脊髄後根神経節の神経細胞における
細胞内カルシウムイオン濃度の増大に対するオレキシン
Aの効果
―疼痛モデルラットを用いた研究―
日本大学大学院松戸歯学研究科歯学専攻
木村 全孝
(指導:河相 安彦 教授)
目 次
Ⅰ
. AbstractⅡ
.緒言
Ⅲ
.実験方法および結果
1. [
研究
1] SNLラットを用いた研究
1)
SNLラットの作製
2
)
[Ca2+]iの測定
3)統計解析
4)結果
2. [
研究
2] CAラットを用いた研究
1)
CAラットの作製
2
)免疫組織化学染色
3)
[Ca2+]iの測定
4)統計解析
5)結果
Ⅳ
.考察
Ⅴ
.結論
Ⅵ
.参考文献
Ⅶ
.図および表
1
Ⅰ
. Abstract PurposeIt is often difficult to demonstrate the analgesic effects of nonsteroidal anti-inflammatory drugs (NSAIDs) in the treatment of neuropathic pain. Thus, elucidating the mechanisms of neuropathic pain is crucial for establishing effective treatments for chronic pain. Orexin-A and B are neurotransmitters that were discovered by Sakurai et al. and that exert a number of physiological roles, such as the regulation of food intake and sleep patterns. Orexin-A is localized on the dorsal root ganglion (DRG), which plays an important role in pain transmission and it has been shown to induce anti-noiceptive effects in animal models of neuropathic pain.
Therefore, orexin-A may induce its analgesic effects through DRG neurons. However, the effects of orexin-A on DRG neural activity are unclear. Intra-cellular free calcium ion concentration ([Ca2+]i) is increased by depolarization that is associated with neuronal firing. The purpose of these two studies was to examine whether orexin-A suppressed high K+-induced [Ca2+]i increases in DRG neurons that were obtained from rat pain models.
Materials and Methods
In study 1, orexin-A was administered to K+-stimulated DRG nerve cells that were derived from an animal model of neuropathic pain, namely segmental spinal nerve ligation (SNL) rats.
The [Ca2+]i were assessed by using a CaF-110 Ca2+ analyzer and confocal laser microscopy.
Nifedipine, an L-type Ca2+ channel inhibitor, and lidocaine, a voltage-gated Na+ channel inhibitor, were also administered for comparison.
In study 2, an animal model of inflammatory pain namely, carrageenan-induced paw inflammation model rat (CA) were used. The expression of the N-methyl-D-aspartate receptor (NMDAR), a glutamate receptor, and orexin-A in the DRG and L4 spinal cord was analyzed immunohistochemically. In addition, orexin-A was administered to K+-stimulated DRG nerve cell that were derived from CA rats to assess the suppressive effect of orexin-A by confocal laser microscopy. In both studies, sham rats were used as controls of SNL and CA.
Results
In study 1, [Ca2+]i increases induced by high K+ stimulation in sham rats was 223.5 ± 35.8%.
Nifedipine and lidocaine significantly suppressed this increase, but orexin-A failed to suppress it.
The high K+ stimulation-induced increase in [Ca2+]i in DRG neurons from SNL rats were inhibited in the presence of orexin-A as well as nifedipine and lidocaine.
In study 2, the expression of NMDAR and orexin-A was observed in the DRG of both sham and CA rats. The NMDAR was expressed at a higher level in CA than in the control. Orexin-A did not suppress the increase in [Ca2+]i that was induced by high K+ stimulation in the control group. However, orexin-A significantly suppressed the increase in [Ca2+]i in the CA group.
2
Conclusions
These findings suggest that orexin-A inhibits Ca2+ influx through L-type Ca2+ channels on the cell membrane by the depolarization of SNL and CA DRG neurons. Furthermore, orexin-A may suppress not only in neuropathic pain but also in inflammatory pain by acting on DRG neurons.
3
Ⅱ
.緒 言
疼痛には神経障害性疼痛と炎症性疼痛があり,前者は非ステロイド性抗炎症薬
(
Non-Steroidal Anti-Inflammatory Drugs: NSAIDs)が鎮痛効果を発揮し難いのに対し,
後者は
NSAIDsによる鎮痛効果が期待できる。神経障害性疼痛でみられる自発痛や痛
覚過敏反応の治療には,抗てんかん薬や抗うつ薬などが用いられるもののその効果は必 ずしも確実とはいえず,めまいなどの副作用も多い。そのため,神経障害性疼痛発現機 構の解明を通じ効果的で安全な治療法の確立が求められている。
神経障害性疼痛の基礎研究で用いられるモデル動物に脊髄神経結紮(
segmental spinalnerve ligation: SNL
)ラット
1)があり,脊髄の部分結紮に伴い神経障害性疼痛様の痛覚過
敏が誘発される
2,3)。こうしたモデル動物では,知覚神経の過剰興奮のほか
4),脊髄後根
神経節(
dorsal root ganglion: DRG)神経細胞の興奮による神経伝達物質の遊離促進も見
られる。したがって,これらの抑制は神経障害性疼痛を含む慢性疼痛治療につながる可 能性がある。
オレキシン
A・
Bは桜井ら
5,6)により発見された神経ペプチドであり,視床下部から 脳の各部位へ神経線維を介して運ばれ,摂食や睡眠パターンの調節などの機能を発揮す ることが報告されている。興味深いことに疼痛伝達系において重要な役割を担う
DRG神経細胞にはオレキシン
Aが認められるうえ,オレキシン
Aは実験動物で抗侵害作用 も示されている
7-10)。これらのことは,オレキシン
Aは,摂食,覚醒以外にも
DRG神 経細胞を介した鎮痛作用を示す可能性を示唆するものである。しかしながら,
DRG神 経細胞の活動にオレキシン
Aが及ぼす影響は明らかでない。細胞内カルシウムイオン 濃度(
[Ca2+]i)の増加が観察される生理現象は多いが,神経細胞では神経発火に伴う脱 分極により
[Ca2+]iが増大する。
そこで本論文では疼痛モデルラットから得た
DRG神経細胞において高
K+刺激が誘 発した
[Ca2+]iの増加をオレキシン
Aが抑制するか否かについて検討した。
すなわち,研究
1では神経障害性疼痛モデル動物として知られる
SNLラット由来の
DRG神経細胞を用い,
K+刺激が誘発した
[Ca2+]iの増加に対するオレキシン
Aの効果に ついて検討した。比較の目的で
L-type Ca2+チャネル阻害薬のニフェジピンと,電位依存 性
Na+チャネル阻害薬のリドカインの投与実験も行った。研究
2では炎症性疼痛モデル 動物
11)として知られるカラゲニン足蹠投与 (
Carrageenan-induced paw inflammation model:CA
)ラットを用い,脊髄ならびに
DRG神経細胞において疼痛に関する神経伝達への関 与 が 推 測 さ れ る グ ル タ ミ ン 酸 受 容 体 の ひ と つ で あ る
N-methyl-D-aspartate receptor(
NMDAR)とともにオレキシン
Aの発現を免疫組織化学的に確認した。さらにこの
CA
ラットから得た
DRG神経細胞を用い,
K+刺激が誘発した
[Ca2+]iの増加に対するオレ キシン
Aの効果を観察した。いずれの研究でも対照群には
sham処置を行ったラットを 用いた。
4
Ⅲ
.実験方法および結果
1. [
研究
1] SNLラットを用いた研究
1)
SNLラットの作製
(
1)実験動物
実験動物は,
10週齢の
Wistar系雄性ラット(三協ラボサービス,日本)を使用した。
飼育は室温
23 ± 1℃,湿度
60 ± 10%,
12時間ごとの明暗サイクルの環境下で,飼 料は固形飼料(
MF,オリエンタル酵母工業,日本)を,飲料水は濾過水道水をそれぞ れ自由摂取させた。なお,本実験は日本大学松戸歯学部実験動物倫理委員会の承認
(
AP12MD025号)を得て,日本大学松戸歯学部実験動物指針に基づいておこなった。
(
2)脊髄神経の結紮
1
週間予備飼育した
Wistar系雄性ラット(
11週齢,体重
221.7 ± 21.8 g)を
1群
5匹とし合計
10匹使用した。脊髄神経結紮による
SNLラットの作製は
Kimら
1)の方法に 従い,ペントバルビタールナトリウム(ソムノペンチル,共立製薬,日本)
30 mg/kgの用量で腹腔内注射を行い,背部側を剃毛し,メスを用いて正中線に沿って縦に長さ
40 mm
の皮膚切開を施した。第
5,第
6(
L5, L6)脊髄神経をステンレス製ワイヤー(リ
ガチャーワイヤー
0.2 mm,トミーインターナショナル,日本)にて結紮,筋層および皮 膚を縫合により閉創した。
Sham群は脊髄神経の結紮以外,
SNL群同様の処置を行った。
(
3)
DRG神経細胞標本の調整
ラットは脊髄神経への手術
24時間後に二酸化炭素を用いて安楽死をさせた。脊柱を 含む後背部組織を一塊として切除後,実体顕微鏡下で脊髄を露出して
DRGを摘出した。
摘出した
DRGは,
collagenase(
3 mg/mL)及び
trypsin(
1 mg/mL)を含む
10%
Fetal Bovine Serum含有
Dulbecco’s Modified Eagle’s Medium溶液(
pH 7.4)で
37℃にて
60分間酵素処
理後,
trypsin inhibitorを添加して,パスツールピペットを用いて十分にピペッティング
して,ラット
DRG神経細胞標本を調整した。
(
4)共焦点レーザー顕微鏡によるラット
DRG神経細胞標本の画像解析
DRG
単一神経細胞は
Hanks緩衝塩類溶液中で,
cell tak処理したカバーグラス上に接 着させ
2 µM fluo-3/AM(励起波長
: 488 nm,蛍光波長
: 518 nm)を添加し,
37℃にて
5%
CO2インキュベーター内で
60分間
incubationすることにより,
Ca2+蛍光指示薬
fluo-3/AMを細胞内に取り込ませた。この
fluo-3/AM負荷
DRG神経細胞に,オレキシン
Aおよび リドカインを作用させた時の
[Ca2+]iの経時的・空間的変化を倒立型共焦点レーザー走査 顕微鏡システム(
TCS4D : Leica Ltd, Germany)を利用して観察した。
5
㸧
㸧>&D
㸰@Lࡢ ᐃ 㸦㸧 ᐃ᪉ἲ
༢㞳ࡉࢀࡓ
'5*ࡢ⣽⬊యࢆ࢞ࣛࢫ࣎ࢺ࣒㸦 PPȭ࣮ࢪ࣮ࢢࣜࢵࣉࢹࢵࢩࣗ
3RO\/O\VLQH㸪ᯇᾉ◪Ꮚओ㸪᪥ᮏ㸧ศὀࡋࡓࠋ)OXR$0
㈇Ⲵ
'5*༢୍⚄⤒⣽⬊ࡢ>&D
@Lࡢ㛫ⓗ࣭✵㛫ⓗኚࢆඹ↔Ⅼ࣮ࣞࢨ࣮㢧ᚤ㙾ࢆ⏝࠸࡚⏬ീゎᯒࡋࡓࠋࡲࡓ
'5*⚄⤒
⣽⬊⺯ගᣦ♧⸆ࡋ࡚
)XUD$0ࢆῧຍࡋ㸪
Υ࡚ศ㛫᧠ᢾࡋ࡞ࡀࡽ
LQFXEDWLRQࡋ࡚㸪⣽⬊ෆ
IXUDࢆ㈇Ⲵࡉࡏࡓ
'5*⚄⤒⣽⬊ࢆ⏝࠸࡚
&D) &D $QDO\]HU㸦᪥ᮏศග㸪᪥ᮏ㸧ࡼࡾ㸪>&D
@Lࢆ ᐃࡋࡓࠋ>&D
@Lࡣບ㉳Ἴ㛗
QP
QPࡢ Ἴ 㛗࡛Ⓨ⏕ࡍࡿ
QPࡢ⺯ගἼ㛗ࢆ ᐃࡋ㸪
))ࡢ
UDWLR್ࡼࡾ>&D
@Lࢆ⟬ฟࡋࡓࠋ ࡲࡓ
IXUD$0࠾ࡼࡧ
IOXR$0ࢆ㈇Ⲵࡋࡓ
'5*⣽⬊⣽⬊እᾮ
P0&DᏑᅾୗ
࡛
P0 .&Oࢆῧຍࡋ࡚>&D
@Lࡢୖ᪼ࢆほᐹࡋࡓࠋ.&O ฎ⨨ ⛊๓ࢽࣇ࢙ࢪࣆࣥ
㸦
î0î0㸧㸪ࣜࢻ࢝ࣥ㸦î0î0㸧㸪࠾ࡼࡧ࢜ࣞ࢟ࢩࣥ$
㸦
î0î0㸧ࢆస⏝ࡉࡏྠᵝグ㘓ࡋࡓࠋࡇࢀࡽ⸆≀ࡢ⁐፹ࡢࢪ࣓ࢳࣝࢫࣝ࣍࢟ࢩࢻ㸦'062㸧ࡣᑐ↷ࡋ࡚స⏝ࡉࡏࡓࠋ
㸦㸧⬺ศᴟᢚไ⋡ࡢ⟬ฟ
㧗
.่⃭ࡼࡿ>&D
@Lࡢ್᭱ࡼࡾ㸪ࡇࡢ>&D
@Lࡢ⸆≀ῧຍࡼࡿᢚไ⋡ࢆୗᘧࡼ
ࡾ⟬ฟࡋࡓࠋ
.&O
่⃭ᚋࡢ>&D
@Lࡣ㸪᭱㧗್ࡢᖹᆒࢆ᥇⏝ࡋࡓࠋ
㸧⤫ィゎᯒ
ᐇ㦂⤖ᯝࡣ㸪ᖹᆒ್ s ᶆ‽೫ᕪ㸦0HDQ s 6'㸧࡛⾲ࡋࡓࠋ6KDP ⩌㸦ᑐ↷⩌㸧࠾ࡼ
ࡧ
&$⩌ࡢ>&D
@Lࢆホ౯ࡍࡿࡓࡵ㸪) ᳨ᐃࡼࡾ➼ศᩓ࡛࠶ࡿࡇࢆ☜ㄆࡋࡓᚋ㸪 ᶆ
ᮏࡼࡿ
W᳨ᐃࢆ⾜ࡗࡓࠋ᭷ពỈ‽ࡣSࡋࡓࠋ
4
)結 果
Sham
群から脊髄
DRGを摘出・単離し,
24時間培養した細胞の画像を図
1に示した。
図
2は培養
7日目のこの細胞に
fluo-3/AMを負荷した後の共焦点レーザー顕微鏡像で,
神経突起の十分な発達が観察された。このことより,研究対象とした細胞が
DRG神経 細胞であることを確認した。
高
K+刺激前を図
3aに,高
K+刺激
2分後の
[Ca2+]i変化を図
3bに示した。高
K+刺激に より細胞膜周辺から核のある中心に向かい細胞全体に
[Ca2+]iの上昇が観察された。刺激 直後から
60秒で核周辺の
[Ca2+]iが高いがそれ以降はほぼ同一の濃度であった。次に高
K+刺激
30秒前にあらかじめリドカイン(
1.0 × 10-4 M)を作用させ同様に記録した。リ ドカイン投与時の
[Ca2+]i変化を図
3cに, 高
K+刺激
2分後の
[Ca2+]i変化を図
3dに示した。
リドカインは高
K+刺激による細胞膜および核周辺の
[Ca2+]i上昇を抑制した。
(
1)
Shamラット(対照群)
a.
高
K+刺激による
[Ca2+]i上昇作用に対するニフェジピンの影響
Sham
群の
fura-2/AM負荷
DRG神経細胞では,静止時の
[Ca2+]iは
60 nM付近で,高
K+(
50 mM)で脱分極させると,
220 nMへ急激に上昇した(図
4a) 。
50 mM KClによ る
[Ca2+]iの増加率は
223.5 ± 35.8%であった。細胞外液中
Ca2+非存在下(
1 mM EGTA添 加)では,
50 mM KCI脱分極による
[Ca2+]iの上昇は完全に消失した。また,
50 mM KClによる
[Ca2+]iの上昇は,ニフェジピン存在下で有意に抑制された(
p < 0.05) 。
1.0 × 10-5 M,
1.0 × 10-4 Mニフェジピンの抑制率はそれぞれ
64.6 ± 9.7%および
81.5 ± 13.1%であった
(図
4b,
6a) 。
b.
高
K+刺激による
[Ca2+]i上昇作用に対するリドカインの影響
高
K+刺激による
[Ca2+]iの上昇反応は,
1.0 × 10-4 M,
1.0 × 10-3 Mリドカイン処置の結果,
有意に抑制された(
p < 0.05) 。抑制率はそれぞれ
35.9 ± 5.2%および
84.1 ± 12.1%であっ た(図
4c,
6a) 。
c.
高
K+刺激による
[Ca2+]i上昇作用に対するオレキシン
Aの影響
高
K+刺激が誘発した
[Ca2+]iの上昇は,オレキシン
A(
1.0 × 10-4 M,
1.0 × 10-3 M)処置に よる目立った影響は認められなかった(図
4d,
6a) 。
(
2)
SNLラット
SNL
群の
fura-2/AM負荷
DRG神経細胞では,静止時の
[Ca2+]iは
80 nM付近で,高
KCl(
50 mM)で脱分極させると,
261 nMへ急激に上昇した。
50 mM KClによる
[Ca2+]iの上 昇の増加率は
243.4 ± 40.5%で,
sham群と比較するとやや高い傾向を示した(図
5a) 。 この
[Ca2+]iは細胞外液中
Ca2+非存在下では完全に消失した。
7
a.
高
K+刺激による
[Ca2+]i上昇作用に対するニフェジピンの影響
50 mM KCl
による
[Ca2+]iの上昇は,
1.0 × 10-5 M,
1.0 × 10-4 Mニフェジピン存在下で有 意に抑制された(
p < 0.05) 。抑制率はそれぞれ
67.1 ± 12.1%および
83.6 ± 11.1%で,
sham群とほぼ同じ値を示した(図
5b,
6b) 。
b.
高
K+刺激による
[Ca2+]i上昇作用に対するリドカインの影響
リドカイン前処置により,
50 mM KClによる
[Ca2+]iの上昇は有意に抑制された(
p <0.05
) 。
1.0 × 10-4 M,
1.0 × 10-3 Mリドカインの抑制率は,それぞれ
51.8 ± 9.1%および
77.3± 10.9%
であった(図
5c,
6b) 。
c.
高
K+刺激による
[Ca2+]i上昇作用に対するオレキシン
Aの影響
50 mM KCl
による
[Ca2+]iの上昇は
1.0 × 10-4 M,
1.0 × 10-3 Mオレキシン
A存在下で有 意に抑制された(
p < 0.05) 。抑制率はそれぞれ
16.2 ± 2.8%および
48.9 ± 9.8%であった(図
5d,
6b) 。
2. [
研究
2] CAラットを用いた研究
1)
CAラットの作製
研究
1と同様に
1週間の予備飼育をおこなった後,一般状態の観察および体重測定を 行い,順調に発育した体重
231.6 ± 17.5 gの動物を使用した。なお,本実験は日本大 学松戸歯学部実験動物委員会の承認(
AP14MD007号)を得て,日本大学松戸歯学部実 験動物指針に基づいておこなった。
CA
ラットの作製は,
Binghamら
10)の方法を基にして,
11週齢の
Wistar系雄性ラット を用いた。ラットを固定台に仰臥位で固定をし,両後肢の足蹠に
2%カラゲニン
0.25 mlを皮下投与した。投与後
24時間経過した個体を
CA群(
6匹)とした。
Control群(
6匹)は同様に溶媒の生理食塩液を投与した。なおそれぞれの群で
1匹ずつを免疫組織化 学染色に使用した。
2
)免疫組織化学染色
カラゲニンおよび生理食塩液投与後
24時間経過したラットは,ペントバルビタール ナトリウム
40 mg/kgにて腹腔内投与による深麻酔をおこなった後に灌流固定し
L4脊髄
と
L5 DRG神経細胞の摘出をおこなった。
免疫組織化学染色を行うため,既固定凍結切片をキシレンで脱パラフィンを行い,次 にエタノール及び
PBSを用いて再水和をした。抗原賦活化は,クエン酸緩衝液内にて
80
℃
pH 6.0の条件下で
40分間熱処理を行った。次にタンパク質ブロック(ジェノスタ
ッフ,日本)及びアビジン
/ビオチンブロックキット(ベクター,日本)と共に
0.3% H2O28
を含むメタノールで
30分間インキュベートし,内因性ペルオキシダーゼのブロックを おこなった。
切片を抗オレキシン
Aマウスモノクローナル抗体
10 μg/mL(
R&D Systems Inc., USA) もしくは抗
NMDAR抗体
2 µg/mL(
RNMDAB 763, Wako Pure Chemical Industries, Japan) を
4℃で一晩インキュベートした。インキュベート後,ビオチン結合抗ウサギ抗体(ダ コ・ジャパン,日本)を
400倍に希釈し室温で
30分間インキュベートした。ペルオキ シターゼ標識のストレプトアビジン(ニチレイ,日本)を室温にて
5分反応させた後,
基質に
DABを用いて発色させ,ヘマトキシリンにて対比染色を行った。その後切片を 脱水・透徹しマリノール(武藤化学,日本)にて封入した。
3
)
[Ca2+]iの測定
研究
1と同じ方法で摘出した
DRGは,
collagenase(
3 mg/mL) および
trypsin(
1 mg/mL) を含む
10% Fetal Bovine Serum(
FBS)含有
Hanks’ Balanced Salt Solution(
HBSS; pH 7.4) で
37℃にて
60分間酵素処理後,
FBS含有
HBSSで
3回洗浄して
DRG神経細胞標本を 調整した。
単離された
DRGの細胞体をガラスボトム(
35 mmφ
Glass Bottom Dishes Poly-d-lysinecoated, Mat Tek Co., USA
)に分注した。
DRG神経細胞に対してプロトコールに従い調整
した最終濃度
2.2 µM fluo-4/AMを添加し,
37℃にて
60分間インキュベートし,細胞内
に
fluo-4/AMを取り込ませた。この
fluo-4/AM負荷
DRG神経細胞に対して,共焦点レ
ーザー顕微鏡(
LSM5 Exciter: Carl Zeiss, Germany)を用いて
[Ca2+]iを測定した。算出し た
[Ca2+]i値から研究
1と,同様に薬物添加による抑制率を算出した。
4
)統計解析
実験結果は,平均値
±標準偏差(
Mean ± SD)で表した。
Sham群および
CA群の
[Ca2+]iを評価するため,
F検定により等分散であることを確認した後,
2標本による
t-検定を行った。有意水準は(
p < 0.05)とした。
5
)結 果
(
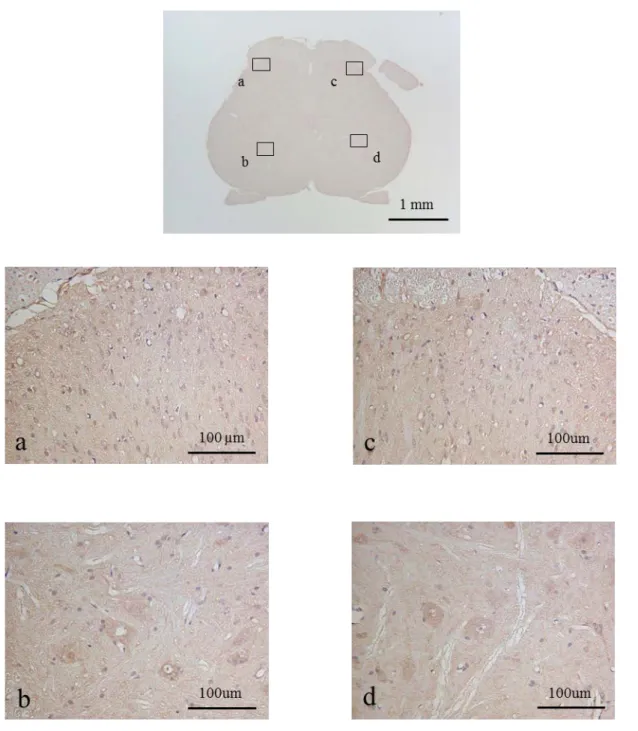
1)免疫組織化学染色
NMDAR
は脊髄の後角両側, および
DRGにおいて発現が認められた (図
7-9) 。
NMDARは前角においてわずかに発現を認めた。
Control群と
CA群を比較すると,
CA群の方が 濃染された細胞が全体的に多く認められた(図
9) 。
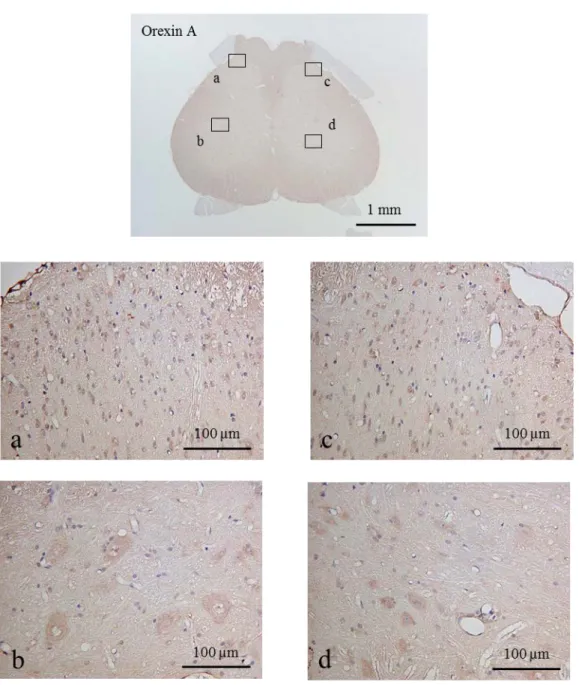
オレキシン
Aは脊髄の後角両側と
DRGにおいて発現がみられた (図
10-12) 。 しかし,
NMDAR
と比較すると濃染された細胞の個数は少なく,染色強度もそれほど高くはなか
った。
Control群と
CA群との差はほとんど認められなかった(図
12) 。
9
(
2)
[Ca2+]i測定
DRG
神経細胞に
fluo-4/AMを負荷し,
DRG神経細胞内の
[Ca2+]iを共焦点レーザー顕 微鏡でとらえた画像を図
13aに示す。細胞外液
1.3 mM Ca2+存在下で,高
K+刺激前を図
13bに示した。高
K+刺激により細胞内が緑から黄色へ急激に変化しており細胞膜周辺か ら核のある中心に向かい細胞全体に
[Ca2+]iの上昇が観察された。次に高
K+刺激
30秒前 にあらかじめオレキシン
A(
1.0 × 10-7 M)を作用させ同様に記録した。オレキシン
A投与時の
[Ca2+]i変化を図
13cに,高
K+刺激
2分後の
[Ca2+]i変化を図
13dに示した。オ レキシン
A非作用時に高
K+刺激により細胞全体は黄色への変化を認めたのに対して,
オレキシン作用時は全体的に緑色が強く脱分極の抑制が起きていることが視覚的に確 認された。
画像解析ソフト
ZEN 2011(
Carl Zeiss Micro Imaging Co. Ltd., Germany)により細胞単 位で
[Ca2+]iを計測した結果,
CA群の
fluo-4/AM負荷
DRG神経細胞では,静止時の
[Ca2+]i
は
70 nM付近で,高
K+(
25 mM)で脱分極させると,約
225 nMへ急激に上昇した
(図
14) 。また,
1.0 × 10-7 Mオレキシン
A存在下では
25 mM KCl脱分極による
[Ca2+]iの上昇は
control群と比較して
29.3 ± 14.4%抑制された(
p < 0.05:図
14, 15) 。
10
Ⅳ
.考 察
[
研究
1]では
SNLラットを用い,高
K+刺激が誘発した
DRG神経細胞の
[Ca2+]iの増加 に対するオレキシン
Aの作用について,
CaF-110 Ca2+ Analyzerおよび共焦点レーザー顕 微鏡(
TCS4D,
LSM5 Exciter)を用いて解析した。
1
)
SNLラット由来の
DRG神経細胞の高
K+刺激による
[Ca2+]i上昇作用に対するニフェ ジピンおよびリドカインの影響
Sham
群から得た
fura-2/AM負荷
DRG神経細胞では,高
K+刺激による
[Ca2+]iの増加
率は
223.5 ± 35.8%であった。この
[Ca2+]iはニフェジピンおよびリドカイン存在下で抑制
され,これらの薬物の
1.0 × 10-4 Mによる抑制率はそれぞれ
81.5 ± 13.1%および
35.9 ± 5.2%であった。また
1 mMの
EGTAを添加し細胞外液中の
Ca2+を除去した細胞外液中
Ca2+非存在条件下では,高
K+刺激が誘発した
[Ca2+]iの上昇は完全に消失した。これらの ことは,本実験条件下で
Ca2+は細胞膜に存在する電位依存性
L-type Ca2+チャネルを介し て細胞外から流入したことを示唆するものである。この
Ca2+流入は用量依存的にニフェ ジピンおよびリドカインにより遮断された。また
SNL群の
DRG神経細胞では,高
K+刺激による脱分極性
[Ca2+]iの増加率は
243.4 ± 40.5%であった。 この
[Ca2+]iの増加は
Sham群と同様にニフェジピンおよびリドカイン存在下で抑制され,両薬物の
1.0 × 10-4 Mに おける抑制率はそれぞれ
83.6 ± 11.1%および
51.8 ± 9.1%だった。これらの結果は,ラッ ト
DRG神経細胞の
L-type Ca2+チャネルを介した
Ca2+流入がリドカインにより用量依存 的に強く抑制されるとする
Whole-cell patch clamp法を用いた研究報告と一致するもの
である
12,13)。
2
)
SNLラット由来の
DRG神経細胞の高
K+刺激による
[Ca2+]i上昇作用に対するオレキ シン
Aの影響
オレキシン
A・
Bはともに視床下部および脊髄に広く分布している。オレキシン受容 体には
orexin 1 receptor(
OX1R)と
orexin 2 receptor(
OX2R)の
2種類のサブタイプが 知られており,オレキシン
Aは
OX1Rおよび
OX2Rに結合し,オレキシン
Bは
OX2Rに結合して効果を発揮する
1,2)。
DRG神経細胞にはオレキシン
Aと
OX1Rが存在し,侵 害刺激伝達の調節に関与していることが報告されている
10,14)。
DRG神経細胞の細胞膜 には脱分極により活性化する電位依存性
Ca2+チャネルが存在し,このチャネルを流入経 路として
[Ca2+]iは上昇し,その結果,神経伝達物質の放出が促進されて中枢に痛みが伝 達される。
SNLラットは神経損傷により,脊髄
DRG神経細胞内で産生される神経伝達 物質や
NK1受容体および
NMDARの発現が増加し,それが痛覚過敏反応の原因になっ
11
ていると考えられている
15)。
桜井らは,オレキシン
Aは
OX1R刺激を介して細胞内ストアを由来とする
[Ca2+]iの増 大を起こすことを報告している
6)。本研究からは対照群とは異なり
SNLラットの
DRG神経細胞では高
K+による脱分極性刺激で上昇する
[Ca2+]iがオレキシン
A投与により有 意に抑制されることが示された。その一因としてオレキシン
Aが,
SNLラットの
DRG神経細胞において細胞膜上の
L-type Ca2+チャネルを介した細胞外からの
Ca2+流入の抑 制を起こす可能性が考えられた。オレキシン
Aは全身投与,脳室内または髄腔内投与 が鎮痛効果を示すことが動物実験から報告されている
10,16-19)。例えば
Binghamら
10)は,
オレキシン
Aをマウスに静脈投与すると熱刺激疼痛とカラゲニン誘発の炎症性痛覚過 敏を抑制すること示している。さらに,オレキシン
Aはラットの脳内への
microinjectionでも熱刺激疼痛に対する鎮痛効果が認められる
16,17)。神経障害性疼痛モデル動物では,
内因性オレキシン
Aの産生促進と
OX1Rの活性化が関わる鎮痛機構も報告されてい
る
18,19)。以上のことから,本研究で観察されたオレキシン
Aによる
DRG神経細胞の
[Ca2+]i
の増加の抑制作用が,行動学的実験から示されているオレキシン
Aの鎮痛作用の 発現に関与する可能性が考えられた。
研究
2では
CAラットを用い,
DRG神経細胞におけるオレキシン
Aの発現を免疫組 織化学的に検索するとともに,高
K+刺激が誘発した
DRG神経細胞の
[Ca2+]iの増加に対 するオレキシン
Aの作用について,共焦点レーザー顕微鏡を用いて解析した。
1
)
NMDARとオレキシン
Aの
CAラット由来の脊髄および
DRG神経細胞での発現
NMDAR
は,
DRGを含む脊髄において痛みに関連する神経伝達に関わるグルタミン
酸の受容体である。
DRGの
NMDARは,実験動物の足蹠へプロスタグランジン
E2を投 与すると増加することを
Ferrariらは指摘している
20)。本研究の
DRG神経細胞の免疫組 織染色では,
NMDAR陽性細胞は対照群に比べ
CAラットの方が多い傾向が認められ た。このことは
CA群の
DRGではグルタミン酸神経伝達が亢進している可能性を示唆 するものである。
Bingham
らは,オレキシン
Aと
OX1Rが
DRG神経細胞に分布していることを報告し
ている
10)。本研究でも,脊髄と
DRG神経細胞でオレキシン
A陽性細胞は,対照群と
CAラットのいずれでも認められた。一方で,
NMDAR陽性細胞の場合とは異なり,オ レキシン
A陽性細胞の染色強度には対照群と
CAラットとの間で目立った違いが認めら れなかった。
2
)
CAラット由来の
DRG神経細胞の高
K+刺激による
[Ca2+]i上昇作用に対するオレキ シン
Aの影響
研究
2では
CAラットを用い,研究
1とほぼ同様の方法で高
K+による
DRG神経細胞
12の
[Ca2+]i上昇の測定を行った。研究
1で高
K+による
DRG神経細胞の
[Ca2+]i上昇は,対 照群ではオレキシン
Aの影響を受けなかったのに対し,
SNLラットではオレキシン
Aで強く抑制された。研究
2でもオレキシン
Aの効果に同様の傾向が認められ,高
K+に よる
DRG神経細胞の
[Ca2+]i上昇は,対照群ではオレキシン
Aの影響を受けなかったの に対し,
CAラットではオレキシン
Aで減弱した。このことは
SNLラットと同様に
CAラットでも,オレキシン
Aが
DRG神経細胞において脱分極に伴う細胞膜上の
L-type Ca2+チャネルを介した
Ca2+流入の抑制を起こす可能性を示唆するものである。つまりオ レキシン
Aによるこの細胞内
Ca2+流入の低下が,痛みに関連する神経伝達物質の
DRGにおける合成や放出の減少に関与する可能性が考えられた。また,神経障害性疼痛のみ ならず炎症性疼痛もオレキシン
Aが
DRG神経細胞に作用して抑制する可能性も示唆さ れた。一方で,対照群と
CAラットとの間でオレキシン
Aの効果に違いが起きた原因は 明らかでない。しかし免疫組織染色の結果,オレキシン
A陽性細胞は対照群でも
CA群 とほぼ同程度認められたことから,少なくとも
DRGにおけるオレキシン
Aの発現が対 照群と
CAラットで著しい差があった可能性は考え難い。神経障害性疼痛モデル動物で は,
OX1Rの活性化が関わる鎮痛機構が報告されている
10)。オレキシン
Bは
OX1Rで はなくむしろ
OX2Rを選択的に刺激するため,本研究は
OX1Rのみならず
OX2Rも刺 激しうるオレキシン
Aを用いた。 研究
1および研究
2で認められた高
K+が誘発した
DRGの
[Ca2+]i上昇のオレキシン
Aによる抑制が起こるメカニズムについて,オレキシン受容 体サブタイプの関与の面からさらなる検討が必要である。
Ⅴ
.結 論
以上の研究
1および研究
2から,オレキシン
Aは生理的状態のラットの
DRG神経細 胞への高
K+刺激が誘発した
[Ca2+]iの増加には影響を与えないことに対し,疼痛モデルラ ットの
DRG神経細胞への高
K+刺激が誘発した
[Ca2+]iの増大は抑制することが示された。
13
Ⅵ
.参考文献
1) Kim SH, Chung JM: An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain, 50: 355-363, 1992.
2) Selzer Z, Dubner R, Shir Y. A novel behavioral model of neuropathic pain disorders produced in rats by partial sciatic nerve injury. Pain, 43: 205-218, 1990.
3) Decosterd I, Woolf CJ: Spared nerve injury: an animal model of persistent peripheral neuropathic pain. Pain, 87: 149-158, 2000.
4) Campbell JN, Meyer RA: Mechanisms of neuropathic pain. Neuron, 52: 77-92, 2006.
5) Sakurai T: The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness.
Nat Rev Neurosci, 8: 171-181, 2007.
6) Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H, Williams SC, Richardson JA, Kozlowski GP, Wilson S, Arch JR, Buckingham RE, Havnes AC, Carr SA, Annan RS, McNulty DE, Liu WS, Terrett JA, Elshourbagy NA, Bergsma DJ, Yanagisawa M: Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell, 92: 573-585, 1998.
7) Cheng JK, Chou RC, Hwang LL, Chiou LC: Antiallodynic effects of intrathecal orexins in a rat model of postoperative pain. J PharmacolExpTher, 307: 1065-1071, 2003.
8) Mobarakeh JI, Takahashi K, Sakurada S, Nishino S, Watanabe H, Kato M, Naghdi N, Yanai K: Enhanced antinociception by intracerebroventricularly administered orexin-A in histamine H1 or H2 receptor gene knockout mice. Pain, 118: 254-262, 2005.
9) Yazdi-Ravandi S, Razavi Y, Haghparast A, Goudarzvand M, Haghparast A: Orexin-A induced antinociception in the ventral tegmental area involves D1 and D2 receptors in the nucleus accumbens. Pharmacol Biochem Behav, 126: 1-6, 2014.
10) Bingham S, Davey PT, Babbs AJ, Irving EA, Sammons MJ, Wyles M, Jeffrey P, Cutler L, Riba I, Johns A, Porter RA: Upton N, Hunter AJ, Parsons AA: Orexin-A, an hypothalamic peptide with analgesic properties. Pain, 92: 81-90, 2001.
11) Tomoyuki K, Keiichi O, Teruyoshi N: How to make the animal models of chronic pain.
Pain Clinic, 30: S73-S79, 2009.
12) Stühmer W, Conti F, Suzuki H, Wang XD, Noda M, Yahagi N, Kubo H, Numa S: Structural parts involved in activation and inactivation of the sodium channel. Nature, 339: 597-603, 1989.
13) Scholz A, Kuboyama N, Hepelmann G, Vogel W: Complex blockade of TTX-resistant Na+
currents by lidocaine and bupivacaine reduce firing frequency in DRG neurons.J Neurophysiol, 79: 1746-1754, 1998.
14) Hervieu GJ, Cluderay JE, Harrison DC, Roberts JC, Leslie RA: Gene expression and 14
protein distribution of the orexin-1 receptor in the rat brain and spinal cord. Neuroscience, 103: 777-797, 2001.
15) Malmberg AB, Basbaum AI: Partial sciatic nerve injury in the mouse as a model of neuropathic pain: behavioral and neuroanatomical correlates. Pain, 76: 215-222, 1998.
16) Jordt SE, Julius D: Molecular basis for species-specific sensitivity to "hot" chili peppers.
Cell, 108: 421-430, 2002.
17) Watson SL, Watson CJ, Baghdoyan HA, Lydic R: Thermal nociception is decreased by hypocretin-1 and an adenosine A1 receptor agonist microinjected into the pontine reticular formation of Sprague Dawley rat. J Pain, 11: 535-544, 2010.
18) Suyama H, Kawamoto M, Shiraishi S, Gaus S, Kajiyama S, Yuge O: Analgesic effect of intrathecal administration of orexin on neuropathic pain in rats. In Vivo, 18: 119-123, 2004.
19) Yamamoto T, Saito O, Shono K, Aoe T, Chiba T: Anti-mechanical allodynic effect of intrathecal and intracerebroventricular injection of orexin-A in the rat neuropathic pain model. Neurosci Lett, 347: 183-186, 2003.
20) Ferrari LF, Lotufo CM, Araldi D, Rodrigues MA, Macedo LP, Ferreira SH, Parada CA:
Inflammatory sensitization of nociceptors depends on activation of NMDA receptors in DRG satellite cells. Proc Natl Acad Sci U S A. 111: 18363-18368, 2014.
15
Ϯ ᅗ࠾ࡼࡧ⾲
ᅗ
ᅗ
&HOOVGHULYHGIURPUDW'5*
ᅗ
ᅗ
3ULPDU\FXOWXUHG'5*QHXURQVZLWKIOXR$0
ᅗ
ᅗ
.&OLQGXFHG LQFUHDVH RI >&D@L UDW LQ '5* QHXURQV H[DPLQHG E\ FRQIRFDO ODVHU VFDQQLQJPLFURVFRSH7KHLPDJHVRIDDQGEVKRZVEHIRUHDQGDIWHU.&OWUHDWPHQWUHVSHFWLYHO\
7KH LPDJHV RI F DQG G VKRZHV HIIHFWV RI OLGRFDLQH RQ EDVDO F DQG WKH .&OLQGXFHG RI
>&D@LG
6KDPJURXS
ᅗ
ᅗ
7LPHFRXUVHRI.&OLQGXFHG>&D@LLQFUHVHLQ'5*QHXURQVRIVKDPUDWVD.&OLQGXFHGLQFUHDVHRI>&D@L7KHDUURZVLQGLFDWHWLPLQJRI'062DQG.&OWUHDWPHQWV E(IIHFWVRIQLIHGLSLQHRQ.&,LQGXFHG>&D@LLQFUHDVH
7KHDUURZVLQGLFDWHWLPLQJRIQLIHGLSLQHDQG.&OWUHDWPHQWV F(IIHFWVRIOLGRFDLQHRQ.&,LQGXFHG>&D@LLQFUHDVH 7KHDUURZVLQGLFDWHWLPLQJRIOLGRFDLQHDQG.&OWUHDWPHQWV G(IIHFWVRIRUH[LQ$RQ.&,LQGXFHG>&D@LLQFUHDVH 7KHDUURZVLQGLFDWHWLPLQJRIRUH[LQ$DQG.&OWUHDWPHQWV
61/JURXS
ᅗ
ᅗ
7LPHFRXUVHRI.&OLQGXFHG>&D@LLQFUHVHLQ'5*QHXURQVRI61/UDWVD.&OLQGXFHGLQFUHDVHRI>&D@L7KHDUURZVLQGLFDWHWLPLQJRI'062DQG.&OWUHDWPHQWV E(IIHFWVRIQLIHGLSLQHRQ.&,LQGXFHG>&D@LLQFUHDVH
7KHDUURZVLQGLFDWHWLPLQJRIQLIHGLSLQHDQG.&OWUHDWPHQWV F(IIHFWVRIOLGRFDLQHRQ.&,LQGXFHG>&D@LLQFUHDVH 7KHDUURZVLQGLFDWHWLPLQJRIOLGRFDLQHDQG.&OWUHDWPHQWV G(IIHFWVRIRUH[LQ$RQ.&,LQGXFHG>&D@LLQFUHDVH 7KHDUURZVLQGLFDWHWLPLQJRIRUH[LQ$DQG.&OWUHDWPHQWV
ᅗ
ᅗ
6 Inhibitory effects of drugs on K+-induced [Ca2+]i increase in DRG neurons derived from sham (a) and SNL (b) rats.The values were expressed as mean ± SD (n = 5/group).
図
7 NMDAR immunohistochemical staining of the spinal cord at the L4 level in the control group.22
図
8 Immunohistochemical staining of NMDAR in the spinal cord at the L4 level of CA group.23
ᅗ
ᅗ
$UURZVLQGLFDWH10'$5SRVLWLYHFHOOV,PPXQRKLVWRFKHPLFDOVWDLQLQJRI10'$5 LQ'5*QHXURQV
図
10 Immunohistochemical staining of orexin-A in the spinal cord at the L4 level of control group.25
図
11 Immunohistochemical staining of orexin-A in the spinal cord at the L4 level of CA group.26
ᅗ
ᅗ
$UURZV LQGLFDWH RUH[LQ$ SRVLWLYH FHOOV ,PPXQRKLVWRFKHPLFDO VWDLQLQJ RI 10'$5LQ'5*QHXURQV
ᅗ
ᅗ
,PDJLQJDQDO\VLVRI.&OLQGXFHGGHSRODUL]DWLRQDQGWKH>&D@LLQFUHDVHLQDVLQJOH UDW'5*QHXURQH[DPLQHGZLWK&RQIRFDOODVHUVFDQQLQJPLFURVFRS\ 'LVWULEXWLRQRI>&D@LLQDVLQJOHUDW'5*QHXURQEHIRUHDDQGDIWHUEVWLPXODWLRQZLWK P0.&O'LVWULEXWLRQRI>&D@LLQDVLQJOHUDW'5*QHXURQEHIRUHFDQGDIWHUGVWLPXODWLRQ ZLWKP0.&ODQGDGGLWLRQRIRUH[LQ$
ᅗ
ᅗ
(IIHFWVRIRUH[LQ$RQ.&OLQGXFHGGHSRODUL]DWLRQDQGWKH>&D@LLQFUHDVHLQWKH FDUUDJHHQDQDQGFRQWUROJURXSV 2UH[LQ$KDGOLWWOHRUQRHIIHFWRQWKHKLJK.LQGXFHGGHSRODUL]DWLRQDQGWKH>&D@LLQFUHDVH LQ WKH SUHVHQFH RI P0 H[WUDFHOOXODU &D LQ FRQWURO QHXURQV D $ KLJK FRQFHQWUDWLRQ RI RUH[LQ$ î í0 LQKLELWHG RSHQLQJ RI YROWDJHGHSHQGHQW &DFKDQQHOV H[SUHVVHG LQ FDUUDJHHQDQUDWVE
ᅗ
ᅗ
(QKDQFHGLQKLELWRU\HIIHFWVRIRUH[LQ$RQWKH.LQGXFHGLQFUHDVHLQ>&D@LLQD '5*QHXURQIURPWKHFDUUDJHHQDQWUHDWHGJURXSQ S