過去のいか流し網漁業データを用いた北太平洋の
アカイカ秋生れ群の資源評価
past driftnet fishery data
一井太郎(遠洋水産研究所)
・Kedarnath Mahapatra(東海大学海洋学部)
Tar
, Kedarnath MAHAPATRA
北太平洋のアカイカ資源は, 1970 年代初めに激減したスルメイカの漁獲量を補うために, いか釣り 漁船によって三陸・道東沖合で開発された. か流し網漁業によるアカイカの本格的な漁獲は 1978 に始まり, いか釣り漁業との競合を避けるため, 日本では 1979 年から東経 170 度以西を釣り漁場, 東を流し網漁場とする規制を行った. その後, いか釣り漁業は縮小したが, いか流し網漁業は 1980 代には毎年12∼22 万トンの漁獲を揚げ, 日本のいか類需要の1/3 を供給する重要な漁業に発展した. た, 韓国と台湾も参入し, いか流し網漁業によるアカイカの全漁獲量は, 1982 から1992 年にかけて 間17∼34 万トンに達した(図 1). 図1 北太平洋のアカイカの国別総漁獲量 1992 年以降の中国の釣り漁獲量は日本のそれと同程度と仮定(過小評価の可能性大) しかし, いか流し網漁業は, ウミガメ, イルカ類などを多く混獲し, これらの資源への悪影響が懸 されるとして、国連決議により1992 年末をもって停止(モラトリアム)となった(谷津, 1995). 一 , いか流し網漁業のアカイカ資源への影響については評価されなかった. アカイカは, 寿命が 1 年 , 北太平洋では発生時期の違いにより, 冬春生れ群と秋生れ群に分けられる(谷津ら, 1998; 長澤ら, 998). 両群とも亜熱帯海域を産卵場, 亜寒帯境界∼移行領域を索餌場とする季節回遊を行い, 後者 海域に漁場が形成される(図2)(村田,1990;村田・中村,1998;谷津,1992). 冬春生れ群の 餌域は日本沖からカナダ沖まで東西に連続するが, 秋生れ群のそれは天皇海山のある 170°E 以東 主に限られる. 従って, 170°E 以東で主に春∼秋に操業した日本のいか流し網漁業においては, 主 に秋生れ群が漁獲された.
Stock assessment of the autumn cohort of neon flying squid
(
Ommastrephes bartramii
) in the North Pacific based on the
o ICHII
い 年 以 年 ま 年 念 方 で 1 の 索 に 0 5 10 15 20 25 30 35 40 74 76 78 80 82 84 86 88 90 92 94 96 98 00 漁獲 量( 万ト ン ) 日本釣り 台湾流し網+釣り 韓国 流し網 日本流し網 中国釣り 年本研究では, 過 る秋生れ群の 資源量の推定を3 つの方法(面 デル)で行い,本漁業の秋生 れ群資源への影響評価を行った いか流し網漁業データ 月・旬毎,緯度 1 度 ×経度1 本漁業が始まった最初の数年間 ( 978-81 年)は 20°∼46°N, 170°E∼145°W と定められ けます類の混獲を最小限 に留める 月毎に定められた(図 ) 漁期は 月であった 流し網の目合は 主に 120mm であった(Yatsu et al., 1993). 韓国および台湾のいか流し網漁業データについては それぞれ ( )および d はなかった 韓国の漁期は ∼ 月 台湾の漁期は ∼ 月であった 流 網の目合は 韓国では 台湾では ∼ であった 漁獲物の外套長データ 日本のいか流し網船に乗船した科学オブザーバーにより アカイカ の外套長データが 年に収集された 法などでは 旬毎のアカイカ外套長( から 以下の式により体重( )に換算し 漁獲(資源)重量から漁獲(資源)尾数への変換を行った r2=0.9925) (一井, 未発表) 去のいか流し網漁業データに基づき, 日本の旧いか流し網漁場におけ 積-密度法, DeLury 法, プロダクションモ .
140°E 150°E 16 0°E 170°E 18 0° 170°W 160°W 150°W 140°W 130°W 120°W
60°N 60°N 秋生れ群の回遊 16 0° 18 120° 50°N 60°N 60°N 冬春生れ群の回遊 50°N 図2 アカイカ秋生れ群と冬春生れ群の回遊
試料と方法
日本のいか流し網漁業データについては, 1982~1992 年の 度毎に集計されたアカイカ漁獲量と努力量を用いた. なお, こうした詳細な漁獲データが収集されなかった. 日本の操業海域は140°E 150°E 16 0°E 170°E 18 0° 170°W 160°W 150°W 140°W 130°W 120°W
20°N 20°N 30°N 30°N 40°N 40°N 産卵場(冬∼春) 産卵場(冬∼春) 索餌場(夏∼冬) 1 , その北限はアカイカの漁獲を維持しつつさ ため, 3 . 6~12 . , 110∼
, Gong et al. 1993 Yeh an
Tung(1993)の月毎の漁獲量を用いた. 日本とは異なり, 両国の操業海域に 170°E 以東という規制 . 4 12 , 4 11 . し , 86 ∼115mm, 80 120mm . アカイカ , 1991 . DeLury , ML ;cm) , BW ;g , . BW = 0.0149 ML 3.1994 ( 20°N 30°N 40°N
140°E 150°E E 170°E 0° 170°W 160°W 150°W 140°W 130°W W
50°N 50°N 40°N 30°N 20°N 索餌場(春∼夏) 産卵場(秋) 産卵場(秋)
14 0°E 15 0°E 160°E 17 0°E 180° 170°W 16 0°W 150°W 14 0°W 13 0°W 120°W 60 °N 60°N 調査流し網データ 17 北海道大学(1979∼1999 年;Hokkaido Univer )が毎年7 月に行っ た流し網調査のアカイカ 調査で用いた流し網 の構成は, 北海道大学の場合は 48∼157mm 目 合13 種類であった(Y l.(2000)にならっ て, 93∼157mm 目合 10 反当 星画像および気候値による表面水温データ 表面水温とアカイカ商業漁獲との関係を調べるため に, NOAA 衛星による AVHRR 水温画像(1985 年∼92 年)と Reynolds の気候値(1982∼1984 年)
の 7 月における月・旬毎の表面水温(SST)データを用いた. いずれのデータもインターネット 図3 日本の流し網漁場 5°30’E 経度ライン(38°30’∼47°30’N)において, sity, 1980-2000)および遠洋水産研究所(2001∼2002 年 漁獲量, 努力量データおよび表面水温データを用いた. 19∼157mm 目合 19 種類で, 遠洋水産研究所の場合は atsu et al., 2000). 秋生れ群の調査 CPUE は, Yatsu et a
160°E 17 0°E 180° 170°W 16 0°W 150°W 14 0°W 170°E 45°N 40°N Proh ibite Prohibited Se p Aug/Sep Ju l/No v Ju n/De c Nov Au g Oct Jul 44°N 42°N 46 °N 43°N 145°W Hawaii Is . 14 0°E 15 0°E 13 0°W 120°W 20 °N 20°N 30 °N 30°N 40 °N 40°N 50 °N 50°N d りの漁獲尾数(あるいは漁獲重量)で表した. 衛 (http://podaac-esip.jpl.nasa.gov/poet/)から入手した. 資源量の推定方法 1. 面積密度法 生物の分布密度を主分布域に引き延ばして, 資源量を推定する方法である. 流し網漁業データから 密度への換算は次のようにして行った. 流し網は, アカイカの北上回遊を遮るように東西方向に海 に一晩敷設し, 夜間浮上して移動するアカ 面 イカを漁獲する(図4). 従って, アカイカの密度 d (g/m2) は次式より求めることができる(田中, 2001).
1
(
C
d
=
(1 ここでは, 流し網 1 反の長さL (m)に, アカイ)
1
)(
00
k
Lvt
) カの北上速度v (m/h)および夜の流し網浸漬時間t (h)を 定している. 乗じ得られる面積に生息するアカイカのうち, 夜間にアカイカが水深 7m(流し網の網丈)以浅に滞在 する時間割合(k %)に見合う分のアカイカC (kg)が漁獲されたと仮定している(図 4). また, アカ イカの羅網率は100%( 網に遭遇したアカイカは網に刺さり, ひれが引っかかって, はずれない)と 仮2. DeLury 法 漁場への移入・漁場からの逸散がなく、資源が漁獲のみによって減少する場合に, 初期資源量を推 定する方法である. 本法では資源量および漁獲量を尾数に変換し, 単位努力量当り漁獲尾数が資源尾 に比例して変化すると仮定している. DeLury 法には幾つかのモデルがあるが, アカイカでは自然死 図4 流し網によるアカイカ漁獲とそれに基づく密度推定 数字は1985 年のアカイカ 度の推定に用いた値。 密 数 亡率(M)を無視できないので(村田・嶋津,1982), Mを取り込んだ次式を用いた.
)
2
/
1
(
)
ln(
C
tX
t+
M
t
+
=ln(
)
(
/
2
)
1 0 0 t t i iX
X
q
qN
−
∑
+
t=0, 1, ··· ,n-1 (2) ここで, Cは漁獲尾数, X は努力量(反), N0は初期資源尾数, qは漁具効率を示す. また, i, tは漁期(旬) を示す添字, 例えば, Ctは, 第t旬の初期の漁獲尾数を示す. 漁獲の機構につ − = いて, t期の漁獲尾数はt 期の漁獲期間平均資源尾数に比例するとしている.ln(
C
tX
t)
+
M
(
t
+
1
/
2
)
を(
/
2
)
1 0 t t i iX
X
+
∑
に 対してプロットすることにより, N0を推定することができ − = る. ョンモデル(ASPIC モデル) 量指数に基づいて資源評価を行うための簡単な資源動態モデルで, 平衡状態の仮定を 使 具効率, 内的自然増加率, 環境収容力は一定である. 単年性アカイカの場合は② 定には問題がないと考えられる. ⑤については, スルメイカ の場合, レジームに応じてパラメータを変化させて適用されている(谷津・木下, 2002). アカイカ秋 3. 非平衡プロダクシ 漁獲量と資源 わずにパラメーターを推定する. イカ類へのの適用例はカナダイレックスやスルメイカの例がある (Hendrickson et al. 1996; 谷津・木下, 2002). ASPIC モデルの基本的な仮定は次の通りである(田 中, 1999):①閉じた漁場である, ②資源の反応に時間遅れがない, ③漁獲開始年齢に変化がない, ④ 年齢組成の変化と資源の増減に関係がない, ⑤漁 , ③, ④の仮 生れ群の場合も, レジームを考慮して適用した.本研究では, CPUE に観測誤差を仮定した次式の ASPIC モデル(Prager,1994)を用いた.
B
qX
r
t t t t)
2−
ここで, Btは時間K
rB
dt
dB
=
(
−
/
(3) tにおける資源量, Kは環境収容力, qは漁具能率, rは内的自然増加率, Xtは漁獲努力 である. また, 単位努力量当り漁獲量(CPUE)はqBt, 漁獲量はqXtBtと表すことができる. なお, 量B0 MSY 与 )で計算 た カ の 生 群 外 長 春 れ 秋には亜熱帯 域 ア 日本, 韓国および台湾による秋生れ群の月別漁獲量をみると, 7月に最も多い傾向にあった(図 6). また日本のデータによる秋生れ群のCPUE(kg/反)も 7 月に最も高くなる傾向を示した(表 1). 従
頻
度
%
(処女資源)はKの近辺とするのが妥当であるので, B1-ratio(計算の開始年における資源量と える資源量の比)を2 に固定して, ASPIC コンピューター・プログラム(Prager,1995 .結果
イカ秋生れ群の漁獲量と CPUE(単位努力量当り漁獲量)の月変化 1991 年の漁獲物の外套長組 月変化をみると, 6~8 月には秋生れ群(外套長 30∼35cm 以上の個体)のみが, 9 月になると秋 (外套長40cm 以上)と冬春生れ群(外套長 30∼38cm)が, 10 月になると冬春生れ群のみ( 30∼40cm)が漁獲されていた(図 5). すなわち, 9 月中旬までは秋生れ群を, それ以降は冬 群を主に漁獲していた. 村田(1990)も, 秋生れ群は 9 月頃に急速に南下を始め, へ移ると報告している. 以上より, 170°E以東の海域において5月初め~9月半ばに漁獲されてい カイカを秋生れ群とみなすことができる. を し ア 成 れ 套 生 水 た6月
0 5 10 15 N=724 0 5 107月
N=1424 0 59月
108月
10 N=116210月
図5 1991 年の日本のいか流し網漁業で漁獲されたアカイカの月別の外套長組成外套長(cm)
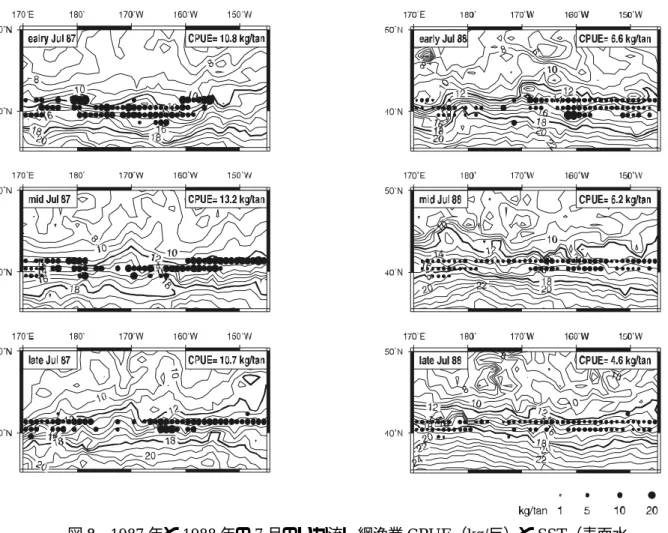
0 5 10 20 30 40 50 10 N=123 0 5 N=472 , って, 秋生れ群の盛漁期は 7 月ということになり, 漁期(5∼9 月)のほぼ中間に位置していた.表1 1982∼1992 年の日本の流し網漁業による月別アカイカ CPUE(kg/反) カイカ秋生 3 つの方法で推定した. . 面積密度法 7 月 との関係でみると, 12.0∼18.0℃ 海域で高い 図7 1979∼2002年7月における表面水温とアカイカ秋生れ群の調査CPUE(尾 数 夏における北上回遊時の適水温は SST12.0∼18.0℃と述べている. いか流し網操業も, 7 月には SST が 積 同月の漁業の平均CPUE を適用することにより, 資源量を推定した. 1991 1992 平均 2.2 4.0 6.3 5.2 7.8 8.2 6.4 7.3 7.0 3.8 4.7 5.3 4.2 5.0 5.3 3.8 2.7 5.2 4.5 4.0 5.3 4.3 5.1 6.1 25 図6 日本, 韓国および台湾の流し網漁業によるアカイカ秋生れ群の月別漁獲量 0 5 1982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 6月 5月 10 15 20 万 年 漁獲量(ト ン ) 7月 8月 9月 れ群の資源量推定 盛漁期(7 月)における秋生れ群の資源量を の秋生れ群の調査流し網CPUE(尾/10 反)を表面水温 (SST) 値を示した(図7). Ichii et al. (2004)も, アカイカ秋生れ群の春から 月 1982 1983 1984 1985 1986 1987 1988 1989 1990 6 8.2 9.8 5.6 7.4 6.3 8.1 8.4 4.5 4.6 7 9.8 12.9 6.8 8.9 5.9 11.5 5.7 5.5 10.1 8 8.0 11.1 4.4 7.0 5.1 8.3 4.3 3.5 12.0 9 8.7 9.8 3.5 5.9 3.6 5.7 3.1 3.7 6.2 10 7.9 6.3 3.9 5.2 4.0 4.0 4.3 7.5 6.4 11 7.2 7.1 4.9 6.0 3.8 5.5 4.3 7.6 4.7 12 5.6 9.5 4.5 5.4 5.2 5.0 4.5 5.9 3.8 平均 7.9 9.5 4.8 6.5 4.8 6.9 4.9 5.5 6.8 0 5 10 15 20 25 30 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 表面水温(℃) 調査C P U E ( 尾/ 1 0 反 ) ア 1 の /10 反)の関係 ほぼ12.0∼18.0℃内の海域で行われていた. 例として, 7 月の CPUE が高かった 1987 年と低かった 1988 年について, いか流し網漁業 CPUE の分布(緯度 1 度×経度 1 度)を SST との関係で旬別にみ た(図8). 以上より, 7 月の秋生れ群の主分布域は SST12.0∼18.0℃と考えられるので, この全面 に
図8 1987 年と 1988 年の 7 月のいか流し網漁業 CPUE(kg/反)と SST(表面水 等温線は12.0 および 18.0℃を示す. CPUE の値は旬別の平均. の北上速度vは, 7 月における適水温の平均 この水温の旬別推移から計算した. ちなみに, 1982~92 標識アカイカの再捕データからの推定値380 m/時(新谷 1987 9 時間で, 本海域におけ , 7m 以浅の滞在時 間割合 は平均 以上より, 1 m2となる. 1982∼1992 年の7 月における であることより, dは0.086 ∼0.330 /m2 7 月の適水温帯の面積は 150∼233 万 km2 . 2. DeLu アカイ 8 月には索餌域となる移行領域に分布する(村田, 1990;Ichii et al., 2004). 漁獲が集中するこの時期, CPUE(kg/反)の推移に減少傾向がみられるので, この期間の 6 旬のデー 温)の旬別分布 太線のSST アカイカの密度は(1)式に基づいて推定した. 1 反の長さLは 50m である. アカイカ 15℃の北上速度に一致すると仮定し, 年の平均vは420m/時と算出され, )とほぼ同様であった. 漁場における 7 月の夜の時間tは約 るアカイカのバイオテレメトリー調査によると, アカイカは夜間には表層に浮上し 25%であった(田中, 未発表). 回の操業での1 反当りの有効面積(Lvt)は144,000~257,000 1 反あたりの漁獲量C(すなわちCPUE)は 5.2∼13.0 kg となる(表2). いか流し網漁場(170°E-145°W)における であるので, 資源量は 15∼63 万トンと推定される(表 2) k g ry 法 カ秋生れ群は, 7~ タを用いて, 7 月 1 日(N0)のアカイカ資源尾数を推定した(図9). なお, CPUE は日本のデータを 用い, 漁獲量Cは日本, 韓国, 台湾の合計を用い, 努力量XはC/ CPUE から求めた. 自然死亡率Mの 値としては北西太平洋の釣り漁場(アカイカ冬春生れ群)で推定された0.07/旬(村田・嶋津,1982) を用いた.
表2 面積密度法におけるパラメーター値と 7 月の資源量推定値 得られた初期(7 月 1 日)資源尾数N0, 漁具効率qを年別に表3 に示した. 1982∼1992 年のN 0.84∼3 1 億尾と推定された. ただし, 1990 年と 1991 年は CPUE に減少傾向がみられず, N0 ができなかった. 面積密度法による推定値(7 月の平均値;表 2)と比較するには、7 月半ばの資源尾 を求める必要がある. N0, Mおよびqより, 7 月 15 日の資源尾数は 0.63∼2.71 億尾と推定され, 重量に換算すると11∼45 万トンとなった(表 3). qについては, N0が小さい年ほど大きくなる傾向 を示した(r2=0.61). 7月の 流し網 アカイカ適水温 夜の時間 夜間アカイカが7m アカイカ 170°E-145°Wでの アカイカ 年 CPUE 1反の長さ 北上速度 以浅に滞在する 分布密度 アカイカ適水温域の 資源量 (kg/反) (m) (m/時) (時間) 時間の割合(%) (g/m2) 面積(万km2) (万トン) 1982 9.8 50 390 9 25 0.223 213 47.7 1983 13 50 350 9 25 0.330 192 63.3 1984 6.8 50 390 9 25 0.155 204 31.6 1985 8.9 50 320 9 25 0.247 215 53.0 1986 5.9 50 490 9 25 0.107 227 24.3 1987 11.5 50 340 9 25 0.301 172 51.7 1988 5.7 50 400 9 25 0.127 150 19.0 1989 5.5 50 570 9 25 0.086 173 14.8 1990 10.1 50 370 9 25 0.243 178 43.2 1991 5.2 50 410 9 25 0.113 193 21.8 1992 7.8 50 550 9 25 0.126 233 29.3 平均 8.2 50 416 9 25 0.187 195 36.3 0は, の推定 数 資源 量, X:努力量, M:自然死亡係数. 7~8 月の 6 旬のデータに基づく. .2 0 0.5 0 500 1000 1500 2000 Cummulative tan (x104) In 1983年 y = -0.000304 x + 2.276562 1.5 2 X )+ (Σ M/2 -0.00 0 0.5 0 500 1000 1500 2000 Cummulative tan (x104) In (C 1986年 1.5 2 X )+ (Σ M/2 ) y = -0.000326 x + 1.593148 0.5 0 500 1000 1500 2000 Cummulative tan (x104) In (C 1989年 2 (Σ 1982年 5 y = -0.000501 x + 2.018769 1 1.5 2. (C/ X )+ (Σ M+ 2 2 (Σ ) (Σ ) 2 M / 2) 0 0.5 0 500 1000 1500 2000 1 2.5 / M+ ) Cummulative tan (x104) In (C 1984年 y = -0.000151 x + 1.470206 0 0.5 0 500 1000 1500 2000 Cummulative tan (x104) In (C 1 2.5 / M+ 1987年 y = -0.000459 x + 1.628569 0 0.5 0 500 1000 1500 2000 Cummulative tan (x104) In (C 1 1.5 X )+ M+ M/ 2) 図9 漁獲と自然死亡を考慮した DeLury モデルによる各年の累積努力量と CPUE の関係 C:漁獲 y = -0.000714 x + 1.792375 0 0.5 0 500 1000 1500 2000 Cummulative tan (x10 ) In 0 0.5 Cummulative tan (x104) (C / M+ y = -0.000358 x + 1.875478 0 0.5 0 500 150 Cummulative tan (x104) I (C / M+ 2 2.5 4 (C/ (Σ 1985年 y = 0317 x + 1.931873 1 1.5 2.5 / X )+ M+ M/2 y = -0.000445 x + 2.238280 1 1.5 2 2.5 0 500 1000 1500 2000 In X )+ (Σ M / 2 ) 1988年 0 1 1.5 2.5 / X )+ M+ M/2 1 1.5 2.5 / X )+ M+M /2) 1992年 1 1.5 2 2.5 1000 0 2000 n X )+ (Σ M / 2)
表3 D 具効率 0 わらず, 調査 CPUE が低くなっている(図 10). ちょうどこの頃に, 北太平 洋で新たなレジームが生じた(自然環境の変化)可能性が示唆されており(見延, 2003), 特に秋生 れ 海域では, 1998 年頃から異変(降水量が減少し, 高塩分傾向)がみられている(Lukas, 海道大学, それ 以降は遠洋水産研究所による. 大型釣り漁船 CPUE は日本の値. eLury 法で用いたパラメーター, 及び推定された資源尾数および漁 年 資源尾数 漁具効率 漁獲尾数 自然死亡率 資源尾数 資源重量 (7月1日) (7∼8月) (7月15日) (7月15日) N0 q C M r2 (×106) (×10-4) (×1 (/旬) (×106) (万トン) 1982 150.3 5.01 33. 0.07 0.758 131.6 21.1 1983 320.5 3.04 58. 0.07 0.720 281.1 45.0 1984 84.1 7.14 37. 0.07 0.930 65.5 10.5 1985 217.7 3.17 55. 0.07 0.543 185.4 29.7 1986 288.1 1.51 44. 0.07 0.736 255.8 40.9 1987 210.7 4.45 70. 0.07 0.788 172.1 27.5 1988 150.9 3.26 53. 0.07 0.898 119.7 19.2 1989 111.0 4.59 52. 0.07 0.987 90.9 14.5 1992 182.2 3.58 47. 0.07 0.240 156.9 25.1 06) 7 3 2 5 6 6 1 2 6 平均 190.6 4.0 50.3 0.1 162.1 25.9 3. 非平衡プロダクションモデル(ASPIC モデル) アカイカ秋生れ群の調査 CPUE は, 1992 年末の流し網モラトリアム以降, 顕著に高くなっている (図10). これは, 流し網による年間 10∼23 万トンの漁獲で減少していた資源が, 流し網の禁止によ り漁獲が激減し, 急速に回復したことを示唆する(Yatsu et al., 200 ). しかし, 1999 年以降になる と, 少ない漁獲にもかか 群の産卵 2001). そこで, このレジームが生じる前の 1979∼1998 年について, 漁獲量および調査流し網 CPUE ( 10 反)データを解析した. その結果, 1982∼1992 年の漁期中期(7 月)における資源量の計算値は 15~30 万トンで(図 11), 環境収容力は37.4 万トン, MSY を与える資源量は 18.7 万トン, MSY は 16.4 万トンと推定された. 0 20 40 60 80 100 120 140 197 9 198 1 198 3 198 5 198 7 198 9 199 1 199 3 199 5 199 7 199 9 200 1 200 3
年
流し 網CP U E ・ 釣 りC P U E 0 5 10 15 20 25 漁獲量 商業流し網 漁獲量(万トン) 漁業釣り 漁獲量(万トン) 大型釣り漁船CPUE (100kg/日) 調査流し網(7月) CPUE(kg/10反) kg/ 図10 アカイカ秋生れ群の漁獲量と CPUE の経年変化 漁獲量は日本, 韓国および台湾の合計値. 調査 CPUE は 1999 年までは北生 れ群の資源量は, 面積密度法では 15∼63 万トン(平均 36 万トン), DeLury 法では 11∼45 万トン(平
平均23 万トン)と推定された. 面積密度法によ
性に影響する. アカイカ科のMについては, 0.033∼0.100/旬と推定
さ
(Pa , 1981)に基づいた値である. カナダイレックス(Illex illecebrosus)では, 0.1/旬が採用され
983)に基づ i, 1984), お . スル 本研究で用いたアカ イ 982)は旬当り 定 (Z)が0.15/ さ れ をみると, M , M=0.07 の時の アカイカ秋生れ群は Mは0.07 よ 本研究や村田・嶋津 (198 は南下回遊に伴う移出があると考えら れ, こ 不確 実性が 図11 170 °E 以東のアカイカ秋生れ群の ASPIC モデルによる資源量推定値
考 察
アカイカ秋生れ群の資源量推定値の信頼性 1982∼ 92年の流し網漁場における7月のアカイカ秋 0 5 10 15 20 25 30 35 1979 1981 1983 1985 1987 1989 1991 1995 1997 年 資源量(万 ト ン --●--) 40 1993 19 均26 万トン), ASPIC モデルでは 15∼30 万トン( る推定値が大きい傾向にあったが, 異なる方法による 3 つの推定値に大きな違いはなかった. ここで は, 各々の方法の推定値の信頼性について検討してみる.ASPIC モデルでは, 資源量の絶対値, MSY を与える資源量(の絶対値), 漁獲係数, MSY を与える
漁獲係数, r, q, Kなどの絶対値の推定精度はよくないとされる(Prager, 1994). また, 加入量が大き く変動する資源には本モデルの当てはまりが悪いことからも(Prager, 1994), 本モデルでの資源量 の推定値15∼30 万トンは不確実性が大きいと考えられる. DeLury 法では, M の推定精度, および漁場への移入・漁場からの逸散がないという仮定がどの程度 満たされているかが推定値の信頼 れている. 例えば, アルゼンチンイレックス(Illex argentinus)では, 0.086/旬が採用されているが (Beddington et al. ,1990), これはさまざまな魚種の自然死亡率, 成長率および生息水温の関係 uly ているが(Hendrickson, 1996), これは, 寿命と自然死亡率の関係の経験則(Hoenig,1
いた 13/旬, 動物のサイズと代謝の関係に基づいた 0.073/旬(Peterson and Wroblewsk
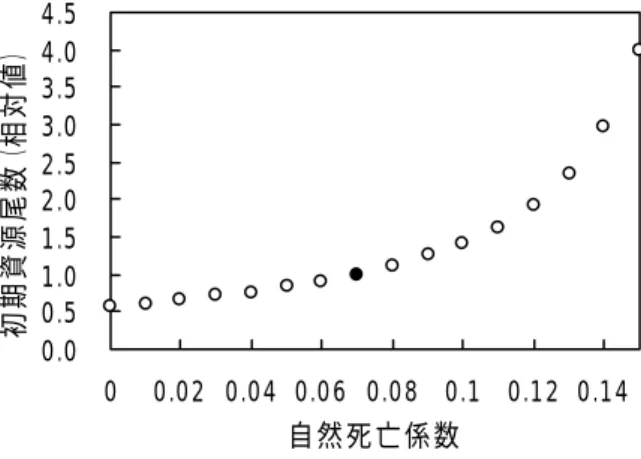
よび近縁種であるアルゼンチンイレックスの0.086/旬(Rosenberg et al., 1990)の平均である メイカでは 0.033/旬が経験的に用いられている(北海道区水産研究所, 2004). カの0.07/旬は, 上記 3 種で用いられているMの平均値に相当する. 村田・嶋津(1 のアカイカ冬春生れ群のMについて, 無視し得る程小さくはないが, 0.07/旬前後以下であろうと推 している. その根拠は, イカ釣り漁場における 2 ヶ月間の CPUE の推移から全減少係数 旬程度と見積もられ, このうち捕食によるアカイカのMは漁獲係数(F)に比べると小さいと推定 , M Zの半分以下の値(0.07/旬以下)であろうというものである. DeLury 法でパラメーターとして与えるMを変化させた時の初期資源量推定値(相対値) が大きくなる程, 資源量が指数的に大きくなっていく. 1986 年を例にとってみると 資源量を1.00 とすると, M=0.03 では 0.71, M=0.00 では 0.57 となる(図 12). 漁場では体長が特大(LL)サイズなので捕食者に食べられることは少ないと考えられ, り小さい可能性が高く, この点では資源量は過大推定となっている. 一方, 0. は 2)では, 漁場からの移出がないと仮定しており, 実際に の点では資源量は過小推定となっている. 以上より, DeLury 法では, Mと移出についての 大きく, 推定資源量の信頼性に問題が残る.
図12 DeLury 法におけるアカイカの自然死亡係数と初期資源尾数との関係 黒丸は本研究で用いた値. は6 月下旬にバイオテレメトリー調査で 追 査を行い, 追跡個体数を増やしてよりk 値 0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 0 0.02 0.04 0.06 0.08 0.1 0.12 0.14 初期資源尾数( 相対値) 自然死亡係数 面積密度法では, ほとんどのパラメーターが観測データに基づいており, その資源量の推定値は他 の 2 つの場合より不確実性が小さいと考えられる. 今後, 面積密度法の推定値の信頼性をさらに高め るためには, まず夜間におけるアカイカの 7m 以浅の滞在時間割合 k の信頼性を高める必要がある. 本研究では, 田中(未発表)による 25%を採用したが, これ 跡した6 個体および 8 下旬~9 上旬月に追跡した 8 個体の計 14 個体の平均である. kと資源量推定 値との関係について1986 年を例にしてみると, kの値が小さくなる程, 資源量は指数的に大きくなっ ていく(図13). kが25%の時の資源量を 1.00 とすると, 15%では資源量がほぼ 2 倍となるが, 35% では資源量が0.90 とあまり変化しない. 今後, 7 月に同様の調 の信頼性を高めていく必要があろう. また, 本研究では,アカイカの羅網率を 100%と仮定している. さけ・ます類については, 回遊方向と直角になるように敷設した流し網の場合, その羅網率(catch efficiency)は 70%と推定されている(Ishida, 1986). アカイカについても, 水中テレビなどの技術 を使って, 実際の羅網率を測定する必要がある. 0.0 0. 1.0 1.5 2.0 2.5 3.0 1 ア イ 資源量( 相対値) 5 0 0 20 30 40 50 60 夜間におけるアカイカの7m以浅の滞在時間(%) カ カ 図13 面積密度法における夜間のアカイカの 7m 以浅の滞在時間とアカイカ資源量推 定値との関係 黒丸は本研究で用いた値 以上, 1982∼1992 年の流し網漁場における 7 月のアカイカ秋生れ群の資源量としては, 面積密度法 による15∼63 万トンが参考となると考えられる. なお, この値は 7 月の平均(すなわち 7 月半ば)の 資源量なので、これに前期(5 月∼7 月前半)の漁獲量を加えたものが, 漁場加入資源重量となり 24 ∼70 万トンと推定される。
いか し網漁獲圧 開発率に基づいて, 1. SPIC モデル 本モデルは 比), 相対漁獲係数 ( msy) これらの値についてみてみる と, 獲量の多か 年以降はFmsyレベルを 上回り1990 年には , 特に 1988 年以降 はB レベルを下回り , アカ 7 月半ばまでの漁獲尾数を加えたものを漁場加入尾数とした. この加入 流 のアカイカ秋生れ群資源への影響評価 ASPIC モデルおよび アカイカ秋生れ群の資源評価をしてみる.
, MSY, 相対資源量(B/Bmsy;資源量と MSY を与える資源量の
などについては, 比較的精度良く推定する(Prager, 1994). った1980~92 年にかけて相対漁獲係数は増加し, 特に 1987 1.6 にまで達した(図 14). 一方, 相対資源量はこの間減少し 1991 年には 0.7 にまで低下した. すなわち, モラトリアム前の数年間は イカ資源は過剰漁獲傾向にあったことになる. 2 2 A F/F 漁 msy --○--) 1.5 (--●--) 図14 ASPIC モデルによるアカイカの相対資源量と相対漁獲係数の経年変化 2. 開発率 開発率 E は, 資源に及ぼす漁獲圧の大きさで, 漁場加入尾数に対する漁獲尾数の割合である.
Sissenwine and Tibbetts(1976)は, シュミレーションの結果, E=40%を最適開発率として奨励して
いる. 0 1979 1981 19 8 3 1985 1987 1989 1991 19 9 3 1995 1997 年 相 0 相 表4 面積密度法における漁場加入尾数と開発率 資源量 資源尾数 前期漁獲尾数 漁場加入尾数 総漁 年 (7月) (7月) (5月∼7月前半) (5月∼ (万トン) (×106) (×106) (×106) ( N' C' N'+C' 1982 47.7 285.5 22.4 307.9 76 1983 63.3 379.3 42.6 421.9 116 1984 31.6 189.5 31.2 220.7 80 1985 53.0 317.6 45.4 363.0 113 1986 24.3 145.5 45.9 191.3 102 1987 51.7 309.5 75.1 384.6 147 1988 19.0 113.6 66.1 179.6 124 1989 14.8 88.9 61.4 150.2 118 1990 43.2 258.9 60.2 319.1 137.6 43.1 1991 21.8 130.3 32.5 本研究で信頼性が最も高いと考えられる面積密度法で推定した資源量を用いた場合, E は次のよう に計算される. まず, この資源量は 7 月の平均値であるので, 7 月半ばの資源量と仮定し, これを尾数 に変換した. これに, 5 月から 平均 1 0.5 1 対資源量( 0.5 1 1.5 対漁獲係数 獲尾数 開発率 9月前半) ×106) (%) C C/(N'+C') .3 24.8 .6 27.6 .7 36.6 .6 31.3 .8 53.7 .9 38.5 .6 69.4 .7 79.0 162.7 86.3 53.0 1992 29.3 175.7 35.8 211.5 89.4 42.3 36.3 217.7 47. 264.8 108.6 45.4
尾数に対する総漁獲尾数(5 月から9月半ばまでの漁獲尾数)の割合, すなわちEは, 平均 45%であ 前には過剰漁獲であった可能性が示唆された. 3. ては今後の課題である. ASPIC モデ . また, 原稿を 読んで有益なコメ , 渡辺光の各氏に深く感 謝する.
Bedd gton J.R., R : Stock assessment and
lkland Islands aters. Fish. Res., 8, 351-365.
im Y.-S. and Hwang S.-J., 1993: Outline of the Korean squid gillnet fishery in the North f .
hortfin squid in the northwest Atlantic during 1993. Northwest Fisheries Science Center
Reference Document, 96-05g, 1-63.
Hoenig J.M., 1983. Empi rates. Fish. Bull., 81:
898-903
北海道区水産研究所 資
源評価,
Hokkaid exploratory
fish
Ichii T., Mahapatr etween the
au n ) related to
the ocea , 1-15.
Ishida Y, 1989: eriment. Bull.
Lukas R, 2001: re associated with
decad 見延庄士郎, 月刊海洋, 35, 45-51. 村 るが, 1982~1989 年にかけては増加傾向を示した(表 4). これは ASPIC 解析における相対漁獲係数 とほぼ同様の増加パターンで, モラトリアム まとめ ASPIC モデルおよび開発率に基づいた評価によると, 大規模に行われていた公海流し網漁業は, アカイカ秋生れ群の適正な漁獲を上回っていた可能性がある. ただし, アカイカ秋生れ群の場合, 漁 場には雌しか分布せず, 雄は未利用となっている. 雄が春∼夏における索餌期にどこにいるのかは定 かでないが, おそらく雌のように北方まで回遊せず産卵場北方の亜熱帯海域に分布すると予想され (Yatsu et al., 1997), その漁獲の可能性につい
謝 辞
ル解析について, 遠洋水産研究所の平松一彦氏にご教示頂き、感謝する ントを下さった同研究所の酒井光夫, 平松一彦, 川原重幸文 献
ozenberg A.A., Crombie J.A. and Kirkwood G.P., 1990 in
the provision of management advice for the short fin squid fishery in Fa w
Gong Y., K
Paci ic Int. N. Pac. Fish. Comm. Bull., 53(I), 45-69.
Hendrickson L.C., Brodziak J., Basson M. and Rago P., 1996: Stock assessment of northern s
rical use of longevity data to estimate mortality .
, 2004: 平成 15 年スルメイカ冬季発生系群の資源評価. 我が国周辺水域の漁業
平成16 年 3 月, 水産庁増殖推進部・独立行政法人水産研究所研究総合研究センター
University, 1980-2000: Data record of oceanographic observations and g. Hakodate, Japan. Faculty of Fisheries, Hokkaido University Nos 22-43. a K., Sakai M., Inagake D. and Okada Y., 2004: Differing body size b and winter-spring cohorts of neon flying squid (Ommastrephes bartramii nographic regime in the North Pacific: a hypothesis. Fish. Oceanogr., 13 Estimation of catch efficiency and chum salmon abundance by parallel gillnet exp Jap. Soc. Sci. Fish. 52, 239-248.
Freshening of the upper thermocline in the North Pacific subtropical gy al changes of rainfall. Geophys. Res. Letters, 28, 3485-3488.
2003: Major regime shift の可能性を秘める北太平洋の 1998/99 年の変化. o in tum 田守, 1990: 北太平洋のおけるいか流し網漁場の海洋環境及びアカイカの分布・回遊.日本海ブロ ック試験研究集録, 17,144-148. 村田守, 嶋津靖彦, 1982: 北西太平洋におけるアカイカの資源特性値いついて.北水研報告 47,1-10.
村田守, 中村好和, 1998: 北太平洋におけるアカイカの季節的回遊および日周鉛直移動,「外洋性大型
イカ類に関する国際シンポジウム講演集」(奥谷喬司編), 海洋水産資源開発センター, 東京,
pp.11-28
長澤和也 森純太, 岡村寛, 1998: 北太平洋のアカイカ系群の生物学的指標としての寄生虫,「外洋性大 類に関する国際シンポジウム講演集」(奥谷喬司編), 海洋水産資源開発センター, 東京,
Prager M. H., 1994: A suite of extensions to nonequilibrium surplus-production model. Fish. Bull. U.S., 90(4), 374-389.
Prager M. H., 1995: User’s manual for ASPIC: a stock-production model incorporating covariates,
rg A.A., Kirkwood G.P., Crombie J. and Beddington J.R., 1990: The assessment of stocks of annual squid species. Fish. Res., 8, 335-350.
新谷久男, 1987: アカイカの分布と回遊. 水産技術と経営, 272, 23-33.
Sissenwine M.P. and Tibbetts A.M., 1976: Simulating the effects of fishing on squid (Loligo and 田中栄
谷津明 (1976-1983 年).研究報告 29,遠
谷津明
谷津明 てパラメーターを変化させたプロダクションモデル:スル
Yatsu atsu H. and Hayase S., 1993: Outline of the Japanese Squid Driftnet Fishery with 谷津明
Yatsu ai Y.,
uring 1979-1998: impact of driftnet fishing and
Yeh S eries in the North Pacific. Int. N.
, 型イカ pp.47-62.
Pauly D., 1981: On the interrelationship between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks. J. Cons. Int. Explor. Mer., 39, 175-192. Peterson I. and Wroblewski J.S., 1984: Mortality rates of fishes in the pelagic ecosystem. Can. J.
Fish. Aquat. Sci., 41,1117-1120.
program version 3.6x. U.S. National Marine Fisheries Service, Southeast Fisheries Science Center, Miami Laboratory, Document MIA-92/93-55, 4th edition, Miami.
Rosenbe
Illex) populations of the Northeastern United States. ICNAF Sel. Pap., 2, 71-84.
次, 1999: 国際会議で用いられている資源評価の手法について. 南西外界の資源・海洋研究, 15, 95-113. 田中博之, 2001: バイオテレメトリーによるアカイカの行動追跡. 海洋と生物,137, 533-539. 彦, 1992: 北太平洋における釣り調査によるアカイカの分布 洋水産研究所,清水,13-37. 彦, 1995: 公海流し網漁業, 「漁業の混獲問題」(松田皎編), 恒星社厚生閣, 東京, pp.52-61. 彦・木下貴裕, 2002: レジームに応じ メイカへの適応例. 平成 13 年度イカ類資源研究会議報告, 29-33. A., Hiram
Notes on the By-Catch. Int. N. Pac. Fish. Comm. Bull., 53(I), 5-24.
彦, 田中博之, 森純太, 1998: 北太平洋におけるアカイカOmmastrephes bartramiiの資源構
造,「外洋性大型イカ類に関する国際シンポジウム講演集」(奥谷喬司編)海洋水産資源開発センタ ー, 東京, pp.29-46.
Yatsu A., Midorikawa S., Shimada T. and Uozumi Y.,1997: Age and growth of the neon flying squid, Ommastrephes bartrami, in the North Pacific Ocean. Fish. Res., 29, 257-270. A., Watanabe T., Mori J., Nagasawa K., Ishida Y., Meguro T., Kamei Y. and Sakur 2000: Interannual variability in stock abundance of the neon flying squid, Ommastrephes
bartramii, in the North Pacific Ocean d
oceanographic conditions. Fish. Oceanogr., 9, 163-170. . and Tung I., 1993: Review of Taiwanese pelagic squid fish