はじめに
感染症は病原微生物の侵入に起因する疾患であ り,ヒトなどの生物に限らず,すべての多細胞生 物において重要な疾患である.多細胞生物は常に 病原微生物の侵入にさらされており,病原微生物 を排除し続けなければ生きてゆくことができな い.そのためヒトに限らず,昆虫などの無脊椎動 物,植物などにおいても感染防御機構が備わって いる.これらの生物における感染防御機構の,分 子レベルでの解明が進んできた結果,病原微生物 を認識し,感染防御反応を誘導する分子が昆虫や 植物で同定されてきた.ハエの Toll はその代表的 な分子である.興味深いことに,Toll に類似する 分子を我々ヒトも持っていることが明らかにな り,我々ヒトから無脊椎動物まで共通する病原体 認識機構,感染防御機構の存在が明らかになった.
それがヒトにおける自然免疫機構であり,中心的 役割をなす認識分子が Toll-like receptor(TLR)で ある.本稿では,TLR についての概説を試みると ともに,感染症にどこまで関わっているかこれま で明らかにされた結果を紹介する.
1.ハエ病原体認識分子Toll
Toll-like receptor(TLR)の発見は昆虫,特にハ エにおける感染防御機構の解析が大きく貢献して いる.TLR は Toll のヒトホモローグとして発見 された1).Toll は,ハエの個体発生の際に背腹軸を 決定するシグナルを伝達する分子として同定され たが,その後 Toll 遺伝子に変異を持つハエが真菌 感 染 に 高 い 感 受 性 を 持 つ こ と が 明 ら か に な っ た2).その原因は,真菌が感染しても感染防御反応 を誘導することができないことに起因する.つま り Toll は真菌の侵入を察知し,感染防御反応を誘 導する分子であることが明らかにされた.Toll は I 型の膜タンパクで,真菌の侵入をロイシン
リッチリピートと呼ばれるタンパクモチーフで構 築された細胞外ドメインで察知し,細胞内ドメイ ンから活性化シグナルを伝達する(図 1).そのシ グナルは抗菌ペプチドの産生を誘導し,真菌の排 除を促す.植物においても同様な病原体認識分子 が報告されている3).シグナル伝達機構は Toll や TLR とは異なっているが,興味深いことに病原体 認識に関わる部位は Toll や TLR と同様にロイシ ンリッチリピートで構築されている.
2.自然免疫病原体認識分子Toll-like receptor(TLR)
Toll に類似したヒトの分子とし て Medzhitov らが 1997 年に TLR4 をク ロ ー ニ ン グ し た1).そ の後現在まで 10 個の TLR がクローニングされ,
フ ァ ミ リ ー を 形 成 し て い る 事 が 明 ら か に な っ た4).ハエの Toll と同様に TLR も病原体を認識 する(図 2).グラム陰性菌由来の膜成分である LPS(lipopolysaccharide)を TLR4 とその細胞外 ドメインに会合する MD-2 が5)〜8),マイコプラズ マや細菌由来リポペプチド,グラム陽性菌のペプ チ ド グ リ カ ン を TLR1!TLR2 や TLR6!TLR2 な どの 2 量体が9)10),鞭毛由来タンパクであるフラ ジェリンを TLR5 が11),細菌由来 DNA を TLR9 が12),それぞれ認識する.ウィルス由来の成分も TLR によって認識される.2 重鎖 RNA を TLR3 が13),TLR7 は抗ウイルス薬として使われている 薬剤に反応することから,類似したウィルス由来 の構造物を認識している可能性が示唆されてい る14).このように TLR ファミリーは細菌ばかり でなく,真菌やウィルスも含めた多様な病原体を それぞれの TLR が特異的に認識し,活性化シグ ナルを伝達する.
3.TLRのシグナル伝達機構
TLR は I 型の膜タンパクで,シグナルを伝達す
Toll-like receptor と感染防御
東京大学医科学研究所感染遺伝学
三 宅 健 介
総 説
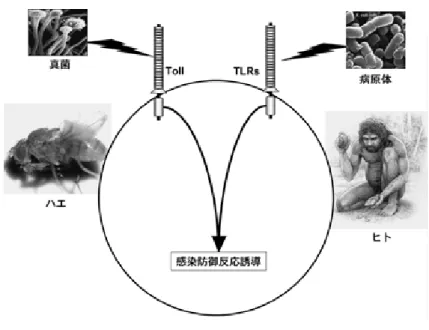
図 1 ハエとヒトに共通する病原体認識分子 Toll と Toll-like receptor ハエとヒトの病原体認識分子 Toll と TLR を示す. ハエは Toll が真菌感染を察知し,
活性化シグナルを伝達し,抗菌ペプチド産生などの防御反応が誘導される.ヒトにお いては Toll-like receptor(TLR)が病原体成分を認識して同様な活性化シグナルを伝達 し,獲得免疫も含めた免疫応答を誘導する.
図 2 Toll-like receptor の構造と特異性
ヒトは 10 個の TLR を持っているがそのうち 9 個まで特異性が決定されている.その 細胞外ドメインはロイシンリッチリピートと呼ばれるタンパクモチーフで,細胞内ド メインは IL-1 レセプターのそれと類似した TIR(Toll-Interleukin 1 receptor)ドメイ ンから構築される.TLR2 は TLR1 や TLR6 と 2 量体を形成して,ペプチドグリカンや リポペプチドに反応する.TLR4 は細胞外ドメインで MD-2 と会合してはじめて,LPS
(Lipopolysaccharide)を認識できる.ほかは本文参照.
る細胞内ドメインは I 型 IL-1 レセプターに類似 しており,TIR(Toll-Interleukin 1 Receptor)ドメ インとよばれる15).TLR の下流でシグナルを伝 達するために必須のアダプター分子が MyD88 で ある(図 3).この分子の KO マウスはほとんどの TLR に お い て シ グ ナ ル 伝 達 に 障 害 が あ る16). MyD88 の下流は,IRAK(IL-1 receptor associated kinase),TRAF6(tumor necrosis factor receptor- associated factor 6)を介して最終的に NF-kB お よび MAP キナーゼ(Mitogen Activated Protein kinases)活性化が誘導される.最近,TIRAP(Toll- interleukin 1 receptor(TIR)domain-containing adapter protein)が MyD88 と協調して TLR2 や TLR4 のシグナルを伝達していることが明らかに
なった17)18).ほかの TLR については TIRAP のよ う な TIR ド メ イ ン を 持 つ ア ダ プ タ ー 分 子 が MyD88 と強調してシグナルを伝達している可能 性が示唆されている.MyD88 非依存性の経路も TLR4 や TLR3 で確認されている.MyD88 非依 存 性 の 経 路 で は TRIF(TIR domain-containing adapter inducing IFN-beta )!TICAM-1 ( Toll- interleukin 1 receptor domain(TIR)-containing adaptor molecule)を 通 し て IRF-3(Interferon Regulatory Factor-3)が活性化され,I 型インター フェロン産生が誘導される(図 3)19)20).I 型イン ターフェロンが抗ウィルス応答などを誘導する.
このように今後は,個々の TLR 間のシグナルの 違いを説明しうる分子基盤の解明に進みつつあ る.病原微生物によって誘導される免疫応答が異 なることが知られているが,その違いを TLR か らのシグナルの違いで説明しうるかもしれない.
4.TLRによって誘導される病原体排除機構
TLR によって誘導される病原体排除機構にお いてもハエとヒトの類似性は存在する.Toll や TLR を介するシグナルは抗菌ペプチドの産生を 誘導する.抗菌ペプチドはアミノ酸 30 個ほどから なるシステインが豊富なペプチドで,陰性に荷電 した菌体膜に作用し,膜に穴をあけたり,菌体内 に入ってプロテアーゼの作用を抑制したりして抗 菌作用を示す21).ヒトの細胞膜の外側は陰性には 荷電していないので,抗菌ペプチドは作用しない.
抗菌ペプチドは病原体侵入の場である消化管や呼 吸器など粘膜組織において産生される.たとえば 腸管を例にとると,パネート細胞における産生が 報告されているが22),その産生の制御に TLR が 関与していると考えられている.また免疫系の細 胞では好中球が多く産生する.最も早く対応する 防御機構において抗菌ペプチドが重要な役割をし ている.
5.獲得免疫活性化におけるTLRの役割 ヒトとハエの免疫機構における違いは,リンパ 球を中心とした獲得免疫にある.ハエが自然免疫 だけで感染を防御するのに対して,我々ヒトは自 然免疫と,リンパ球を中心とした獲得免疫とを融 合させた免疫システムを築きあげてきた.獲得免 図 3 TLR のシグナル伝達経路
TLR4-MD-2 を LPS が刺激すると,TIR ドメインから 活性化シグナルが伝達される.その直下で機能する分 子が MyD88 や TIRAP ( Toll-interleukin 1 receptor
(TIR)domain-containing adapter protein)で あ る.
MyD88 の 下 流 で,IRAK(IL-1 receptor associated kinase),TRAF6(Tumor necrosis factor receptor as- sociated factor 6)が機能し,最終的に NF-kB,MAPK
(Mitogen activated protein kinase)の活性化が誘導さ れ,防御反応が誘導される.MyD88 非依存性の経路も TLR4 や TLR3 で確認されている.MyD88 非依存性の 経 路 で は TRIF(TIR domain-containing adapter in- ducing IFN-beta)!TICAM-1(Toll-interleukin 1 recep- tor domain(TIR)-containing adaptor molecule)を通し て IRF-3(Interferon Regulatory Factor-3)が活性化さ れ,I 型インターフェロン産生が誘導される.
疫活性化は,細胞レベルでは樹状細胞にゆだねら れる23).樹状細胞は骨髄由来のマクロファージ系 の細胞で,広範な組織に分布する.病原体が侵入 すると樹状細胞は成熟,活性化されるとともに所 属リンパ節に遊走し,リンパ節に遊走してきた T 細胞を活性化する(図 4).樹状細胞は抗原に感作 されたことのない T 細胞を活性化することので きる唯一の抗原提示細胞であり,T 細胞活性化を 介して細胞性免疫,体液性免疫の誘導を制御して いる.TLR は樹状細胞においても発現されてお り,そのシグナルはサイトカインの産生,CD80!86 など補助刺激分子の発現誘導を通じて T 細胞活 性化を促す1).獲得免疫はタンパク抗原を認識す る機構であるが,タンパクそれ自体はあまり免疫 原性がない.タンパクに対する免疫応答を誘導す るために我々は免疫賦活剤アジュバントを用い る.最も強いアジュバントは結核菌などの病原体 成分である.病原体の成分が樹状細胞の TLR を 活性化することで,サイトカインの産生や補助刺
激分子の発現が誘導され,タンパク抗原に対する 獲得免疫応答が誘導される.TLR はいってみれば アジュバントのレセプターである.TLR は自然免 疫の病原体排除機構を誘導するばかりでなく,獲 得免疫応答の誘導にも深く関与している.つまり 生体に侵入した病原体に対し,獲得免疫も含めた すべての免疫応答を誘導するかどうかの決定に深 く関わる分子であると考えられる.
6.Th1!Th2誘導とTLR
樹状細胞は,獲得免疫発動に重要であるばかり でなく,誘導する獲得免疫の種類にも影響を与え る(図 4).獲得免疫はγインターフェロン産生に よって誘導される Th1 型と IL-4,IL-6,IL-9,IL-13 などのサイトカインによる Th2 型に分けられる.
前者はマクロファージの活性化や IgG2a 抗体産 生が特徴で,後者は IgE や IgG1 抗体産生を特徴 とする.興味深いことに病原体の種類によって誘 導される獲得免疫応答が異なる.病原体細菌,特 に細胞内で増殖する細菌の場合にはγインター 図 4 獲得免疫活性化における TLR の役割
獲得免疫の活性化に重要な役割を果たしている樹状細胞も TLR を発現している.病 原体由来のタンパクは貪食され,組織適合抗原(Major histocompatibility complex,
MHC)と複合体の形で,T 細胞レセプター(T cell receptor,TCR)によって認識さ れる.一方病原体成分が TLR によって認識され,活性化シグナルが伝達される.その シグナルは,CD80 や CD86 などの補助刺激分子の発現を誘導し,T 細胞の CD28 を刺 激する.抗原刺激と補助刺激の両方がそろって始めて T 細胞は活性化され,Th1 か Th2 に分化し,獲得免疫応答が誘導される.
フェロンによるマクロファージの活性化が重要で あるが,ある種の寄生虫やウィルスの場合には IgE などの抗体産生が優位となる.どちらの応答 が優位に誘導されるかは,樹状細胞によって大き く影響される.Huang らは大腸菌,真菌,インフ ルエンザウィルスで樹状細胞を刺激し,誘導され る遺伝子を,DNA チップを用いて調べた.全部で 6,800 程度の遺伝子を調べているが,すべての病原 体に共通して誘導される遺伝子もあればそれぞれ の 病 原 体 に 特 異 的 に 誘 導 さ れ る 遺 伝 子 も あ っ た24).つまり病原体の種類によって樹状細胞の活 性化が異なることになる.病原体が獲得免疫に及 ぼす影響の少なくとも一部は,樹状細胞上の TLR からのシグナルを介している事が予想される.実 際に MyD88 の欠損マウスでは Th1 への誘導に 著しい障害が認められるが,Th2 への誘導には影 響がないことが報告されている25).TLR は一般 に Th1 への誘導に寄与するのかもしれない.今 後,TLR 間のシグナル伝達経路の違いが明らかに なるにつれて,その違いと Th1!Th2 誘導との関 係が明らかにされることが期待される.
7.TLRと感染防御
TLR は上皮細胞など最前線の臓器において,抗 菌ペプチド産生などを介して防御反応の誘導を促 す.また上皮細胞やマクロファージに発現した TLR はサイトカイン産生を誘導することで,好中 球を中心とした迅速な防御反応誘導を促す.さら に樹状細胞に発現する TLR は,樹状細胞を活性 化し,T 細胞活性化を通じて獲得免疫応答を誘導 する.このように侵入した病原体を TLR が認識 し,活性化シグナルを伝達することは,その病原 体を生体全体として排除するかどうかの決定に直 結しており,いうなれば TLR は防御反応を誘導 するかどうかの決定に大きく関与する分子である とも言える.実際に TLR が欠損すると,感染症に 高い感受性を示す例が報告されている.感染防御 という点では,TLR の中でも TLR4 と TLR2 が特 に重要な役割を果たしている.TLR2 は TLR1 や TLR6 とヘテロ 2 量体を形成してグラム陰性菌の ペプチドグリカンやリポペプチドを認識する(図 2)10).一方 TLR4 はその細胞外ドメインで MD-2
という分子と会合し,TLR4-MD-2 がグラム陰性 菌の外膜を構成する脂質であるエンドトキシン
(LPS,Lipopolysaccharide)を認識する8).TLR2 や TLR4 が病原体を認識できないと,その病原体 に対する免疫応答が著しく低下する.たとえば TLR4 と MD-2 のどちらの分子が欠損してもグラ ム陰性菌感染に対して高い感受性を示すことがマ ウスにおいて報告されている9)26).TLR4 の場合 はヒトにおいても TLR4 の Mutation(299 番目の アスパラギンがグリシン,あるいは 399 番目のス レオニンがイソロイシン)が ICU(Intensive Care Unit)でのグラム陰性菌感染の患者に多い傾向が あることが報告されている27)28).CD14 にはその ような相関は認められていない.一方 TLR2 を欠 損したマウスはグラム陽性菌感染に高い感受性を 示す29).ヒトにおいても TLR2 遺伝子の変異が報 告されており,その変異とグラム陽性菌感染との 関連が疑われている30).TLR4 や TLR2 について はほかの感染症との関連も報告されている31)〜33). これら TLR に変異のある場合には対応する病原 体の認識に問題があり,防御反応誘導が低下して いることが高い感受性の原因であると予想され る.
おわりに
自然免疫機構は特異的な病原体認識から病原体 排除までのすべてを備えており,それ自体で完結 したシステムである.実際ハエなどの種では自然 免疫機構だけで,感染防御機構を構築している.
ヒトは自然免疫の上にリンパ球を中心とした獲得 免疫を加えたシステムを組み上げてきた.TLR は自然免疫に属するというよりも,自然免疫,獲 得免疫を含めた免疫システムの制御に深く関与す る分子といえる.したがって病原微生物による感 染症の感受性に大きく影響する分子であることは 容易に想像できる.
文 献
1)Medzhitov R, Preston-Hurlburt P, Janeway CA Jr:A human homologue of the Drosophila Toll protein signals activation of adaptive immunity.
Nature 1997;388:394―7.
2)Lemaitre B, Nicolas E, Michaut L, Reichhart JM, Hoffmann JA:The dorsoventral regulatory gene
cassette spatzle!Toll!cactus controls the potent antifungal response in Drosophila adults . Cell 1996;86:973―83.
3)Dixon MS, Jones DA, Keddie JS , Thomas CM , Harrison K, Jones JD:The tomato Cf-2 disease resistance locus comprises two functional genes encoding leucine-rich repeat proteins. Cell 1996;
84:451―9.
4)Medzhitov R, Janeway C Jr:Innate immunity. N Engl J Med 2000;343:338―44.
5)Hoshino K, Takeuchi O, Kawai T, Sanjo H, Ogawa T, Takeda Y,et al.:Cutting edge:Toll-like re- ceptor 4(TLR4)-deficient mice are hyporespon- sive to lipopolysaccharide:evidence for TLR4 as the Lps gene product. J Immunol 1999 ; 162 : 3749―52.
6)Qureshi ST, Lariviere L, Leveque G, Clermont S, Moore KJ, Gros P,et al.:Endotoxin-tolerant mice have mutations in Toll-like receptor 4(Tlr4).J Exp Med 1999;189:615―25.
7)Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X,et al.:Defective LPS signaling in C3H! HeJ and C57BL!10ScCr mice:mutations in Tlr4 gene. Science 1998;282:2085―8.
8)Shimazu R, Akashi S, Ogata H, Nagai Y, Fuku- dome K, Miyake K,et al.:MD-2, a molecule that confers lipopolysaccharide responsiveness on Toll-like receptor 4. J Exp Med 1999 ; 189 : 1777―82.
9) Takeuchi O , Hoshino K , Kawai T , Sanjo H , Takada H, Ogawa T,et al.:Differential roles of TLR2 and TLR4 in recognition of gram-negative and gram-positive bacterial cell wall components.
Immunity 1999;11:443―51.
10)Ozinsky A, Underhill DM, Fontenot JD, Hajjar AM, Smith KD, Wilson CB,et al.:The repertoire for pattern recognition of pathogens by the in- nate immune system is defined by cooperation between toll-like receptors . Proc Natl Acad Sci U S A 2000;97:13766―71.
11)Hayashi F, Smith KD, Ozinsky A, Hawn TR, Yi EC, Goodlett DR,et al.:The innate immune re- sponse to bacterial flagellin is mediated by Toll- like receptor 5. Nature 2001;410:1099―103.
12)Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H,et al.:A Toll-like receptor recognizes bacterial DNA. Nature 2000;408:740―5.
13) Alexopoulou L , Holt AC , Medzhitov R , Flavell RA : Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3.
Nature 2001;413:732―8.
14)Hemmi H, Kaisho T, Takeuchi O, Sato S, Sanjo H,
Hoshino K,et al.:Small anti-viral compounds ac- tivate immune cells via the TLR 7 MyD 88- dependent signaling pathway . Nat Immunol 2002;3:196―200.
15)O Neill LA:Signal transduction pathways acti- vated by the IL-1 receptor!toll-like receptor su- perfamily. Curr Top Microbiol Immunol 2002 ; 270:47―61.
16)Kawai T, Adachi O, Ogawa T, Takeda K, Akira S:Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity 1999;11:115―22.
17)Yamamoto M, Sato S, Hemmi H , Sanjo H , Ue- matsu S, Kaisho T,et al.:Essential role for TI- RAP in activation of the signalling cascade shared by TLR2 and TLR4. Nature 2002;420:
324―9.
18)Horng T, Barton GM, Flavell RA, Medzhitov R:
The adaptor molecule TIRAP provides signalling specificity for Toll-like receptors. Nature 2002;
420:329―33.
19)Oshiumi H, Matsumoto M, Funami K, Akazawa T, Seya T:TICAM-1, an adaptor molecule that participates in Toll-like receptor 3-mediated interferon-beta induction . Nat Immunol 2003 ; 4:161―7.
20) Yamamoto M , Sato S , Mori K , Hoshino K , Takeuchi O, Takeda K,et al.:Cutting edge:a novel Toll!IL-1 receptor domain-containing adapter that preferentially activates the IFN-beta promoter in the Toll-like receptor signaling. J Im- munol 2002;169:6668―72.
21) Ganz T : Defensins and host defense . Science 1999;286:420―1.
22) Ayabe T , Wulff H , Darmoul D , Cahalan MD , Chandy KG, Ouellette AJ:Modulation of mouse Paneth cell alpha-defensin secretion by mIKCa1, a Ca2+-activated, intermediate conductance po- tassium channel. J Biol Chem 2002;277:3793―
800.
23)Mellman I, Steinman RM:Dendritic cells:spe- cialized and regulated antigen processing ma- chines. Cell 2001;106:255―8.
24)Huang Q, Liu D, Majewski P, Schulte LC, Korn JM, Young RA,et al.:The plasticity of dendritic cell responses to pathogens and their compo- nents. Science 2001;294:870―5.
25) Schnare M , Barton GM , Holt AC , Takeda K , Akira S, Medzhitov R,et al.:Toll-like receptors control activation of adaptive immune responses.
Nat Immunol 2001;2:947―50.
26) Nagai Y , Akashi S , Nagafuku M , Ogata M , Iwakura Y, Akira S,et al.:Essential role of MD-2
in LPS responsiveness and TLR 4 distribution . Nat Immunol 2002;3:667―72.
27)Agnese DM, Calvano JE, Hahm SJ , Coyle SM , Corbett SA, Calvano SE,et al.:Human toll-like receptor 4 mutations but not CD 14 polymor- phisms are associated with an increased risk of gram-negative infections . J Infect Dis 2002 ; 186:1522―5.
28)Lorenz E, Mira JP, Frees KL , Schwartz DA : Relevance of mutations in the TLR4 receptor in patients with gram-negative septic shock. Arch Intern Med 2002;162:1028―32.
29)Echchannaoui H, Frei K, Schnell C, Leib SL, Zim- merli W , Landmann R : Toll-like receptor 2- deficient mice are highly susceptible to Strepto- coccus pneumoniae meningitis because of re- duced bacterial clearing and enhanced inflamma- tion. J Infect Dis 2002;186:798―806.
30) Lorenz E , Mira JP , Cornish KL , Arbour NC ,
Schwartz DA:A novel polymorphism in the toll- like receptor 2 gene and its potential association with staphylococcal infection . Infect Immun 2000;68:6398―401.
31)Netea MG, Van Der Graaf CA, Vonk AG, Ver- schueren I, Van Der Meer JW, Kullberg BJ:The role of toll-like receptor(TLR)2 and TLR4 in the host defense against disseminated candidiasis. J Infect Dis 2002;185:1483―9.
32)Abel B, Thieblemont N, Quesniaux VJ, Brown N, Mpagi J, Miyake K,et al. :Toll-like receptor 4 ex- pression is required to control chronic Mycobac- terium tuberculosis infection in mice. J Immunol 2002;169:3155―62.
33)Kang TJ, Chae GT:Detection of Toll-like recep- tor 2(TLR2)mutation in the lepromatous leprosy patients. FEMS Immunol Med Microbiol 2001;
31:53―8.
Toll-like Receptors and Their Roles in Defense Responses Against Infection Kensuke MIYAKE
Division of Infectious Genetics, Department of Microbiology and Immunology, The Institute of Medical Science, The University of Tokyo
〔J.J.A. Inf. D. 77:473〜479, 2003〕