若手の会自由集会報告
ヤドカリの貝殻という生息環境―貝殻を介した共生関係―
Gastropod shells used by hermit crabs, as secondary habitats for various invertebrates
吉川晟弘・後藤龍太郎・朝倉彰
Akihiro Yoshikawa, Ryutaro Goto, and Akira Asakura
はじめに 十脚甲殻類・異尾下目に属するヤドカリ類は,巻 貝の空き殻をシャルターとして利用するユニークな 生態を持っている.彼らは深海から潮間帯,海岸付 近の陸上などにも生息しており,海洋を中心に様々 な環境への適応を成功させた生物のひとつだといえ る(Reese, 1969).ヤドカリは多様な動物と,貝殻を 介して共生関係を築いている (Williams & McDer-mott, 2004).本来ならば巻貝の死殻はヤドカリに使 用されない限り海底の基質に埋没してしまうが,ヤ ドカリがこれを住処として利用することにより,そ の他の動物も貝殻を生息場所として利用できる.つ まり,ヤドカリは深海から潮間帯にかけての様々な 海洋環境において,他の生物が利用できる環境を作 り出すエコシステムエンジニアとしての重要な役目 を果たしている(Williams & McDermott, 2004).
これまで550種以上の動物が,ヤドカリの貝殻内 部 や 外 側 か ら 発 見 さ れ て い る(Williams & McDer-mott, 2004).彼らはヤドカリと共生することによ り,安定的な食料の供給や移動能力の獲得,基質へ の埋没の回避,繁殖機会の増加等の様々なメリット を受けるとされる(Williams & McDermott, 2004).し かし,これほど多くの共生生物が知られているにも 関わらず,彼らの宿主利用様式(ヤドカリおよびヤ ドカリが利用している貝殻への選好性)や貝殻内部 という狭隘空間への形態的適応方法などはほとんど 解明されていない.そこで,私たちは共生生物がヤ ドカリの貝殻という生息環境をどのように利用し, ヤドカリとの共生のためにどのように形態的・行動 的に適応しているのかを解明すべく研究を行なっ た.本稿では著者らが行った“貝殻を介したヤドカ リとの共生”に関する2題の研究,“ヒラフネガイ の 宿 主 利 用 様 式 と 形 態 的 適 応(Yoshikawa et al., 2018a)”および“カブトヤドカリによるモンバンイ ソギンチャクの移送行動(Yoshikawa et al., 2018b)” を紹介する. ヒラフネガイの宿主利用様式と形態的適応 ヤドカリの背負う貝殻を住処として利用している 動物の中で,軟体動物門は最も種数が多いグループ である(Williams & McDermott, 2004).これまで多板 綱からは1種,腹足綱からは36種,二枚貝綱からは 25種の動物がヤドカリの貝殻から報告されており, そ のほ とん ど種 が片 利共 生で ある とさ れて いる (Williams & McDermott, 2004).ヤドカリ共生性の軟 体動物の中でも腹足綱の種がその大半を占めてお り,その中で特にカリバガサガイ科の種がその大半 を占めている(Williams & McDermott, 2004).ちなみ に, 同 科 のCrepidula属 お よ びErgaea属,Grandi-crepidula属,Maoricrypta属でヤドカリの貝殻を住 処とする種が報告されている(Vermeij, 1989; Mar-shall, 2003; Williams and McDermott, 2004; Low and Tan, 2014).
今回私たちは,カリバガサガイ科の一種ヒラフネ 京都大学フィールド科学教育研究センター瀬戸臨海
実験所
〒649–2211 和歌山県西牟婁郡白浜町459
Seto Marine Biological Laboratory, Field Science Educa-tion and Research Center, Kyoto University, 459 Shiraha-ma, Nishimuro, Wakayama 649–2211, Japan
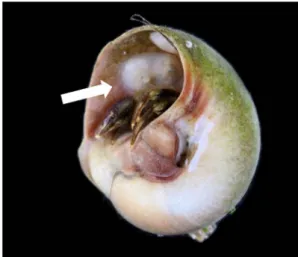
ガイErgaea walshi (Fig. 1)を対象に,宿主利用様式 およびヤドカリ貝殻内部への形態的適応についての 研究行った.ヒラフネガイの採集は三重県鳥羽市菅 島の砂泥および転石帯と,和歌山県和歌山市和歌浦 の干潟で行った.両採集地点からは,24種で構成 される合計419個体の巻貝が採集され,うち363個 体が7種のヤドカリにより使用されていた.採集さ れた巻貝の中で,エゾタマガイやツメタガイ,スク ミリンゴガイ,アカニシ,イボニシなどの比較的大 型の種から2個体以上のヒラフネガイが確認され た.また,採集された7種のヤドカリのうち,6種 (コブヨコバサミ,ケブカヒメヨコバサミ,ユビナ ガホンヤドカリ,テナガツノヤドカリ,ベニホンヤ ドカリ)が利用する貝殻内部からヒラフネガイが発 見され,比較的小型の種であるブチヒメヨコバサミ が利用する貝殻からは発見されなかった. ヒラフネガイの出現を最も良く説明できる要因を 明らかにするために,一般化線形混合モデルを用い て解析した.ヒラフネガイの有無を目的変数,ヤド カリの種および体サイズ,貝殻の種およびサイズ (殻長および殻幅)を説明変数に用いたモデルを作 成し,各モデルの当てはまりの良さを,それぞれの AICを比較することで検証した.その結果,巻貝の 横幅およびヤドカリの体サイズを説明変数として用 いたモデルの順に,本種の出現を良く予測してい た.Figure 2にも示すように,どちらのサンプリン グ地点においても,比較的横幅の大きな巻貝殻から Fig. 1. テナガツノヤドカリが利用する貝殻に共 生するヒラフネガイ(白矢印).
Fig. 2. ヒラフネガイの貝殻利用パターン(Yoshikawa et al., 2018aのデータを元に作図);(i), 三重県鳥羽市菅島
で採集された巻貝,(ii), 三重県鳥羽市菅島におけるヒラフネガイの貝殻利用パターン,(iii), 和歌山県 和歌浦で採集された巻貝,(iv), 和歌山県和歌浦におけるヒラフネガイの貝殻利用パターン.
ヒラフネガイが見つかる事が多い.つまり,本研究 の結果は,ヒラフネガイはある特定の種(ヤドカ リ,巻貝共に)と共生しているわけでは無く,体サ イズの大きなヤドカリおよび殻幅の大きな巻貝に着 底できたものが生存している可能性を示唆している (Yoshikawa et al., 2018a).
続いて,ヒラフネガイの形態的適応様式を調べる ために,本種の殻高に着目し,殻高と接地面積の成 長パターンを他のカリバガサガイ科の23種(非ヤド カリ共生性,19種;ヤドカリ共生性,4種)と比較 した.すると他のカリバガサガイ科の種は成長に合 わせて接地面積および貝殻高を継続的に成長させて いるのに対して,ヒラフネガイは接地面積の成長は 他のカリバガサガイ科と同じく継続的な成長を見せ るものの,貝殻高の成長はある時期(5.0 mm付近) で抑制されている事が示唆された.ヤドカリは自ら の成長に合わせてさらに大きな貝殻へと引っ越すた め,ホストがいない貝殻内部に取り残されること は,ヒラフネガイにとっては,砂や泥などへの付着 基盤の埋没に加え,食料の獲得,繁殖参加,酸素の 獲得等の面で大きな不利益を被るものと思われる. したがって,特徴的な成長様式を持つ事で貝殻内部 の体積をなるべく広く保ち,ヤドカリが貝殻を交換 することを防いでいるのかもしれない. ヤドカリによるモンバンイソギンチャクの移送 行動 ヤドカリと相利共生関係を築いている生物の最も 有名な例としてイソギンチャクが知られている.少 なくとも3科15属41種のヤドカリがイソギンチャ クと共生しており,7科14属35種のイソギンチャ クがヤドカリと共生する (Williams & MacDermott, 2004; Antoniadou et al., 2013).ヤドカリは成長に合 わせて貝殻を交換しなければならず,その際にヤド カリはハサミ脚と歩脚を用いてイソギンチャクに一 定間隔の刺激を与える.するとイソギンチャクはそ れに応じるように接着部分を緩め,ヤドカリにより 移送される.このようにヤドカリ–イソギンチャク 間では,お互いが共生関係を維持するために特徴的 な行動を進化させている.しかし,ヤドカリの貝殻 上の共生性イソギンチャクの移送に関するほとんど の研究は,クビカザリイソギンチャク科のイソギン チャクに関するもので,そのほかの共生性イソギン チャクがヤドカリからの刺激に反応するかどうかに ついての研究は少ない.そこで著者らは,ナゲナワ イソギンチャク科に属するモンバンイソギンチャク とその宿主のヤドカリにおいても(Fig. 3), 移送行動 が見られるかどうかを確かめた (Yoshikawa et al., 2018b). カブトヤドカリとその利用貝殻についているモン バンイソギンチャクを和歌山県西牟婁郡白浜町畠島 の転石帯で採集した.貝殻交換を促すためにヤドカ リが使用している巻貝の貝殻をペンチやニッパーを 用いて少し破壊し水槽内に戻した (Fig. 3).続いて ヤドカリの体サイズに合った巻貝空殻を水槽内に置 いた.そしてビデオカメラを用いてその後の行動を 記録した.するとモンバンイソギンチャクもヤドカ リにより移送され,かつヤドカリによる刺激に反応 す る よ う に 接 地 部 分を緩 め ること が 観察さ れ た (Yoshikawa, et al., 2018b).ヤドカリはモンバンイソ ギンチャクを貝殻交換前と同様の位置,すなわち貝 殻 の 入 り 口 に 再 度 取り付 け ること が 確認さ れ た (Fig. 4).今回の観察結果は,これまで知られていな いイソギンチャクの系統でもヤドカリの刺激に対す る協調的な反応が進化していることを示唆してい る.カブトヤドカリはベニヒモイソギンチャクと共 生する場合も知られるが,その際カブトヤドカリは ベニヒモイソギンチャクを貝殻の外側に取り付け Fig. 3. カブトヤドカリ(SL=6.3 mm; female) が利 用する貝殻の入り口付近に取り付けられ たモンバンイソギンチャク.
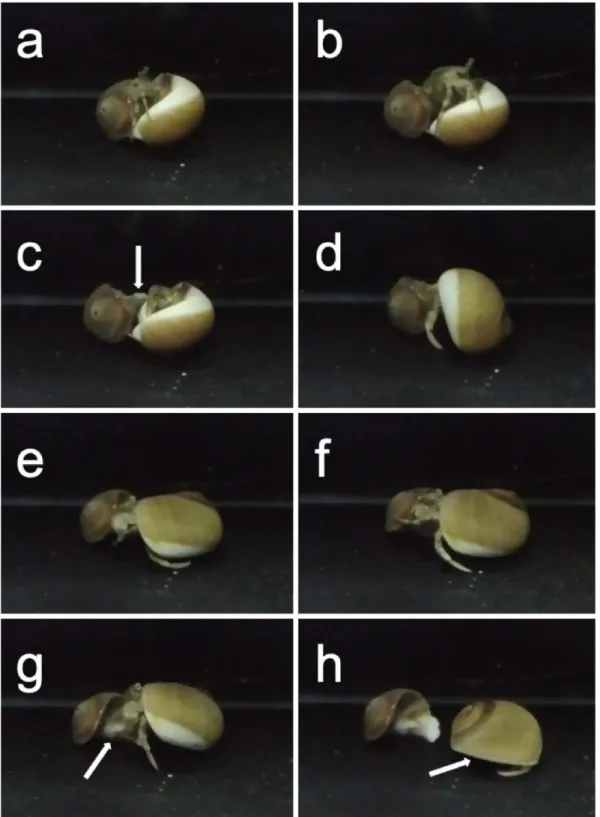
Fig. 4. カブトヤドカリによるモンバンイソギンチャクの移送行動(Yoshikawa et al., 2018bのデータを元に作
図)(白矢印=モンバンイソギンチャクの位置):a, ヤドカリによる貝殻の選定;b~c, ヤドカリの引越 し行動;d~f, ヤドカリのモンバンイソギンチャクへの突き行動;g, ヤドカリによるモンバンイソギン チャクの取り外し;hヤドカリによるモンバンイソギンチャクの取り付け行動.
る.したがってカブトヤドカリは共生イソギンチャ クの種によって取り付ける位置を変更している可能 性がある.もしくはイソギンチャクのサイズごとに 取り付ける位置を変更している可能性も考えられ る.また,ヤドカリにとって貝殻の入り口は,個体 間の闘争,および天敵からの捕食の際には最大の弱 点となり得る.したがって,カブトヤドカリは本種 を取り付けることで,この弱点を補完している可能 性がある.しかし,本種が貝殻の入り口付近に取り 付けられることにより,外側に取り付けられる種と 同様の機能(例えば天敵であるタコから捕食の回 避,Ross, 1971)を果たしているかを観察した例は 無く,今後検証する必要がある. 文 献
Antoniadou, C., Vafeiadou, A. M., & Chintiroglou, C., 2013. Symbiosis of sea anemones and hermit crabs in temper-ate seas. In Camisão, A. F. & Pedroso, C. C., (eds), Symbiosis: Evolution, Biology and Ecological Effects. NOVA Science, New York, 95–118.
Low, MEY., & Tan, SK., 2014. Ergaea H. Adams & A. Ad-ams, 1854, the correct genus for Crepidula walshi Reeve, 1859, with nomenclatural notes on Syphopatella
Lesson, 1831, Siphonipatella L. Agassiz, 1846, and the incorrect subsequent spelling “Siphopatella” of authors (Gastropoda: Calyptraeoidea: Calyptraeidae).Occ Mollusc Paper, Publ Malac Sun Reg 3: 11–14. Marshall, B. A., 2003. A review of the recent and late
Ceno-zoic Calyptraeidae of New Zealand (Mollusca: Gas-tropoda).Veliger 46(2): 117–144.
Reese, E. S., 1969. Behavioral adaptations of intertidal her-mit crabs. Am Zool 9: 343355.
Ross, D. M., 1971. Protection of hermit crabs (Dardanus spp.) from octopus by commen- sal sea anemones (Cal-liactis spp.).Nature, 230: 401–402.
Vermeij, G. J., 1989. Habitat and form of Crepidula grandis in Japan, with comments on habitat specialization in ca-lyptraeid gastropods. Nautilus 103: 89–91.
Williams, J. D., & McDermott, J. J., 2004. Hermit crab bio-coenoses: a worldwide review of the diversity and natu-ral history of hermit crab associates. Journal of Experi-mental Marine Biology and Ecology, 305:1–128. Yoshikawa, A., Goto R., & Asakura, A., 2018a. Morphology
and habitats of the hermit-crab-associated calyptraeid gastropod Ergaea walshi. Zoological Science, 35 (6): 494–504.
Yoshikawa, A., Goto R., & Asakura, A., 2018b. Transfer of the gatekeeper sea anemone Verrillactis sp. (Cnidaria: Actiniaria: Sagartiidae) between shells by the host her-mit crab Dardanus deformis (H. Milne Edwards, 1836) (Decapoda: Anomura: Diogenidae).Crustacean Re-search, 47: 55–64.