二極性および四極性担子菌におけるブラー現象に関する研究
(
S
t
u
d
i
e
s
on t
h
e
B
u
l
l
e
r
phenomenon
inb
i
p
o
l
a
r
and t
e
t
r
a
p
o
l
a
r
mushrooms)
野 上 友 美
A
D
i
s
s
e
r
t
a
t
i
o
n
For D
o
c
t
o
r
Degree
S
u
p
e
r
v
i
s
o
r
:
P
r
o
f
.
Yutaka
Ki
tamoto
The United Graduate S
c
h
o
o
l
o
f
A
g
r
i
c
u
l
t
u
r
a
l
S
c
i
e
n
c
e
s
,
T
o
t
t
o
r
i
U
n
i
v
e
r
s
i
t
y
,
Department o
f
M
i
c
r
o
b
i
a
l
B
i
o
t
e
c
h
n
o
l
o
g
y
目次 第1章 緒言一一一一一一一一一一一---一一一一一一一一一一 1 第 2章二極性担子菌きのこ
P
h
o
l
i
o
t
anameko
のダイ・モン交配 における核の選択 一一一一一一一一一一一一一一一一一一一一一一一 8 第3章 四極性担子菌きのこ Flammulinav
e
l
u
t
i
p
e
s
のダイ・モン 交配における核選択の規則性 一一一一一一一一一一一一---一一一ーー一一 20 第 4章 四極性担子菌Len
加u
l
ae
d
o
d
e
s
およびP
l
e
u
r
o
t
u
so
s
t
r
e
a
t
u
s
の ダイ・モン交配における核選択の規則性一一一一一一一一---時一 37 第5章 四極性きのこ Flammu
必'lav
e
l
u
t
i
p
e
s
の不和合性ダ、イ・ モン交配における不和合性因子の体細胞紐換による新核 形成の検討 一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一 48 摘要 一一一一一一一一一---一一---一一一一一一一一一一一一一一一---一一---一一 61 Abstract 一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一---一一一一 65 謝 辞 一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一 71 引用文献一一---一ー一一一一一一一一一一一一一一一一一一一一一一一一一 72 公表論文のリスト 一一一一一一一一一一一一一一一一一一一一一一一一---一一一 76第1章 緒論 多 く の 食 用 き の こ は 性 的 に ヘ テ ロ タ リ ズ ム の 担 子 蓄 に 属 し て い る (Whitehouse, 1949).その菌糸体や子実体の細胞の核相は,一つの細胞に 異なったふたつの半数体の核で構成されるヘテロカリオンである (Fig1-I)• 一般的な担子菌の生活環を Fig1-2に示す.きのこにおけるー核菌糸体の 二核化は交配型が異なる二つのー核菌糸体,例えばこ極性きのこの場合で
は AIXA2,四撞性きのこの場合は AIBIXA2B2あるいは AIB2XA2Bl
などの相補的な不和合性因子をもっ一核菌糸細胞同士の接合により交配が 行われることで完遂する. 一方, Buller(1930, 1931)は ヒ ト ヨ タ ケ 属 の 四 撞 性 き の こ で あ る Coprinus lagopusのー核菌糸体の周縁部に同菌の二核菌糸体を少量接種 したところ,一核菌糸体が速やかに二核化し,その菌糸体コロニーの全周 縁部に二核菌糸体が新生することを見い出した.この現象はその後,多く の き の こ で も 見 ら れ る こ と が わ か り , 発 見 者 の 名 に 国 ん で Buller現 象
(Quintanilha, 1937),もしくはダイ・モン(Di-mon)交配(Papazian,1950)
と呼ばれている.この現象は,二核菌糸体からー核蕗糸体へ核が移動して 新しい供役核の組み合わせができ,二核化するものである.ダイ・モン交 配にはいろいろな組み合わせがあるが, Raper(1966)は次のように分類し ている. まず,ー核菌糸体の核が二核菌糸体の両方と和合できる場合,例えば, 1
-Heterotha1lisrn
B
i
p
o
l
a
r
(
A
f
a
c
t
o
r
)
A
l
-一+
A
2
一一+T
e
t
r
a
p
o
l
a
r
(
A
a
n
d
B
f
a
c
t
o
r
s
)
Hornotha1lisrnP
r
i
m
a
r
y
S
e
c
o
n
d
a
r
y
A
I
B
l
A
I
B
2
一一+
A
2
B
l
-ー+A
2
B
2
一件。。。宵
z令 め
→
e
。?ぉ布石〉
F
i
g
"
1・L.M
a
t
i
n
g
s
y
s
t
e
m
s
i
n
B
a
s
i
d
i
o
m
y
c
e
t
o
u
s
m
u
s
h
r
o
o
m
s
.
2-¥pm
円ヤ
og阻 y向
包
Y
γ
e
的iおo
悶向
V
γ
抑 問p
oru凶u血laヴ
げ
げ
γ
ω
ω
N) Fruit-body/
β
p討IDordium 的)でこ , 、 、 f . oidia_¥.&
-一
…
¥ フ ? 官ω) - - ¥ ¥ 、、 /、 、〆 、、 -、、 J〆 、 、 、 、 、 、 - - - " " . . . . 、、、 、、
、
、
、
、
、
、 、 、
、 、
、 、、
1 t、
1 、 1¥i
. ' i J J ' J J J J 〆 a〆 え了 /' / 4ノヒ Haploid basidiospores 2 0 . 包 a 叫 e G l i l t V ぬ Ym v e
AyByHdU
H
併
MU
/ラ /" oidia(N) ノ :: Asexual cycle、
、 、、 、 AxBx Fusion Monokal'yotic mycelia (N)F
i
g
.
1・2.L
i
f
e
c
y
c
l
e
o
f
b
a
s
i
d
i
o
m
y
c
e
t
o
u
s
mushrooms.
3二擾性きのこの場合 (Al+A2)
x
A3, 四極性きのこの場合 (A1Bl+A2B2)x
A3B3という和合性組み合わせのダイ・モン交配である.この場合,二極
性 き の こ で は (Al+A3)と(A2+B3), 四 極 性 き の こ で は (A1Bl+A3B3)と
(A2B2+ A3B3)の2種類の二核菌糸体が同じ頻度で出現すると考えられる が,二核菌糸体の二つの核は一核菌糸体の核と等しい確率で移動せず,二 核菌糸体の核の構成によりー核菌糸体に二核菌糸体の核のどちらか一方が 移動し,二核化の割合に偏りが生じることが明らかにされた(Quintanilha, 1939 ; Oikawa, 1939; Papazian, 1950;木村, 1966, 1975). つぎに,ー核菌糸体の核が二核菌糸体の一方の核とだけ和合できる場合, 例えばこ極性きのこの場合(Al+A2)XAlまたは A2,四極性きのこの場合
(A1Bl+A2B2)XA1Bl, A1B3, A3Blとし、う半和合性組み合わせのダイ・
モン交配である.この場合では二核菌糸体の二つの核のうち和合性を示す 核のみが一核菌糸体に移動して二核化が生じるものと考えられているが (Buller, 1931; Quintanilha, 1939),ニ核菌糸体の二つの核とも移動し たと考察した相反する報告もある(Dickson,1934, 1936; Quintanilha, 1939 ;木村, 1958 b). さらに,一核菌糸体の核が二核菌糸体のどちらの核とも和合できない場 合,二極性きのこの場合は該当組み合わせはないが,四極性きのこの場合
では (A1Bl+A2B2)
x
A1B2または A2Bl とし1う不和合性組み合わせのダイ・モン交配が存在する.この組み合わせでも一核菌糸体の二核化が生じ,
その場合,二核菌糸体の二核開で体細胞組換えにより和合性の新核が生じ
る仮説(Quintanilha,1939; Ellingboe, 1964),あるいは二核菌糸体の二
-つ の 核 と も 移 動 し た と す る 相 反 す る 仮 説 が 提 起 さ れ て い る(Papazian, 1950). 本 研 究 で は , ま ず , こ の ダ イ ・ モ ン 交 配 に つ い て 二 掻 性 担 子 菌 で あ る
P
h
o
l
i
o
t
a
nameko
および四極性拒子菌であるFlammulina v
e
l
u
t
i
p
e
s
とL
e
n
t
i
n
u
l
a
edodes
を供試菌として,和合性および半和合性ダイ・モン交配 時に二核菌糸体と一核菌糸体が接合してー核菌糸体の二核化が生じるか, またそのこ核化の規則性について検討した. ところで,二極性担子菌のきのこであるP
h
o
l
i
o
t
anameko
(Arita, 1964, 1979 Masuda et al., 1995)お よ び 四 極 性 担 子 菌 の き の こ で あ るFlamm
u
l
i
n
a
v
θ,'
l
u
t
i
p
e
s
(Ashan, 1952; Aschan-Aberg, 1960)の二核菌糸体コロニーの先端部で、は一核菌糸体が生じることが報告されている.また,
P
h
o
l
i
o
t
a
nameko
の二核菌糸体コロニー外縁部から分離した一核菌糸体の 大部分は二核菌糸体の片方の核型を示したことから,二核菌糸体のー核化において核の選択に比較優位性が機能すること,その比較優位性に序列が
あることが見いだされている(Masudaet al., 1995). また, Fig1合 に 示
すように
Flammulinav
e
l
u
t
i
p
e
s
(Brodie, 1936),Coprinus C
I
n
e
r
e
u
s
(Raoand Niederpruem, 1969),
P
h
o
l
i
o
t
a
nameko
(Arita, 1979; Cao et al. ,1999)の二核菌糸体から形成されるオイディアの多くは一核性であること が 報 告 さ れ て い る . さ ら に ,
P
h
o
l
i
o
t
a
nam
θ'
k
o
(Cao et al., 1999)Flammulina v
e
l
u
t
i
p
e
s
(Kitamoto et al., 2000)の二核菌糸体からの一核 性オイディア形成における核選択にも同様な比較優位性の序列が見いださ れている.一方,二核菌糸体が一核化しない,例えばPleurotuso
s
t
r
e
a
t
u
s
5 -,.O
轡。
砂喝事。
。
ゆ吋毎 秒4静一 ① @ ① ① ① ①
F
i
g
1-3.O
i
d
i
u
l
n
f
o
r
m
a
t
i
o
n
by t
h
e
segmentation o
f
c
e
l
l
u
n
i
t
s
from branched m
y
c
e
l
i
a
o
f
P
h
o
l
i
o
t
a
nameko.
-などのきのこにおいてもダイ・モン交配における親二核菌糸体での核の選 択的移動が考えられるが,それを例証した論文は見られない. 以上の報告から,ダイ・モン交配は二核菌糸体と一核菌糸体が接合して 生じるのではなく,二核菌糸体コロニーの先端部が脱二核化して生じたー 核菌糸体あるいは一核性オイディア由来の一核菌糸体と相補的な核をもっ 一核菌糸体が接合して生じる可能性も考えられる.そこで,二種の担子菌
Pho
Jio
t
a
nameko
およびFlammu
Jinav
e
l
u
t
i
p
e
s
の二核菌糸体においてー 核菌糸体を生じることが普遍的に起きているのかを調査した.さらに,一 核化における核選択の序列とダイ・モン交配における序列とを比較検討し た.また,親二核株の一核化が例証されていないL
θn
t
i
n
u
l
aedodes
につい ても核選択の序列について検討を行った. さらに,不和合性ダイ・モン交配において,体締胞組換が生じて和合性 の核が新生するかどうかを ,Flammu
Jina v
e
l
u
t
i
p
e
s
を供試材料として,そ のー核性オイディアを分離して不和合性因子を解析することにより解明し た. - 7第2章 二極性担子菌きのこ Pholiota namekoのダイ・モン交配における核の選択 緒言 多 く の 食 用 き の こ は 性 的 に ヘ テ ロ タ リ ズ ム の 担 子 菌 類 に 属 し て い る (Whitehouse, 1949).その菌糸体や子実体の細胞は,一つの細胞に異なった 交配型の二つの半数体の核で構成されるヘテロカリオンである.きのこにお ける一核菌糸体の二核化は交配型が相補的な二つのー核菌糸体により交配 が行われることにより完遂する. Buller (1930, 1931)は,自極性のきのこ である Coprinuslagopusのー核菌糸体の周縁部に向菌の二核菌糸体を接種 したところ,一核菌糸体が速やかに二核化し,一核菌糸体の全周縁部に二核 菌糸体が新生することを見出した.それ以誇,多くの研究者が間様の二核化
について報告している (Dickson,1934;木村, 1957a, 1957b, 1958a, 1958b ;
Oikawa 1939 ; Quintanilha, 1937, 1939). このように,二核菌糸体(親ニ
核株)によってー核菌糸体(親ー核株)が二核化する現象はダイ・モン交配
(Papazian, 1950)あるいは発見者の名にちなんで Buller現象と呼ばれて
きた (Quintanilha,1937).自極性きのこのダイ・モン交配には,和合性 (AB
X (ab+ A ) B} ) ) ,半和合性 (ABX(AB十ab)),不和合性 (ABX(Ab十aB))の
組み合わせが存在する.[9極性きのこにおいて Quintanilhaにより見いださ
れた Buller現象に関するこつの主課題をまとめると, (a)和合性・半和合
-性組み合わせによるダイ・モン交配において,親二核株の二つの核うちの一
つの核が選択されるかどうか. (b)体細胞組換え,すなわち不和合性ダイ・
モン交配時に和合性の遺伝子型が新生するのかどうかである (Raper,1966).
しかし,二極性きのこの場合,二核菌糸体と一核菌糸体のすべての交配組み
合わせは,和合性 (Al
x
(A2+ A3) )および半和合性 (Alx
(Al十A2))の二種類のダイ・モン交配しか存在しない. Raper (1966) によると二極性交配系 は四極性の交配系よりも核選択の研究において,四極性きのこで生じる可能 性のある交配型A国子と B因子聞の相互作用を避けることができる有用な実 験系と言及している. Pholiota namekoはアジア諸国,特に日本で栽培されているこ掻性の食 用きのこである.本菌は菌糸体培養時にコロニーの周縁部分に常に菌糸体の ー核化が生じる (Arita, 1964, 1979;Masuda et a1. ,1995). 二核菌糸体の ー核化は,本菌のオイディア形成でも生じる (Caoet a1. , 1999). 本菌にお け る 二 核 菌 糸 体 あ る い は オ イ デ ィ ア 形 成 に よ る 一 核 化 で は 核 の 選 択 が 介 在 する (Masudaet a1., 1995 ; Cao et a1., 1999). すなわち,二核菌糸体コロ ニ ー の 外 縁 部 か ら 分 離 さ れ た ほ と ん ど の 一 核 菌 糸 体 は 親 二 核 株 の 片 方 の 核 型を示す (Masuda et a1., 1995). それ故,菌糸体のー核化における優位核 の選択が介在する.菌糸体のー核化における親二核株からの核選択の比較優 位性には序列が見いだされている.また,同ーの核選択の規則が本菌の一核 性オイディア形成においても見いだされている (Cao et a1., 1999). そこで本研究では,不和合性因子を異にする栄養要求性変異株を用い,二 撞性きのこの和合性および半和合性のダイ・モン交配において同様な核選択 9
-の機能の介在を検証した.さらに本菌における和合性および半和合性組み合 わせのダイ・モン交配における核選択の比較優位性の序列について検討を行 った. 材料および方法 供試菌株 本研究には ,P. namekoのー核株 NAll,NF8, NX6を紫外線照射することに より作出した三つの栄養要求性突然変異株 NA11-26 (Al/ade-), NF8-267 (A5/pdx-), NX6-582 (A6/his-)を用いた.栄養要求性突然変異株の作出は 衣笠および北本(1
9
9
7
)
らの方法に従った.菌糸体の培養は 12mlのPDA
培地 (Nissui Pharmaceutical Co. Ltd) を分注したプラスチック製のペト リ皿 (Iwaki,9
0
x 15m
m
)
で、行った.保存菌株から切り出した寒天のブロッ ク (3x
3x
3m
m
)
を接種し 20'"'-'220C
,暗黒下で培養した. 二核交雑株の作出 ダイ・モン交配に用いるこ核菌株は,和合性を示す二つの栄養要求性一核 株同士を交配し,作出した.PDA
平板培地の中央に4mm
離して二つの和 合性を示す二つの一核株を接種した.約 10日間, 250C
で培養後,PDA
平 板培地上のコロニーを検鏡し,クランプ結合を確認したものを,二核菌糸体 と見なした.このように二核化を確認した後,PDA
斜面培地に接種し,2
週間, 250C
で培養し,実験に使用するまで 50C
で保守字した. ダイ・モン交記におけるこ核化の検証 10-まず,栄養要求性マーカーを付与したー核菌糸体をP D A平板培地の中央 部分に接種して 250
C
, 5日間培養した.生長したー核菌糸体に二つの異なっ た栄養要求性マーカーをもっ二核菌糸体を接種し,ダイ・モン交配させた. 10日間塔養後,一核菌糸体側のコロニー外縁部から 10サンプルの菌糸体を 切り出し, P D A斜面培地に移植した.培養後,小片を最少培地 (MM)お よびMM~こアデニン (ade),ピリドキシン (pdx),ヒスチジン (his)をそれぞ れ 10mg/l添加した培地に接種し,培養した.一核菌糸体は生長のためにこ れらの物質を必要とすることから, ade-, pdx-, his-と分類した.M Mの組 成は, グノレコース 20 g/l, (NH4) zP04 1.5 g/,l KHzP04 0.46 g/,lKzHPO 4 1.0 g/,lMgSO 4・7Hz 0 O. 5 g/l,チアミン塩酸塩 120μg/l,精製寒天 15 g/lであり,殺菌前に pHを5.5に調整した.供試菌株がー核菌糸体の状態 ならば菌糸体の生長に ade,pdxもしくは hisを要求するが,菌糸体細胞中 に二種類の核が存在すれば栄養要求性を相補することによりM Mで生育可 能である. ダイ・モン交配による二核菌糸体からのー核性オイディア形成における核型 の決定 ダイ・モン交配により作出した新生二核菌糸体の核型はそれからのー核性 オイディア形成により生じる優位核と従罵核の核型の解析によって判定し た (Caoetal, 1999). ダイ・モン交配により作出したこ核菌糸体からのー 核性オイディアの分離はCaoetal.(2000) に従った.オイディアの培養は 250C
,暗黒牝で、行った.オイディア細胞は通常 3日後に発芽し,菌糸体コロ ニーを形成する.コロニーの直径が 2mm程度に生長した頃,平板培地から 1i 噌3 ム 箇圃阻分離し,その栄養要求性マーカーを利用して核型を決定するために
M M
およ びMM~こ三つの化学物質をそれぞれ添加した平板培地に移植した .MM~こは 生育せず,M M
にこれらの物質の一つを含むものでは生長することにより菌 糸体の核型を判断した. 二核菌糸体のー核菌糸体化の検証 親二核株のー核化を供試菌株の栄養要求性検定で検証した.異なる栄養要 求性マーカーを付与した二核菌糸体をペトリ皿の端から 6mm離して接種し 培養した.9'"'-' 12日間培養後,菌糸体コロニーの先端部分から接種部分に向 かつて 1 mm切り出し, P D A斜面培地に接種した.さらに,分離したー核 菌糸体の核型を,それぞれの栄養要求性マーカーによるM M
とM M
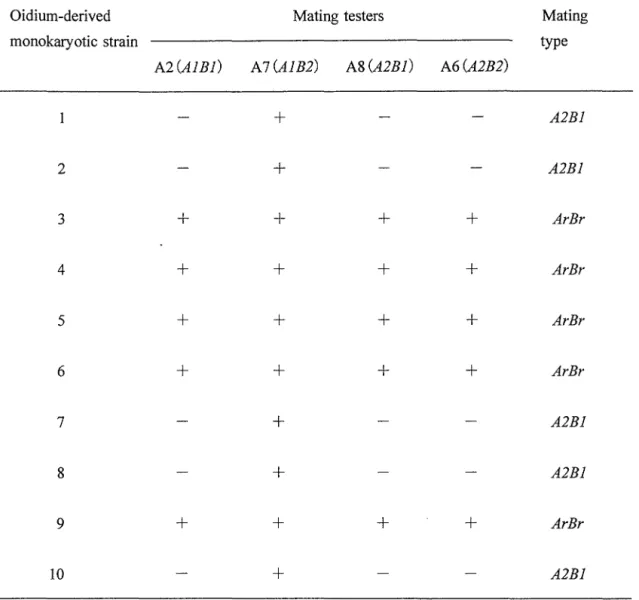
に各種栄 養素を添加した培地での生長テストで決定した. 結果 和合性ダイ・モン交配における核の選択 三 つ の 異 な る 栄 養 要 求 性 一 核 菌 糸 体 と 両 方 と も 異 な る 不 和 合 性 因 子 を 宥 する和合性組み合わせの栄養要求性二核菌糸体との需のダイ・モン交配結果 をTable 2-1に示す. 親 一 核 株 NX6-582 (A6/his-) と親二核株 NAl1-26(
A
l
/
ade-)+
NF8-267 (A5/pdx-)間での交配では,一核菌糸体側のコロニーから分離したすべて のサンプルでー核菌糸体の二核化が確認された.同様に和合性組み合わせの つ 山 11 ム 』・・・ーーーーー←ート 4 ι心 Table 2-1. Nuclear selection in compatible the di-mon matings between auxotrophic monokaryons and dikaryons of a bipolar mushroom , Pholiota nameko. Parental monokaryon Parental dikaryon Dikaryotization of parental monokaryon* NX6-582 (A6/his-) NAll-26 (Al/ade-) 十 NF8-267 (A5/pdx-) 10/10 NF8-267 (A5/pdx-) NAll-26 (Al/ade-)
+
NX6-582 (A6/his-) 10/10 NAll-26 (Al/ade-) NF8-267 (A5/pdx-)+
NX6-582 (A6/his-) 10/10 *Isolates were subcultured on PDA slants before their nuclei were examined. Nuclear type of Nuclear type of oidial monokaryon** dikaryon obtainedo
(his-) :96 (pdx-) NF8-267 + NX6-582 2 (his 一) :66 (pdx-) NX6-582 + NF8-267o
(ade-) :19 (pdx-) NF8-267 + NAll-26 **Nuclear types were identified by nutritional requirement tests described in Materials and Methods.圃圃圃圃圃
親 一 核 株
N
F
8
-
2
6
7
(A5/pdx-) と親二核株N
A
l1-
2
6
(Al/ade-)+NX6-582
(A6/his-)および親一核株
N
A
l
l
-
2
6
(Al/ade-)と親二核株N
F
8
-
2
6
7
(A5/pdx-)十
N
X
6
-
5
8
2
(A6/his-)の二つの異なる組み合わせにおいても分離したすべて のサンプルで、二核化が確認された.最初の和合性ダイ・モン交配の組み合わ せで二核化した菌糸体の核型を調査したところ,分離した9
6
のー核菌糸体 はすべてピリドキシン要求性を示した.この結果から,形成されたこ核菌糸 体は親ー核株N
X
6
-
5
8
2
(A6/his-) の核と二核親株N
F
8
-
2
6
7
(A5/pdx-) の核 とで構成されることが推測された.最初の組み合わせのダイ・モン交配の二 核化のプロセスにN
A
l
l
-
2
6
(Al/ade-) とN
F
8
-
2
6
7
(A5/pdx-) の間に核選択 の序列が介在した.次の組み合わせのダイ・モン交配では,二核化した菌糸 体の核型は6
6 (
p
d
x
-
)
:
2
(
h
i
s
-
)
の分離比であった.すなわち,形成され た二核株はN
X
6
-
5
8
2
(A6/his-) とN
F
8
-
2
6
7
(A5/pdx-)の二つの核によって 構 成 さ れ た . 三 番 目 の 組 み 合 わ せ で の ダ イ ・ モ ン 交 配 の 核 型 はN
A
l1-
2
6
(Al/ade-)の核と親二核株からのN
F
8
-
2
6
7
(A5/pdx-)の核とで構成される と推測した.以上の三つの結果から,ダイ・モン交配によるー核菌糸体の二 核化において親二核株の核の選択が関与することが示された.和合性ダイ・ モン交配に供試した親二核株を構成する三つの核問の核選択にはN
F
8
-
2
6
7
→N
X
6
-
5
8
2
とN
X
6
-
5
8
2
→N
A
l1-
2
6
の序列が推測された. 半和合性ダイ・モン交配における核の選択 三つの異なった栄養要求性一核菌糸体と一つの核型が親一核株と同じ不 和 合 性 因 子 を 構 成 す る 核 を 有 す る 栄 養 要 求 性 二 核 窟 糸 体 簡 の 半 和 合 性 ダ 14-イ・モン交配について調査した結果を Table 2-2に示す. 親一核株 NAll-26 (Al/ade-) と親二核株 NAll-26 (Al/ade-) 十NF8-267 (A5/pdx-)とのダイ・モン交配では親一核株側から分離したすべてのサン プノレで二核化が確認された. し か し な が ら , 一 方 の 親 一 核 株 NF8-267 (A5/pdx-)と閉じ親二核株とのダイ・モン交配では二核化は確認できなか った.また,親一核株 NAll-26 (Al/ade-) と親二核株 NAll-26 (Al/ade-) 十NX6-582 (A6/his-) とのダイ・モン交配では 10サンプルとも二核化が確 認されたが, NX6-582 (A6/his-) と親二核株 NAll-26(Al/ade-)十NX6-582 (A6/his-) とのダイ・モン交配の場合は 5サンプルで、しか二核化が認めら れなかった.親一核株 NF8-267 (A5/pdx-) と親二核株 NF8-267 (A5/pdx-) 十NX6-582 (A6/hi s-) とのダイ・モン交配ではすべてのサンプルに二核化が 認められたが,親一核株 NF8-267(A5/pdx-)側から分離したサンプノレは 30% しか二核化していなかった.これらの結果から,核選択の序列において相補 核が優位核の場合も従属核の場合でも二核化のための供与核となり得た.し かし,親二核株中の相補核が核選択の序列において従属核の場合,親二核株 の優位核と比較して二核化の確率は低いものとなった.この点から考察する と,半和合性ダイ・モン交配の場合,核の選択において 3つのこ核菌糸体を 構成するこつの核聞には NF8-267→NX6-582とNX6-582→NAll-26の序列があ ると推測された. 栄養要求性を付与した二核菌糸体の一核化における核選択の規則性 三つの二核菌株 NAll-26十NF8-267,NAl1-26十NX6-582,NF8-267十NX6-582 ζ J 1 i
-
・
・
ー
Table 2-2. Nuclear selection in hemicompatible di-mon mating between auxotrophic monokaryons and dikaryons with one nucleus of a similar to the incompatibility factor of the parental monokaryon. Parental monokaryon Parental dikaryon Dikaryotization of parental monokaryon* 10/10 0/10 10/10 5/10 3/10 10/10 * Isolates were subcultured on PDA slants before their nuclei were examined. Nuclear type of dikaryon obtained NF8-267 + NAll-26 NX6-582 + NAll-26 NX6-582 + NAll-26 NX6-582 + NF8-267 NX6-582 + NF8-267
のコロニー先端部から 1 mm内 側 の 菌 糸 体 小 片 を 切 り 出 し て ー 核 化 を 調 査 した結果を
T
a
b
1
e2
-
3
に示したが,一核化の割合はそれぞれ48%
,5%
,83%
であった.また,三つの二核菌糸体からー核化した菌糸体の核型はそれぞれN
F
8
-
2
6
7
,N
X
6
-
5
8
2
,N
F
8
-
2
6
7
であった.これらの結果から,二核麗糸体の 一核化において,三つの核の比較優位性の序列をN
F
8
-
2
6
7
→N
X
6
-
5
8
2
お よ びN
X
6
-
5
8
2
→N
A
l
l
-
2
6
と決定した. 考察 多くの研究者が四極性きのこの和合性ダイ・モン交配において親二核株の 二 つ の 核 の う ち の 一 つ の 核 が 期 待 値 以 上 の 頻 度 で 二 核 化 に 関 わ る こ と を 見 い 出 し た が , こ の 核 選 択 の 規 則 性 に 関 す る 報 告 は 見 当 た ら な い ( 木 村 ,1
9
6
6
,1
9
7
5
;
O
i
k
a
w
a
,1
9
3
9
;
P
a
p
a
z
i
a
n
,1
9
5
0
;
Q
u
i
n
t
a
n
i
1
h
a
,1
9
3
9
)
.
二極 性きのこである P.namekoの和合性ダイ・モン交配では,親二核株の二つの 核のうちのひとつの核が選択され,親一核株に移動し,二核化が完遂すると 推測された.和合性ダイ・モン交配に供試した三つの核における比較優位性 の序列はN
F
8
-
2
6
7
→N
X
6
-
5
8
2
およびN
X
6
-
5
8
2
→N
A
l
l
-
2
6
と決定した.この序列 は同菌の二核麗糸体のー核化の序列(
M
a
s
u
d
ae
t
a
1. ,1
9
9
5
)
およびー核性オ イディア形成の序列(
C
a
oe
t
a
1
.
,1
9
9
9
)
と一致した. 二極性きのこである P.namekoの半和合性ダイ・モン交配では,親二核株 の 相 補 核 が 二 核 菌 糸 体 の 一 核 化 に お け る 核 選 択 に つ い て 優 位 核 で も 従 罵 核 の場合でも二核化の供与核となり得た.しかし,親二核株の相補核が核選択 ウ ー 吋l よ-Table 2-3. Nuclear types of monokaryotized isolates in mycelial monokaryotization from dikaryons in di-mon matlng. 臼 ikaryon Number of monokaryons/total isolated from the margins of dikaryotic colonies* fム o shkp e ホ pas vdn 十uo rη aa eLK品 、EEム、,J cnu uo Nm NAll-26 (Al/ade-) 十 NF8-267 (A5/pdx-) 23/48
o
(NAll-26) :23 (NF8-267) NAll-26 (Al/ ade-)+
NX6-582 (A6/hi s-) 2/44o
(NAll-26) :2(NX6-582) ト 4 αコ NF8-267 (A5/pdx-)+
NX6-582 (A6/his-) 40/48 40 (NF8-267) :0 (NX6-582) *The number of monokaryons from mycelial isolates dissected into l-mm sections starting from the farthest edge of the dikaryotic mycelial colonies. **Nuclear types were identified by the nutritional requirement tests described in Materials and Methods.の序列において二核菌糸体の他方の核よりも優位性を示す場合,親ー核株側 から分離したすべてのサンプノレにおいて二核化が認められた.親二核株の棺 補核が核選択の序列が従属性を示す場合,親一核株側から分離したサンプノレ の二核化の割合は 0-50%にすぎなかった. Masudaらは本菌のいくつかのこ 核蕗株で菌糸体の一核化において出現する核型に偏りが生じることを報告 している (Masuda et al., 1995).本研究において供試した二核菌株におい ても優位核よりも従属核が出現する割合は大変低いという同様な核選択の 偏りが確認された.半和合性ダイ・モン交配において見られた従属核の関与 による見かけ上低頻度を示した二核化の例は,従属核が移動することによる ごく稀に生じる二核菌糸細胞がそれにつづく核分裂と核の移動により親一 核株のコロニーの二核細胞領域に拡大したものと推測された. 本菌では一般的に二核菌糸体の生長中に二核菌糸の一核化が生じる.菌糸 体のー核化は興味深く,担子菌の二核締胞中での共役核分裂における不安定 要因となり,供役核分裂のメカニズムを解析するいとぐちとなりうる.先に 二核菌糸体の細胞分裂の際,最初に分裂する核(ー核化細胞中で確認された 核)を主導核とし,他方の核を従属核とする Cascadeプロセスが介在すると する仮説を提起した (Masuda et al., 1995). ナメコのダイ・モン交配にお ける詳細はまだ解明されていないが,本研究で得られた知見ならびに既住の 知見に基づき,親一核株と親二核株のー核化菌糸から産出されるー核細胞と の交配の可能性が示唆された. Q J 1 tム
圃圃
後〉 幽 圃 圃 第 3 章 四 極 性 担 子 菌 き の こ
Flammulina v
θlutipes
の 和 合 性 お よ び 半 和 合 性 ダ イ ・ モ ン 交 配 に お け る 核 選 択 の 規 則 性 緒 言 思 極 性 き の こCoprinuslagopus
に お け る 二 核 菌 糸 体 に よ る ー 核 菌 糸 体 の 二 核 化 は , Bulier (1930,1931) に よ り 発 見 さ れ た . そ の 後 , 同 様 の 二 核 化 現 象 が 多 数 報 告 さ れ て い る(Dicson, 1934; Quintanilha, 1937, 1939; Oikawa, 1939 ;
木村, 1957a, 1957b, 1958a, 1958b). 親 二 核 株 に よ る 親 ー 核 株 の 二 核 化 は ダ イ ・ モ ン 交 配 (Papazian, 1950), あ る い は Quintanilha (1937) に よ り 発 見 者 の 名 に ち な ん で ブ ラ ー 現 象 と 呼 ば れ て き た . 自 極 性 き の こ に お け る ブ ラ ー 現 象 に 関 連 し た 未 解 明 の 主 た る 課 題 は , (a) Legitimateな 組 み 合 わ せ の 状 態 で の ダ イ ・ モ ン 交 配 ( 和 合 性 お よ び 半 和 合 性 ) で 親 二 核 株 の 二 核 の う ち の ー 核 の 選 択 が 生 じ る の か どうか, (b) 不 和 合 性 組 み 合 わ せ の ダ イ ・ モ ン 交 配 の 際 に 和 合 性 の 交 配 型 因 子 が 体 細 胞 組 換 え に よ り 新 生 す る の か ど う か で あ る (Raper, 1966). 我 々 は 二 極 性 き の こ で あ る
P
.nameko
の 和 合 性 ダ イ ・ モ - 20一ン 交 配 に お い て 親 二 核 株 の ー 核 が 選 択 さ れ , そ の 優 位 核 が 親 ー 核 株 へ 移 動 し 二 核 化 す る と 推 測 し た (Nogami et al., 2002) . こ の ダ イ ・ モ ン 交 配 に 供 試 し た こ 核 菌 糸 体 株 を 構 成 す る 2 核 開 で の 比 較 優 位 性 の 序 列 が 示 さ れ , そ の 結 果 は 本 菌 に お け る こ 核 菌 糸 体 (Masuda et al., 1995) お よ び オ イ デ ィ ア (Caoet al., 1999) の ー 核 化 の 序 列 と 一 致 し た . 半 和 合 性 ダ イ ・ モ ン 交 配 の 場 合 , 相 補 性 を 示 す 親 二 核 株 の 核 が 二 核 化 時 の 供 与 体 と な り 得 た . し か し , 親 一 核 株 と 相 補 す る 核 が 他 の 二 核 株 の 核 よ り 序 列 の 低 い 場 合 は , 親 - 核 株 側 コ ロ ニ ー か ら 採 取 し た サ ン プ ル の 二 核 化 は 0%'"'-'50% で あ っ た . さ ら に , 和 合 性 ダ イ ・ モ ン 交 配 に 見 ら れ る 同 一 の 核 選 択 の 規 則 が 半 和 合 性 ダ イ ・ モ ン 交 配 に お い て も 機 能 し て い る と 推 測 さ れ た . 本 菌 に お け る い わ ゆ る ダ イ ・ モ ン 交 配 は , 親 一 核 株 と 親 二 核 株 の ー 核 化 菌 糸 体 か ら 産 出 さ れ た 一 核 細 胞 と の 交 配 に よ る こ と が 示 唆 さ れ た . こ れ ら の 結 果 が 四 極 性 き の こ で も 適 用 さ れ る か ど う か , 比 較 検 証 す る 必 要 が あ ろ う . と こ ろ で , き の こ に は 二 核 菌 糸 体 か ら 有 性 生 殖 過 程 を 経 ず に ー 核 化 さ れ る こ と に 関 し て , 二 種 類 の ー 核 化 現 象 が 報 告 さ れ て い る . 最 初 の 事 例 は ,
P
.
nameko (
有田, 1964;Arita, 1979) Masuda et al., 1995) で 報 告 さ れ て い る 二 核 菌 糸 体 コ ロ ニ ー の 外 縁 部 で 脱 二 核 化 が 生 じ , 一 核 菌 糸 体 が 形 成 11 ム 円 〆 山-・
ι
--・-←
さ れ る も の で あ る . こ の よ う な 二 核 菌 糸 体 の ー 核 化 は
Ashan ( 1952) お よ び Aschan-Aberg( 1960) より
Collybia
(=Flammu
1
ina) velutipes
で も 報 告 さ れ て い る . も う 一つ 事 例 は ,
Coprinus cinereus
(Rao et al, 1969),Favolus
arcularius
(Kitamoto, unpublished data),F
.
velutipes
(Brodie, 1936; Takemaru, 1954; Kitamoto et al., 2000),
P
.
nam
θko
(Cao et al., 1999 Arita, 1979) な ど の き のこ で , 二 核 菌 糸 体 の オ イ デ ィ ア 形 成 を 介 し た 一 核 化 で あ る .
F
.
velu tipes
で は , 一 核 性 オ イ デ ィ ア の 核 型 分 析 に よ り オ イ デ ィ ア の 核 構 成 が 同 定 さ れ , 二 核 菌 糸 体 の オ イ デ ィ ア 形 成 を 介 し た ー 核 化 に お け て 核 の 選 択 が 一 定 の 序 列 に よ っ て 行 わ れ る こ と が 示 さ れ て い る (Kitamoto et al., 2000). 本 研 究 で は , 四 極 性 き の こ ,F
.
velutipes
の ダ イ ・ モ ン こ う は い に お け る 上 述 の 諸 課 題 の 究 明 を 目 的 に , 不 和 合 性 因 子 の 異 な る 栄 養 要 求 性 突 然 変 異 株 を 用 い て 和 合 性 お よ び 半 和 合 性 ダ イ ・ モ ン 交 配 で の 親 二 核 株 に お け る 核 選 択 の 規 則 を 解 析 し た . ま た , 四 極 性 き の こ に お け る 和 合 性 お よ び 半 和 合 性 ダ イ ・ モ ン 交 配 を 用 い た 二 核 菌 糸 体 の ー 核 化 に お い て 2核 う ち の 1核 が 選 択 さ れ る か ど う か を 検 討 し た . さ ら に Legitimateな ダ イ ・ モ ン 交 配 の プ ロ セ ス に こ 核 菌 糸 体 の ー 核 化 が 介 在 す る か ど う か に に つ い て も 検 討 し た . -22-材 料 お よ び 方 法
供 試 菌 株
本 研 究 に は ,F. velu tipθ sの ー 核 株 A2,A6, D4を 紫 外
線 す る こ と に よ り 得 ら れ た 栄 養 要 求 性 突 然 変 異 株 A2・10
(A1Bl/pab-), A6・9 (A2B2/met-), D4・2 (A4B4/ade-)
を 用 い た . 菌 糸 体 は 12 ml の PDA 培 地 (Nissui Pharmaceutical
Co.
Ltd.) を 分 注 し た プ ラ ス チ ッ ク 製 ペ ト リ 皿 (Iwaki,90X15 mm) で 培 養 し た . 保 存 菌 株 か ら 菌 糸 体 小 片 (3X3X3mm) を 切 り 出 し , 250 C暗 所 で 培 養 し た . 二 核 交 雑 株 の 作 製 ダ イ ・ モ ン 交 配 に 用 い る 親 二 核 株 は 和 合 性 の 栄 養 要 求 性 一 核 株 二 つ を 交 配 し て 作 製 し た . PDA平 板 培 地 の 中 央 に 4 m m離 し て 二 つ の ー 核 株 を 接 種 し た . 250 C, 10日 間 の 培 養後, PDA平 板 培 地 上 の コ ロ ニ ー を 検 鏡 し , ク ラ ン プ 結 合 を 調 査 す る こ と に よ り 二 核 菌 糸 体 を 確 認 し た . 確 認 し た 二 核 菌 糸 体 を PDA斜 面 培 地 に 移 植 し , 250 Cで 2週 間 培 養 後 , 50 Cで 保 存 し た . ダ イ ・ モ ン 交 配 に お け る こ 核 化 の 検 証 栄 養 要 求 性 マ ー カ ー を 付 与 し た ー 核 菌 糸 体 を PDA平 板 培 っ d つ 山.
圃
-
-』』 地 の 中 央 に 接 種 し ,
25
0C
で ー 逓 間 培 養 し た . 生 長 し た ー 核 菌 糸 体 の コ ロ ニ ー 屑 縁 部 に 二 核 と も 栄 養 要 求 性 マ ー カ ー を 有 す る こ 核 菌 糸 株 を 接 種 し た . 1週 間 培 養 後 , ー 核 菌 糸 体 側 の コ ロ ニ ー の 外 縁 部 か ら 約10 m m
内 側 の 菌 糸 体 小 片 を10
値 切 り 出 し , PDA斜 面 培 地 に 移 植 し た . 生 長 し た 菌 糸 体 を 最 少 培 地(MM)
とM M
に10 mg
/ l の ア デ ニ ン , メ チ オ ニ ン , も し く は p-ア ミ ノ 安 患 香 酸 を 加 え た 平 板 培 地 に 接 種 し , 菌 糸 体 の 栄 養 要 求 性 を 試 験 し た . こ れ ら の 栄 養 素 を 必 要 と す る ー 核 菌 糸 体 を そ れ ぞ れade¥met-
,pab-
と 分 類 し た .M M
の 組 成 は20g
/lglucose
, 1.5
g
/l(NH
4 ) 2HPO
4 '0.46
g
/lK H
2P0
4, 1.0 g
/lK
2HP0
4,0.5 g
/lMgS0
4 ・7H
20
,120μg
/ l チ ア ミ ン 塩 酸 塩 ,15 g
/ l 精 製 寒 天 で あ り , 殺 菌 前 に pH
を5.5
に 調 整 し た . 分 離 培 養 さ れ た ー 核 菌 糸 体 はade
幽met-
, あ る い はpab
- で あ る が , 二 種 類 の 核 を 持 つ 二核 細 胞 は 栄 養 要 求 の 相 補 性 に よ り
M M
で 生 長 可 能 で あ る . ダ イ ・ モ ン 交 配 に よ る 一 核 蕗 糸 体 の 二 核 化 は , 各 試 験 区 毎 に10
個 の サ ン プ ノ レ を 用 い て 検 証 し た . ダ イ ・ モ ン 交 配 に よ っ て 新 生 し た こ 核 菌 糸 体 の 核 型 の 決 定 ダ イ ・ モ ン 交 配 に よ っ て 新 生 し た こ 核 菌 糸 体 の 核 型 は , そ れ か ら 作 出 さ れ た ー 核 性 オ イ デ ィ ア の 核 型 の 分 析 か ら 同 定 し た(Kitamoto e
t
a
l.,2000).
二 核 菌 糸 体 か ら の ー 核 -24-性 オ イ デ ィ ア の 分 離 は , Kita盟 oto et al. (2000) ら の 方 法 に 従 っ た . オ イ デ ィ ア は 250 C, 暗 黒 下 で 培 養 し た . オ イ デ ィ ア の 細 胞 は 通 常 3 日 後 に 発 芽 し , 菌 糸 体 コ ロ ニ ー を 形 成 す る . コ ロ ニ ー の 直 径 が 2 m m程 度 に 伸 長 し た 後 , 平 板 培 地 か ら 移 植 し , 栄 養 要 求 性 を 検 定 す る た め に , M Mお よ び M Mに 一 つ の 化 学 化 合 物 を 加 え た こ つ の 平 板 培 地 に 接 種 し た . 分 離 培 養 さ れ た 菌 糸 体 の 核 型 は , ま ず M Mで は 生 育 し な い こ と を 確 認 し , つ ぎ に 閉 じ 化 合 物 を 加 え た M Mで の 生 育 テ ス ト に よ り 検 証 し た . 二 核 菌 糸 体 に お け る 菌 糸 体 の ー 核 化 の 検 証 ダ イ ・ モ ン 交 配 で 作 出 し た こ 核 菌 糸 体 コ ロ ニ ー の 周 縁 部 に お け る の 菌 糸 体 の ー 核 化 の 成 否 を 栄 養 要 求 性 試 験 で 検 証 し た . 二 つ の 栄 養 要 求 性 マ ー カ ー を 付 与 し た 二 核 菌 糸 体 を ペ ト リ 皿 の 端 か ら 6 mm離 し て 接 種 し 培 養 し た . 9'"'-' 12 日 間 培 養 後 , 菌 糸 体 コ ロ ニ ー の 先 端 部 分 か ら 接 種 部 分 に 向 か つ て 1 mm切 り 出 し , フ ッ ク 細 胞 の 有 無 を 検 鏡 し た . さ ら に 分 離 し た 一 核 菌 糸 体 の 核 型 を , そ れ ぞ れ の 栄 養 要 求 性 マ ー カ ー に よ る M Mと M Mに 各 種 栄 養 素 を 添 加 し た 培 地 で の 生 長 テ ス ト で 決 定 し た . 結 果 z i u q L -・・ー
-和 合 性 ダ イ ・ モ ン 交 配 に お け る 核 の 選 択 異 な る 栄 養 要 求 性 マ ー カ ー を 付 与 し た 三 つ の ー 核 菌 糸 体 と 両 方 と も 異 な る 不 和 合 性 因 子 を 有 す る 栄 養 要 求 性 二 核 醤 糸 体 の 和 合 性 ダ イ ・ モ ン 交 配 の 結 果 を Table 3・1に 示 す .
A6
幅9
(A2B2/met-)
とA2
・10
(A1Bl/pab-)
+D4
・2
(
A4B4/ade
幽 ) の 交 配 に つ い て10
組 の 試 験 を 行 っ た 結 果 , す べ て の 交 配 試 験 で 親 一 核 株 の 二 核 化 が 確 認 さ れ た . 同 様に , 親 一 核 株
A2-10
(A1Bl/pab-)
と 親 二 核 株A6-9
(A2B2/met-)
十D4
・2
(A4B4/ade-)
お よ び 親 一 核 株D4
・2
(
A4B4/ade-)
と 親 二 核 株A2
・10
(A1Bl/pab-)
+A6
桐9
(A2B2/met-)
の 二 つ の 組 み 合 わ せ で で も す べ て の サ ン プ ル で 試 験 二 核 化 が 認 め ら れ た . 最 初 の 組 み 合 わ せ の 和 合 性 ダ イ ・ モ ン 交 配 に よ り 得 ら れ た 新 生 二 核 菌 糸 体 の ー 核 性 オ イ デ ィ ア の 核 型 は pab幽株が 53株 分 離 さ れ た が met-株 は わ ず か 4 株 分 離 さ れ た の み で あ っ た . こ れ は , 親 二 核 株 か らA2-10
(A1Bl/pab-)
の 核 の み が 親 - 核 株 側 に 移 動 し た こ と を 示 す 結 果 で あ る . し た が っ て , 最 初 の 和 合 性 ダ イ ・ モ ン 交 配 の 二 核 化 の 過 程 に , 親 二 核 株 中 のD4
・2
(A4B4/ade-)
の 核 よ り もA2
・10(A1Bl/pab-)
の 核 が 選 択 的 に 移 動 す る 機 能 が 介 在 す る こ と が 示 唆 さ れ た .2
番 目 の 組 み 合 わ せ で は , 新 生 二 核 菌 糸 体 か ら の ー 核 性 オ イ デ ィ ア の 核 型 の 割 合 は 38 f o つ ム ... ー←N h 斗 Table 3-1. Nuclear selection in compatible the di-mon matings between auxotrophic monokaryons and dikaryons of a tetrapolar mushroom , Flammulina v , θ lutip θS. Parental monokaryon Parental dikaryon Dikaryotization of Nuclear type of Nuclear type of parental monokaryon* oidial monokaryon 料 dikaryon obtained A6-9 (A2B2/met-) A2-10 (A1Bl/pab-)+D4-2 (A4B4/ade-) 10/10 4 (met-) :53 (pab-) A2-10+A6-9 A2-10 (A1Bl/pab ー) A6-9 (A2B2/met-)+D4-2 (A4B4/ade 一) 10/10 22 (ade-) :38 (pab-) D4-2+A2-10 D4-2 (A4B4/ade-) A2-10 (A1Bl/pab-)+A6-9 白 2B2/met-) 10/10 25 (ade 一) : 32 (pab-) A2-10+D4-2 *Isolates were subcultured on PDA slants before their nuclei were examined. **Nuclear types were identified by nutritional requirement tests described in Materials and Methods.
pab
-株で22
株 がade
- 株 で あ っ た . こ の 結 果 か ら ,A2
・10
(A1Bllpab-)
とD4-2
(A4B4/ade-)
の 核 が 新 生 二 核 菌 糸 体 に 内 在 し , そ れ ぞ れ の 核 は 親 一 核 株 と 親 二 核 株 由 来 の 核 で あ る こ と が 判 明 し た . 3番 自 の 組 み 合 わ せ で 得 ら れ た こ 核 菌 糸 体 の 核 はD4
・2(A4B4/ade-)
の 核 と 親 二 核 株 の A2・10
(A1Bllpab-)
の 核 で 構 成 さ れ て い た . こ れ ら の 結 果 は , す べ て の 組 み 合 わ せ に お い て , 和 合 性 ダ イ ・ モ ン 交 配 に よ る 一 核 蔭 糸 体 の 二 核 化 に は 親 二 核 株 か ら 移 動 す る 核 の 選 択 性 が 介 在 す る こ と を 示 し て い る . 供 試 し た 3 株 の 親 二 核 株 を 構 成 す る 三 つ の 核 が 供 与 核 と な る 場 合 に 生 じ る 核 選 択 の 序 列は, A2 -1 0→ D4・2お よ び D4・2→ A6・9で あ っ た . 半 和 合 性 ダ イ ・ モ ン 交 配 に お け る 核 の 選 択 異 な る 栄 養 要 求 性 マ ー カ ー を 付 与 し た 三 つ の ー 核 菌 糸 体 と , 親 二 核 株 の 一 つ は 同 一 の 不 和 合 性 国 子 を 有 す る 核 を , も う 一 つ は 相 補 性 の 不 和 合 性 因 子 を 有 す る 核 を も っ 栄 養 要 求 性 を 有 す る こ 核 菌 糸 体 の 間 で 半 和 合 性 ダ イ ・ モ ン 交 配 の 成 否 を 検 討 し た . そ の 結 果 をTable 3-2
に 示 す . 親 一 核 株D4
・2(A4B4/ade
同 ) と 親 二 核 株 A2・10
(A1Bllpab-)
+D4・2(A4B4/ade-)
を 交 配 さ せ た と こ ろ ,10
株 す べ て の サ ン プ ル で 二 核 化 が 生 じ た . し か し , 親 一 核 株A2-10(A1Bllpab-)
に 閉 じ 親 二 核 株 を 交 配 さ せ た と こ ろ - 28-rtv にο Table 3-2. Nuclear selection in hemicompatible di-mon mating between auxotrophic monokaryons and dikaryons with one nucleus of a similar to the incompatibility factor of the parental monokaryon in Flammulina velutipes. Parental monokaryon Parental dikaryon A2-10(AIBl/pab-) A2-10 (A1B1/pab-) +D4-2 (A4B4/ade-) D4-2 (A4B4/ade-) A6-9 (A2B2/met-) A6-9 (A2B2/met-)+D4-2 (A4B4/ade-) D4-2 (A4B4/ade-) A2-10(AIBl/pab-) A2-10 (AIBl/pab-) +A6-9 (A2B2/met-) A6-9 (A2B2/met-) Dikaryotization of parental monokaryon* 4/10 10/10 10/10 5/10 4/10 10/10 *Isolates were subcultured on PDA slants before their nuclei were examined. Nuclear type of dikaryon obtained
... ー← 二 核 化 の 証 拠 と な る M Mに 生 育 し た の は わ ず か 40%で あ っ た . 親 二 核 株
A6
・9 (A2B2Imet-)+D4-2
((A4B4Iade-)と 親 ー 核 株A6
・9 (A2B2Imet-) の 組 み 合 わ せ の 交 配 で は , す べ て の サ ン プ ル で 二 核 化 が 生 じ た . 一 方 , 親 一 核 株D4
・2
(A4B4Iade-) と の 交 配 で は , 全 サ ン プ ノ レ の 50%で 二 核 化 が 観 察 さ れ た . 親 二 核 株 A2-10 (A1Bllpab-)+
A6
・9
(A2B2Imet-) と 親 ー 核 株A2
・10
(A1Bllpab幽 ) の 組 み 合 わ せ の 交 配 で は , 40%の サ ン プ ル が 二 核 化 し た の み で あ っ た が , 同 じ 親 二 核 株 と 親 一 核 株 A6-9 (A2B2Imet-) の 組 み 合 わ せ の 交 配 で は , す べ て の サ ン プ ル が 二 核 化 し た . し た が っ て , 本 菌 で は , 半 和 合 性 ダ イ ・ モ ン 交 配 に お い て , 例 え 親 二 核 株 の 2つ の 核 が 核 選 択 の 序 列 に お い て 優 位 あ る い は 劣 位 で あ っ て も , 親 一 核 株 と 相 補 性 を 示 す 親 二 核 株 の 核 が 二 核 化 の 供 与 核 と な り う る こ と が 確 認 さ れ た . し か し , 親 一 核 株 と 相 補 す る 核 が 核 選 択 の 序 列 で 従 属 核 の 場 合 は , 親 一 核 株 の 優 位 核 と 比 較 し て 二 核 化 の 頻 度 は 低 率 で あ っ た . ま た , 半 和 合 性 ダ イ ・ モ ン 交 配 に お け る 核 選 択 の 序 列 は 和 合 性 ダ イ ・ モ ン 交 配 の 序 列 と 同 じ で あ る と 推 測 し た . 栄 養 要 求 性 を 付 与 し た 二 核 菌 糸 体 の 一 核 化 に お け る 核 選 択 の 規 則 性Collybia (=Flamm ulina) vθlu tipesは 生 長 の 周 縁 部 で 二
30-核 菌 糸 体 か ら オ イ デ ィ ア 形 成 を 経 由 す る こ と な し に 一 30-核 菌 糸 細 胞 を 産 生 す る こ と が 知 ら れ て い る (Ashan, 1952 Aschan-Aberg, 1960). こ れ は , 本 四 極 性 菌 に お い て , 和 合 性 お よ び 不 和 合 性 ダ イ ・ モ ン 交 配 に お け る 親 二 核 菌 糸 体 の ー 核 化 の プ ロ セ ス が ダ イ ・ モ ン 交 配 に 介 在 す る 可 能 性 を 示 唆 す る も の で あ る . そ こ で , 親 二 核 株 と し て 用 い た 三 つ の 栄 養 要 求 性 二 核 菌 糸 体 コ ロ ニ ー の 周 縁 部 か ら 分 離 し た サ ン プ ル の 一 核 化 を 検 討 し た . そ の 結 果 を Table3・3に 示 す . 3株 の 二 核 菌 糸 体 の コ ロ ニ ー 先 端 部 か ら 1 m mの 小 片 を 切 り 出 し , 一 核 菌 糸 体 の 割 合 を 検 査 し た と こ ろ , そ れ ぞ れ , A2・10 (A1Bllpab-) +D4・2 (A4B4/ade胴)では 13%,A6幽 9
( A2B2Imet-) +D4・2 (A4B4/ade-) で は 5
%
,
A2-10(A1Bllpab-) +A6・9 (A2B2Imet-) で は 10%の 頻 度 で ー 核
菌 糸 体 が 検 出 さ れ た . こ の 結 果 か ら , こ れ ら の こ 核 菌 糸 体 の コ ロ ニ 一 周 縁 部 の 菌 糸 先 端 細 胞 で 、 は 一 核 住 が 生 じ て い る も の と 推 測 し た . 3株 の 二 核 菌 糸 体 か ら ー 核 化 し た 菌 糸 体 の 核 型 は , そ れ ぞ れ A2・10(A1Bllpab-), D4・2(A4B4/ade -), A2・10 (A1Bllpab-) で あ っ た . 菌 糸 体 の ー 核 化 に お け る こ れ ら の 核 の 比 較 優 位 性 の 序 列 を 総 括 す る と , A2・10
(A1Bllpab-) → D4・2 (A4B4/ade-), D4・2 (A4B4/ade-)
→ A6・9 (A2B2Imet-) と な っ た . す な わ ち , こ の 序 列 は 和 合 性 お よ び 半 和 合 性 ダ イ ・ モ ン 交 配 の 核 選 択 の 序 列 と 一 致 円 、 υ
-
-
-
-w N Table 3-3. Nuclear types of monokaryotized isolates in mycelial monokaryotization from dikaryons in di-mon mating in Flammulina velutip θS. Oikaryon A2-10 (AIBl/pab-) +04-2 (λ 4B4/ade-) A6-9(A2B2/met-)+04-2(A4B4/ade-) A2-10 (AIBl/pab-)+A6-9 (A2B2/met-) Number of monokaryons/total isolated from the margins of dikaryotic colonies* 5/40 2/40 4/40 Nuclear types of monokaryons** 5 (A2-10) :0(04-2)
o
(A6-9) :2(04-2) 4 (A2-10) :0 (A6-9) *The number of monokaryons from mycelial isolates dissected into l-mm sections starting from the farthest edge of the dikaryotic mycelial colonies. **Nuclear types were identified by the nutritional requirement tests described in Materials and Methods.ー
ー
ー
ー
←
ー
す る も の で あ っ た . 考 察 四 極 性 き の こ の ダ イ ・ モ ン 交 配 は , 親 一 核 株 と 親 二 核 株 間 の そ れ ぞ れ が 有 す る Aお よ び B交 自 己 型 因 子 の 組 み 合 わ せ に よ り , 和 合 性 , 半 和 合 性 不 和 合 性 お よ び 不 和 合 性 ダ イ モ ン 交 配 の 3 種 類 の カ テ ゴ リ ー に 分 類 さ れ る . 本 章 で はFlammu
1
ina
velutipesの legitimateな 和 合 性 お よ び 半 和 合 性 不 和 合 性 ダ イ ・ モ ン 交 配 に つ い て 論 じ , 不 和 合 性 ダ イ モ ン 交 配 に つ い て は 第 5章 で 論 じ た い . 四 極 性 き の こ の 和 合 性 ダ イ ・ モ ン 交 配 に つ い て は , 一 核 菌 糸 体 が ニ 核 化 す る プ ロ セ ス で , 偶 然 生 じ る と 期 待 さ れ る 以 上 の 高 い 頻 度 で 親 二 核 菌 糸 体 の 二 つ の 核 の う ち の ど ち ら か が 親 一 核 菌 糸 体 に 移 動 す る こ と が , 多 く の 研 究 に お い て 報 告 さ れ て い る (Quintanilha, 1937, 1939 Papazian, 1950 ;木村, 1958a, 1966, 1975). し か し な が ら , 著 者 は ,F
.
velutipes
の 和 合 性 ダ イ ・ モ ン 交 配 に お い て , 親 二 核 株 の 2核 の ー 核 の み が 選 択 さ れ , 親 一 核 株 へ 移 動 し て い く こ と で 二 核 化 が 完 成 す る こ と を 栄 養 要 求 性 突 然 変 異 株 を 用 い て 検 証 し た . 二 極 性 き の こ で あ るP
. nameko
の 和 合 性 ダ イ ・ モ ン 交 配 に お い て も 同 様 な 結 果 が 得 ら れ て い る . ま た , -33--
-
ー
F
.
velutipes
の 和 合 性 ダ イ ・ モ ン 交 配 に お い て , 供 試 二 核 菌 糸 体 を 構 成 す る 3 種 の 核 の 間 で , こ の よ う な 核 の 選 択 に 関 して, A2・10→ D4欄 2お よ び D4-2→ A6・9 と い う 比 較 優 位 性 の 序 列 が 介 在 す る こ と が 示 さ れ た . さ ら に , こ の 序 列 は , 本 菌 の 二 核 菌 糸 体 の ー 核 化 の 序 列 と 一 致 す る こ と が 示 さ れ た. 西 極 性 き の こ の 半 和 合 性 ダ イ ・ モ ン 交 配 に お い て , Buller ( 1931) はCoprinuslagopus
に お い て , 親 二 核 株 に 存 在 す る 相 補 核 が 親 一 核 株 へ 移 動 し , 二 核 化 が 完 遂 さ れ る と 推 測 し た . し か し , Dickson( 1934,1936), Quintanilha( 1939) お よ び 木 村 (1958b ) ら は , そ れ ぞ れCoprinus
spha
θrosporus
お よ びCoprinus
macrorhizus f
.
microsporus(= C
.
cinereus)
に お い て , 半 和 合 性 ダ イ ・ モ ン 交 配 で は 親 二 核 株 の 両 方 の 核 が 一 核 株 に 移 動 し , 新 し い こ 核 菌 糸 株 が 形 成 さ れ た こ と を 示 唆 す る 報 告 を 行 っ て い る . 著 者 はF
. velutipes
に お い て , 親 二 核 株 中 の 親 一 核 株 に 対 し て 相 補 性 の 核 の み が こ 核 化 の 供 与 核 と な り う る こ と を 示 し た . そ の 際 , 親 二 核 株 の 相 補 核 が 菌 糸 体 の ー 核 化 に お い て 示 さ れ た 序 列 で 優 位 核 あ る い は 従 属 核 の い ず れ で あ っ て も 親 一 核 株 の 二 核 化 の 完 遂 が 可 能 で あ っ た . し か し , 相 補 核 が 二 核 菌 糸 体 の 他 核 よ り 序 列 が 低 い 場 合 は , 親 一 核 株 恨 ! J か ら 分 離 し た サ ン プ ル の 40'"'-'50%し か 二 核 化 し な て い な -34-か っ た . こ の 結 果 は , 二 核 茜 糸 体 中 の 劣 位 核 が 誤 っ て 選 択 さ れ る こ と が 低 頻 度 で は あ る が 生 じ る た め , 相 補 核 が 劣 位 核 で あ っ て も ダ イ ・ モ ン 交 配 が 完 遂 す る も の と 仮 定 さ れ る . 相 補 核 が 劣 位 核 で あ っ て , 極 低 頻 度 し か ダ イ ・ モ ン 交 配 に よ る 二 核 菌 糸 体 の 形 成 が 生 じ な い 場 合 で も , そ れ に つ づ い て 惹 起 さ れ る 共 役 核 分 裂 と コ ロ ニ ー に お け る 核 の 移 動 に よ り 新 生 二 核 菌 糸 体 の 領 域 が 拡 大 し , ダ イ ・ モ ン 交 配 で 生 じ る こ 核 化 が 生 じ る 確 率 よ り も 大 き く 増 幅 さ れ , そ の 結 果 , 見 か け 上 , 優 位 核 の 場 合 と そ れ ほ ど 劣 ら な い 二 核 株 作 出 率 と な っ た と 推 測 さ れ た . ま た , 半 和 合 ダ イ ・ モ ン 交 配 に お け る 核 選 択 の 序 列 は 和 合 性 の 場 合 と 同 ー で あ り , 和 合 性 ダ イ ・ モ ン 交 配 で 起 き て い る 間 じ 核 選 択 の プ ロ セ ス が 宇 和 合 ダ イ ・ モ ン 交 配 で も 介 在 す る も の と 推 測 さ れ た . 二 極 性 き の こ
P
.
namθko
お よ び 四 極 性 き の こF
. velutipes
に お い て は , ニ 核 菌 糸 体 コ ロ ニ ー 屑 縁 部 で の ー 核 菌 糸 細 胞 の 形 成 が 報 告 さ れ て い る ( 有 田 1964, 1979; Masuda et al., 1995; Ashan, 1952; Aschan-Aberg, 1960). オ イ デ ィ ア 形 成 に お け る こ 核 細 胞 の ー 核 化 もP
. nameko
(Arita,1964, 1979 Kitamoto et al., 2000) お よ び
F
. velu tipes
で 報 告 が あ る (Caoet al., 1999; Kitamoto et al. , 2000). 著 者 ら の 研 究 グ ル ー プ は , 二 核 繭 糸 体 の 細 胞 分 裂 に お い て , ま ず 最 初 に 分 裂 す る 核 が 主 導 核 と し て 機 能 し , も う 一 方 の F h u q 司 U ... ーー←
核 は 従 属 核 と し て 機 能 す る 共 役 核 分 裂 が “
Cascade"
し て 起 こ る と い う 仮 説 を 先 に 提 起 し て い る (M a
s
u
da
e
ta
1
.
,1
99 5
)
.
二 核 菌 糸 体 の 一 核 化 は 共 役 核 分 裂 に お け る 不 安 定 さ を 示 す も の で あ り , 担 子 菌 に お け る 共 役 核 分 裂 の 解 析 の 糸 口 と な り う る 興 味 深 い 現 象 で あ る . ダ イ ・ モ ン 交 配 の 様 相 はF
.
Vθlu tipes
に お い て も 詳 細 の 解 明 に 至 っ て い な い が , 親 一 核 株 と 親 二 核 株 か ら の ー 核 化 菌 糸 と の 関 の 交 配 は , 実 際 上 , か な り の 頻 度 で 起 こ り う る と 推 測 さ れ た . 続 F む つ d-
-
-第 4 章
四 極 性 担 子 菌 Lentinulaedodesお よ び P1 euro tus os trea tus
の 和 合 性 お よ び 半 和 合 性 ダ イ ・ モ ン 交 配 に お け る 核 選 択 の 規 制
性
緒 言
著 者 は , 第 2 章 お よ び 第 3 章 に お い て , 二 核 菌 糸 体 の コ
ロ ニ 一 周 縁 部 で 一 核 化 菌 糸 体 が 生 じ る こ 極 性 お よ び 四 極 性
き の こ Pholiota namekoお よ び Flammulina velutip θ Sに
お け る 和 合 性 お よ び 半 和 合 性 ダ イ ・ モ ン 交 配 を 行 い , そ れ
ぞ れ の き の こ に お い て 親 二 核 株 か ら ー 核 化 し た 菌 糸 細 胞 と
親 ー 核 菌 糸 体 の 間 で 交 配 が 起 こ り う る ケ ー ス を 指 摘 し た .
こ れ ら の き の こ は , ど ち ら も 一 核 性 オ イ デ ィ ア を 形 成 す る
能 力 が 報 告 さ れ て い る ( 有 田 1979; Cao et a1., 1999;
Brodie, 1936; 五itamoto et a1. 2000) . し か し な が ら ,
き の こ に お け る オ イ デ ィ ア 形 成 は 担 子 菌 の ご く 一 部 と 考 え
ら れ , 例 え ば ,Lθntinula θdodesお よ び Pleurotusostreatus
で は , オ イ デ ィ ア 形 成 は 観 察 さ れ て い な い し , ま た , 二 核 菌 糸 体 の 一 核 化 も 報 告 が な い . そ こ で , 本 章 で は , こ れ ら の 四 掻 性 担 子 菌 を 供 試 材 料 と ヴ i つ d
-
ー
ー
-幽幽~ し て , 親 一 核 株 と 親 二 核 株 を 用 い て 真 性 と 考 え 得 る 和 合 性 お よ び 半 和 合 性 ダ イ ・ モ ン 交 配 を 行 い , 第 2章 お よ び 第 3 章 と 同 様 に し て ダ イ ・ モ ン 交 配 に お け る ー 核 菌 糸 体 の 二 核 化 に お け る 核 選 択 の 規 則 性 に つ い て 検 討 し た . 材 料 お よ び 方 法 供 試 菌 株 本 研 究 に は L.edodesの 二 核 株
X-1
お よ び #110
株 をu
v
照 射 し て 作 出 し た 栄 養 要 求 性 変 異 株 X-1-248(A4B3/his-),X-1-449(A3B3/pab-), X-1-650(A3B4/ile-),X-1-667(A4B3/
ade-)
,X-1-829
(A4B4/met-),#
110-666
(A1B2/ade-),#
110-822
(A1B2/met-),#
110-872
(A1Bl/trp-)を 用 い た . ま た ,P. ostreatusの 栄 養 要 求 性 変 異 株 と し て ,
TD33-100
(A2B2/nic
一, TD33-118(A1B2/ade-),)τ
D33-138(A2Bl/met-),TD33-336(A1B2/met-)を 用 い た . 栄 養 要 求 性 突 然 変 異 株 の 作 出 は
衣 笠 お よ び 北 本 ( 1
99
7 ) の 方 法 に 従 っ た . 菌 糸 体 の 培 養 は12
ml
の P D A培 地(NissuiPharmaceutical
Co.Ltd.)
を 分 注 した プ ラ ス チ ッ ク 製 の ペ ト リ 皿
(Iwak
,i90
x
15 mm)
で 行 っ た .保 存 茜 株 か ら 切 り 出 し た 寒 天 の ブ ロ ッ ク
(
3
x
3
x
3 mm)
を 接種 し
20-22
0C
, 暗 黒 下 で 培 養 し た .
38--.幽』ー 二 核 交 雑 株 の 作 製 ダ イ ・ モ ン 交 配 に 用 い る 二 核 菌 株 は , 和 合 性 を 示 す 二 つ の 栄 養 要 求 性 一 核 株 同 士 を 交 配 し て 作 出 し た . P D A平 板 培 地 の 中 央 に
4
m離 し て 二 つ の 和 合 性 を 示 す 二 つ の 一 核 株 を 接 種 し た . 約 10日間, 250 Cで 培 養 後 , P D A平 板 培 地 上 の コ ロ ニ ー を 検 鏡 し , ク ラ ン プ 結 合 を 確 認 し た も の を , 一 核 菌 糸 体 と み な し た . こ の よ う に 二 核 化 を 確 認 し た 後 P D A斜 面 培 地 に 接 種 し 2週間, 250 Cで 培 養 し 50 Cで 保 存 し た . ダ イ ・ モ ン 交 配 に お け る こ 核 化 の 検 証 ま ず , 栄 養 要 求 性 マ ー カ ー を 付 与 し た - 核 菌 糸 体 を P D A平 板 培 地 の 中 央 部 分 に 接 種 し て 250 C, 5日 間 培 養 し た . 生 長 し た ー 核 菌 糸 体 コ ロ ニ ー の 周 縁 部 の ー カ 所 に 二 つ の 異 な っ た 栄 養 要 求 性 マ ー カ ー を も っ 二 核 菌 糸 体 を 接 種 し , ダ イ ・ モ ン 交 配 さ せ た . 10日 間 培 養 後 , 接 種 点 と は 反 対 側 で あ る - 核 菌 糸 体 の コ ロ ニ ー 外 縁 部 か ら 10サ ン プ ル の 菌 糸 体 を 切 り 出 し , P D A斜 面 培 地 に 移 植 し た . 培 養 後 , 小 片 を 最 少 培 地 ( MM ) お よ び M Mに ア デ ニ ン , ヒ ス チ ジ ン , ニ コ チ ン 酸 ア ミ ド , メ チ オ ニ ン , ト リ プ ト フ ァ ン p-ア ミ ノ 安 息 香 酸 あ る い は イ ソ ロ イ シ ン を そ れ ぞ れ 10 mg/l添 加 し た 培 地 を 用 い て 培 養 し た . 一 核 菌 糸 体 は 生 長 の た め こ れ ら の-39--ーーーー一←ー
物 質 を 必 要 と す る こ と か ら ,
ade-
,his
一,n
i
c
,一met
,一tr
p-
,pab-
お よ びile
ー と 同 定 し た.MM
の 組 成 は , グ ル コ ー ス20
g/
,l (刊H4)2P04
1.5 g/
,l 五百2P04 0.46 g/1
,K2
註P04
1.0 g/l
,MgS04
・7H20 0.5 g/l
, チ ア ミ ン 塩 酸 塩120μ
g/
l , 精 製 寒 天15g
/1
で あ り , 殺 菌 前 にpH
を5.5
に 調 整 し た . 供 試 菌 株 が ー 核 菌 糸 体 の 状 態 な ら ば 菌 糸 体 の 生 長 に 上 記 の 物 震 を 要 求 す る が , 菌 糸 体 細 胞 中 に 二 種 類 の 核 が 存 在 す れ ば 栄 養 要 求 性 を 相 補 す る こ と に よ りM M
で 生 育 可 能 で あ る こ と か ら ,M M
で の 生 育 の 可 否 に よ り 二 核 化 の 検 証 を 行 っ た . な お , 親 一 核 株 の 二 核 化 は , 検 鏡 に よ っ て ク ラ ン プ の 存 在 を 確 認 す る 方 法 を 併 用 し た . 結 果 和 合 性 ダ イ ・ モ ン 交 配 の 二 核 化 の 確 認 L.θdodesの 親 二 核 株 と 異 な る 不 和 合 性 因 子 を 有 す る 親 一 核 株 と の 間 で ダ イ ・ モ ン 交 配 を 行 い , 親 一 核 株 側 の 二 核 化 に つ い て 調 べ た 結 果 を ,Table 4-1
に 示 す L.edodes の 親 二 核 株 X-1-248(A4B3/his一)+
X-1-650 (A3B4/ile-)
と 親 一 核 株 #110-822(A1B2/met-)
, 親 二 核 株X-1-650(A3B4/ile-)
+
X-1-667(A4B3/ade
) と 親 一 核 株 #110-872 (A1Bl/trp-)
, お よ び 親 二 核 株X-1-449
(A3B3/pab-)+
X-1-829
(A4B4/met-)と -40-広 h ト 4 Table 4-1. The results of compatible di-mon mating between auxotrophic monokaryons and dikaryons in Lentinula edodes. Parental dikaryon Parental monokaryon X-1-248 (A4B3/his-) +X-1-650 (A3B4/ile-) 詳 110-822 (AIB2/met-) X-1-650 (A3B4/ile-) +X-1-667 (A4B3/ade-) 枠 110-872 (AIBl/trp-) X-1-449 (A3B3/pab-) +X-1-829 (A4B4/met-) れ 10-666 (AIB2/ade-) Rate of dikaryotization of parental monokaryon 10/10 10/10 10/10