http://repository.osakafu-u.ac.jp/dspace/
Title
定量的RT-PCR法によるヒト常在細菌叢の解析Author(s)
倉川, 尚Editor(s)
Citation
Issue Date
2015URL
http://hdl.handle.net/10466/14608Rights
大阪府立大学博士(獣医学)学位論文
定量的 RT-PCR 法によるヒト常在細菌叢の解析
倉川 尚
目次
第
1 章 ヒト常在細菌叢とその解析技術…………..………1
第
2 章
Clostridium coccoides group の詳細な定量測定系の構築および応用
序論………..7
材料および方法………..8
結果………14
考察………16
図表………19
第
3 章 ヒト腸内における腸内細菌科細菌の菌種構成の解析
序論………26
材料および方法………26
結果………29
考察………30
図表………32
第
4 章 腸管病原細菌の高感度定量システムの構築
序論………36
材料および方法………36
結果………39
考察………41
図表………43
第
5 章 定量的 RT-PCR 法の膣内細菌叢解析への応用
序論………52
材料および方法………53
結果………56
考察………58
図表………61
総括……….………68
引用文献……….………70
謝辞……….………90
略語一覧
AAHC Antibiotic-associated hemorrhagic colitis
AFLP Amplified fragment length polymorphism
BHI Brain Heart Infusion
BV Bacterial vaginosis
Cq Quantification cycle
CFU Colony forming unit
DAPI 4’, 6-diamidino-2-phenylindole
DMEM Dulbecco's modified eagle medium
DNA Deoxyribonucleic acid
dNTP Deoxyribonucleotide triphosphate
EB Elementary body
EDTA Ethylenediamine tetraacetic acid
ELISA Enzyme-linked immunosorbent assay
FBS Fetal bovine serum
FISH Fluorescence in situ hybridization
FMT Fecal microbiota transplantation
IBD Inflammatory bowel desease
IFU Inclusion forming unit
IPTG Isopropyl--thiogalactopyranoside
NGS New generation sequencer
PBS Phosphate buffered saline
PCR Polymerase chain reaction
qPCR Quantitative PCR
RB Reticulate body
RNA Ribonucleic acid
rRNA Ribosomal RNA
RT-qPCR Quantitative RT-PCR
SDS Sodium dodecyl sulfate
SPG Sucrose-phosphate-glutamate buffer
TE Tris-EDTA
T-RFLP Terminal Restriction Fragment Length Polymorphism
VBNC Viable but non-culturable
1
第
1 章 ヒト常在細菌叢とその解析技術
宿主と細菌の関わり
ヒトの腸, 膣, 皮膚, 口腔, 咽頭等の各部位には, 多種多様な細菌が棲息しており, 常在 細菌叢を構成している [1]。 これらの常在細菌は, 宿主と共存共栄関係にあるが, 何らかの きっかけで常在細菌叢が破綻すると, 宿主の健康に悪影響を及ぼす。 例えば, 腸内細菌叢 のバランスが崩れると, 下痢, 便秘, 過敏性腸症候群等の機能異常や, 炎症性腸疾患 (IBD), 大腸癌等の疾患を引き起こし [2-4], 膣内細菌叢の破綻は, 細菌性膣炎 (BV) の原因となる [5]。また, 病原性を有する細菌が宿主に感染すると様々な病気を引き起こす。 コレラ菌 (Vibrio cholerae), 腸炎ビブリオ (Vibrio parahaemolyticus), およびカンピロバクター(Campylobacter jejuni ) 等の腸管病原細菌は, 腸管内で感染し、毒素を産生するなど種々
のメカニズムにより腸炎を引き起こす [6-8]。 クラミジア (Chlamydia trachomatis) は,
性器に感染することにより子宮頸管炎を引き起こす [9]。さらに, 普段は宿主に危害を与え ないが, 免疫力が低下したときに増殖し, 宿主に病気を引き起こす常在細菌 (日和見感染
菌) も存在する。大腸菌 (Escherichia coli ) や緑膿菌 (Pseudomonas aeruginosa) 等が日
和見感染菌として知られている [10,11]。 一方で, 細菌叢を改善することにより, 宿主の健 康状態が回復することも知られている。 その典型例がプロバイオティクス (宿主に有益な 作用をもたらす生きた微生物) である。 Lactobacillus属菌やBifidobacterium属菌等のプ ロバイオティクスは腸内環境を改善し, IBD の治療にも効果を発揮する [12]。 また, 健康 な 人 の 細 菌 叢 を 移 植 す る こ と に よ り 病 気 を 治 療 す る 試 み (Fecal microbiota transplantation; FMT) も近年注目されている [13]。 このように, 常在細菌叢は宿主と密 接な関係を有し, これを正常に維持することが宿主の健康の維持につながる。
ヒト腸内細菌叢
ヒトの腸内には 1,000 種類以上, 約 1014個の細菌が棲息しており、複雑な微生物生態系 を形成している [14]。 腸内細菌叢は免疫系の成熟 [15], ビタミン等の必須栄養素の合成 [16], 難消化性多糖の分解 [17], 病原菌の定着阻害 [18] 等の有益な生理作用を有する。 ヒ2
トの腸内細菌叢の形成は出生直後から始まる。 出生後まもなく, Escherichia 属菌や
Streptococcus属菌等の通性嫌気性菌群が生着し増加するが, 出生して約 1 か月後に, これ
らは偏性嫌気性菌群に置き換わる [19]。 ヒト腸内細菌叢を高い菌数レベルで占有する最優
勢 嫌 気 性 菌 群 と し て, Clostridium coccoides group, Clostridium leptum subgroup,
Bacteroides fragilis group, Bifidobacterium属菌, Atopobium cluster およびPrevotella属
菌の6 菌群が知られている [20]。 松木らは, 健常日本人の糞便の解析を行い, これら 6 菌
群のうち, C. coccoides group が腸内細菌叢の 29%を占めており, 続いて C. leptum
subgroup が 15%, B. fragilis group が 11%を占めていることを報告している [20]。 腸内細
菌叢に最も大きな影響を及ぼす因子は食事であり, 食習慣の異なる地域では, 全く異なる 腸内細菌叢となっている [21]。例えば, 高炭水化物および高繊維食を中心とするアフリ カ・ブルキナファソに居住する子供では, Prevotella属菌が腸内細菌叢の50%以上を占めて おり, 高タンパクおよび高脂肪食を摂取する西洋都市部に居住する子供とは大きく異なる 細菌構成となっている [22]。 マウスを用いた実験では, 脂肪, 人工甘味料, 乳化剤等の食 餌成分の違いが, 腸内細菌叢の構成に影響を与えることが示唆されている [15,23,24]。 ま た, 抗生物質投与によりE. coliを代表とするProteobacteria 門の細菌が増加すること [25], 喫煙者と非喫煙者では腸内細菌叢が異なっており, 喫煙者が禁煙することにより腸内細菌 叢が変化すること [26], スポーツ選手では腸内細菌叢の多様性が大きいこと [27] 等が報 告されており, 食事以外にも様々な環境要因が腸内細菌叢に影響を与えていることが示唆 されている。 さらに, 高齢者で E. coli が増加すること [28] , 男性では女性と比較して Bacteroides 属菌の菌数が高いこと [28] が報告されており, 年齢および性別等の遺伝的要 因も腸内細菌叢に影響を及ぼすと考えられている。 腸内細菌叢による代謝産物は, 腸のみにとどまらず全身に影響を与えている。例えば, Firmicutes 門の細菌が増加すると食品からのエネルギー摂取効率が高まり肥満になりやす くなること [29], 腸内細菌叢の多様性が低い乳児はアレルギー疾患に罹患しやすいこと [30], 腸内細菌の代謝産物であるトリメチルアミン-N-オキシドが動脈硬化を誘導すること [31], さらにPrevotella属菌およびCoprococcus属菌の菌数の減少と腸内細菌叢の多様性の 低下が自閉症と関与していること [32] 等が報告されている。 このように, 腸内細菌叢の 影響は多岐にわたっており, そのメカニズムを解明するためにも, 腸内細菌叢を正確に把 握することが極めて重要である。
3
細菌叢の解析技術
細菌叢解析の黎明期には, 19 世紀にロベルト・コッホによって開発された分離培養法が 用いられてきた。 本手法は, 様々な選択培地上に細菌をコロニーとして増殖させ, 得られ た細菌の形状や生化学的性状をもとに細菌種を判別し, 各細菌種を定量する方法である。 嫌気チャンバーおよび多種類の選択培地の開発により, 培養技術が著しく向上したが, そ れでもなお培養困難な細菌が多く存在するうえ, 培養操作には多大な労力と洗練された技 術が必要であり, 生化学的性状のみでは菌種同定が困難な細菌が存在する等, 多くの問題 点があった。 このような状況のもと, Polymerase Chain Reaction (PCR) 法 [20],Fluorescence in situ hybridization (FISH) 法 [33], Terminal Restriction Fragment
Length Polymorphism (T-RFLP) 法 [34], クローンライブラリー法 [35] 等, 細菌に普遍
的に存在するRibosomal RNA (rRNA) 遺伝子の塩基配列の違いを利用して、細菌叢を解析
する手法が開発された。 PCR 法は, 標的細菌に特異的なプライマーを用いることにより, 標的細菌を定性的または定量的に解析する手法である。 FISH 法は, 蛍光標識した標的細 菌特異的プローブを用いることにより, 標的細菌を定性的または定量的に解析する手法で ある。 T-RFLP 法は, 全細菌由来の DNA を PCR により増幅した後, 制限酵素処理によっ て生じる菌特異的 DNA 多型を検出する手法である。 クローンライブラリー法は, 細菌の 16S rRNA 遺伝子の共通領域を標的としたプライマーを用いて PCR を行い, 得られた増幅 産物の塩基配列をもとに菌種構成を解析する手法である。これらの分子生物学的手法は, 培 養を介さないことから, 煩雑な手間を省力化でき, 培養困難な細菌の定量も容易となった。 しかし, これらの手法では検出感度に限界があり, サブドミナントに棲息する菌群を検出 できないという欠点がある。 さらに近年, 次世代シークエンサー (NGS) の発展により, 網 羅的な細菌叢解析が可能となった [22,36]。 NGS はクローンライブラリー法と同様の原理 で細菌種の構成を解析する手法であるが、大規模な塩基配列解析により, 詳細な菌種構成情 報を得ることができる。 また, NGS はメタゲノム解析による機能遺伝子の解析も可能であ り, 腸内細菌叢の機能に関わる研究にも利用されている [37]。 しかし, NGS により得られ る細菌叢データは, 個々の細菌種の割合であり, 個々の細菌数ではない。 正確な細菌叢の
4 把握のためには, 多種多様な細菌の数を正確かつ高感度に定量する方法の確立が必要であ る。
腸内細菌叢解析としての定量的
RT-PCR 法およびその応用
腸内細菌叢解析における細菌数定量法として, 近年, 16S または 23S rRNA 分子を標的と した定量的RT-PCR 法 (RT-qPCR) が開発された [38]。 本手法の特長として, 1 細胞内に 約10,000 分子存在する rRNA を標的とするため, 細菌 1 ゲノムあたり数コピーしか存在 しないrRNA 遺伝子を標的とする定量的 PCR 法 (qPCR) と比較して, 約 1,000 倍高い検出 感度で標的細菌を定量可能であることが挙げられる。 この特長により, これまで qPCR で は検出が困難であった腸内細菌科細菌および緑膿菌等, 腸内においてサブドミナントに存 在する菌群の定量が可能となった [38]。 このような利点から, 本手法は腸内常在菌の解析 のみにとどまらず, 様々な臨床研究に応用されている。 例えば, 発熱性好中球減少症患者 の血液や, 胆道がん患者の手術後の腸管膜リンパ節を対象とした細菌検出に, RT-qPCR が 適用可能であることが示されている [39,40]。 本手法が2007 年に初めて報告されて以来, 様々な菌群および菌種特異的プライマーが作 製され, 本手法に適用されてきた (Table 1-1) [38,40-43]。 しかし, rRNA という限られた 長さの配列を標的とするため, 特異的プライマーの作製が困難な場合もあり, 腸内細菌叢 の構成細菌種を完全に網羅できるには至っていない。 このような状況のもと, 本研究では ヒト腸内細菌叢で最優勢に棲息する菌群の1 つであるC. coccoides group に焦点を当て, こ の菌群を詳細に解析するためのサブグループおよび菌種特異的プライマーの作製を試みた。 そして, これらのプライマーを用いて, 健常幼児, 成人および高齢者の腸内における本菌群 の分布を詳細に解析した。 次に, これまで正確な定量が困難であったE. coliに特異的なプ ライマーを作製し, 腸内における本菌の定量を試みた。 さらに, RT-qPCR とクローンライ ブラリー法を組み合わせることにより, ヒト腸内における腸内細菌科細菌の詳細な解析を 行った。 また, 代表的な腸管病原細菌に特異的なプライマーを作製することで, RT-qPCR を高感度な腸管病原細菌診断へ応用した。 最後に, RT-qPCR の膣内細菌叢解析への応用を 試みた。 膣内は腸内とは異なった細菌叢を形成しているが, これまで研究がほとんど進ん5
でいなかった。そこで, 膣に固有の菌種を標的とした特異的プライマーを作製し, これらを
6
Target Representative species Targeted rRNA Reference
Atopobium cluster
Bacteroides fragilis group B. fragilis, B. ovatus , B. thetaiotaomicron , B. uniformis , B. vulgatus 16S [42]
Bifidobacterium B. breve, B. bifidum, B. catenulatum, B. dentium 16S [42]
Clostridium coccoides group
Clostridium difficile C. difficile 23S [43]
Clostridium leptum subgroup C. leptum, Faecalibacterium prausnitzii , Ruminococcus albus 16S [42]
Clostridium perfringens C. perfringens 16S [38]
Enterobacteriaceae
Enterococcus E. avium, E. caccae, E. casseliflavus, E. cecorum, E. faecalis 16S [38]
Enterococcus avium subgroup E. avium, E. pseudoavium, E. malodoratus, E. gilvus, E. raffinosus 23S [41]
Enterococcus caccae E. caccae 16S [41] Enterococcus casseliflavus subgroup E. casseliflavus, E. gallinarum, E. flavescens 16S [41]
Enterococcus cecorum E. cecorum 16S [41]
Enterococcus dispar E. dispar 16S [41]
Enterococcus faecalis E. faecalis 16S [41] Enterococcus faecium E. faecium 16S [41] Enterococcus faecium subgroup E. faecium, E. durans, E. mundtii, E. hirae 23S [41] Enterococcus sulfureus subgroup E. sulfureus, E. italicus 16S [41]
Lactobacillus brevis L. brevis 16S [42]
Lactobacillus casei subgroup L. casei, L. rhamnosus, L. zeae 16S [42]
Lactobacillus fermentum L. fermentum 16S [42]
Lactobacillus fructivorans L. fructivorans 16S [42]
Lactobacillus gasseri subgroup
Lactobacillus plantarum subgroup L. plantarum, L. pentosus 16S [42]
Lactobacillus reuteri subgroup L. reuteri, L. oris, L. panis, L. vaginalis 16S [42]
Lactobacillus ruminis subgroup L. ruminis, L. animalis, L. mali, L. salivarius, L. satsumensis 16S [42]
Lactobacillus sakei subgroup L. sakei, L. curvatus, L. graminis 16S [42]
Lactococcus lactis subgroup L. lactis, L. garvieae 16S [41]
Lactococcus pisium subgroup L. pisium, L. plantarum, L. raffinolactis 16S [41]
Prevotella P. buccae, P. intermedia, P. melaninogenica, P. oralis 16S [42]
Pseudomonas P. aeruginosa, P. putida 16S [38]
Staphylococcus S. aureus, S. epidermis, S. schleiferi 16S [38]
Streptococcus
Streptococcus agalactiae S. agalactiae 16S [40] Streptococcus pneumoniae/ mitis S. pneumoniae, S. mitis 23S [40] Streptococcus pyogenes S. pyogenes 16S [40] Streptococcus salivarius/ thermophilus S. salivarius, S. thermophilus 16S [41]
S. agalactiae, S. pneumoniae, S. mitis, S. pyogenes, S. salivarius,
S. thermophilus 23S [40]
Escherichia coli, Enterobacter cloacae, Proteus mirabilis,
Klebsiella pneumonieae, Serratia marcescens 23S [38]
L. gasseri, L. jensenii, L. johnsonii, L. crispatus, L. acidophilus,
L. iners 16S [42]
A. parvulum, Collinsella aerofaciens , C. intestinalis , C. stercoris ,
Eggerthella lenta 16S [42]
Blautia producta, Eubacterium hallii , C. symbiosum,
Roseburia intestinalis, Ruminococcus lactaris , Coprococcus eutactus 16S [42]
7
第
2 章
Clostridium coccoides group の詳細な定量測定系の構築および応用
1. 序論
C. coccoides group は, 偏性嫌気性菌から構成されるグループであり, ヒト腸内細菌のう
ち, 25~60%を占めると言われている [20,44-46]。 本グループには, Clostridium 属,
Blautia属, Dorea属, Eubacterium属, Ruminococcus属, Anaerostipes属, Roseburia属お
よび Coprococcus 属等の細菌が含まれる [44,46]。 また, 本グループには多くの難培養細
菌が含まれていることが, クローンライブラリー解析により明らかとなっている [35]。 近
年においても培養技術の発展に伴い, Fusicatenibacter saccharivorans等, 本グループの新
たな菌種の分離報告が相次いでいる [47]。
C. coccoides group に属する細菌は, 宿主に様々な影響を与えることが知られている。
Eubacterium rectale, Eubacterium hallii, Eubacterium ramulus, Roseburia intestinalis
およびAnaerostipes caccaeは, 酪酸産生菌として知られている [48,49]。 腸内で産生され た酪酸は宿主のエネルギー源になる他, 腸管の粘膜バリア機能の強化や抗炎症作用に関わ
っている [50]。 Clostridium scindensおよびClostridium hylemonaeは, 胆汁酸 7-脱水
酸化能を有しており, デオキシコール酸やリトコール酸等、宿主に有害な二次胆汁酸を産生 し, 大腸癌を誘導する [51]。 さらに, ある種のC. coccoides group 菌種は, マウス腸管内 において免疫応答の恒常性維持に重要な制御性 T 細胞の分化を誘導することが知られてい る [52]。 以上のことから, C. coccoides group は, 栄養源の供給, 有害物質の産生, 免疫機 能の恒常性等に影響を与えており, 宿主の健康と密接な関係を有していると考えらえる。 このように, C. coccoides group は腸内において高い占有率を有するだけでなく, 生理学的 に重要な細菌群であるが, ヒトの腸内における詳細な構成は明らかとなっていない。
本研究では, 16S rRNA 配列に基づいてC. coccoides group を便宜的に 3 サブグループお
よび19 菌種に系統化し, それぞれに特異的なプライマーを新規に構築した。 さらに, 96 名
の日本人ボランティア (32 名の幼児, 32 名の成人および 32 名の高齢者) の, 腸内における
C. coccoides group の構成を RT-qPCR により解析し, それぞれの年齢層における細菌叢の 特徴を明らかとした。
8 2. 材料および方法
使用菌株および培養法
本試験で使用した菌株をTable 2-1 に示した。 下記の 38 菌株について, 1%グルコース加
変法GAM ブロス (日水製薬) により, 嫌気条件下, 37℃で静置培養した。 培養時間につい
ては、下記の通りである。A. caccae, Dorea formicigenerans, E. ramulus, およびBlautia
producta : 18 時間; Clostridium hathewayi, Clostridium symbiosum, F. saccharivorans,
Ruminococcus gnavus, Ruminococcus lactaris, E. rectale, お よ び Eubacterium ventriosum : 20 時間; Clostridium nexile およびE. hallii : 22 時間; Clostridium indolis,
Clostridium oroticum, Eubacterium eligens, Clostridium celerecrescens, Clostridium sphenoides, Blautia hydrogenotrophica, Blautia schinkii, Blautia obeum, Blautia coccoides, Blautia hansenii, Blautia luti, C. hylemonae, C. scindens, Ruminococcus torques, Coprococcus eutactus, Coprococcus comes, R. intestinalis, Clostridium asparagiforme, Bacteroides vulgatus, Bifidobacterium longum, Collinsella aerofaciens,
Prevotella melaninogenica, Clostridium perfringens, およびClostridium difficile : 24 時 間; Faecalibacterium prausnitzii : 72 時間。 Lactobacillus acidophilusについては, MRS
ブロス (Becton Dickinson) により, 嫌気条件下, 37℃で 24 時間静置培養した。 E. coli,
Enterococcus faecalisおよびStaphylococcus aureusについては, Brain Heart Infusion
(BHI) ブロス (Becton Dickinson) により, 好気条件下, 37℃で 16 時間振盪培養した。 C.
jejuniについては, Preston 培地 (1.0% [wt/vol] Bacto peptone [Difco Laboratories], 1.0% [wt/vol] Lab lemco powder [Oxoid], 1.0% [wt/vol] PBS (-) [日水製薬], 0.025% [wt/vol] sodium pyruvate [関東化学], 0.025% [wt/vol] sodium disulfite [関東化学], 0.025% [wt/vol] iron(III) sulfate n-hydrate [関東化学]) により, 微好気条件下, 37℃で 16 時間振盪培養し た。 なお, 一連の嫌気培養操作は嫌気チャンバー (Coy Laboratory Products) 内で実施し た。 また, CampyPak plus micro-aerophilic システム (Becton Dickionson) を用いて, 微 好気条件を作出した。
被験者
8 名の健常成人 (ID: A-H, 年齢: 39±8 歳) から採取した糞便を用いて, RT-qPCR, qPCR
9
0.1 歳), 32 名の健常成人 (ID: A1-A32, 年齢: 39±11 歳) および 32 名の健常高齢者 (ID:
E1-E32, 年齢: 82±6 歳) から採取した糞便を, RT-qPCR によるC. coccoides group の解析
に用いた。 ヘルシンキ宣言に基づいて, すべての被験者は試験についての十分な説明を受 け, すべての被験者より試験への参加に関するインフォームドコンセントを得た。 本試験 はヤクルト中央研究所ヒト倫理審査委員会の承認を得た。
純培養菌液からのRNA 抽出
純培養菌液 100 l に RNAlater (Ambion) 200 l を加えて室温で 10 分間静置し, RNA を
固定した。 固定後の菌液を遠心分離 (13,000 g, 4℃, 10 分間) した。 遠心上清を除去した
後, Buffer RLT (Qiagen) 346.5 l, -メルカプトエタノール (Sigma-Aldrich) 3.5 l,
Tris-EDTA (TE) バッファー (Wako) 100 l, ガラスビーズ (直径 0.1 mm) (BioSpec
Products) 300 mg を加えて, ShakeMaster Auto (BioMedical Science) により 5 分間激しく
振盪して菌体を破砕した。 振盪後の菌液に, 水飽和フェノール (Wako) 500 l を加えた後, 60℃で 10 分間インキュベートした。 インキュベート後, クロロホルム-イソアミルアルコ ール (24:1) 100 l を加えて, 遠心分離 (13,000 g, 4ºC, 10 分間) した。 遠心上清 470 l を 回収し, 等量のクロロホルム-イソアミルアルコール (24:1) を加えて, 遠心分離 (13,000 g ,4ºC, 10 分間) した。 遠心上清 400 l を回収し, 等量のイソプロピルアルコールおよび 1/10 量の 3M 酢酸ナトリウム (pH 5.2) を加えて転倒混和した後, 遠心分離 (20,000 g, 4 ºC, 10 分間) した。 遠心上清を除去した後, 80% エタノール 500 l を加えて, 遠心分離 (20,000 g, 4ºC, 5 分間) した。 遠心上清を除去した後, 風乾させた。 得られた核酸ペレッ
ト (沈渣) に, Nuclease-free water (Ambion) 100 l を加えて溶解した。
純培養菌液からのDNA 抽出
純培養菌液 200 l に, Tris-SDS 溶液 (167 mM Tris-HCl, 66.7 mM EDTA [pH 9.0],
1.67% [wt/vol] ドデシル硫酸ナトリウム) 300 l, TE 飽和フェノール (Sigma-Aldrich) 500
l およびガラスビーズ(直径 0.1 mm) 300 mg を加えて, ShakeMaster Auto により, 15 分
間激しく振盪した。 振盪後の菌液を遠心分離 (20,000 g, 4ºC, 5 分間) した。 遠心上清 400
l を回収し, フェノール-クロロホルム-イソアミルアルコール (25:24:1) 400 l を加えて,
10 ル 300 l および 1/10 量の 3M 酢酸ナトリウム (pH 5.2) を加えて転倒混和した後, 遠心分 離 (20,000 g, 4ºC, 10 分間) した。 遠心上清を除去した後, 80% エタノール 500 l を加え て, 遠心分離 (20,000 g, 4ºC, 5 分間)した。 遠心上清を除去した後, 風乾させた。 得られ た核酸ペレットにTE バッファー 200 l を加えて溶解した。 DAPI 染色 純培養菌液 100 l に 4%パラホルムアルデヒド溶液 300 l を加えて, 4℃で 16 時間固定 した。 この固定液を PBS (-) により適宜希釈し, 希釈菌液 10 l を 8 枠付き MAS コート スライドグラス (松浪硝子工業) に一様に塗沫した。 風乾後, 各検体に VECTORSHIELD
with DAPI (Vector Laboratories) 4.8 l を反応させて封入した。 蛍光画像の取得には, ラ
イカイメージングシステム (Leica Microsystems) を用いた。 蛍光画像の解析には, Image-Pro Plusv. 4.5 (Media Cybernetics) を用いた。 1 検体につき 10 視野の菌数を計測 し, その平均値から純培養菌液の菌数濃度を算出した。
糞便処理
核酸抽出用に, 約 0.5 g の糞便を, RNAlater 2 ml の入った採便チューブに採取した。 ま
た, FISH 解析用に, 別の約 0.5 g の糞便を, 空の採便チューブに採取した。
核酸抽出用の糞便サンプルについては, 重量を測定した後, 10 倍希釈濃度となるように RNAlaterを加えて, 糞便 RNAlater懸濁液を調製した。 RNA 抽出用に, 糞便懸濁液 200
l を別のチューブに移し, PBS (-) 1 ml を加えて遠心分離 (20,000 g, 4ºC, 10 分間) した。
遠心上清を除去して, 得られたペレット (沈渣) を RNA 抽出に用いた。 RNA 抽出の方法 については, 下記に記載する事項以外を上述の「純培養菌液からの RNA 抽出」に記載した 方法に従った:菌体破砕には, FastPrep FP 120 (BIO101) を用いて, パワーレベル 5.0 で 60 秒間振盪した。 最終溶解に用いる Nuclease-free water 量を 1 ml とした。
また, DNA 抽出用に糞便 RNAlater懸濁液 200 l を別のチューブに移し, PBS (-) 1 ml
を加えて遠心分離 (20,000 g, 4ºC, 10 分間) した。 遠心上清 1 ml を除去した後, PBS (-) 1
ml を加えて遠心分離 (20,000 g, 4ºC, 10 分間) した。 遠心上清 1 ml を除去した後, さらに
PBS (-) 1 ml を加えて遠心分離 (20,000 g, 4ºC, 10 分間) した。 遠心上清 1 ml を除去し, 得
11 のDNA 抽出」に記載した方法に従い, 最終溶解に用いる TE バッファー量を 1 ml とした。 FISH 解析用の糞便サンプルの調製については, 検体の重量を測定した後, 10 倍希釈濃度 となるようにPBS (-) を加えた。 この糞便懸濁液 100 l を別のチューブに移し, 4%パラホ ルムアルデヒド溶液300 l を加えて, 4℃で 16 時間静置した。 この溶液を FISH 解析用検 体とした。 FISH 解析 上述の糞便処理で調製したFISH 解析用検体を, PBS (-) により適宜希釈し, 希釈菌液 10 l を 8 枠付き MAS コートスライドグラスに塗沫した。 風乾後, 蛍光標識したC. coccoides
group 特異的 DNA プローブ Erec482 (5’-GCTTCTTAGTCARGTACCG-3’) [53] 4.5 g /ml
を含むハイブリダイゼーション溶液 (750 mM NaCl, 100 mM Tris-HCl [pH 7.6], 5 mM
EDTA [pH 8.0], 0.01% BSA, 10% dextran sulfate) を, スライドグラスに 20 l 滴下して
カバーグラスを載せた。 スライドグラスを湿潤箱内で, 40℃, 16 時間, 暗所で静置した。 45℃の Wash 溶液 (50 mM NaCl, 4 mM Tris-HCl [pH 7.6], 0.02 mM EDTA [pH 8.0]) でス
ラ イ ド グ ラ ス を 洗 浄 し た 後, 滅 菌 水 を 用 い て 洗 浄 し た 。 風 乾 後 , 各 検 体 に
VECTORSHIELD with DAPI 4.8 l を添加し, 反応させて封入した。 画像の取得および解
析については, 上述の「DAPI 染色」と同様の方法で実施した。
プライマーの設計
ヒト腸管内から分離が報告されているC. coccoides group 菌種 [14,35,46], およびE. coli
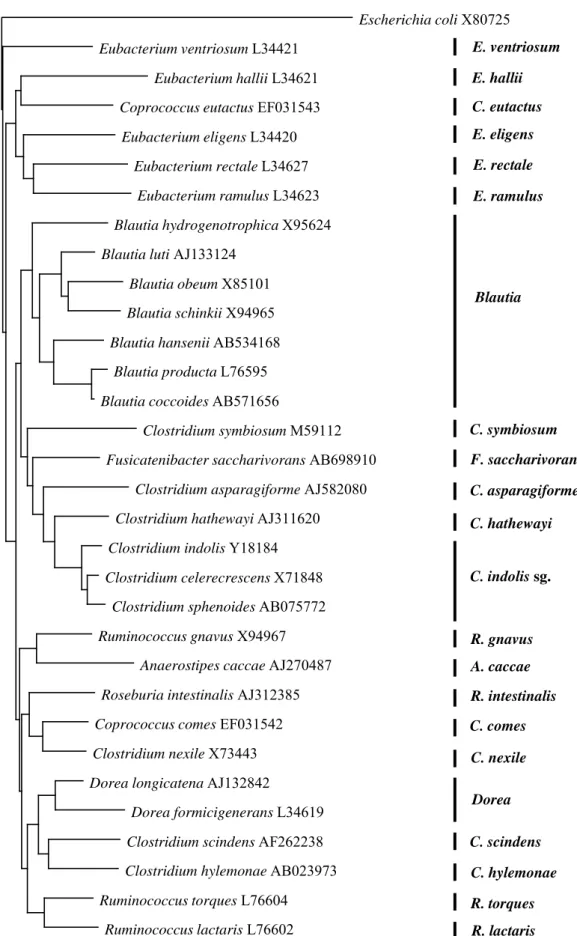
について, DDBJ/Genbank/EMBL データベースより 16S rRNA 配列を取得した。 Clustal X ソフトウェア [54] を用いて得られた配列の多重整列を行い, Tree View ソフトウェアを 用いて, 近隣接合法による系統樹を作成した。 作成した系統樹をもとに, ヒト腸管に棲息 するC. coccoides group を 3 菌群 (Blautia属, C. indolisサブグループ, Dorea属) および
19 菌種 (E. ventriosum, E. hallii, C. eutactus, E. eligens, E. rectale, E. ramulus, C.
symbiosum, F. saccharivorans, C. asparagiforme, C. hathewayi, R. gnavus, A. caccae, R. intestinalis, C. comes, C. nexile, C. scindens, C. hylemonae, R. torques, および R. lactaris) に系統化した (Figure 2-1)。 整列配列の比較から, それぞれのサブグループおよ び菌種に特異的な配列を同定し, それらをもとに特異的プライマーを設計した (Table 2-2)。
12
設 計 し た プ ラ イ マ ー 配 列 に つ い て は, Ribosomal Database Project (RDP-II)
(http://rdp.cme.msu.edu/) の Probe Match プログラムを用いて, データベース上の rRNA 遺伝子配列との交叉がないことを確認した。
RT-qPCR
RT-qPCR には, ABI PRISM 7900HT (Life Technologies), 384-Well Reaction Plate (Life
Technologies) および Qiagen OneStep RT-PCR kit (Qiagen) を用いた。 1 反応 (10 l) に
つ き, 1× Qiagen OneStep RT-PCR buffer, 0.5× Q-solution buffer, dNTP 400 M,
1/100,000 容 量 の SYBR green I (BioWhittaker Molecular Applications), Qiagen
OneStep RT-PCR enzyme mixture 0.4 l および RNA 溶液 5 l を用いた。 s-Acac-F/R 以
外のプライマーセットについては, 1 反応あたり 0.6 M となるように供試した。 s-Acac-F/R については, 0.6 M の濃度では増幅効率が低かったため, 2.1 M となるように 供試した。 逆転写反応として, 50℃, 30 分間反応させた後, 逆転写酵素を失活させるために, 95℃, 15 分間反応させた。 続いて, 94℃, 20 秒/ 55 ℃または 60℃, 20 秒/ 72℃, 50 秒を 1 反復として, 45 反復反応させた。 その反復ごとに, 増幅産物の量を SYBR Green I の蛍光 強度として測定した。 引き続いて, PCR の特異性を検証するために, 増幅産物の変性温度 の測定を行った。 94℃, 15 秒間反応させた後, 60℃から 95℃まで毎秒 0.2℃の速度で緩や かに温度を上昇させ, 増幅産物の乖離曲線を取得した。 また, 得られた増幅曲線について, 蛍光強度のベースラインおよび閾値を設定し, 増幅曲線と閾値が交差するサイクル数
(Quantification cycle: Cq値) を求めた。 データ解析には, Sequence Detection System ソ
フトウェア (Life Technologies) を用いた。 RT-qPCR による細菌数の測定 各標的細菌の標準曲線の作成には, Table 2-2 に記載した標準菌株を使用した。 それぞれ の標準菌株の純培養液を, 上述の方法により DAPI 染色し, 菌数濃度を測定した。 純培養 液から抽出したRNA の 10 倍段階希釈液を作製し, 供試 RNA 量と Cq値との比較により近 似曲線を作成した。 この近似曲線において, 直線性が認められる範囲のものを標準曲線と した。 糞便中の標的細菌数の測定については, 上述の方法で調製した RNA の 1/100, 1/1,000, 1/10,000 希釈液を作製し, それぞれ 5 l の溶液を RT-PCR 1 反応に供試した。 反
13
応後, 得られた Cq値を標的細菌の標準曲線に代入し, 鋳型 RNA あたりの菌数を算出して,
これをもとに, 糞便 1 g あたりの標的細菌数を求めた。 なお, 標準曲線については, 反応の たびに毎回作成した。
qPCR
qPCR には, ABI PRISM 7900HT および 384-Well Reaction Plate を用いた。 1 反応 (10
l) につき, 1× PCR buffer (タカラバイオ), dNTP 200 M, g-Ccoc-F/R プライマー (Table
2-2) 0.2 M, MgCl2 2.5 mM, 1/75,000 容量の SYBR Green I, Takara Taq (タカラバイオ)
0.02 units/l, TaqStart antibody (タカラバイオ) 5.5 ng/l および DNA 溶液 5 l を用いた。
95℃, 5 分間反応させた後、94℃, 20 秒/ 55℃, 20 秒/ 72℃, 50 秒を 1 反復として, 45 反復反 応させた。 その反復ごとに, 増幅産物の量を SYBR Green I の蛍光強度として測定した。 引き続いて, PCR の特異性を検証するために, 増幅産物の変性温度の測定を行った。 94℃, 15 秒間反応させた後, 60℃から 95℃まで毎秒 0.2℃の速度で緩やかに温度を上昇させ, 増幅 産物の乖離曲線を取得した。 また, 得られた増幅曲線について, 蛍光強度のベースライン および閾値を設定し, 増幅曲線と閾値が交差するサイクル数 (Quantification cycle: Cq値)
を求めた。 データ解析には, Sequence Detection System ソフトウェアを用いた。
qPCR による細菌数の測定 B. producta JCM 1471Tを標準曲線の作成に使用した。 上記菌株の純培養菌液を, 上述 の方法によりDAPI 染色し, 菌数濃度を測定した。 純培養菌液から抽出した DNA の 10 倍 段階希釈液を作製し, 供試 DNA 量と Cq値との比較により近似曲線を作成した。 この近似 曲線において, 直線性が認められる範囲のものを標準曲線とした。 糞便中の標的細菌数の 測定については, 上述の方法で調製した DNA の 1/10, 1/100, 1/1,000 希釈液を作製し, それ ぞれ5 l の溶液を PCR 1 反応に供試した。 反応後, 得られた Cq値を標的細菌の標準曲線 に代入し, 鋳型 DNA あたりの菌数を算出して, これをもとに糞便 1 g あたりの標的細菌数 を求めた。 なお, 標準曲線については, 反応のたびに毎回作成した。 プライマーの特異性確認 プライマーの特異性については, Table 2-1 に示す菌株を用いて, RT-qPCR により解析し
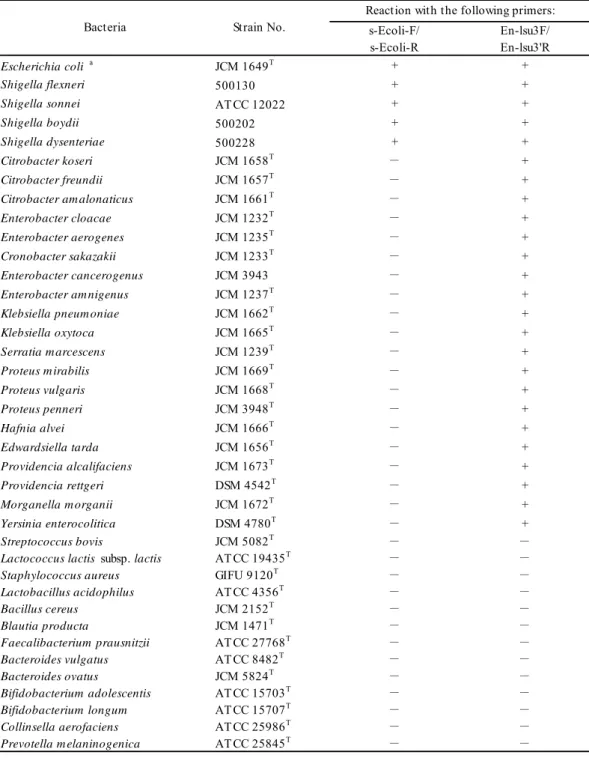
14 た。 標準 RNA については 1 反応あたり 1×105 個相当を供試した。 104 個相当以上の反 応性を示した菌株については陽性 (+), 100 個以上 104 個未満の反応性を示した菌株につい ては偽陽性 (±), 100 個未満の反応性を示した菌株については陰性 (-) として、特異性の判 定を行った。 増幅産物のシークエンシング解析
糞便RNA を鋳型として, s-Casp-1F/2R (Table 2-2) を用いて RT-qPCR を行った後の反
応液 10 µl を High Pure PCR Product Purification Kit(Roche Diagnostics)により精製 した。 それらを鋳型として, 先の RT-qPCR に使用したプライマーおよび Big Dye Terminator v3.1 Cycle Sequencing kit(Applied Biosystems)を用いて PCR を行った。 増
幅された DNA をエタノール沈殿し, ホルムアミド 15 µl に溶解後, ABI PRISM 3130
(Applied Biosystems)に供した。 シークエンシング解析により得られた塩基配列の相同 性検索を, Ribosomal Database Project(http://rdp.cme.msu.edu/)の Sequence Match を 用いて実施した。 統計解析 細菌数の統計解析には対数値を用い,検出限界未満の検体については, 検出限界値の半値 を代入した。細菌数の検定には, JMP v. 9.0 ソフトウェア (SAS Institute) を用い, Steel-Dwass による多重比較を行った。 また, 検出頻度の検定には Fisher の正確確率検定 後, Holm 補正を実施した。 多変量解析については, 統計ソフトウェア R 2. 1. 5 を用いて主 成分分析を実施した。 ヒートマップの作成には, R の “heatmap” パッケージを用いた。 ク ラスタリングには, ユークリッド距離に基づく Ward 法を用いた。 3. 結果
RT-qPCR, FISH および qPCR 法により測定した, ヒト腸内におけるC. coccoides group 菌
数の比較
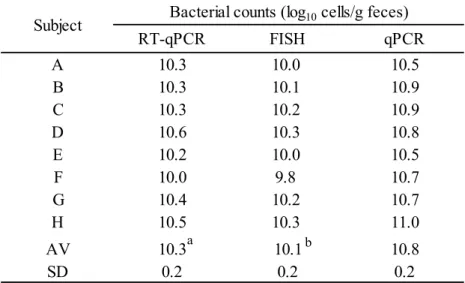
8 名の健常成人における糞便中の C. coccoides group 菌数を, RT-qPCR, FISH および
qPCR 法により解析した (Table 2-3)。その結果, RT-qPCR 法による測定菌数は, FISH 法に よる測定菌数と統計学的に同等であった。これに対して、RT-qPCR 法による測定菌数は,
15
qPCR 法による測定菌数と比較して, 有意に低かった (P <0.01)。
プライマーの特異性検討
Table 2-1 に示した 43 菌株から RNA を抽出し, 細菌 105個相当の RNA を鋳型として
RT-qPCR を行い, プライマーの特異性を検討した。 その結果, それぞれのプライマーは, 標的細菌種に特異的に反応した(Table 2-1)。 g-Blau-F/R, s-Csym-F/R, s-Casp-F/R およ
びs-Rgna-F/R は, 標的以外の細菌種と弱く交叉反応を示したが, これは標的細菌種と比較
して1/10,000 程度であるため, 標的細菌種の定量に差し支えないと考えられた。
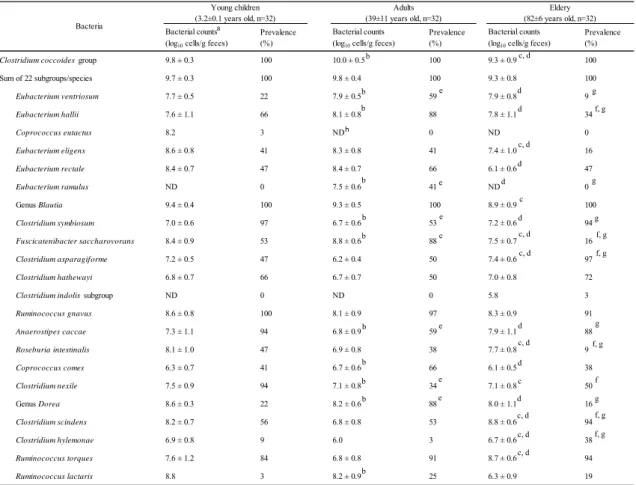
幼児, 成人および高齢者の腸内におけるC. coccoides group 構成の比較
幼児, 成人および高齢者, それぞれ 32 名の糞便中のC. coccoides group 構成を RT-qPCR
により解析した (Table 2-4)。 幼児および成人の糞便におけるC. coccoides group 菌数は,
糞便1 グラムあたり, それぞれ 109.8±0.3および1010.0±0.5個であった。 また, 高齢者の糞便 におけるC. coccoides group 菌数は, 糞便 1 g あたり, 109.3±0.9個であり, 幼児および成人に おける菌数と比較して有意に低かった (P <0.01)。 幼児, 成人および高齢者における, 22 プ ライマーによる測定菌数の総和は, 糞便 1 g あたり, それぞれ 109.7±0.3個 (C. coccoides group プライマーによる測定菌数の 99.4%), 109.8±0.4個 (同 99.8%) および 109.3±0.8個 (同 100%) であった。 Blautia属菌は, いずれの年齢層においても, 22 菌群/菌種の中で最も菌 数が高かったことから, 本属は年齢に関わらずヒト腸内の最優勢菌であることが示唆され た。 R. gnavusおよびR. torquesは, いずれの年齢層においても検出頻度が 84%~100% と高かった。 また, 成人におけるF. saccharivoransおよびDorea属菌の検出頻度 (それぞ れ88%および 88%) は, 幼児 (それぞれ 53%および 22%) ならびに高齢者の検出頻度 (それ ぞれ16%および 16%) と比較して, 有意に高かった (P <0.01)。 E. ramulusは成人の41% から検出されたが, 他の年齢層からは検出されなかった。 一方, 幼児におけるC. nexileの 検出率は 94%と高く, これは成人 (34%) および高齢者における検出頻度 (47%) と比較し て、有意に高かった (P <0.01)。 高齢者におけるC. scindensおよびC. hylemonaeの検出 率 (94%および 38%) は, 幼児 (56%および 9%) ならびに成人における検出頻度 (53%およ び 3%) と比較して, 有意に高かった (P <0.01)。 その他, E. ventriosum, E. hallii, E.
16
C. comes, R. torquesおよびR. lactarisの菌数または検出頻度についても, 年齢層による違
いが認められた (P <0.05)。
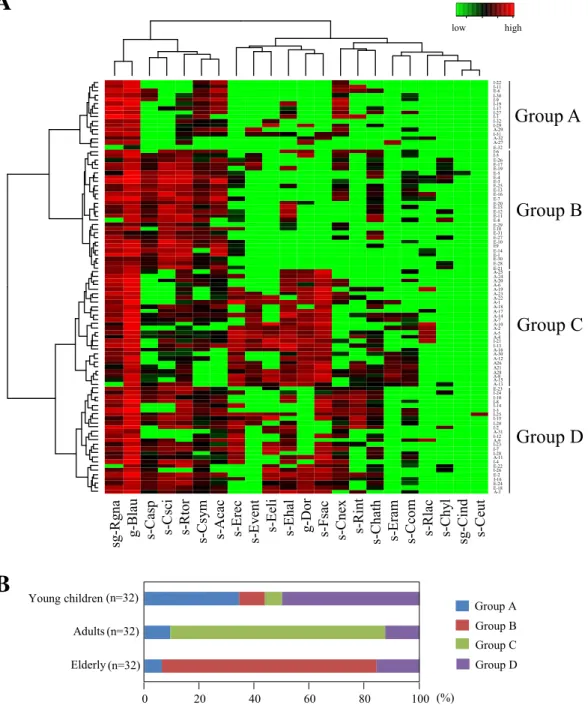
菌数データに基づいて, 96 名の被験者のクラスタリング解析を行った結果, これらは 4 グ ループ (Group A – D) に分類された (Figure 2-2)。 成人の 78%は Group C に分類された のに対して, 高齢者の 78%は Group B に分類された。 また, 幼児の 34%および 50%がそ れぞれGroup A および GroupD に分類された。 さらに, 主成分分析を行った結果, 年齢層 によるフローラの違い, 特に成人と高齢者の間の違いが顕著に認められ, クラスタリング 解析を支持する結果となった (Figure 2-3)。 4. 考察 これまで, 培養法および分子生物学的手法により, 多くのヒト腸内細菌叢解析が実施さ れてきた。 しかし, 適切な選択培地および菌種特異的プライマーが存在しなかったため, C. coccoides group のみに着目した解析はほとんど報告されていなかった。 そこで, 本研究で は本グループを便宜的に3 サブグループおよび 19 菌種に系統化し, それぞれを標的とした 特異的プライマーを構築した。
RT-qPCR により測定した, ヒト腸内におけるC. coccoides group 菌数は, FISH 法による
測定菌数と統計学的に同等であったが, qPCR 法による測定菌数と比較して有意に低かった (Table 2-3) 。 これは, それぞれの手法における標的分子の違いによるものと考えられた。 すなわち, RT-qPCR および FISH 法の標的分子は RNA であるのに対して, qPCR 法の標的 分子はDNA であることに起因すると考えられた。 死菌において RNA 分子は速やかに分 解されるのに対して, DNA は比較的安定して存在することから, RNA は生菌の指標となり 得る [55,56]。 嫌気性および好気性細菌を用いたin vitroの試験において, RT-qPCR によ り測定された菌数と, 培養法により測定された菌数の間に高い相関が認められている [42]。 DNA は死菌中でも比較的安定に存在することから, qPCR 法による測定菌数には, 生菌の みでなく死菌も含まれる可能性がある。 したがって, 本研究における RT-qPCR および qPCR 法により測定された菌数の差は, 死菌の数を反映していると考えられた。 また, RT-qPCR および FISH 法は, いずれも rRNA を標的としているにも関わらず, 両者により
測定された菌数にもわずかな差 (100.1 – 100.3 cells/ g feces) が認められた(Table 2-3)。 こ
17 度を有する細菌を陽性, これに満たない細菌を陰性と判定して細菌数を測定するのに対し て, RT-qPCR 法では, 総 rRNA 量を細菌数に換算することから, 両者による測定菌数に若干 の差が生じたと考えられた。生菌および死菌の判定には, RNA 量の他, 細胞膜透過性 [57], および呼吸活性 [58] といった指標が用いられるが, 生菌および死菌の閾値の設定には主 観的判断が伴う。 RT-qPCR 法は FISH 法と異なり, 上記の閾値設定による問題がなく, 客 観的な評価が可能だと考えられる。 ヒトの生後まもなく, 腸管には通性嫌気性細菌が速やかに定着する [19]。 これらの細菌 による酸素消費に伴って, 偏性嫌気性細菌が通性嫌気性細菌に代わって優勢に増殖すると 言われている [59]。 C. coccoides group は生後の比較的早い段階から腸内に定着し, それ らは永続的に存在すると考えられている [19,60]。 本研究においても, 幼児および成人にお けるC. coccoides group 菌数は同レベルであったことから (Table 2-4), 本試験は上記の説 を支持する結果となった。 しかし、本グループの構成については, 幼児および成人で異な ることが明らかとなった。 例えば, 幼児では全く検出されなかったE. ramulusが, 成人の 41%から検出された。 E. ramulusは, フラボノイドの 1 種であるケルセチン-3-グリコシド (イソケルセチン) をエネルギー源として利用することが知られている [61]。 フラボノイド は植物由来の食品および飲料に含まれるポリフェノール化合物であり [62], in vivo試験に おいて, フラボノイド摂取がE. ramulusの増殖を促進することが報告されていることから [63], 成人の食物にこの物質が多く含まれている可能性が考えられた。 一方, 抗菌物質産生 菌と言われているC. nexile [64] は, 成人 (34%) と比較して, 幼児 (94%) において高頻度 で検出された。 このことから, C. nexileは幼児の腸内細菌叢を特徴づけている可能性が考 えられた。 幼児が通常摂取することがないと考えられるニンニクは, C. nexileの増殖を抑 制することが報告されている [65]。 以上の結果から, 各年齢層における摂取食物の差異が 腸内のC. coccoides group 分布を規定する重要な因子である可能性が示唆された。 腸内のC. coccoides group は, 抗生物質や抗炎症薬投与等の影響を受けやすく, 高齢者に おいて, その細菌数が減少することが報告されている [66-69]。 本研究においても, 日本, イタリアおよびフィンランドにおける試験結果 [28,70,71] と同様, 乳児および成人と比較
して, 高齢者におけるC. coccoides group の菌数は有意に低かった (Table 2-4)。 いずれの
年齢層においても, C. coccoides group の中ではBlautia属菌が最優勢で検出されたことか
18 と考えられた。 Blautia 属は近年, 分類体系の再編により提唱された新属で [72], これま での多くの研究においても, 本属菌種がヒト腸内から高頻度に検出されている [14,35,73]。 加齢に伴う Blautia 属菌の減少は過去に報告されており [35], 本結果はこれと合致する。 Blautia属菌は1 型糖尿病や肝硬変との関連が示唆されていることから [74,75], 本属菌の 定量解析は宿主の健康状態を知る上でも重要であると考えられる。 クラスタリング解析および主成分分析の結果から, 高齢者における腸内の C. coccoides group 細菌叢パターンは, 成人のそれと大きく異なることが示唆された (Figures 2-2, 2-3)。
高齢者ではC. scindensおよびC. asparagiformeが高頻度で検出され, 逆にDorea属菌お
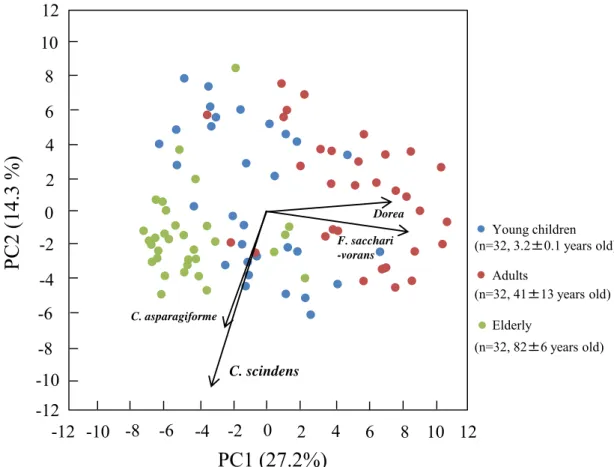
よびF. saccharivoransの検出頻度が低いという特徴が観察された。 このことから, これら の菌種が高齢者菌叢のマーカーになり得る可能性が示唆された。 F. saccharivorans は近 年分離された菌種であるが [47], Dorea 属菌とともに生体内における機能はほとんど明ら かとなっておらず, 宿主の健康との関わりについて興味がもたれる。 一方, C. scindensは、 胆汁酸7-脱水酸化能を有しており, デオキシコール酸やリトコール酸等の二次胆汁酸の産 生に関与することが報告されている [51]。 乳児および成人と比較して, 高齢者において検 出頻度が高かったC. hylemonaeについても, C. scindensと同様の機能を有することが報告 されている [76]。 二次胆汁酸は, 大腸ガンおよび肝細胞ガン発症に関与していることが知 られていることから [77,78], 高齢者におけるガン発症リスクの増大に, これらの二次胆汁 酸産生菌種の増加および二次胆汁酸濃度増加が関連している可能性を示唆するものとして 興味深い。 また, 主成分分析における幼児のプロットは, 成人および高齢者と比較して, PC1 および PC2 軸沿いに広く分散していたことから (Figure 2-3), 幼児では個人間の多様 性に富んでいることが示唆された。 さらに, クラスタリング解析においても, 多くの幼児 は成人および高齢者と異なるクラスターに配置されたことから, その細菌構成は成人およ び高齢者と大きく異なっていることが明らかとなった。 このことから, C. coccoides group 細菌叢は幼児期において個人間の多様性に富んでおり, 成人および高齢者となるにつれて, その細菌叢も年齢層に応じた安定した構成に移行することが示唆された。 本研究の被験者 数および年齢層は限られており, さらに多くの年齢層の被験者を用いた大規模な細菌叢解 析により, 一生涯の細菌叢のより詳細な変遷が明らかになると考えられる。
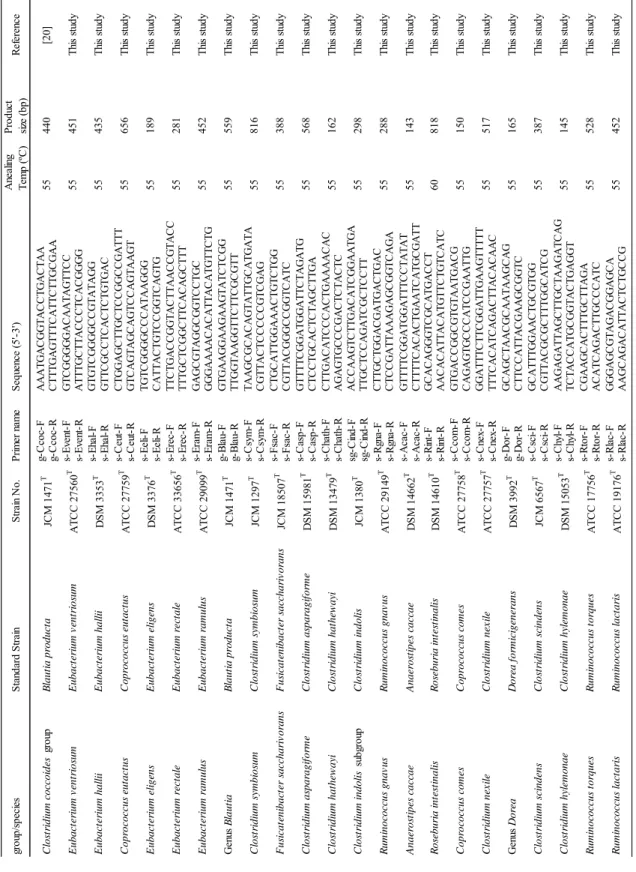
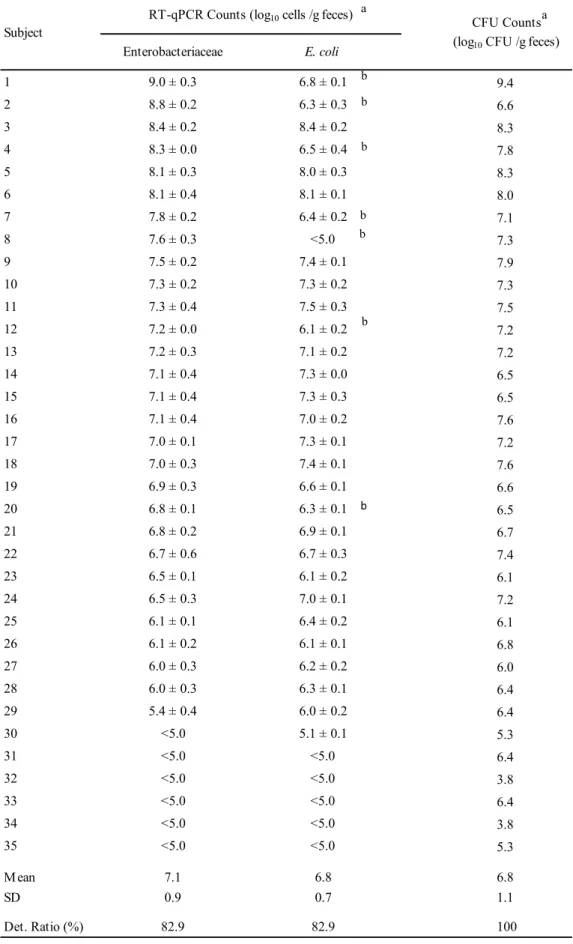
19 T abl e 2 -1 Pr im er spe ci fici ty s-E ve nt s-E ha l s-Ce ut s-E el i s-E re c s-E ra m g-Bl au s-Cs ym s-F sa c s-Ca sp s-Cha th sg -Ci nd s-Rg na s-A ca c s-Ri nt s-Cc om s-Cne x g-D or s-Cs ci s-Chy l s-Rt or s-Rl ac -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R -F /R Cl os tr idi um c oc coi de s g roup E ubac te ri um v ent ri os um A T CC 27560 T + -± ± -E ubac te ri um hal li i D SM 3353 T -+ -Copr oc oc cus e ut ac tus A T CC 27759 T -+ -E ubac te ri um e li ge ns D SM 3376 T -+ -E ubac te ri um r ec tal e A T CC 33656 T -+ -± -E ubac te ri um r am ul us A T CC 29099 T -+ -± -B laut ia hy dr oge not rophi ca D SM 10507 T -+ -B laut ia l ut i D SM 14534 T -+ -R um inoc oc cus obe um A T CC 29174 T -+ -B laut ia s chi nk ii D SM 10518 T -+ -B laut ia hans eni i A T CC 27752 T -+ -B laut ia pr oduc ta JCM 1471 T -+ -B laut ia c oc coi de s JCM 1395 T -+ -Cl os tr idi um s ym bi os um JCM 1297 T -± + -F us ic at eni bac te r s ac char iv or ans JCM 18507 T -+ -Cl os tr idi um as par agi for m e D SM 15981 T -± -+ -Cl os tr idi um hat he way i D SM 13479 T -+ -Cl os tr idi um i ndol is JCM 1380 T -± -+ -Cl os tr idi um c el er ec re sc ens D SM 5628 T -± -+ -Cl os tr idi um s phe noi de s JCM 1415 T -± -+ -R um inoc oc cus gnav us A T CC 29149 T -± -+ -A nae ros ti pe s c ac cae D SM 14662 T -± -+ -R os ebur ia i nt es ti nal is D SM 14610 T -+ -Copr oc oc cus c om es A T CC 27758 T -+ -Cl os tr idi um ne xi le A T CC 27757 T -± -+ -D or ea f or m ic ige ne rans D SM 3992 T -+ -Cl os tr idi um s ci nde ns JCM 6567 T -± -+ -Cl os tr idi um hy le m onae D SM 15053 T -+ -R um inoc oc cus t or que s A T CC 17756 T -+ -R um inoc oc cus l ac tar is A T CC 19176 T -+ Cl os tr idi um or ot ic um JCM 1429 T -± -O the r g roup s F ae cal ibac te ri um pr aus ni tz ii A T CC 27768 T -B ac te roi de s v ul gat us JCM 5824 T -B if idobac te ri um l ongum A T CC 15707 T -Col li ns el la ae rof ac ie ns A T CC 25986 T -P re v ot el la m el ani noge ni ca A T CC 25845 T -Cl os tr idi um pe rf ri nge ns JCM 1290 T -L ac tobac il lus ac idophi lus A T CC 4356 T -E sc he ri chi a c ol i A T CC 11775 T -E nt er oc oc cus f ae cal is A T CC 19433 T -St aphy loc oc cus aur eus A T CC 12600 T -Cl os tr idi um di ff ic il e D SM 1296 T -Cam py lobac te r j ej uni A T CC 33560 T -St ra in Re ac tio n w ith th e fo llo w in g pri m ers a a The sp ecifi cit y of the RT -q PC R as sa y for t he tar get bact er ia w ith each pr im er w as i nv es tigat ed usi ng R N A ext ract s cor res pondi ng to 10 5 ce lls of each st rai n. Spe cifi cit y w as judged usi ng t he cr iter ia des cr ibed i n t he t ext .
20 g-C coc -F A A A TG A C G G TA C C TG A C TA A g-C coc -R C TT TG A G TT TC A TT C TT G C G A A s-Ev en t-F G TC G G G G G A C A A TA G TT C C s-Ev en t-R A TT TG C TT A C C C TC A C G G G G s-Eh al-F G TG TC G G G G C C G TA TA G G s-Eh al-R G TT C G C C TC A C TC TG TG A C s-C eu t-F C TG G A G C TT G C TC C G G C C G A TT T s-C eu t-R G TC A G TA G C A G TC C A G TA A G T s-Ee li-F TG TC G G G G C C C A TA A G G G s-Ee li-R C A TT A C TG TC C G G TC A G TG s-Er ec -F TT C TG A C C G G TA C TT A A C C G TA C C s-Er ec -R TT TG C TC G G C TT C A C A G C TT T s-Er am -F G A G C G TA G G C G G TC C TG C s-Er am -R GGGAAAAC AC AT TAC AT GT TC TG g-Bl au -F G TG A A G G A A G A A G TA TC TC G G g-Bl au -R TT G G TA A G G TT C TT C G C G TT s-C sy m -F TA A G C G C A C A G TA TT G C A TG A TA s-C sy m -R C G TT A C TC C C C C G TC G A G s-Fs ac -F C TG C A TT G G A A A C TG TC TG G s-Fs ac -R C G TT A C G G G C C G G TC A TC s-C as p-F G TT TT C G G A TG G A TT C TA G A TG s-C as p-R C TC C TG C A C TC TA G C TT G A s-C ha th -F C TT G A C A TC C C A C TG A A A A C A C s-C ha th -R A G A G TG C C C G A C TC TA C TC sg -C ind-F A C C A A G TC TT G A C A TC G G A A TG A sg -C ind-R TT G C TC C A G A TC G C TC C TT s-Rg na -F C TT G C TG G A C G A TG A C TG A C s-Rg na -R C TC C G A TT A A A G A G C G G TC A G A s-A ca c-F G TT TT C G G A TG G A TT TC C TA TA T s-A ca c-R C TT TT C A C A C TG A A TC A TG C G A TT s-Ri nt -F G C A C A G G G TC G C A TG A C C T s-Ri nt -R A A C A C A TT A C A TG TT C TG TC A TC s-C com -F G TG A C C G G C G TG TA A TG A C G s-C com -R C A G A G TG C C C A TC C G A A TT G s-C ne x-F G G A TT TC TT C G G A TT G A A G TT TT T s-C ne x-R TT TC A C A TC A G A C TT A C A C A A C g-D or -F G C A G C TA A C G C A A TA A G C A G g-D or -R C TT C C A TT A C G A A G C G G TC s-C sc i-F G C A TT TG G A A C TG C G TG G s-C sc i-R C G TT A C G C G C TT TG G C A TC G s-C hy l-F A A G A G A TT A G C TT G C TA A G A TC A G s-C hy l-R TC TA C C A TG C G G TA C TG A G G T s-Rt or -F C G A A G C A C TT TG C TT A G A s-Rt or -R A C A TC A G A C TT G C C C A TC s-Rl ac -F G G G A G C G TA G A C G G A G C A s-Rl ac -R A A G C A G A C A TT A C TC TG C C G R um inoc oc cus l ac tar is R um inoc oc cus l ac tar is A TC C 191 76 T 55 452 Th is s tu dy R um inoc oc cus t or que s R um inoc oc cus t or que s A TC C 177 56 T 55 528 Th is s tu dy C los tr idi um hy le m ona e C los tr idi um hy le m ona e D SM 150 53 T 55 145 Th is s tu dy C los tr idi um s ci nde ns C los tr idi um s ci nde ns JC M 656 7 T 55 387 Th is s tu dy G en us D or ea D or ea f or m ic ige ne rans D SM 399 2 T 55 165 Th is s tu dy C los tr idi um ne x il e C los tr idi um ne x il e A TC C 277 57 T 55 517 Th is s tu dy C opr oc oc cus c om es C opr oc oc cus c om es A TC C 277 58 T 55 150 Th is s tu dy R os ebur ia i nt es ti nal is R os ebur ia i nt es ti nal is D SM 146 10 T 60 818 Th is s tu dy A nae ros ti pe s c ac cae A nae ros ti pe s c ac cae D SM 146 62 T 55 143 Th is s tu dy R um inoc oc cus gna v us R um inoc oc cus gna v us A TC C 291 49 T 55 288 Th is s tu dy C los tr idi um i ndo li s su bg rou p C los tr idi um i ndo li s JC M 138 0 T 55 298 Th is s tu dy C los tr idi um hat he w ay i C los tr idi um hat he w ay i D SM 134 79 T 55 162 Th is s tu dy C los tr idi um as par agi for m e C los tr idi um as par agi for m e D SM 159 81 T 55 568 Th is s tu dy F us ic at eni bac te r s ac char iv or ans F us ic at eni bac te r s ac char iv or ans JC M 185 07 T 55 388 Th is s tu dy C los tr idi um s ym bi os um C los tr idi um s ym bi os um JC M 129 7 T 55 816 Th is s tu dy G en us B laut ia B laut ia pr odu ct a JC M 147 1 T 55 559 Th is s tu dy E uba ct er ium r am ul us E uba ct er ium r am ul us A TC C 290 99 T 55 452 Th is s tu dy E uba ct er ium r ec tal e E uba ct er ium r ec tal e A TC C 336 56 T 55 281 Th is s tu dy E uba ct er ium e li ge ns E uba ct er ium e li ge ns D SM 337 6 T 55 189 Th is s tu dy C opr oc oc cus e ut ac tus C opr oc oc cus e ut ac tus A TC C 277 59 T 55 656 Th is s tu dy E uba ct er ium hal li i E uba ct er ium hal li i D SM 335 3 T 55 435 Th is s tu dy E uba ct er ium v ent ri os um E uba ct er ium v ent ri os um A TC C 275 60 T 55 451 Th is s tu dy Pr odu ct siz e ( bp) Re fe re nc e C los tr idi um c oc coi de s g rou p B laut ia pr odu ct a JC M 147 1 T 55 440 [20] gr ou p/ spe cie s St an da rd S tra in St ra in N o. Pr im er n am e Se qu en ce (5' -3' ) A ne ali ng Te m p ( ºC ) T abl e 2 -2 Pr im er inf or m at ion
21
Table 2-3 Comparison of C. coccoides group population levels in
human feces determined by RT-qPCR, FISH and qPCR
a Indicates a significant difference between RT-qPCR and qPCR with
Steel-Dwass test (P <0.01) RT-qPCR FISH qPCR A 10.3 10.0 10.5 B 10.3 10.1 10.9 C 10.3 10.2 10.9 D 10.6 10.3 10.8 E 10.2 10.0 10.5 F 10.0 9.8 10.7 G 10.4 10.2 10.7 H 10.5 10.3 11.0 AV 10.3 10.1 10.8 SD 0.2 0.2 0.2
Subject Bacterial counts (log10 cells/g feces)
b a
b Indicates a significant difference between FISH and qPCR with
22 Bacterial counts
(log10 cells/g feces) Prevalence(%)
Bacterial counts
(log10 cells/g feces) Prevalence(%)
Bacterial counts
(log10 cells/g feces) Prevalence(%)
Clostridium coccoides group 9.8 ± 0.3 100 10.0 ± 0.5 100 9.3 ± 0.9 100
Sum of 22 subgroups/species 9.7 ± 0.3 100 9.8 ± 0.4 100 9.3 ± 0.8 100 Eubacterium ventriosum 7.7 ± 0.5 22 7.9 ± 0.5 59 7.9 ± 0.8 9 Eubacterium hallii 7.6 ± 1.1 66 8.1 ± 0.8 88 7.8 ± 1.1 34 Coprococcus eutactus 8.2 3 ND 0 ND 0 Eubacterium eligens 8.6 ± 0.8 41 8.3 ± 0.8 41 7.4 ± 1.0 16 Eubacterium rectale 8.4 ± 0.7 47 8.4 ± 0.7 66 6.1 ± 0.6 47 Eubacterium ramulus ND 0 7.5 ± 0.6 41 ND 0 Genus Blautia 9.4 ± 0.4 100 9.3 ± 0.5 100 8.9 ± 0.9 100 Clostridium symbiosum 7.0 ± 0.6 97 6.7 ± 0.6 53 7.2 ± 0.6 94 Fuscicatenibacter saccharovorans 8.4 ± 0.9 53 8.8 ± 0.6 88 7.5 ± 0.7 16 Clostridium asparagiforme 7.2 ± 0.5 47 6.2 ± 0.4 50 7.4 ± 0.6 97 Clostridium hathewayi 6.8 ± 0.7 66 6.7 ± 0.7 50 7.0 ± 0.8 72
Clostridium indolis subgroup ND 0 ND 0 5.8 3
Ruminococcus gnavus 8.6 ± 0.8 100 8.1 ± 0.9 97 8.3 ± 0.9 91 Anaerostipes caccae 7.3 ± 1.1 94 6.8 ± 0.9 59 7.9 ± 1.1 88 Roseburia intestinalis 8.1 ± 1.0 47 6.9 ± 0.8 38 7.7 ± 0.8 9 Coprococcus comes 6.3 ± 0.7 41 6.7 ± 0.6 66 6.1 ± 0.5 38 Clostridium nexile 7.5 ± 0.9 94 7.1 ± 0.8 34 7.1 ± 0.8 50 Genus Dorea 8.6 ± 0.3 22 8.2 ± 0.6 88 8.0 ± 1.1 16 Clostridium scindens 8.2 ± 0.7 56 6.8 ± 0.8 53 8.8 ± 0.6 94 Clostridium hylemonae 6.9 ± 0.8 9 6.0 3 6.7 ± 0.6 38 Ruminococcus torques 7.6 ± 1.2 84 6.8 ± 0.8 91 8.7 ± 0.6 94 Ruminococcus lactaris 8.8 3 8.2 ± 0.9 25 6.3 ± 0.9 19 Bacteria Young children
(3.2±0.1 years old, n=32) (39±11 years old, n=32)Adults (82±6 years old, n=32)Eldery
d f, g c, d c a h e e e e e e e g f, g g g g f, g f, g g f, g f f, g b b b b b b b b b b d d d d c, d c, d d c, d d c d c, d c, d c, d b c, d
Table 2-4 Comparison of C. coccoides group populations among different age groups
a Data are expressed as the means and standard deviations.
b Indicates a significant difference between infants and adults with Steel-Dwass test (P <0.05)
c Indicates a significant difference between infants and elderly with Steel-Dwass test (P <0.05)
d Indicates a significant difference between adults and elderly with Steel-Dwass test (P <0.05)
e Indicates a significant difference between infants and adults with Fisher's exact probability test
after Holm correction (P <0.05)
f Indicates a significant difference between infants and elderly with Fisher's exact probability test
after Holm correction (P <0.05)
g Indicates a significant difference between adults and elderly with Fisher's exact probability test
after Holm correction (P <0.05)
23 E. ventriosum E. hallii E. eligens E. rectale E. ramulus Blautia C. symbiosum F. saccharivorans C. hathewayi C. indolis sg. Dorea R. lactaris C. nexile R. gnavus A. caccae C. comes R. intestinalis C. asparagiforme C. eutactus R. torques C. hylemonae C. scindens 0.1 Escherichia coli X80725 Eubacterium ventriosum L34421 Eubacterium hallii L34621 Coprococcus eutactus EF031543
Eubacterium eligens L34420 Eubacterium rectale L34627
Eubacterium ramulus L34623 Blautia hydrogenotrophica X95624 Blautia luti AJ133124
Blautia obeum X85101 Blautia schinkii X94965 Blautia hansenii AB534168
Blautia producta L76595 Blautia coccoides AB571656
Clostridium symbiosum M59112 Fusicatenibacter saccharivorans AB698910
Clostridium asparagiforme AJ582080 Clostridium hathewayi AJ311620 Clostridium indolis Y18184 Clostridium celerecrescens X71848
Clostridium sphenoides AB075772 Ruminococcus gnavus X94967
Anaerostipes caccae AJ270487 Roseburia intestinalis AJ312385 Coprococcus comes EF031542 Clostridium nexile X73443 Dorea longicatena AJ132842
Dorea formicigenerans L34619 Clostridium scindens AF262238 Clostridium hylemonae AB023973 Ruminococcus torques L76604
Ruminococcus lactaris L76602
Figure 2-1 Phylogenetic tree showing the relationships among 16S rRNA gene sequences of
the Clostridium coccoides group. Scale bar represents 0.1 substitutions per nucleotide position. The Escherichia coli sequence was used as an outgroup for rooting the tree.
24 Young children Adults Elderly Group A Group B Group C Group D
B
0 (n=32) (n=32) (n=32) 20 40 60 80 100 (%)A
sg -R gna g-B lau s-C asp s-C sc i s-R tor s-C sy m s-Ac ac s-Er ec s-Eve nt s-Ee li s-Eha l g-Dor s-Fsa c s-C ne x s-R int s-Cha th s-Er am s-C com s-R lac s-C hy l sg -C ind s-C eut A-3 E-18 E-24 I-16 E-2 I-26 E-22 I-4 A-11 I-28 I-7 I-23 A-9 I-12 A-31 I-2 I-20 I-15 I-25 I-3 I-14 I-8 I-10 I-24 E-23 A-13 A-15 A-8 A28 A21 A26 A-12 A-30 A-16 I-13 I-21 A-4 A-5 A-2 A-10 A-7 A-14 A-17 A-18 A-1 A-22 A-23 A-19 A-6 A-20 A-24 A-25 E-21 E-28 E-30 E-1 E-14 E9 E-10 E-27 E-31 I-18 E-29 E-8 E-11 E-12 E-15 E-20 E-7 E-16 E-13 E-25 E-3 E-4 E-5 E-19 E-17 E-26 I-5 I-6 E-32 A-27 A-32 I-31 A-29 I-29 I-32 I-1 I-27 I-17 I-19 I-9 I-30 E-6 I-11 I-22Group A
Group C
Group D
high lowGroup B
Figure 2-2 Heatmap analysis of the counts of C. coccoides group populations enumerated
by using 22 primer sets in 32 young children, 32 adults, and 32 elderly. (A) Hierarchical clustering with a heatmap representation based on Ward's minimum variance method and a Euclidean distance metric. Subject IDs of 32 young children (I-1 to I-32), 32 adults (A-1 to A-32), and 32 elderly (E-1 to E-32) are shown at the right side of the heatmap. The subjects tested were classified into 4 groups (Groups A to D) by hierarchical clustering. Colors ranging from green to red indicate low to high population levels. (B) Classification of subjects by the hierarchical clustering. Most of the adults (78%) were classified into Group C and most of the elderly (78%) into Group B, Specific clusters were not as apparent in children.
25
2
0
2
4
6
8 10
-2
-4
-6
-8
-8
-6
-4
-2
0
4
6
8
PC1 (27.2%)
PC2
(1
4.3
%)
Young children Elderly Adults (n=32, 3.2±0.1 years old) (n=32, 41±13 years old) (n=32, 82±6 years old)10
-10
-10
F. sacchari -vorans Dorea C. scindens-12
12
12
-12
C. asparagiformeFigure 2-3 Principal component analysis (PCA) of bacterial counts determined by using
22 primers in 32 young children, 32 adults, and 32 elderly. Blue, red, and green plots show young children, adults and the elderly, respectively. Arrows indicate characteristic vectors of the upper 4 factor loadings.