特徴はある が、完全には一致しないと考えられる。wortmannin については、PI3K の阻害剤であるので、 PI3K の関与するリン酸化反応あるいは関連の経路の阻害により PCFT の修飾を引き起こし、 回転速度の低下及び親和性の上昇を生じている可能性が考えられる。したがって、myricetin については、同様のメカニズムが部分的に関与している可能性を否定できないが、一方で、 別のメカニズムの関与も推察される。
Table 2. Effect of Wortmannin Concentration for Pretreatment on the Uptake of Folate by PCFT Wortmannin concentration (µM) Uptake rate (% of control) Folate (5 nM) Folate (5 µM) 0 (control) 100.0 ± 4.2 100.0 ± 9.9 1 86.7 ± 3.3 63.4 ± 1.5* 2 86.2 ± 4.6 60.6 ± 2.8* 5 84.7 ± 5.0 59.6 ± 5.4*
Mean ± S.E. (n = 4); specific uptake of [3H]folate (5 nM or 5 µM) for 2 min at 37°C and pH
5.5; pretreatment in buffer for 60 min at pH 5.5 and 37°C in the presence of reagent or in its absence (control); control, 366 fmol/min/mg protein and 62.9 pmol/min/mg protein, respectively, for 5 nM and 5 µM; * p < 0.05.
9 6 まとめ
ヒトPCFT 安定発現 MDCKII 細胞において、PCFT による folate 取り込みに対する myricetin (50 µM)の持続性阻害作用は、5 µM の folate 濃度において、前処理開始後 10 min の時点 から認められた。阻害は前処理時間依存的に増強され、60 min の時点で、ほぼ最大レベルの 阻害率(約80%)に達した。前処理の終了後には、この阻害作用は、徐々に減弱しつつも 30 分まで持続したが、90 分後には、輸送活性はほぼ完全に回復した。このことから、可逆的 な機構により阻害を生じている可能性が示唆された。さらに、この阻害作用は、myricetin 濃 度依存的であり、アルコール飲料(ワイン)中に含まれる濃度の範囲(10-100 µM 程度)で 生じ得ることも示唆された。 速度論的解析では、myricetin の濃度上昇に伴い最大輸送速度(Vmax)が低下し、阻害作用 の要因となっていることが明らかとなった。しかし一方で、ミカエリス定数(Km)の低下も みられ、folate に対する PCFT の親和性が上昇していることが示唆された。このため、Kmを 大きく下回る低基質濃度での輸送活性を表す Vmax/Kmに大きな変化はみられなかった(Fig. 2, Table 1)。この速度論的特性は、GFP 付加体 PCFT を用いた先の研究での結果と一致する ものであった。先に行われたGFP 付加体を用いた検討では、PCFT の発現量及び細胞膜局在 に変化は無いことが確認されていることに加え、今回、比較的短時間で回復するという特徴 が見出されたことから、この阻害作用は可逆的なPCFT 分子の修飾等のメカニズムによるも のである可能性が考えられる。 wortmannin(PI3K 阻害剤)での前処理により、myricetin での前処理の場合と同様の阻害 作用が見出された。しかし、阻害率は40%に留まり、myricetin による阻害(80%程度)の全 体を説明することはできないとみられた。よって、myricetin による阻害作用のメカニズム として、PI3K の関与するリン酸化反応あるいは関連の経路への作用が部分的に関与してい る可能性を否定できないが、一方で、別のメカニズムの関与が推察される。
10
第2 章 myricetin の持続性阻害作用のヒト PCFT 特異性
1 myricetin の即時性阻害作用
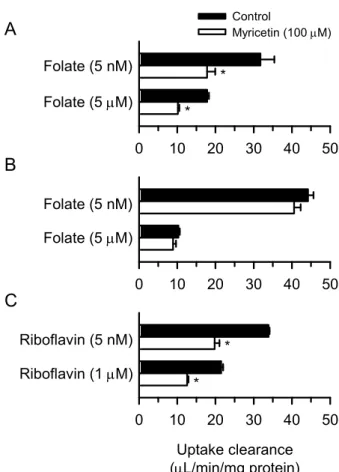
myricetin の持続性阻害作用のヒト PCFT(hPCFT)特異性を探るため、他のトランスポー ターへの影響を検討することとした。比較対象としては、汎用の実験動物であるラットの PCFT(rPCFT)とヒトの riboflavin transporter 3(hRFVT3/RFT2/SLC52A3)を取り上げた。 hRFVT3 は、小腸での riboflavin(vitamin B2)の吸収に働くトランスポーターであり、hPCFT と同様にmyricetin に曝露され得るトランスポーターの例として取り上げた。また、hRFVT3 の安定発現株を樹立することが不可能であったため(安定的遺伝子導入による細胞死のた め)、ここではHEK293 細胞一過性発現系を用いた。さらに、持続性阻害作用との関連性の 有無を探る目的で、即時性阻害作用についても検討した。rPCFT を用いた検討における試験 folate 濃度は、rPCFT による葉酸輸送の Km(2.4 µM)3)がhPCFT での値と同等であるので、 hPCFT での試験濃度と同じく、5 nM 及び 5 µM とした。hRFVT3 を用いた検討では、PCFT の場合と同様に、Km(0.77 µM)27)を大きく下回る線形領域の低riboflavin 濃度(5 nM)及び 輸送速度がほぼ Vmaxレベルに達する高riboflavin 濃度(1 µM)において阻害作用の評価を行 った。 取り込み試験液にのみmyricetin(100 µM)を添加し、前処理を行うことなく即時性阻害 作用を検討したところ、hPCFT では、低 folate 濃度(5 nM)及び高 folate 濃度(5 µM)で、 同程度の阻害(40%程度の阻害率)が認められ、先に見出されているように非競合阻害を生 じていることが確認された(Fig. 6A)。その一方で、rPCFT では、いずれの folate 濃度にお いてもmyricetin の阻害作用は認められなかった(Fig. 6B)。また、hRFVT3 では、hPCFT の 場合と同様に、低riboflavin 濃度(5 nM)及び高 riboflavin 濃度(1 µM)で、同程度の阻害 (40%程度の阻害率)がみられ、非競合阻害を生じていることが示唆された(Fig. 6C)。し たがって、即時性阻害作用の面ではrPCFT のみ myricetin 非感受性であることが明らかとな った。
11 0 10 20 30 40 50 Uptake clearance (L/min/mg protein) 0 10 20 30 40 50 Folate (5 nM) Riboflavin (5 nM) Folate (5 M) Folate (5 nM) Folate (5 M) Riboflavin (1 M) 0 10 20 30 40 50 Control Myricetin (100 M) * * * *
A
B
C
Fig. 6. Transient Effect of Myricetin on the Transport Function of hPCFT (A), rPCFT (B), and hRFVT3 (C) in HEK293 Cells Transiently Expressing the Designated Transporter
(A) The specific uptake of folate by hPCFT was evaluated in hPCFT-expressing cells for the initial 2-min period at pH 5.5 and 37ºC in the presence of myricetin (100 M) or in its absence for control. (B) The specific uptake of folate by rPCFT was similarly evaluated in rPCFT-expressing cells. (C) The specific uptake of riboflavin by hRFVT3 was evaluated in hRFVT3-expressing cells for the initial 1-min period at pH 6.0 and 37ºC in the presence of myricetin (100 M) or in its absence for control. Data are presented as the means ± SE (n = 4). * p < 0.05 compared with control, as assessed by Student's t-test.
12 2 myricetin の持続性阻害作用
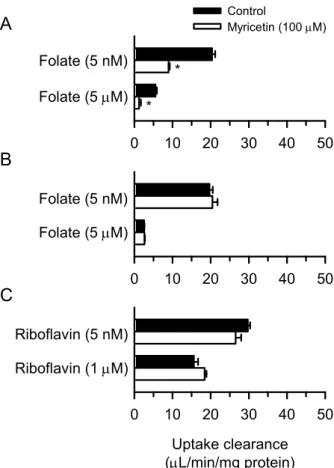
myricetin(100 µM)での 60 分間の前処理による持続性阻害作用を検討したところ、hPCFT では、低folate 濃度(5 nM)及び高 folate 濃度(5 µM)で阻害作用が認められた(Fig. 7A)。 また、高folate 濃度での阻害作用が低 folate 濃度での阻害作用より強く、MDCKII 細胞安定 発現系の場合と同様の阻害作用が確認された。その一方で、rPCFT では、いずれの folate 濃 度においてもmyricetin の阻害作用は認められず、即時性阻害作用と同様に hPCFT と rPCFT との間で、myricetin 感受性に種差があることが示唆された(Fig. 7B)。また、hRFVT3 にお いては、低riboflavin 濃度(5 nM)及び高 riboflavin 濃度(1 µM)で myricetin による持続性 阻害作用は認められなかった(Fig. 7C)。したがって、rPCFT と hRFVT3 については、持続 性阻害作用の面でhPCFT とは対照的に myricetin 非感受性であることが明らかとなった。
13 0 10 20 30 40 50 Uptake clearance (L/min/mg protein) 0 10 20 30 40 50 Folate (5 nM) Riboflavin (5 nM) Folate (5 M) Folate (5 nM) Folate (5 M) Riboflavin (1 M) * *
A
B
C
0 10 20 30 40 50 Control Myricetin (100 M)Fig. 7. Sustained Effect of Myricetin on the Transport Function of hPCFT (A), rPCFT (B), and hRFVT3 (C) in HEK293 Cells Transiently Expressing the Designated Transporter
(A) The specific uptake of folate by hPCFT was evaluated in hPCFT-expressing cells for the initial 2-min period at pH 5.5 and 37ºC in the absence of myricetin after preincubation for 60 min in the presence of myricetin (100 M), or in its absence for control, and subsequently for 5 min in its absence for washout. (B) The specific uptake of folate by rPCFT was similarly evaluated in rPCFT-expressing cells. (C) The specific uptake of riboflavin by hRFVT3 was evaluated in hRFVT3-expressing cells for the initial 1-min period at pH 6.0 and 37ºC in the absence of myricetin after preincubation for 60 min in the presence of myricetin (100 M), or in its absence for control, and subsequently for 5 min in its absence for washout. Data are presented as the means ± SE (n = 4). * p < 0.05 compared with control, as assessed by Student's t-test.
14 3 まとめ
myricetin(100 µM)による即時性阻害作用に関しては、hPCFT では、Vmax/Kmへの影響を
評価できる低folate 濃度(5 nM)及び Vmaxへの影響を評価できる高folate 濃度(5 µM)に
おいて、folate 輸送は同程度に阻害され(40%程度の阻害率)、先に見出されているように非 競合阻害を生じていることが確認された。しかし、rPCFT では、低 folate 濃度(5 nM)及び 高folate 濃度(5 µM)のいずれにおいても阻害がみられず、myricetin に対して非感受性であ ることが示唆された。一方、hRFVT3 でも、hPCFT の場合と同様に、低 riboflavin 濃度(5 nM)及び高 riboflavin 濃度(1 µM)において、riboflavin 輸送は同程度に阻害され(40%程度 の阻害率)、非競合阻害を生じていることが示唆された。 myricetin(100 µM)での 60 分間の前処理による持続性阻害作用に関しては、hPCFT では、 低及び高folate 濃度において有意な folate 輸送の阻害を生じた。また、高 folate 濃度での阻 害率(80%程度)が低 folate 濃度での阻害率(60%程度)よりも大きい点で、これまでに見 出されているhPCFT 阻害の特徴が確認された。しかし、rPCFT 及び hRFVT3 では、阻害は みられず、両トランスポーターはmyricetin に対して非感受性であることが示唆された。 以上の結果から、myricetin による非競合型の即時性阻害作用は、hPCFT と hRFVT3 に共 通する特異的な分子構造特性により生じているものと考えられる。即時性阻害作用を生じ なかった rPCFT は、それに関わる部分の分子構造が異なるものとみられる。持続性阻害作 用については、rPCFT と hRFVT3 では阻害を生じなかったことから、hPCFT に対する特異 性が高い機構により惹起されているものと考えられる。hPCFT と同様の即時性阻害作用を 生じた hRFVT3 が非感受性である点から、即時性阻害作用との関連性はないものと考えら れる。極めて相同性が高く(87%, Fig. 8)、輸送機能もほぼ同様である hPCFT と rPCFT との 間でmyricetin 感受性が異なる点は、興味深い。わずかなアミノ酸残基の差異ないしそれに 関連した分子構造の差異により、rPCFT は myricetin 非感受性となっているものと考えられ る。なお、この種差のために、汎用実験動物であるラットをPCFT の myricetin 感受性の解 析には利用できず、臓器ないし個体レベルでの解析に際しての課題となるとみられる。
15
Fig. 8. Alignment of Amino Acid Sequence of hPCFT with That of rPCFT
The amino acid sequence of hPCFT was aligned with that of rPCFT, using the program of ClustalW, and processed to visualize using the program BOXSHADE.
16 第3 章 PCFT の myricetin 感受性に関わるアミノ酸残基の同定 1 hPCFT-rPCFT 間でのキメラ体を用いた解析 hPCFT の持続性阻害作用に関する myricetin 感受性のメカニズムの解明のため、非感受性 のrPCFT との比較解析により、感受性に関わるアミノ酸残基の特定に取り組むことにした。 まず、hPCFT-rPCFT 間でキメラ体を作製し、folate 輸送に及ぼす myricetin の影響を評価する ことで、hPCFT の myricetin 感受性に関与する領域を絞り込むこととした。そこで、A-D の 4 つのキメラ体を作製した(Fig. 9A)。A、B は Spe I の制限酵素サイトをヒトとラットの PCFT にそれぞれアミノ酸配列を変えることなく導入し、そこを基点として組み換えること で作製した。C、D は rPCFT に存在する Sph I の制限酵素サイトを利用し、そこを基点とし て組み換えることで作製した。作製した4 種のキメラ体を HEK293 細胞に一過性発現させ、 myricetin(100 µM)での前処理(60 min)の影響を検討した結果、キメラ体の B(83-459 番 目がhPCFT に相当)及び C(1-186 番目が hPCFT に相当)において、有意な阻害作用が認 められた(Figs. 9B and 9C)。また、hPCFT での結果と一致して、低 folate 濃度(5 nM)より も高folate 濃度(5 µM)で強い阻害作用が認められた。したがって、両者に共通の hPCFT 配列に当たる83-186 番目の領域内に、myricetin 感受性に関与するアミノ酸残基が存在する ことが示唆された。
17 human rat A B C D
A
B C
Uptake rate (% of control)
0 20 40 60 80 100 120 140 160 A B C D * *
Uptake rate (% of control)
0 20 40 60 80 100 120 140 160
Control Myricetin
* *
Fig. 9. Effect of Pretreatment with Myricetin on the Uptake of Folate by Chimeric PCFTs Transiently Expressed in HEK293 Cells
(A) Four chimeric PCFTs between human and rat. Construct A, B, C and D contain amino acid sequence of hPCFT at 1-82, 83-459, 1-186 and 188-459 respectively. (B, C) Mean ± S.E. (n = 4); specific uptake of [3H]folate [5 nM (B) or 5 µM (C)] for 2 min at 37C and pH
5.5; pretreatment in buffer for 60 min at pH 5.5 and 37C in the presence of myricetin (100 µM) or in its absence (control); control values in panel B, 52.6, 90.7, 130 and 44.2 fmol/min/mg protein, respectively, for construct A, B, C and D; control values in panel C, 6.30, 24.2, 43.3 and 3.86 pmol/min/mg protein, respectively, for construct A, B, C and D; * p < 0.05.
18
2 部位特異的変異導入による hPCFT の myricetin 感受性アミノ酸残基の同定
83-186 番目の領域内でヒトとラットの PCFT のアミノ酸配列を比較すると、10 箇所でア ミノ酸残基が異なっていることが明らかとなった(Fig. 10)。そこで、該当する hPCFT のア ミノ酸残基をrPCFT の対応アミノ酸残基に置換した変異体を作製し、各々の変異体を一過 性発現させたHEK293 細胞において folate 輸送における myricetin 感受性を評価した。その 結果、10 種の変異体のうち、9 つの変異体については、野生型の hPCFT と同様に myricetin 前処理によるfolate 取り込みの顕著な低下が認められた(Table 3)。その一方で、hPCFT の 158 番目のグリシン残基(G158)を rPCFT の対応アミノ酸残基であるアスパラギン残基(N) に置換したG158N の変異体では、両 folate 濃度において myricetin 前処理による folate 取り 込みの低下は認められず、myricetin 感受性が失われるという結果が得られた(Fig. 11)。 逆に、rPCFT の 158 番目のアスパラギン残基(N158)を hPCFT の対応アミノ酸残基であ るグリシン残基(G)に置換した N158G の変異体では、野生型の rPCFT では現れなかった 阻害作用がみられた(Fig. 12)。したがって、hPCFT/G158N での検討と合わせて、158 番目 のアミノ酸残基がPCFT の myricetin 感受性に関与するアミノ酸残基であり、ヒトとラット における1 アミノ酸置換が PCFT の myricetin 感受性を劇的に変化させることが明らかとな った。
19
Fig. 10. Alignment of Amino Acid Sequence of hPCFT with That of rPCFT (Amino Acids 83-186)
The amino acid sequence of hPCFT was aligned with that of rPCFT, using the program of ClustalW, and processed to visualize using the program BOXSHADE.
Table 3. Effect of Pretreatment with Myricetin on the Uptake of Folate by Mutant hPCFTs Transiently Expressed in HEK293 Cells
Uptake rate
Amino acid (fmol/min/mg protein) (pmol/min/mg protein) substitution Folate (5 nM) Folate (5 µM)
None (control) Myricetin None (control) Myricetin S100W 147.6 ± 2.7 27.8 ± 2.3* 43.5 ± 1.4 ND S110R 154.1 ± 8.4 61.6 ± 3.7* 45.9 ± 1.8 9.1 ± 1.0* L129V 153.5 ± 3.0 76.7 ± 2.8* 62.1 ± 2.6 10.2 ± 0.6* V132I 146.5 ± 7.7 91.2 ± 4.0* 63.8 ± 4.4 18.7 ± 0.9* V141I 150.9 ± 3.1 72.6 ± 7.1* 53.4 ± 3.0 5.1 ± 0.8* Y143F 147.8 ± 2.7 72.7 ± 3.6* 57.7 ± 5.5 12.4 ± 1.3* I149A 120.9 ± 1.5 36.4 ± 2.6* 40.3 ± 3.9 5.8 ± 1.3* S174N 149.3 ± 6.0 60.1 ± 9.3* 54.1 ± 5.7 11.3 ± 1.7* R175H 160.8 ± 4.0 70.4 ± 3.2* 50.3 ± 3.1 7.1 ± 0.7* Mean ± S.E. (n = 4); specific uptake of [3H]folate for 2 min at 37°C and pH 5.5; pretreatment
in buffer for 60 min at pH 5.5 and 37°C in the presence of a myricetin (100 µM) or in its absence (control); * p < 0.05. ND, not detected.
20
Uptake rate (% of control)
0 20 40 60 80 100 120 140 160
A B
Uptake rate (% of control)
0 20 40 60 80 100 120 140 160
None (control)
Myricetin
Fig. 11. Effect of Pretreatment with Myricetin on the Uptake of Folate by Mutant hPCFT (G158N) Transiently Expressed in HEK293 Cells
Mean ± S.E. (n = 4); specific uptake of [3H]folate [5 nM (A) or 5 µM (B)] for 2 min at 37°C
and pH 5.5; pretreatment in buffer for 60 min at pH 5.5 and 37°C in the presence of myricetin (100 µM) or in its absence (control); control, 86.7 fmol/min/mg protein and 7.22 pmol/min/mg protein, respectively, for 5 nM and 5 µM; * p < 0.05.
Uptake rate (% of control)
0 20 40 60 80 100 120
*
A B
Uptake rate (% of control)
0 20 40 60 80 100 120
None (control)
Myricetin *
Fig. 12. Effect of Pretreatment with Myricetin on the Uptake of Folate by Mutant rPCFT (N158G) Transiently Expressed in HEK293 Cells
Mean ± S.E. (n = 4); specific uptake of [3H]folate [5 nM (A) or 5 µM (B)] for 2 min at 37°C
and pH 5.5; pretreatment in buffer for 60 min at pH 5.5 and 37°C in the presence of myricetin (100 µM) or in its absence (control); control, 90.1 fmol/min/mg protein and 25.2 pmol/min/mg protein, respectively, for 5 nM and 5 µM; * p < 0.05.
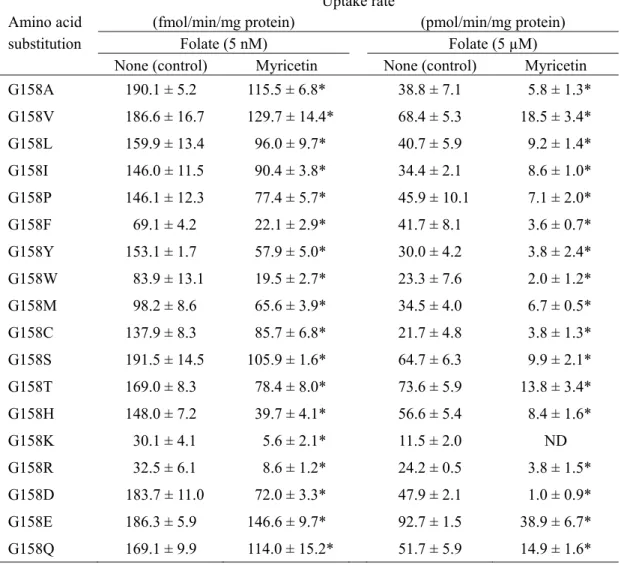
21 3 hPCFT の G158 の各種アミノ酸による置換の影響 さらなる詳細な解析のため、hPCFT の G158 を N 以外の 18 種のアミノ酸に置換した変異 体についても、myricetin 感受性を検討した。その結果、18 種全ての変異体で myricetin 前処 理による阻害作用が認められた(Table 4)。このように、hPCFT は G158 を N に置換した場 合にのみmyricetin 感受性を失うことから、G への特異的な作用等ではなく、何らかの間接 的な機構によりmyricetin 感受性阻害を生じている可能性が考えられる。 また、hPCFT の G158 は膜貫通領域に局在すると共に、基質結合ポケットの内部またはそ の周辺に局在し、基質輸送の際のコンフォメーション変化に関与する残基であることが示 唆されている28)。したがって、ヒトとラットにおけるmyricetin 感受性の差異は、PCFT の 立体構造の違いにより生じているものと推察される。 なお、本研究の持続性阻害試験では、基質輸送時に myricetin が細胞外液中に存在しない ことを考慮すると、前処理過程において細胞内に取り込まれたmyricetin が惹起する作用に より、G158 を含む基質結合ポケットに影響を生じているものと推察される(Fig. 13)。
22
Table 4. Effect of Mutation at G158 in hPCFT Transiently Expressed in HEK293 Cells on Myricetin Sensitivity
Uptake rate
Amino acid (fmol/min/mg protein) (pmol/min/mg protein) substitution Folate (5 nM) Folate (5 µM)
None (control) Myricetin None (control) Myricetin G158A 190.1 ± 5.2 115.5 ± 6.8* 38.8 ± 7.1 5.8 ± 1.3* G158V 186.6 ± 16.7 129.7 ± 14.4* 68.4 ± 5.3 18.5 ± 3.4* G158L 159.9 ± 13.4 96.0 ± 9.7* 40.7 ± 5.9 9.2 ± 1.4* G158I 146.0 ± 11.5 90.4 ± 3.8* 34.4 ± 2.1 8.6 ± 1.0* G158P 146.1 ± 12.3 77.4 ± 5.7* 45.9 ± 10.1 7.1 ± 2.0* G158F 69.1 ± 4.2 22.1 ± 2.9* 41.7 ± 8.1 3.6 ± 0.7* G158Y 153.1 ± 1.7 57.9 ± 5.0* 30.0 ± 4.2 3.8 ± 2.4* G158W 83.9 ± 13.1 19.5 ± 2.7* 23.3 ± 7.6 2.0 ± 1.2* G158M 98.2 ± 8.6 65.6 ± 3.9* 34.5 ± 4.0 6.7 ± 0.5* G158C 137.9 ± 8.3 85.7 ± 6.8* 21.7 ± 4.8 3.8 ± 1.3* G158S 191.5 ± 14.5 105.9 ± 1.6* 64.7 ± 6.3 9.9 ± 2.1* G158T 169.0 ± 8.3 78.4 ± 8.0* 73.6 ± 5.9 13.8 ± 3.4* G158H 148.0 ± 7.2 39.7 ± 4.1* 56.6 ± 5.4 8.4 ± 1.6* G158K 30.1 ± 4.1 5.6 ± 2.1* 11.5 ± 2.0 ND G158R 32.5 ± 6.1 8.6 ± 1.2* 24.2 ± 0.5 3.8 ± 1.5* G158D 183.7 ± 11.0 72.0 ± 3.3* 47.9 ± 2.1 1.0 ± 0.9* G158E 186.3 ± 5.9 146.6 ± 9.7* 92.7 ± 1.5 38.9 ± 6.7* G158Q 169.1 ± 9.9 114.0 ± 15.2* 51.7 ± 5.9 14.9 ± 1.6* Mean ± S.E. (n = 4); specific uptake of [3H]folate for 2 min at 37°C and pH 5.5; pretreatment
in buffer for 60 min at pH 5.5 and 37°C in the presence of a myricetin (100 µM) or in its absence (control); * p < 0.05. ND, not detected.
23
24 4 各動物種における PCFT の myricetin 感受性種差 hPCFT の G158 に相当する箇所に着目し、各動物種で比較すると Fig. 14 のようになって いることが明らかとなった。そこで、対応アミノ酸残基がそれぞれG、S(セリン)、N とな っているサル、ウシ、マウスのPCFT について、HEK293 細胞に一過性発現させ、myricetin での前処理の影響を検討した。その結果、サル、ウシにおいてはmyricetin の阻害作用が認 められ、hPCFT での結果と一致して、低 folate 濃度よりも高 folate 濃度でより強い阻害作用 がみられた(Fig. 15)。その一方で、対応アミノ酸残基がラットと同様に N となっているマ ウスでは、いずれのfolate 濃度においても阻害作用は認められなかった。したがって、hPCFT の G158 に対応するアミノ酸残基が N となっている種では、ラット及びマウスと同様に myricetin 感受性が失われている可能性が考えられ、実験動物の利用に際して、注意が必要 になるとみられる。
Fig. 14. Alignment of Amino Acid Sequence of hPCFT with Those of agmPCFT, bPCFT, rPCFT and mPCFT
The amino acid sequence of hPCFT was aligned with those of agmPCFT, bPCFT, rPCFT and mPCFT, using the program of ClustalW, and processed to visualize using the program BOXSHADE.
25
Uptake rate (% of control)
0 20 40 60 80 100 120 * * Bovine Mouse Monkey
Uptake rate (% of control)
0 20 40 60 80 100 120 Control Myricetin * * A B
Fig. 15. Effect of Pretreatment with Myricetin on the Uptake of Folate by PCFTs of Various Animal Species in Transiently Transfected HEK293 Cells
Mean ± S.E. (n = 4); specific uptake of [3H]folate [5 nM (A) or 5 µM (B)] for 2 min at 37°C
and pH 5.5; pretreatment in buffer for 60 min at pH 5.5 and 37°C in the presence of myricetin (100 µM) or in its absence (control); control values in panel A, 131, 94.8 and 90.7 fmol/min/mg protein, respectively, for monkey, bovine and mouse; control values in panel B, 39.1, 53.7 and 13.2 pmol/min/mg protein, respectively, for monkey, bovine and mouse; * p < 0.05.
26 5 まとめ hPCFT と rPCFT との myricetin 感受性の差異を利用した比較解析により、hPCFT の G158 が持続性のmyricetin 感受性阻害に関与していることが見出された。hPCFT の G158 を rPCFT の対応アミノ酸残基であるN に置換すると、myricetin 感受性が失われ、rPCFT 様の特性を 示した。一方で、rPCFT の N158 を hPCFT の対応アミノ酸残基である G に置換すると、 myricetin 感受性となって hPCFT 様の特性を示したことから、この 1 アミノ酸残基の違いに より、ヒトとラットとの間でのPCFT の myricetin 感受性種差が生じていることが示唆され た。また、hPCFT の G158 を N 以外のアミノ酸に置換した場合には、myricetin 感受性は維 持されたことから、G への特異的な作用ではなく、間接的な機構により myricetin が持続性 阻害作用を惹起していることも示唆された。さらに、hPCFT の G158 に対応するアミノ酸残 基がそれぞれG、S となっているサル、ウシでは myricetin 感受性がみられた一方で、N と なっているマウスではmyricetin 感受性がみられなかったことから、G158 に対応するアミノ 酸残基がN となっている動物種では、myricetin 感受性が失われている可能性が示唆された。 したがって、それらの動物種については、関連研究での利用に際して、注意が必要になると みられる。
27
結論
本研究では、ヒトPCFT(hPCFT)に対する myricetin の持続性阻害作用の解析に取り組み、 以下の知見を得た。 1. hPCFT に対する myricetin の持続性阻害作用は、myricetin への曝露時間に依存して増強 される性質を示したが、一方で、可逆的なものであることが示唆された。さらに、この 阻害作用はmyricetin 濃度に依存的であり、アルコール飲料(ワイン)中に含まれる濃 度の範囲(10-100 µM 程度)で生じ得ることも示唆された。 2. ラット PCFT(rPCFT)は、hPCFT との相同性が高いにも関わらず、myricetin による即 時性及び持続性阻害を生じず、myricetin 感受性に種差があることが明らかとなった。ま た、hRFVT3 でも、myricetin による持続性阻害を生じず、持続性阻害作用は hPCFT に 対する特異性の高い機構により惹起されていることが示唆された。一方、hRFVT3 は、 即時性阻害作用については、hPCFT と同様に myricetin 感受性であった。このことから、 即時性阻害作用と持続性阻害作用に関連性はないことが示唆された。 3. myricetin に対して感受性である hPCFT と非感受性である rPCFT との比較解析により、 持続性阻害作用に関して、hPCFT の G158 が myricetin 感受性に関わるアミノ酸残基で あることが明らかとなった。 本研究の成果は、folate 類の吸収阻害の原因となり得る、myricetin 類による持続性 PCFT 阻害作用の特性の把握及び機構の解明を大きく進展させるものである。また、folate 及び関 連医薬品を用いた経口薬物療法の最適化のための基盤情報として役立つものと期待される。28
実験の部
I 実験材料
1 試薬
[3H]folate(27.2 Ci/mmol)、[3H]riboflavin(24 Ci/mmol)、[14C]inulin(1.2 mCi/gram)は、Moravek
Biochemicals, Inc.(Brea, CA, U.S.A.)より購入した。非標識の folate 及び riboflavin は、Wako Pure Chemical Industries, Ltd.(Osaka, Japan)製のものを用いた。myricetin は、Tokyo Chemical Industry Co., Ltd.(Tokyo, Japan)より購入して用いた。液体シンチレーションカクテルとし ては、Nacalai Tesque, Inc.(Kyoto, Japan)製の Clear-sol I を、遺伝子操作用キットとしては、 Sigma-Aldrich(St. Louis, MO, USA)製の TRI REAGENT® BD、QIAGEN(Valensia, CA, U.S.A.)
製のQIAEX®IIGel Extraction Kit、Thermo Fisher Scientific(Waltham, MA, U.S.A)製の PureLink™
HiPure Plasmid Midiprep Kit を用いた。DNA polymerase としては、TOYOBO(Osaka, Japan) 製のKOD-Plus-Neo、Takara Bio, Inc.(Shiga, Japan)製の PrimeSTAR® Max DNA Polymerase を
用いた。ベクターは、Promega(Fitchburg, WI, U.S.A.)製の pCI-neo ベクターを用いた。ま た、逆転写酵素はReverTra Ace(TOYOBO)を用いた。その他の試薬は全て市販特級品を使 用した。
2 培養細胞
MDCKII 細胞(Madin-Darby canine kidney II cells)及び MDBK 細胞(Madin-Darby bovine kidney cells)は RIKEN BioResource Center(Ibaraki, Japan)より提供を受けた。HEK293 細胞 (human embryonic kidney 293 cells)及び Cos-7 細胞は Cell Resource Center for Biomedical Research, Tohoku University(Miyagi, Japan)より提供を受けた。
II 実験方法
1 各種トランスポーターのクローニング
1-1 human proton coupled folate transporter(hPCFT)
hPCFT プラスミドは、先に当研究室にて調製されたものを用いた2)。これは、RT-PCR 法
29 1-2 rat proton coupled folate transporter(rPCFT)
rPCFT プラスミドについては、先に当研究室にて RT-PCR 法によりクローニングされた rPCFT の cDNA を pGH ベクターに組み込むことにより調製されたものを用い、今回、pCI-neo ベクターに組み換えることにより調製した3)。
1-3 human riboflavin transporter 3(hRFVT3)
hRFVT3 プラスミドは、先に当研究室にて調製されたものを用いた 29)。これは、RT-PCR 法によりクローニングしたhRFVT3 の cDNA を pCI-neo ベクターに組み込んだものである。 1-4 hPCFT-rPCFT 間のキメラ体 hPCFT の部分配列と rPCFT の部分配列を組み合わせることによりキメラ体を作製した。 hPCFT/pCI-neo または rPCFT/pCI-neo を鋳型とし、制限酵素サイトを組み込んだプライマー を用いてPCR を行った。polymerase は KOD-plus-Neo を用いた。得られた PCR 産物を制限 酵素で処理し、その後電気泳動を行い、QIAEX®IIGel Extraction Kit を用いて精製、抽出し
た。哺乳類発現ベクターであるpCI-neo ベクターも同様の制限酵素処理を行った後、精製し た。得られたhPCFT 及び rPCFT の各部分配列と pCI-neo ベクターを T4 DNA Ligase を用い てトリプルライゲーションし、ヒートショック(42ºC, 1 min)によって大腸菌(DH5α)に導 入した。その大腸菌をampicillin の添加された LB プレートに播き、37ºC で一晩(約 12 時 間)培養した後、プレート上のコロニーを採取した。そこからプラスミド DNA を抽出し、 制限酵素処理後、電気泳動を行うことによりインサート部分を確認した。予想された長さの インサート部分が確認されたコロニーから得られたプラスミドは、シークエンスを確認し、 インサート部分の塩基配列に変異がないことを確認した(ABI PRISM® 3130-Avant Genetic
Analyzer)。
hPCFT の部分配列の作製には、以下のプライマーを使用した。 1-82 番目のアミノ酸配列
forward primer: 5'-CTCTCCACAGGTGTCCACTC-3' reverse primer, + Spe I site(下線): 5'-GTGACTAGTAAGGGTCTCCACTT-3' 83-459 番目のアミノ酸配列
forward primer, + Spe I site(下線): 5'-CTTACTAGTCACTGGACCCTCTA-3' reverse primer: 5'-TTCACTGCATTCTAGTTGTGG-3' 1-186 番目のアミノ酸配列
forward primer: 5'-CTCTCCACAGGTGTCCACTC-3' reverse primer, + Sph I site(下線): 5'-GATGCATGCTTCCAGCAGGGCCAT-3'
30 187-459 番目のアミノ酸配列
forward primer, + Sph I site(下線): 5'-GAAGCATGCATCGGGGTGGCT-3' reverse primer: 5'-TTCACTGCATTCTAGTTGTGG-3'
rPCFT の部分配列の作製には、以下のプライマーを使用した。 1-82 番目のアミノ酸配列
forward primer: 5'-CTCTCCACAGGTGTCCACTC-3' reverse primer, + Spe I site(下線): 5'-GTGACTAGTCAGGGTCTCCACTT-3' 83-459 番目のアミノ酸配列
forward primer, + Spe I site(下線): 5'-CTGACTAGTCACTGGACCCTCTA-3' reverse primer: 5'-TTCACTGCATTCTAGTTGTGG-3' 1-186 番目のアミノ酸配列
forward primer: 5'-CTCTCCACAGGTGTCCACTC-3' reverse primer, + Sph I site(下線): 5'-AATGCATGCTTCCAGCAGAGCCAT-3' 187-459 番目のアミノ酸配列
forward primer, + Sph I site(下線): 5'-GAAGCATGCATTGGTGTGGCC-3' reverse primer: 5'-TTCACTGCATTCTAGTTGTGG-3'
1-5 hPCFT 及び rPCFT の変異体
変異体を作製するにあたり、hPCFT/pCI-neo または rPCFT/pCI-neo を鋳型とし、以下に示 したプライマーを用いてPCR を行った(Tables 9 and 10)。polymerase は PrimeSTAR® Max
DNA Polymerase を用いた。得られた PCR 産物をヒートショック(42ºC, 1 min)によって大 腸菌(DH5α)に導入した。その後は、キメラ体の場合と同様の操作を行い、クローンを得 た。
31
Table 9. Primer Sequences Used for Amplification of the Mutant Genes of hPCFT
Mutant Orientation Sequence (5' to 3') S100W (TCG→TGG) S110R (AGT→CGC) L129V (CTA→GTG) V132I (GTT→ATC) V141I (GTC→ATC) Y143F (TAC→TTC) I149A (ATC→GCC) G158N (GGT→AAC) G158A (GGT→GCC) G158V (GGT→GTG) G158L (GGT→CTG) G158I (GGT→ATC) G158P (GGT→CCC) G158F (GGT→TTC) G158Y (GGT→TAC) G158W (GGT→TGG) G158M (GGT→ATG) G158C (GGT→TGC) G158S (GGT→AGT) G158T (GGT→ACC) G158H (GGT→CAC) Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse TGGTCCACCCTGCTGGGAGCTT GAAGAGCCCCACCAGGAA AGCGACCGCGTGGGCCGCCGCCCGCTG GCCCACGCGGTCGCTCCAAGCTCCCAG GTGGTGTCCGTTTTTGTGGTGCA GGCCTGGAGCAGCAGGCCCA GTGTCCATCTTTGTGGTGCAGCTGCAG CACAAAGATGGACACTAGGGCCTGGAG ATCGGCTACTTCGTGCTGGGT GTGGAGCTGCAGCTGCACCA GTCGGCTTCTTCGTGCTGGGTCGCATC CACGAAGAAGCCGACGTGGAGCTGCAG GCCCTTTGTGCCCTCCTCGG GCGACCCAGCACGAAGTA GACTTCAACGGCCTTCTGGCTGCTAGC AAGGCCGTTGAAGTCGCCGAGGAGGGC GACTTCGCCGGCCTTCTGGCTGCTAGC AAGGCCGGCGAAGTCGCCGAGGAGGGC GACTTCGTGGGCCTTCTGGCTGCTAGC AAGGCCCACGAAGTCGCCGAGGAGGGC GACTTCCTGGGCCTTCTGGCTGCTAGC AAGGCCCAGGAAGTCGCCGAGGAGGGC GACTTCATCGGCCTTCTGGCTGCTAGC AAGGCCGATGAAGTCGCCGAGGAGGGC GACTTCCCCGGCCTTCTGGCTGCTAGC AAGGCCGGGGAAGTCGCCGAGGAGGGC GACTTCTTCGGCCTTCTGGCTGCTAGC AAGGCCGAAGAAGTCGCCGAGGAGGGC GACTTCTACGGCCTTCTGGCTGCTAGC AAGGCCGTAGAAGTCGCCGAGGAGGGC GACTTCTGGGGCCTTCTGGCTGCTAGC AAGGCCCCAGAAGTCGCCGAGGAGGGC GACTTCATGGGCCTTCTGGCTGCTAGC AAGGCCCATGAAGTCGCCGAGGAGGGC GACTTCTGCGGCCTTCTGGCTGCTAGC AAGGCCGCAGAAGTCGCCGAGGAGGGC GACTTCAGTGGCCTTCTGGCTGCTAGC AAGGCCACTGAAGTCGCCGAGGAGGGC GACTTCACCGGCCTTCTGGCTGCTAGC AAGGCCGGTGAAGTCGCCGAGGAGGGC GACTTCCACGGCCTTCTGGCTGCTAGC AAGGCCGTGGAAGTCGCCGAGGAGGGC To be continued.
32
Mutant Orientation Sequence (5' to 3') G158K (GGT→AAG) G158R (GGT→AGA) G158D (GGT→GAC) G158E (GGT→GAG) G158Q (GGT→CAG) S174N (AGT→AAT) R175H (CGC→CAC) Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse Forward Reverse GACTTCAAGGGCCTTCTGGCTGCTAGC AAGGCCCTTGAAGTCGCCGAGGAGGGC GACTTCAGAGGCCTTCTGGCTGCTAGC AAGGCCTCTGAAGTCGCCGAGGAGGGC GACTTCGACGGCCTTCTGGCTGCTAGC AAGGCCGTCGAAGTCGCCGAGGAGGGC GACTTCGAGGGCCTTCTGGCTGCTAGC AAGGCCCTCGAAGTCGCCGAGGAGGGC GACTTCCAGGGCCTTCTGGCTGCTAGC AAGGCCCTGGAAGTCGCCGAGGAGGGC AGCTCCAATCGCAGCCGCACCTTCCGG GCTGCGATTGGAGCTGACATCTGCCAC TCCAGTCACAGCCGCACCTTCCGGATG GCGGCTGTGACTGGAGCTGACATCTGC
Table 10. Primer Sequences Used for Amplification of the Mutant Gene of rPCFT
Mutant Orientation Sequence (5' to 3') N158G (AAC→GGT) Forward
Reverse
GATTTCGGTGGCCTTCTTGCTGCTAGC AAGGCCACCGAAATCTCCCAGAAGGGC
1-6 african green monkey proton coupled folate transporter(agmPCFT)
agmPCFT の遺伝子導入用プラスミドの作製にあたっては、Cos-7 細胞(アフリカミドリザ ル腎臓由来)より抽出したRNA から、ReverTra Ace を用いて逆転写することによって cDNA を得た。このcDNA を鋳型とし、agmPCFT(GenBank accession no., XP_008008973.1)の cDNA を得るためにPCR を行った。polymerase は PrimeSTAR® Max DNA Polymerase を用いた。プ
ライマーは以下のものを使用した。
forward primer: 5'-CAGTCCCAGGCAGCCAGTCG-3'
reverse primer: 5'-AAGGAAGAACTGACTCCTGTCCCGA-3'
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーは以下のものを使用した。
forward primer, + EcoRI site(下線): 5'-GCTGAATTCAGACATGGAGGGGAGCGCG-3' reverse primer: 5'-AAGGAAGAACTGACTCCTGTCCCGA-3'
得られた PCR 産物を、制限酵素 EcoRI と XbaI で処理し、その後電気泳動を行い、 QIAEX®IIGel Extraction Kit を用いて精製、抽出した。哺乳類発現ベクターである pCI-neo ベ
クターも同様の制限酵素処理を行った後、精製した。これらをT4 DNA Ligase を用いてライ ゲーションし、ヒートショック(42 ºC, 1 min)によって大腸菌(DH5α)に導入した。その 後は、キメラ体の場合と同様の操作を行い、クローンを得た。
33 1-7 bovine proton coupled folate transporter(bPCFT)
bPCFT の遺伝子導入用プラスミドの作製にあたっては、MDBK 細胞(ウシ腎臓由来)よ り抽出したRNA から、ReverTra Ace を用いて逆転写することによって cDNA を得た。この cDNA を鋳型とし、bPCFT(GenBank accession no., NP_ 001073053.1)の cDNA を得るために PCR を行った。polymerase は PrimeSTAR® Max DNA Polymerase を用いた。プライマーは以
下のものを使用した。
forward primer: 5'-CTGCAAGAAACCCGAAAGCA-3' reverse primer: 5'-ACTGACTTATGATGATTGGCACT-3'
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーは以下のものを使用した。
forward primer, + XhoI site(下線): 5'-AAGCTCGAGAGCATGGAGGGACGCGCGAA-3' reverse primer, + NotI site(下線): 5'-TCAGCGGCCGCTCAGGGGCTCTGGGAAAAC-3'
得られたPCR 産物を、制限酵素 XhoI と NotI で処理し、その後は、agmPCFT の場合と同 様の操作を行い、クローンを得た。
1-8 mouse proton coupled folate transporter(mPCFT)
mPCFT の遺伝子導入用プラスミドの作製にあたっては、マウス(ddY、雄性)の小腸より 抽出した RNA から、ReverTra Ace を用いて逆転写することによって cDNA を得た。この cDNA を鋳型とし、mPCFT(GenBank accession no., NP_ 081016.2)の cDNA を得るために PCR を行った。polymerase は PrimeSTAR® Max DNA Polymerase を用いた。プライマーは以
下のものを使用した。
forward primer: 5'-CCTCCCGCAGCTGGTTCCGAG-3' reverse primer: 5'-TGCTGGGCTTTAGGCATACATCGTC-3'
これによってできたPCR 産物を鋳型として、制限酵素サイトをデザインしたプライマー を用い、再びPCR を行った。プライマーは以下のものを使用した。
forward primer, + EcoRI site(下線): 5'-CCGAATTCGAGCATGGAGGGGCGCGT-3' reverse primer, + XbaI site(下線): 5'-TCCTCATGGCTTCTAGAAGATCAGGG-3'
その後は、agmPCFT の場合と同様の操作を行い、クローンを得た。
2 細胞の培養方法
MDCKII 細胞の培養には、TPP(Transadingen, Switzerland)製の組織培養用フラスコ、24 well プレート(底面積 1.91 cm2/well)及び 96 well(底面積 0.31 cm2/well)プレートを用いた。
34
コートした24 well プレートを用いた。組織培養メディウムとして 10% FBS(Sigma-Aldrich)、 1% penicillin/streptomycin(Wako Pure Chemical Industries, Ltd.)を含む DMEM を使用し、37ºC, 95% air-5% CO2インキュベーター内で培養した。
3 遺伝子導入
3-1 安定発現系の作製:MDCKII 細胞
MDCKII 細胞は継代に際し、ビュルケルチュルク盤を用いて細胞数を計測した後、4.0 × 105 cells/ml となるように調製し、24 well プレートに 500 µl/well ずつ播種した。その直後、
lipofectamine 2000(Invitrogen)を用いてリポフェクション法により hPCFT のプラスミド DNA を導入した。6 時間後、抗生物質を含む培地に交換し、さらにその 24 時間後、1 well 中の全 細胞を剥がし、300 倍に希釈し、96 well プレートに 100 µl/well ずつ細胞を播いた。その 24 時間後0.8 mg/ml の G418(Sigma-Aldrich)を含んだ培地に交換し、遺伝子導入細胞のセレク ションを10 日間ほど行った。G418 を含んだ培地は、2、3 日おきに交換した。セレクショ ンを行った後は、G418 を含まない培地で培養し、それを安定発現株として用いた。また、 遺伝子を導入しないmock 細胞として、pCI-neo vector のみを導入し、上記の手順で作製した ものを用いた。
3-2 一過性発現系の作製:HEK293 細胞
HEK293 細胞は継代に際し、ビュルケルチュルク盤を用いて細胞数を計測した後、2.0 × 105 cells/ml となるように調製し、poly-L-lysine コートした 24 well プレートに 1 ml/well ずつ
播種した。12 時間培養後、細胞が 90%コンフルエントになったのを確認し、抗生物質の入 っていない培地に交換した。その後、lipofectamine 2000 を用いてリポフェクション法により 目的のプラスミドDNA を導入した。6 時間後、抗生物質を含む培地に交換し、さらにその 42 時間後、取り込み実験に用いた。また、遺伝子を導入しない mock 細胞として、pCI-neo vector のみを導入し、上記の手順で作製したものを用いた。 4 取り込み実験 あらかじめ、24 well プレート上でコンフルエントになるように培養した細胞を用意した。 取り込み実験用緩衝液として、10 mM MES を加えた Hanks’ buffer(136.7 mM NaCl, 5.36 mM KCl, 0.952 mM CaCl2, 0.812 mM MgSO4, 0.441 mM KH2PO4, 0.385 mM Na2HPO4, 25 mM
D-glucose)を調製した。pH の調整には、2N NaOH を用いて pH 5.5(PCFT の場合)または pH 6.0(RFVT3 の場合)に調整した。この緩衝液中に[3H]folate または[3H]riboflavin を加え、取
35 り込み試験液とした。
持続性阻害効果の検討においては、まず well 中の培地を吸引除去し、試験化合物を添加 したHanks’ buffer(1 ml)を加え、プレインキュベーションした(37°C, 60 min または所定 時間)。その後、これを取り除き、試験化合物を含まないHanks’ buffer(1 ml)を加え、プレ インキュベーションをした(37°C, 5 min)。そして、これを取り除き、取り込み試験液(0.25 ml)を加えて取り込みを開始させた。所定時間経過後に、氷冷した Hanks’ buffer(stop solution, 2 ml)を加えることにより取り込みを停止させ、stop solution(2 ml)で細胞を 2 回洗浄した。 続いて0.5% SDS を含む 0.2 N NaOH(0.5 ml)を加え、細胞を可溶化した。1 時間経過後、 マイクロピペットで well 中をよく撹拌し、その全量を放射活性測定用サンプルとしてカウ ンティングバイアルに入れた。そのサンプルを入れたカウンティングバイアルに液体シン チレーションカクテル3 ml を加え、ボルテックスミキサーにより撹拌した。その放射活性 を液体シンチレーションカウンター(LSC-5100, Aloka Co., Tokyo, Japan)により測定した。 また、BCA 法によりタンパク質定量を行い、well 中のタンパク質量を算出した。なお、Thermo Fisher Scientific, Inc.(Waltham, MA, U.S.A)より BSA を購入し、標準タンパク質として用い た。
即時性阻害効果の検討においては、まずwell 中の培地を吸い取り、37°C の Hanks’ buffer (1 ml)を加え、5 分間プレインキュベーションをした。その後、これを取り除き、試験化 合物を添加した取り込み試験液(0.25 ml)を加えて取り込みを開始させた。所定時間経過後 に、氷冷したHanks’ buffer(stop solution, 2 ml)を加えることにより取り込みを停止させ、 stop solution(2 ml)で細胞を 2 回洗浄した。以後は、持続性阻害効果の検討と同様の操作を 行った。 III データ解析 1 取り込みデータ処理 細胞内への薬物の取り込み量は以下の式を用いて算出した。 v ct s
A
A
s
A
Uptake
(1)ここで、Uptake は取り込み量(pmol/mg protein)、Asはサンプルの放射能(dpm/well)、Act
は取り込み試験液の放射能(dpm/µl)、Avは細胞のタンパク量(mg/well)、s は取り込み試験
液の基質濃度(µM)である。
取り込み量(Uptake)を取り込み時間(t)で割ることにより、取り込み速度(v)を求め た。また、遺伝子導入細胞での v から、mock 細胞での v を差し引き、トランスポーターに
36 よる特異的な v を算出した。さらに v を s で割ることにより、取り込みクリアランス(CLup) を求めた。
t
Uptake
v
(2)s
v
CL
up
(3) hPCFT による飽和性の輸送の解析においては、Michaelis-Menten 型の担体輸送を仮定して 解析を行った。s
K
s
V
v
m max・
(4) Vmaxは最大取り込み速度、KmはMichaelis 定数である。非線形最小 2 乗法プログラムであるWinNonlin(Pharsight, CA, U.S.A.)を用い、v と C との関係に(4)式を当てはめ、Vmax、Km
を算出した。当てはめ計算に際しては、重みを1/v2とした。
2 統計処理
データは平均値 ± 標準誤差として示した。2 群間の比較には Student’s t-test を用いた。多 重比較を要する場合には、ANOVA の後に Dunnet’s test を用いた。
37
謝辞
本研究を行うにあたり、六年半もの長きにわたり御指導、御鞭撻を賜りました名古屋市立 大学大学院薬学研究科 湯浅 博昭 教授に深く感謝いたします。 また、本論文作成にあたり、種々の有益な御助言と御校閲を賜りました、名古屋市立大学 大学院薬学研究科 林 秀敏 教授、頭金 正博 教授ならびに岩尾 岳洋 准教授に深 く感謝いたします。 本研究を進めるにあたり、熱心な御指導及び御助言を賜りました東京薬科大学薬学部 井上 勝央 教授、金城学院大学薬学部 太田 欣哉 准教授、名古屋市立大学大学院薬学 研究科 保嶋 智也 助教に深く感謝いたします。 本研究に御協力を賜りました金城学院大学薬学部 故 林 弥生 教授に深く感謝いた します。 研究途上種々の御便宜を図ってくださいました名古屋市立大学大学院薬学研究科 薬物 動態制御学分野の卒業生、在室生及び研究員、RI 施設員、動物舎施設員各位に感謝いたし ます。 長井記念薬学研究奨励支援事業に採用していただき、日本薬学会長井記念薬学研究奨励 金(Nagai Memorial Research Scholarship from the Pharmaceutical Society of Japan)をはじめと した、多大なる御支援をいただきました公益社団法人日本薬学会関係各位の皆様に深く御 礼申し上げます。38
参考文献
1. Qiu A, Jansen M, Sakaris A, Min SH, Chattopadhyay S, Tsai E, Sandoval C, Zhao R, Akabas MH, Goldman ID. Identification of an intestinal folate transporter and the molecular basis for hereditary folate malabsorption. Cell, 127: 917-928 (2006).
2. Nakai Y, Inoue K, Abe N, Hatakeyama M, Ohta KY, Otagiri M, Hayashi Y, Yuasa H. Functional characterization of human proton-coupled folate transporter/heme carrier protein 1 heterologously expressed in mammalian cells as a folate transporter. J. Pharmacol. Exp. Ther., 322: 469-476 (2007).
3. Inoue K, Nakai Y, Ueda S, Kamigaso S, Ohta KY, Hatakeyama M, Hayashi Y, Otagiri M, Yuasa H. Functional characterization of PCFT/HCP1 as the molecular entity of the carrier-mediated intestinal folate transport system in the rat model. Am. J. Physiol. Gastrointest. Liver Physiol., 294: G660-G668 (2008).
4. Halsted CH, Villanueva JA, Devlin AM, Chandler CJ. Metabolic interactions of alcohol and folate. J. Nutr., 132: 2367S-2372S (2002).
5. Said HM. Intestinal absorption of water-soluble vitamins in health and disease. Biochem. J., 437: 357-372 (2011).
6. Medici V, Halsted CH. Folate, alcohol, and liver disease. Mol. Nutr. Food Res., 57: 596-606 (2013).
7. Koury MJ, Horne DW. Apoptosis mediates and thymidine prevents erythroblast destruction in folate deficiency anemia. Proc. Natl. Acad. Sci. U.S.A., 91: 4067-4071 (1994).
8. Koike H, Hama T, Kawagashira Y, Hashimoto R, Tomita M, Iijima M, Sobue G. The significance of folate deficiency in alcoholic and nutritional neuropathies: analysis of a case. Nutrition, 28: 821-824 (2012).
9. Bailey DG, Malcolm J, Arnold O, Spence JD. Grapefruit juice-drug interactions. Br. J. Clin. Pharmacol., 46: 101-110 (1998).
10. Song J, Kwon O, Chen S, Daruwala R, Eck P, Park JB, Levine M. Flavonoid inhibition of sodium-dependent vitamin C transporter 1 (SVCT1) and glucose transporter isoform 2 (GLUT2), intestinal transporters for vitamin C and Glucose. J. Biol. Chem., 277: 15252-15260 (2002). 11. Huang S-M, Hall SD, Watkins P, Love LA, Serabjit-Singh C, Betz JM, Hoffman FA, Honig P,
Coates PM, Bull J, Chen ST, Kearns GL, Murray MD. Drug interactions with herbal products and grapefruit juice: a conference report. Clin. Pharmacol. Ther., 75: 1-12 (2004).
12. Kwon O, Eck P, Chen S, Corpe CP, Lee JH, Kruhlak M, Levine M. Inhibition of the intestinal glucose transporter GLUT2 by flavonoids. FASEB J., 21: 366-377 (2007).
39 Clin. Pharmacol., 70: 645-655 (2010).
14. Alvarez AI, Real R, Pérez M, Mendoza G, Prieto JG, Merino G. Modulation of the activity of ABC transporters (P-glycoprotein, MRP2, BCRP) by flavonoids and drug response. J. Pharm. Sci., 99: 598-617 (2010).
15. Dolton MJ, Roufogalis BD, McLachlan AJ. Fruit juices as perpetrators of drug interactions: the role of organic anion-transporting polypeptides. Clin. Pharmacol. Ther., 92: 622-630 (2012). 16. Brantley SJ, Argikar AA, Lin YS, Nagar S, Paine MF. Herb–drug interactions: challenges and
opportunities for improved predictions. Drug Metab. Dispos., 42: 301-317 (2014).
17. An G, Wang X, Morris ME. Flavonoids are inhibitors of human organic anion transporter 1 (OAT1)-mediated transport. Drug Metab. Dispos., 42: 1357-1366 (2014).
18. Lemos C, Peters GJ, Jansen G, Martel F, Calhau C. Modulation of folate uptake in cultured human colon adenocarcinoma Caco-2 cells by dietary compounds. Eur. J. Nutr., 46: 329-336 (2007). 19. Alemdaroglu NC, Wolffram S, Boissel JP, Closs E, Spahn-Langguth H, Langguth P. Inhibition of
folic acid uptake by catechins and tea extracts in Caco-2 cells. Planta Med., 73: 27-32 (2007). 20. Furumiya M, Inoue K, Nishijima C, Yamashiro T, Inaoka E, Ohta K, Hayashi Y, Yuasa H.
Noncompetitive inhibition of proton-coupled folate transporter by myricetin. Drug Metab. Pharmacokinet., 29: 312-316 (2014).
21. Furumiya M, Yamashiro T, Inoue K, Nishijima C, Ohta K, Hayashi Y, Yuasa H. Sustained inhibition of proton-coupled folate transporter by myricetin. Drug Metab. Pharmacokinet., 30: 154-159 (2015).
22. Shirasaka Y, Shichiri M, Murata Y, Mori T, Nakanishi T, Tamai I. Long-lasting inhibitory effect of apple and orange juices, but not grapefruit juice, on OATP2B1-mediated drug absorption. Drug Metab. Dispos., 41: 615-621 (2013).
23. Greenblatt DJ, von Moltke LL, Harmatz JS, Chen G, Weemhoff JL, Jen C, Kelley CJ, LeDuc BW, Zinny MA. Time course of recovery of cytochrome p450 3A function after single doses of grapefruit juice. Clin. Pharmacol. Ther., 74: 121-129 (2003).
24. German JB, Walzem RL. The health benefits of wine. Annu. Rev. Nutr., 20: 561–593 (2000). 25. Vuorinen H, Määtta K, Törrönen R. Content of the flavonols myricetin, quercetin, and
kaempferol in finnish berry wines. J. Agric. Food Chem., 48: 2675-2680 (2000).
26. Babu PV, Liu D, Gilbert ER. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem., 24: 1777-1789 (2013).
27. Fujimura M, Yamamoto S, Murata T, Yasujima T, Inoue K, Ohta KY, Yuasa H. Functional characteristics of the human ortholog of riboflavin transporter 2 and riboflavin-responsive expression of its rat ortholog in the small intestine indicate its involvement in riboflavin absorption. J. Nutr., 140: 1722-1727 (2010).
proton-40
coupled folate transporter function as assessed by the substituted cysteine accessibility method. Am. J. Physiol. Cell Physiol., 304: C1159-C1167 (2013).
29. Yamamoto S, Inoue K, Ohta KY, Fukatsu R, Maeda JY, Yoshida Y, Yuasa H. Identification and functional characterization of rat riboflavin transporter 2. J. Biochem., 145: 437-443 (2009).