Tech Bull Fac. Agr Kagawa Univ Vol 28, No 60, 67-74, 1977
DECOMPOSITION O F SOYBEAN OLIGOSACCHARIDES BY INTESTINAL BACTERIA
VIII. p-Fructosidase Activity of Some Intestinal Bacteria*
Sin'itir 6 KAWAMURA,
* *
Tadasi KASAI** *
and SumizB TANUSI****~ ~ P J M I O ,5 6 Ercherichia coli 9 &, Lactobacillus acidophilus 2 #, b d; U; Streptococcus faecalis 1 # l E 9 & &
d8@ ( Y m = t - x , $ 4 ~ 7 1. -.x, . 7 , ~ . / / ~ - . - . x , , y . . . y k a , - x , 9 9 r & - , x , 7 7 4 / - x ) &3f.~6$ithiEb tf&@H$q. (L%S@)
a
5U;lC&NJYdrn (gLg@g!L?gkCd; & ) 4 241L.fi,-*
(,V y 7 .-.e') D&,hJ*;m4,7:. $,& &'"iCt@EObIb$j&j5<, 4 2 4 1 k f i -,+'D&&&#,&f=, 7 4 1 k f i --.t*ht a-Y/k=t V k ?.-+'( 2
f i d d / i " ~ = t ~ b I . ' 4 ?.N/kfi-+') ' T i t % ( P-h-71kP b >fi',-..ti'(EC3.,2 1. 26) ' T $ j . ~ f z . sVII$$i$'TtEfl+% Lf: a-8'7 7 1- >fiS-+' ((EC3..2. 1 22) 2 k Q l E C i f = b % , h 3 9 ? D f & D B . % l E
%a&&
7 7 4 / % , % f l+
7j.-X, ~ I V J ( % X - - X ~ ~ E ~ ~ ~ ~ U T ~ ~ > F $ S & , t I . ~ E ~ ; ~ T ~ - ( : Q ~ ~ I W $ ~ I ~ S & & ~ ~ E * ~ ~ ~ ~ & & ,X ~ ~ ~ ~ ~ ~ @ I . ~ ( F G - , J a - 1 2 3 ) D&$$%:$T&& ~ 3 ~ b f ~ f : b l f = ; l j 4 I . ~ I & % 3 j ~ & ,
Nine strains of E~chericlzza colz, two strains of Lactobaczllur aczdophzlus, and one strain of Streptococcur faecalzs were examined for their formations ol invcrtase on thc media containing various sugars
Some of them formed morc or less potent invertasc, which was shown to be P-h-fructosidase (EC 3 2 1 26) rathcr than a-glucosidosaccharasc. This cnzymc togethcr with a-galactosidase (EC 3 2 1 22) of intestinal bacteria might decomposc raffinose, stachyosc, and verbascose con- tained in soybeans and other food legumcs, when eaten by mcn
Introduction
I n Parts I-VII(1-7) of this series we reported the studies exclusively on a-galactosidase (EC 3.2.1.22) of intestinal bacteria. I t was revealed that some strains of E~cherzchza coli(5.6) and other species, notably Streptococcus faecalzr,(7) produced this enzyme adaptively When one mole of U-galactosylsucroses such as rafinose, stachyose, and verbascose is hydrolyzed with this enzyme, the products should be 1, 2, or 3 moles of galactose and 1 mole of sucrose,
Sucrase (or invertase) found in human and other mammalian intestinal rnucosa, acts as a-D-
glucosidasc and is called glucosaccl~aiase(8) or glucosido-invertasc (9) Raffinose family oligosac-
*
This resea~ch has bcen financed in part by the grant (FG-Ja-123) made by thc U S Department of Agriculture under Public Law 480 Part V I I is shown as the rcference (7)**
Present address: Kagawa-kcn Meizen Junior College, Takamatu 760, Japan( @ J I I L ~ ~ ~ ~ f l J ] k P )
* *
*
T o whom correspondence should be addressed****
Present address: Department of Domestic Scicncr, The Tokushima Univctsity of Arts and Sciencc, Tokushima 7'70, Japan (%&f(Bk%%iEfiY$3).68 Sin'itirB KAWAMUKA, Tadasi KASAI and Sumiz8 TANUSI Tcch. Bull Fac Agr Kagawa Univ
charides are not attacked by such a-glucosidasc T h e other kind of invertase acts as 8-fructo-
furanosidase or p-D-fructofuranoside fructohydrolase (EC 3 2 1 26) When we distinguish the
enzyme from inulin-type p-fructosidase, wc call it p-h-fructosidase
Now, even if intcstinal bacteria producc a-galactosidase, the problem rcrnains concerning the
hydrolysis of sucrose formcd When sucrose is freed from galactosyl residues, it is easily decom-
posed by glucosaccharase. There might be, however, hardly no such enzyme in the lower in- testine
Thus we examined the intcstinal bacteria about the ability of decomposing sucrose with p-h-
fructosidase According to NEUBERG and M A N D L , ~ ~ ~ ) "Many bacteria contains invertase (i e.
p-h-fructosidase), e g acetic acid and lactic acid bacteria (Notc that Lactobaczllur aczdophzlur
cxarnined here bclongs to the latter), though not all strains The amounts vary in
E
colz butrather uniform in Thermobacterzum mobzle and sulfur bactcria.(ll>lZ) T h c lower the nitrogen con-
tent of the E colz the lower the invertase activity (I3)"
GOTTSCHALK(~) stated, "Maltase preparations obtained from Sckzzoraccharomycer octosporus,
Nezsserza menzngztzdzr, and E colz(14-18) arc without any effect on sucrose,'' thus indicating the dis-
tinction of two types of invertase
We could not make thorough literature search for p-fructosidase of E colz and other intestinal
bactcria However, this enzyme was cxtcnsively studied on ycasts and seemingly it did not
attract any interests by biochemists studying with E colz
1. Experimental Procedures 1.1 B-,Fraxctosidase activity of E. coli on the sucrose medium
Sucrase activity was examined on the 9 strains of
E
colz, i e Nos 6, 10, 13, 14, 15, 16, 17, 19,and 20 (See Part II(2)). They were selectcd because they consumed sucrose more or less rapidly by settling culture, when the residual sucrose was estimated in the culture liquid. See Table 3
of Part I11 (3)
1.1.1 Cultzuatzon The culture mcdium was cssentially thc same with that uscd f ~ r m c r l ~ : ( ~ )
it consisted of 0 2 g (NH4&304, 0 7 g K2HP04, 0.3 g K H 2 P 0 4 , 0 5 g pcptonc, 0 5 g sucrose, and
0 002 g M g S 0 4 " 7 H z 0 filled up to 100 ml with well watcr. I t was sterilized by autoclaving at
120" for 10 mirl Cultivation was madc a t 37' by shaking for 24 and 48 hours in test tubes
(10 ml).
1.1.2 A5say
of
sucrase actzuzty Centrifuge (12,000 rpm for 20 min at 0-5') after 24 or 48 hours of cultivation to separate into the supernatant (culture liquid) (I) and the precipitated cells.Wash the latter with 10 ml distilled water Centrifuge again Suspend in about 5 ml 100 m M
phosphate buffer (pH 6 8) Sonicate for 15 min Cent1 ifuge to obtain the supernatant (11).
The enzyme in I and PI is called, as before, cxtracellular and intracellular, respectively.
The reaction mixture consisted of 2 ml sucrose solution (1 mglml buffer), 1 ml buffer, and 2 ml
of crude enzyme extract I or 11.
Enzymatic reaction was carried out a t 37' foi 0 5, 3, and 24 hours Add 0.5 ml 5% ZnSOc
and then 0 5 ml 0 2 N Ba(0H)z to 1 ml of the reaction mixture Centrifuge Measure reduc-
ing sugar by the SOMOGYI method observing O D (optical density) a t 530 nm on the supernatant
As the blank was used the mixture heated a t 100' for 5 min to inactivate the enzyme,
1.1.3 Substrate speczjczty of the sucrase contazned zn the rtrains Nos 14, 15, 16, and 20 They were se-
Vol 28, No 60 (1977) ,!3-Fructosidase of Intestinal Bacteria 69
strains and the culture liquid of the strain No 16 were cxamined As the substrates were used (1) maltose, (2) rafinose, and (3) p-nitrophenyl-a-D-galactoside (PNPG)
1.1.3.1 Maltore The reaction mixture consisted of 0 4 ml maltose solution (3 mg/ml buffer), 0 2 ml buffer, and 0 4 ml crude enzyme solution I t was incubated a t 37' for 20 min Glucose formed was determined by glucose osidase/peroxidase method of BAR TON(^^) by measuring O D at 525 nm
1.1.3.2 Rajinore The rcaction mixture consisting of 0 4 ml raffinose solution (4.6 mg/ml buffer), 0 2 ml buffer, and 0 4 ml crude enzyme solution, was similarly incubated as in 1.1.3.1. The reducing sugar was estimated by observing O D at 530 nm
1.1.3.3 PNPG The mixturc consisting of 0 1 ml 10 m M PNPG, 0.05 ml buffer, and 0 35 ml crude cnzyme solution, was incubated a t 37' for 60 min The liberated p-nitroplienol was estimated from O D a t 400 nm
1.1.4 Paper chromatography
of
reaclzon productr from rafinore Incubate 0 5 M raffinose in buffer plus crude enzyme solution from the strains Nos 14, 15, 16, and 20 a t 37' for 20 min Add ZnS04 and Ba(OH)2 to stop the enzymatic reaction. Concentrate, centrifuge, deionize, con- centrate again, and cl~romatograpli on paper by butanol/pyridine/O 1 N HC1 ( 5 : 3 : 2) twice upwards Colorize with 3% p-anisidine-HCl in H20-saturated butanol1.2 B-Fructosidase activity of the strains Nos. 14, 16, and 20 of
E.
coli, the strains Nos. 22 and 23 of Lactobacillus acidophilus, and the strain No. 24 of Streptococcus faecalis, cultivated on the media containing 6 different sugars1.2.1 Cultzvation This was the same as in 1.1.1, except that sucrose was replaccd by one of tlie 6 following sugars: glucose, galactose, fructose, sucrose, maltose, and raffinose The sonicate extract was used as the crude (intracellular) enzyme solution, after cultivation by shaking a t 37' for 24 hours (for 15, 24, and 48 hours for tlic strains Nos 22--24).
1.2.2 Enzyme asray According to the substrate appropriate methods were used as described above For sucrose, maltose, rafinose, and PNPG, see 1.1.2, 1.1.3.1, 1.1.3.2, and 1.1.3.3, respec- tivcly
2. Results and Discussion
'The decomposition of sucrose (and other sugars) by the strains Nos. 21-24 are shown in Figs 1-4 of Part VII (7) Though the strain No. 21 (Proteur mzrabzlzs) decomposed sucrose very slow- ly, this species was omitted in this experiment concerning p-fructosidase.
2.1 B-Fructosidase activity of
E.
coli on the sucrose medium2.1.1 p-Fructoszdare actzvzty
of
11 rtraznr The results of the measurement of p-fructosidase activity described in 1.1.2 is shown in Tables 1 and 2 Thus high sucrase activity was noted for E colz strains Nos 14, 15, 16, and 20 I n general extracellular enzyme activity (a) was low, except in case of the strain No 16 (24 hour cultivation) Intracellular enzyme activity (b) was higher in 24 hour cultivation than in 48 hour cultivation with the strains Nos. 14, 16, and 20, while it was lower after 24 hours than after 48 hours with the strain No 15.Comparison of the data in Tables 1 and 2 in rclation to the time of incubation for enzyme assay shows that the enzyme activity per unit time is highest in case of 30 min. Thus it is presumed that the monosaccharide once formed may be further metabolized by other enzymes contained in the crude enzyme solutions Therefore, the other strains also may have produced sucrase, but the reducing sugar may have been converted to other nonreducing compounds.
Sin'itirB KAWA~IURA, 'I adasi I<AMI and SutnizB TANUSI Tcch Bull Fac Agl Icagawa Univ Table 1 Rclative sucrase activity of (a) the culturc liquid and (b) sonicatc cxtract of 9 strains
of E colz cultivated for 24 hours
-- -- -. - - -.- .-- - - - -- - -- -- - - - Strain No 6 10 13 14 15 16 17 19 20 (a) 0 5 h 022 029 020 032 030 149 02 5 030 ,040 3 03 7 024 020 018 019 .437 024 .028 035 24 "035 066 030 03 5 056 02 1 038 096 03 1 (b) 0 5 h 010 - 012 056 ,720 -- 001 -440 - 010 008 .645 3 - 003 .OOO 345 .825 .001 .a00 015
-
005 3 3 2 24 005 .002 006 013 01 5 01'7 004 004 005Notcs: Thc data are difference of OD,,, values of each test and blank test 0 5, 3, and 24 hours arc the time of incubation for enzyme assay
Tablc 2 Rclativc sucrase activity oi (a) the culturc liquid and (b) sonicatc cxtract of 9 strains of E colz cultivated for 48 hours
.---... -. -. -.~. -. Strain No. 6 10 13 14 15 16 17 19 20 (a) 0 5 h 040 039 035 030 03 5 032 040 042 03 5 3 03 7 051 02 5 034 055 116 036 043 .022 24 024 034 01 7 020 029 020 024 019 022 (b) 0 5 h 044 005 004 .641 .700 195 004 00 1 "275 3 000 000 069 .840 .800 .615 000 000 .562 24 008 005 020 048 019 012 003 062 004
Notcs: Sec Tablc 1.
2.1.2 Substrate speczjicip
of
tlze sucrare of the 4 straznr The result of the experiment 1.1.3 is shown in Table 3. Thus thcsc crude enzyme solutions had only vcry low ability to hydrolyze maltose or PNPG Thus it scems not to contain considerablc tu-qlurosidasc or a-galactosidasc Then this sucrase should bc coilsiclcrcd as p-fructofurai~osidasc, sincc rafiinose was always llydrolyzcd.2.1.3 Paper chr~mato~graplqy OJ reaclzon products from raflnose Thc result of the papcr chromato-
graphy conducted as in 1.1.4 is shown in Tablc 4 T11e parallcl cxpcrimcnt showcd that standard raffinose, melibiose, and fructose showcd R, values and colors of 0 14 (YB), 0 18 (B), and 0 56
(Y),
respectively. Thus the spots Nos 1-3 correspond to raffinose, melibiose, and fructose, re- spectively. The spot No. 4 could not be identified Thus it may be concluded that these crudeTable 3 Hydrolysis of sucrose, maltose, raffinose, and PNPG by E colz strains Nos 14, 15, 16, and 20
Strain No 14 15 16 16 20 Timc of cultivation (h) 24 48 24 24 24 (a) or (b) (b) (b) (b) (a) (b) Sucrose 965 92 7 5 79 090 829 Maltosc 042 - 007 - 002 01 1 050 PNPG 002 00 1 - 002 002 00 1
Notes: PNPG stands for p-nitrophenyl-a-D-galactoside (a) is extra- and (b) is intracellular activity. The values are for respective OD's (cf 1.1.3.1-1.1.3.3) In case of sucrose the measurement was made on reducing sugar as with raffinose.
Vol 28, No.. 60 (1977) /3-Fr uctosidase of Intestinal Bacteria 71 Table 4 The R, values of paper chromatoglams
Strain No. 14 15 16 20
Time of cultivation (h) 24 24 48 24
Spot No 1 12 (YB) 12 (YB) 13 (YB) 1 4 (YB)
2 15 (R) 14 (B) 16 (B) 18 (B)
3 57 (Y) 58 (Y) 56 (?) "57 (Y)
4 70 (VB) 70 (VB) 68 (VB) 70 (VB)
Notes: I n parentheses are given colors: B, brown; V, violet; and Y yellow (with p-anisidine).
enzyme solution contained p-fructosidase, catalyzing hydrolysis of raffinose into melibiose and fructose. There could be found no spots corresponding to glucose or galactose
2.2 ,9-Fructosidase activity of
E.
coli cultivated on media containing 6 different sugars2.2.1
T h e result for Nos 14, 16, and 20 ( E colz) The result of the experiment1.2
forE
ciolis shown in Table 5 The data of Table 5 cannot be compared among different substrates.
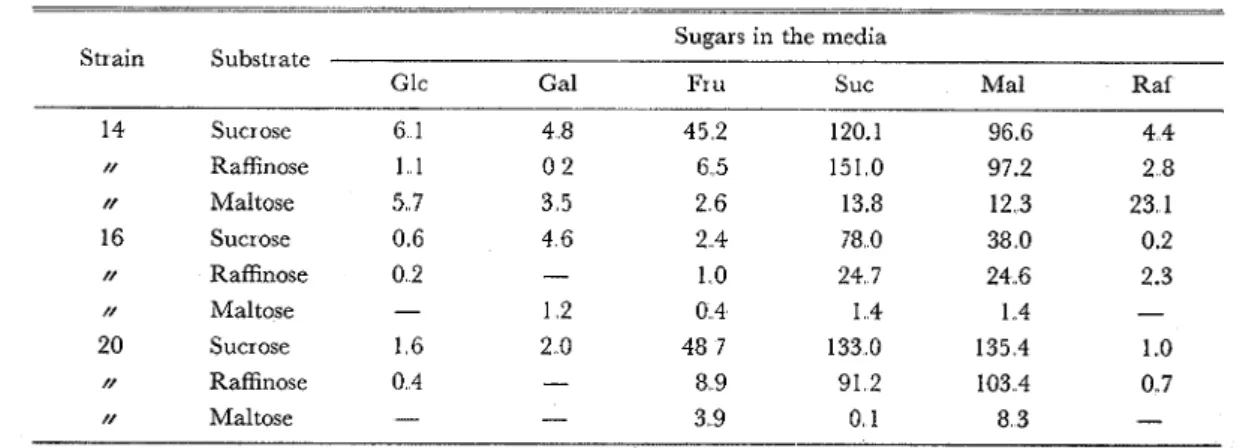
Table 5 Enzyme activities of E colz cultivated on the media containing different sugars
Sugars in the media
Substrate Strain
-
-
Glc Gal Fr u Suc Ma1 Raf
---
-
Sucrose 14 458 360 ( 340) ( 900) ( 724) 330 N 16 04 5 348 182 ( 587) ( 285) 016 N 20 122 153 ( 365) (1.143) (1 016) 0 74 Maltose 14 223 136 102 54 1 48 1 907 // 16-
04 6 016 053 056 - x 20-
- 153 002 .324 - Raffinose 14 082 014 494 (1 153) ( 744) 2 0 9 // 16 026 - 079 1 895 1 894 .I 75 N 20 032 - 68 5 .698 .792 055 PNPG* 14 - 001 0 0 - 002 00 1 00 1 N 20 .OO 1 - 002 - "003 002 - 001 .001Notes: The data are the difference of OD values of each tcst minus blank test The values in paren- theses are those of incubation (in enzyme assay) of 6 min, and the other values are of 60 min
*
I n case of PNPG (p-nitrophenyl-a-D-galactoside) the incubation continued 6 hours.They were calculated to the enzyme activity in the unit mPM substrate decomposed per min
per ml of crude enzyme solution Sincc the a-galactosidase activity on the synthetic substrate
PNPG is negligible, the values for the three sugar substrates are given in Table 6. Thus general-
ly sucrase activity goes parallel with raffinose-decomposing activity, while it has no relation with
very low maltase activity Again this result proves that the E. colz sucrase is not a-glucosidase
but b-fr uctosidase.
As to the sugars of the medium, sucrose was always most effective in producing this fructosidase
in E.. coli. It appeared that maltose had considerable effect in producing this enzyme, but this
72 Sin'itirB KAWAMURA, Tadasi KASAI and SumizB TANUSI Tech. Bull. Fac.. Agr.. Kagawa Univ.
Table 6 The substrates decomposed by E colz cultivated on different sugars (mpM/min/ml)
Sugars in the media Strain Substrate
Glc Gal Fru SUC Ma1 Raf
Sucrose RafEnose Maltose Sucrose Raffinose Maltose Sucrose Raffinose Maltose
has no effect on sucrose (*) Glucose and galactose in the medium had no effect, while fructose
appeared to stimulate the formation of @-fructosidase.
RUTTLOFF et a1 (20) detected p-fructosidase activity in one strain of E colz by using raffinose as
the substrate, though the same strain showed no a-galactosidase activity The other strain of
E.
colz isolated from human feces showed neither p-fructosidase nor a-galactosidase. Also fromour experience there are great differences among the strains of E. colz in forming these enzymes.
2.2.2
The result
forNor.
22-24
See Tables 7-9 for Nos 22 and 23 (Lacidophzlus)
and No24
(S.
faecalis)
.
These results are difficult to evaluate. However, it is clear that all these strains produce
p-
fructosidase, since both substrates sucrose and raffinose are more or less decomposed. This
activity is highest with
S
faecalzr
Sucrose and raffinose in the medium appear to induce the for-mation of this enzyme These enzyme solutions were crude and they seem to contain also some
a-galactosidase and a-glucosidase
Table 7 Relative enzyme activities of Lactobaczllur aczdophzlur, No 22, cultivated on the media con- taining different sugars (OD calculated for 1 hour-incubation in enzyme assay)
Time of Sugars in the media
Substrate cultivation Glc Gal Fru Suc Ma1 Raf
Sucrose 1 5 h -
-
-+
-
- N 24 070 02 1 010-
.011 - x 48 018+
010 - -+
Maltose 15-
--
-- 04 7-
// 24 - 023-
- 030 .023 // 48 008 044 01 1 013 062 .072 Raffinose 15 030 009 --
- - // 24 - - 012+
+
.010 N 48 - - - - - PNPG 48+
+
- -+
fVol 28, No 60 (1977) ,8-Fructosidase of Intestinal Bacteria 73
Table 8 Relative enzyme activities of L acidophzlur, No. 23, cultivated on the media containing different sugars (OD calculated for 1 hour-incubation in enzyme assay)
- . - -- -- - -- - - - - - - - - - -- -- - - -
Time of Sugars in the media
Substrate --- -
-Glc Gal Fru SUC Ma1 Raf
-- Sucrose 15 h - -I- 01 1 - - - // 24
+
- 139 .011 - - // 48 .157 043 030 .943+
.206 Maltose 15 --
+
+
-+
N 48 f - f-
+
Raffinosc 15 056 - f .028 -+
I / 24-
+
023 007 03 1-
PNPG 48 i- ,011 ,045+
-
+
Table 9 Relative enzyme activities of Streptococcur faecalzr, No 24, cultivated on the media con- taining different sugars (OD calculated for 1 hour-incubation in enzyme assay)
Sugars in the media
Substrate
,
Gal Fr u SUC Ma1 Raf
- Sucrose 1 5 h 219 - .153 1.105 065 .032 // 48 188 043 030 .943 003 .206 Maltose 15 - -
+
07 1 064 I / 24+
- - - 007 038 Raffinose 15 --
004 .057 - .001 // 24 00 7+
+
.052+
.004 N 48+
012 018 .040 - - PNPG 48 005 112 003 007 007 033 3. ConclusionSome strains of E coli formed powerful enzyme catalyzing the hydrolysis of sucrose. This
enzyme was proved to be P-fructofuranosidase rather than a-glucosidase This is very con-
venient in decomposing sucrose as well as galactosyl sucroses (raffinose and stachyose) in distal part of the intestine
Stre#tococcur faecalzr (No 24) produced /3-fructosidase induced by sucrose and raffinose Lac-
tobaczllur aczdophzlur (Nos 22 and 23) produced this enzyme in smaller amounts Presence of
small amounts of a-galactosidase and a-glucosidase was noted
A study(21) made in collaboration with YOSHIDA of in vivo utilization of soybean oligosac-
charides by rats supported our presumption, that at least a part of raffinose and stachyose was
utilized by a-galactosidase and p-fructosidase produced by some intestinal bacteria I t showed
that about 90% of soybean oligosaccharides were absorbed, while only about 50% of them were
74 Sin'itirB KAWAMURA, Tadasi KASAI and SumizB TANUSI Tech Bull Fac Agr Kagawa Univ
corresponding to the amount of raffinose and stachyose) occurred owing to absorption through the enzymatic activity of intestinal bacteria of rats.
Acknowledgments
Thanks are duc to the members of Northern Rcgional Research Laboratory of the U
S
Department of Agriculture, Peoria, Illinois, especially to DrR J
DIMLER, DrJ
C COWAN, and Dr W J WOLF, for their encouragementsWe are thankful to Dr. I<ei ARIMA and Dr. Hiroshi IIZUKA, Professors of the University of Tokyo, who donated us 20 strains of Ercherichia colz and 4 other strains of intestinal bacteria, Proteur mzrabzlzr, Lactobaczllus aczdophzlus, and Streptococcur faecalzr used in this research.
Valuable discussions were given by Dr. Hiroshi SUZUKI, Dr Kei YAMANAKA, and the late Dr Teiiti NARASAKI of Kagawa University.
REFERENCES ( 1 ) KAWAMURA, S
,
MIYAKE, T., NARASAKI, T :Tech Bull Fac Agrzc. Kagama Unzv (Kagawa
Daigaku Nogakubu Gakuzyutu Hokoku) 20, 2 5-32 (1 968).
( 2 ) KAWAMURA, S
,
KASAI, T : Ibid 33-40 (1968)( 3 ) KAWAMURA, S
,
KASAI, T. : Ibzd. 41 -8 ( 1968) ( 4 ) KAWAMURA, S., TADA, R,
KASAI, T. : Ibzd21, 90-103 (1970)
( 5 ) KAWAMURA, S
,
TORIGOE, H,
KASAI, T :Ibzd 104-9 (1 970)
( 6 ) KAWAMURA, S
,
KASAI, T,
TANUSI, S, : Agrzc Biol Chem 40, 64 1-8 (1976)( 7 ) KAWAMURA, S
,
KASAI, T,
TANUSI, S : Tech Bull Iiac Agrzc Kagawa Unzv 27, 1 17-24 (1976)( 8 ) GOTIISCHALK, A : a-D-Glucosidases The Enzymer, 1st ed (ed SUMNER, J B
,
MYRBACK, K ) 1, 551-82 (1950)( 9 ) MYRBACK, K : Invertase The Entymer, 2nd ed (ed BOYER, P D , LARDY, H , MYRBACK, K ) 4, 379-96 (1960)
(10) NEUBERG, C
,
MANDL, I : Invcrtase TheENZYMES, 1st ed
,
1, 527-50 (1950).(1 1) FORSSMAN, S : Bzochem Z 264, 231 (1933) ;
through (10)
(12) HOFMANN, E : Ibzd 273, 198 (1934) ; through (10)
(13) VIRIANEN, A I , LEY, J : Arch Biochem 16,
169 (1948); through (10).
(14) KARSTROM, H : Bzochem 2. 231, 399 (1931) ; through (8)
(15) VIRIANEN, A. I : Ibzd 235, 490 (1931); through (8)
(16) MYRBACK, K.: Z Phy~zol Chenz 198, 196 (1931); through (8)
(17) TAUBER, H
,
KLEINER, I S : J Gen. Phy- rzol. 16, 767 (1937); through (8)(18) HOICHKISS, M : J Bacterzol 29, 391 (1935);
through (8)
(19) BARTON, R R : Anal Biochern. 14, 358-60
(1966).
(20) RUIILOFF, H
,
TAUFEL, A , HAENEL, H,
TAUFEL, K : Nahrung 11, 47-54 (1967).(21) YOSHIDA, A , UMAI, A , KURAIA, Y , KAWAMURA, S : Eyo To Slrokuryo (J Jpn
Soc Food Nutr ) 22,262-5 (1969) (in Japanese) (Received September 30, 1976)