2015年 9 月 2 日受付;2015年12月24日受理. 1
国 立 研 究 開 発 法 人 水 産 総 合 研 究 セ ン タ ー 増 養 殖 研 究 所 志 布 志 庁 舎(National Research Institute of Aquaculture, Fisheries Research Agency, Shibushi Laboratory, Natsui, Shibushi, Kagoshima 899-7101, Japan).
2
国立研究開発法人水産総合研究センター西海区水産研究所奄美庁舎(Seikai National Fisheries Research Institute, Fisheries Research Agency, Amami Laboratory, Setouchi, Oshima, Kagoshima 894-2414, Japan).
3
東 京 農 業 大 学 生 物 産 業 学 部 ア ク ア バ イ オ 学 科(Department of Aquatic Bioscience, Faculty of Bio-industry, Tokyo University of Agriculture, Abashiri, Hokkaido 099-2493, Japan).
4
農 林 水 産 技 術 会 議 事 務 局 研 究 推 進 課(Research Promotion Division, Agriculture, Forestry and Fisheries Research Council, Kasumigaseki, Chiyoda-ku, Tokyo 100-8907, Japan).
5
国立研究開発法人水産総合研究センター西海区水産研究所八重山庁舎(Seikai National Fisheries Research Institute, Fisheries Research Agency, Yaeyama Laboratory, Fukai-Ohta, Ishigaki, Okinawa 907-0451, Japan).
6
国 立 研 究 開 発 法 人 水 産 総 合 研 究 セ ン タ ー 日 本 海 区 水 産 研 究 所 宮 津 庁 舎(Japan Sea National Fisheries Research Institute, Fisheries Research Agency, Miyazu Laboratory, Odashukuno, Miyazu, Kyoto 626-0052, Japan).
7
富山県農林水産総合技術センター水産研究所(Fisheries Research Institute, Toyama Prefectural Agricultural, Forestry & Fisheries Research Center, Takatsuka, Namerikawa, Toyama 936-8536, Japan).
8
国 立 研 究 開 発 法 人 水 産 総 合 研 究 セ ン タ ー 増 養 殖 研 究 所 南 伊 豆 庁 舎(National Research Institute of Aquaculture, Fisheries Research Agency, Minami-izu Laboratory, Irouzaki, Kamo, Shizuoka 415-0156, Japan).
*
連絡先(Corresponding author): Tel, (+81) 99-472-0730; Fax, (+81) 99-472-1544; E-mail, [email protected](T. Tomoda).
資 料
海上網生簀における天然プランクトンを用いた
マダラの半粗放的種苗生産
友田 努
1,*・久門一紀
2・渡邉研一
3・荒井大介
4・
小磯雅彦
5・手塚信弘
6・堀田和夫
7・桒田 博
8Semi-extensive larviculture of pacific cod Gadus macrocephalus
utilizing wild zooplankton in the sea net-cage
Tsutomu T
OMODA1,*Kazunori K
UMON2, Ken-ichi W
ATANABE3, Daisuke A
RAI4,
Masahiko K
OISO5, Nobuhiro T
EZUKA6, Kazuo H
OTTA7and Hiroshi K
UWADA8Abstract: The present study intends to develop larviculture techniques for mass production of
Pacific cod Gadus macrocephalus juveniles possessing better seed quality and lower cost. We evalu-ated the efficiency of semi-extensive larviculture of Pacific cod utilizing wild zooplankton in the sea net-cage. Main food was light-allured zooplankton and supplementary foods were enriched Artemia nauplii, commercial frozen copepoda and dry feed. Metamorphosing cod larvae (12 and 13 mm in total length, TL) were raised until approximately 30 mm TL, and larvae and metamorphosed juve-niles (17, 19, and 20 mm TL) were raised until 45 to 50 mm TL. Survival rates ranged from 46.4 to 48.7% in the former groups, and from 51.4 to 76.5% in the latter groups. A total of 204.8 and 183.7 thousand juveniles were produced in each semi-extensive trial. Moreover, semi-extensive method offered the benefits of labor saving and cost reduction. One juvenile with 30 mm and 45 to 50 mm TL cost only 6.4 yen and 6.2 to 10.5 yen, respectively. These results suggest that cod larviculture utilizing wild zooplankton in the sea net-cage provides faster growth, higher survival rate, and lower cost. We propose that this rearing method may be applicable to various fish species for stock enhancement.
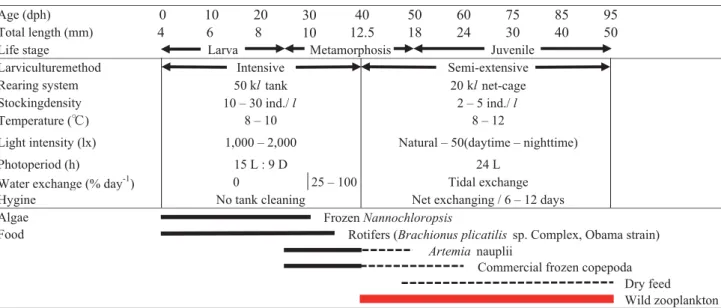
マダラ Gadus macrocephalus は北部太平洋・日本 海・オホーツク海等に分布する冷水系の底棲性魚類で あり,日本海中部の近海では重要な漁獲対象種となっ ている。1990年以降の漁獲量の低迷状態(藤田・堀田 1998)から富山県や石川県では栽培漁業等による資源 回復が強く望まれ,様々な対策が講じられてきた(小 關 1947; 町田 1985)。水産総合研究センター日本海区 水産研究所能登島庁舎(能登島庁舎,旧.日本栽培漁 業協会能登島事業場)では,日本栽培漁業協会能登島 事業場開所時から本種の親魚養成と種苗生産技術開発 に取り組み(中野 1986; 與世田 1993),2003年には50 万尾以上の種苗を生産・放流できる技術水準に達した (久門ら 2003; 荒井 2005)。ここに至るまでには,種々 の要素解明(竹内ら 1994; 鄭ら 1995, 1996, 1997; 渡辺 2003; Matsunari et al. 2005)が進められ,近年は仔魚 期における省力化型の集約的種苗生産(通称,ほっと け飼育,友田ら 2009; 友田・團 2014)や粗放的種苗生 産手法(大野 1992, 2001; 森岡 2002)で育成した仔稚 魚の 健苗性 (日本栽培漁業協会 1996)についても 報告された(荒井ら 2006, 2009; Tomoda et al. 2009)。 すなわち,集約的種苗生産は環境制御された陸上のコ ンクリート水槽に仔魚を高密度で収容して,大量培 養・栄養強化した餌料生物と配合飼料を利用する飼育 方法,粗放的種苗生産は自然環境下の素掘り池など に仔魚を低密度で収容して,自然発生した天然餌料 生物のみを利用する飼育方法として一般に定義され る(大野 2001; Shields 2001)。一方で,種苗期後期に おける省コスト・省力化型の飼育方法として,天然プ ランクトンを主要餌料として活用しつつ,培養した 餌料生物と市販の飼餌料を補助的に利用するハタハ タ Arctoscopus japonicus 方式の半粗放的種苗生産(日 本栽培漁業協会 2002; 友田 2008)も検討された。そ の結果,量産手法としての実用性がうかがわれたも のの,概要の一部が報告されたにすぎない(久門ら 2003)。特に,量産規模におけるマダラ属の研究報告 は欧米の大西洋マダラ G. morhua(Brown et al. 2003; Rosenlund and Halldórsson 2007)に比べて国内には 見当たらない。このことから,量産現場における実用 的な飼育管理のノウハウと技術レベルを記載しておく ことは,本種に限らず国内における栽培漁業対象魚種 の技術開発を進めるうえで極めて重要であると考え る。そこで,本稿では海上網生簀における天然プラン クトンを活用したマダラの半粗放的飼育事例を報告 し,飼育成績と省コスト・省力化の面から本生産手法 を評価するとともに,海上筏施設で考慮すべき飼育管 理上の留意点についても若干の考察を加えた。 材料および方法 親魚養成と採卵 親魚は,氷見漁業協同組合魚市場(氷見漁協)に水 揚げされた天然魚を富山県農林水産総合技術センター 水産研究所(富山水試,旧.富山県水産試験場)の深 層水利用研究施設で 1 ∼ 3 年間養成した個体および 氷見漁協で水揚げした直後の個体を用いた。養成と採 卵方法は渡辺ら(2005)に従った。2003年度は 1 月14 日から 2 月 6 日の間,および2004年度は2003年12月30 日から2004年 2 月17日の間,搾出媒精法による人工授 精または自然産卵による採卵を試みた。採卵は,養成 個体の場合は富山水試で,水揚げ直後の個体の場合は 能登島庁舎で実施した。富山水試で得られた受精卵は 陸上輸送により能登島庁舎に搬入し,20 l ハッチング ジャー(MPC-20,アース)1 ∼ 2 基に収容後,毎日 死卵を除去しながらふ化前日まで自然水温10℃前後で 卵管理した(手塚 2006)。 沖出しまでのふ化仔魚の陸上水槽飼育 2003年度は 1 月19,25日および2004年度は12月30 日,2 月 6 ,10日に採卵した受精卵を50 kl 八角型コン クリート水槽(実容量45 kl)に浮かべた20 l ハッチ ングジャーに収容し,ふ化まで自然水温で卵管理し た。得られたふ化仔魚を沖出しサイズまで陸上水槽 で飼育して,海上網生簀での半粗放的飼育試験に供 した。Fig. 1 に現状におけるマダラ飼育手順の概要を 示した。飼育用水には,電解殺菌装置(電解バリア ESF-030,荏原実業)で殺菌処理した砂濾過海水を用 いた。水槽中央の水面上部には船舶用耐震電灯(片 口天井灯 M2 号15×1PL,110V-60 W 電球)2 個を設 置し,飼育期間中を通して昼夜点灯状態にした。餌 料には,ふ化後 4 ∼ 40日齢にシオミズツボワムシ Brachionus plicatilis sp. complex(L 型 小 浜 株, 平 均 背甲長250.0μm),20 ∼ 60日齢にアルテミア Artemia sp. 幼生(北米産,太平洋貿易),25 ∼ 60日齢に後述 のエアーリフト装置により灯火採集した天然プランク トン,45 ∼ 60日齢に冷凍養成アルテミア(全長1.2 ∼ 1.5 mm)および50 ∼ 70日齢に市販の配合飼料(初期 飼料協和 A-400, B-400, 700,C-700, 1000,協和発酵; アンブローズ200, 400, 600,日本配合飼料)を用いた。 生物餌料の栄養強化および陸上水槽での飼育方法は 既報(竹内ら 1994; 久門ら 2003; 渡辺 2003; 荒井 2005; Matsunari et al. 2005)に準じた。 仔稚魚の沖出しサイズの検討 実用的な半粗放的飼育を目的として,ハタハタの
種苗生産(友田 2008)に準じた収容密度( 4 ∼ 6 万 尾/面)で適正沖出しサイズを検討した。供試魚に は,50 kl コンクリート水槽で飼育中のマダラ仔稚魚 を適宜抽出した。2003年度は 3 月19,20,21日(各 42,49,44日齢)に変態中期のマダラ仔魚 2 群(全 長12,13 mm 半粗放群)合計43.46万尾を網生簀 8 面 (大きさ3.0×3.0×2.5 m,実容量20 kl)に沖出しした。 半粗放的飼育における網生簀 1 面当たりの収容密度は 2,345 ∼ 3,640尾/kl であった。2004年度は 3 月 3 ,9 , 12日(各54,60,63日齢)に変態後期および変態完 了後の仔稚魚 3 群(17,19,20 mm 半粗放群)合計 38.51万尾を網生簀 8 面に沖出しした。半粗放的飼育 における網生簀 1 面当たりの収容密度は1,326 ∼ 4,661 尾/kl であった。なお,両年度とも対照区として網生 簀 1 面当たりに2,000尾(100尾/kl)を収容する無給 餌の粗放的飼育事例(13,17 mm 粗放群)を設けた。 コンクリート水槽からの取り揚げには,テトロンラッ セル(T-90,目合い2.5 mm,アース)の巻き網,カ ナライン A ホース(内径50.6 mm,アース)と種苗 移送ポンプ(ピンピンZ-65L,松坂製作所)を用いて 500 l ポリカーボネイト水槽(SPS-500,アース)に収 容した。魚数計(FCH-10 稚魚カウンター,日本海洋) を用いて,おおよその尾数を収容しておき,改めて水 槽内に強通気を施しながら容量法により計数を行っ た。すなわち,2 l 計量カップを用いてポリカーボネ イト水槽の 3 定点から合計 6 l の海水を採取した。そ の後,生存している仔稚魚数を計数,1 l 当たりの密 度を確定して総収容尾数を算出した。沖出しは,ポリ カーボネイト水槽 1 ∼ 2 基を能登島庁舎の栽培漁業調 査船「のとじま丸」(4.5トン)に積載し,酸素通気を 施しながら庁舎地先の海上筏施設まで低速航走して 行った。航走中は,船のローリング・ピッチングによ る水槽からのオーバーフローを防ぐため,輸送水量を 400 l まで減水して水面に10 l バケツ 7 個を浮かべ波 打ちを緩衝した。海上筏施設では水槽水量を約250 l まで減水して,10 l バケツを用いて海水ごと仔稚魚を すくい取り網生簀に収容した。沖出し作業は正規職員 5 名と臨時職員 5 名で行った。 海上網生簀における半粗放的飼育 飼育には,網生簀(ナイロンモジ網 N22-380,300, 240,220,200径,日東製網)を用いた。飼育手順は Fig. 1 に準じた。網生簀中央の水面上部には船舶用耐 震電灯 1 個を設置し,陸上水槽と同様に昼夜点灯状態 にして主要餌料となる天然プランクトンを蝟集させた (Fig. 2)。補助的餌料として,アルテミア幼生を変態 完了(全長18 ∼ 19 mm )に至るまで 1 日 1 回(13時 半)与えた。網生簀 1 面当たりの給餌量を500 ∼ 1,500 万個体/日とした。アルテミア耐久卵は脱殻処理(吉 松 1999)し,水温24℃で48時間ふ化管理した後に幼 生を分離した。アルテミア幼生は栄養強化剤(スー パーカプセル A1,クロレラ工業)を250 ml/億個体の 割合で添加して,水温20℃で20時間栄養強化した。ま た,従来の陸上飼育で与えている冷凍養成アルテミ アの代替として市販の冷凍コペポーダ 2 種類(Cyclop-eeze,Argent Chemical Laboratories; 雅 1 号,サイエン テック)を用いた。冷凍コペポ−ダは凍結状態のまま タマネギ袋に入れ,網生簀中央の水面下網カゴに垂 下して 1 日 2 回( 9 時半,16時半)与えた。網生簀 1 面当たりの給餌量を100 ∼ 600 g /日とした。配合飼料 (初期飼料協和 B-400, 700, N-400, 700, C-700, 1000,協 和醗酵; アンブローズ200, 400, 600, 800,日本配合飼料; オリエンタルなぎさ No. 5, 7,オリエンタル酵母工業) は仔稚魚の成長にあわせて 2 ∼ 4 種類を混合し,全長 17 mm 前後から網生簀中央の水面上部に設置した自 動給餌器(YDF-160SO およびコントローラ A ,ヤマ ハ発動機)を用いて給餌した。給餌方法は,ハタハタ (友田 2008)に準じて飼育初期は夜間中心の間欠給餌 (18時より翌朝 6 時まで 2 時間間隔で計 7 回/日),後 期は連続給餌( 2 時間間隔で計12回/日)とした。網 生簀 1 面当たりの給餌量を25 ∼ 900 g /日とした。網 生簀の交換は 6 ∼ 12日間隔で行い,仔稚魚の成長に あわせてモジ網の目合いを拡大した。網替え頻度を少 なくするため,側面の網地を外側から適宜デッキブラ シ掛けすることで付着物を除去し,潮通しを改善し た。また,飼育初期における減耗要因のひとつである 波浪対策として,網生簀を垂下する沈子重量を従来 ( 1 面当たり 1 kg コンクリート製沈子 8 個)よりも増 加させ,四隅に 3 kg,側面中央に 2 kg の鉄製沈子を 各 4 個ずつ(計 8 個)を設置した。飼育期間中,作業 時以外は網生簀上面を防鳥網(大きさ3.5×3.5 m,目 合い 3 cm)で覆い,海鳥による食害を防除した。そ の他,海上網生簀での飼育方法は既報(日本栽培漁業 協会 2002; 友田 2008)に準じた。毎朝 9 時に箱メガネ で網生簀の底面を観察しながら死亡魚をタモ網ですく い取り計数した。なお,取り揚げ時における生残尾数 の計数は重量法により行った。次式に従い死亡尾数の 補正を行い,毎日の生残率を求めた。 M1× C,M2× C,M3× C, …,Mt × C:1,2,3, …, t 日目における死亡尾数 た だ し,M1,M2,M3, …,Mt:1,2,3, …, t 日目における死亡魚の計数値 C = D/D'(D≧D'):補正係数 N0:収容尾数 Nt:t日目の取り揚げ尾数 S = Nt/N0:試験期間を通した正確な生残率
D = 1-S:試験期間を通した正確な死亡率 D' = (M1+M2+M3+ … +Mt)/N0:死亡魚の計数 値に基づく死亡率 なお,M1,M2,M3,…,Mt は試験期間中のどの 時点の死亡が大きいのかを明らかにすることを目的と しているので,見落とし率が一定になるように死亡魚 の取り揚げ努力量を一定にした。また,補正係数 C は網生簀毎に求めた。毎日のルーチンワーク(死亡魚 除去,給餌など)は正規職員 1 名と臨時職員 1 名,不 定期の網替え作業は正規職員 2 名と臨時職員 3 名で 行った。 天然餌生物調査 網生簀内の天然餌生物の出現状況を確認するため, 飼育期間中は毎日餌生物の出現数調査を行った。マダ ラ仔稚魚を収容していない網生責 2 面(ナイロンモジ 網 N22-380 径)の電灯直下に蛸集した餌生物をダイヤ フラムブロワー(DF-150,アース)に接続したエアー リフト装置(吐出量15 l/分)を用いて採集した。夕 方 4 時から翌朝 9 時までの17時間連続でナイロンプラ ンクトンネット(17 XX,目合い80μm,アース)に 採集し,容量法で計数して餌生物数を求めた(森岡 2002; 荒井ら 2006)。網生簀 2 面の平均値を 1 晩にお ける 1 面当たり出現数とした。
Fig. 2. Semi-extensive larviculture of Pacific cod Gadus macrocephalus in sea net-cages with plankton-alluring lamps.
a, Daytime; b, Nighttime.
Fig. 1. Current larviculture protocols for juvenile Pacific cod Gadus macrocephalus in Notojima Laboratory, JSNFRI, FRA.
海上網生簀における生産コストの試算 経費は,マダラ種苗生産にかかる直接経費のうち餌 料費,備品費,資材消耗費,光熱水費,人件費につ いて算出した。本稿では,生産事例毎に沖出し時か ら取り揚げ時までの種苗期後期の飼育期間中に要し た総経費を取り揚げ尾数で除して,海上網生簀にお ける種苗 1 尾当たりの生産コスト(岩本ら 2006; 中川 ら 2006)として試算した。なお,施設(海上筏,橋 形クレーン,冷凍冷蔵庫)や高額機材(船舶,フォー クリフト,高圧洗浄機,種苗移送ポンプ,魚数計)な どの減価償却費については,ハタハタ種苗生産・配布 業務やワムシ高密度輸送業務との共用,および他庁舎 からの借用物品であるため,試算には含めなかった。 一方,マダラ種苗生産のみで使用する備品(自動給餌 器,コントローラ)は償却年数 6 年,資材消耗品(網 生簀,沈子,船舶用耐震電灯,防鳥網,タモ網,バケ ツ,ロープ,キャブタイヤコード,デッキブラシ,箱 メガネ,計量カップなど)は償却年数 3 年として,1 年間に要した費用として算出した(中川ら 2006)。自 動給餌器,コントローラ,電灯および高圧洗浄機の稼 働に要する電気代と網生簀の洗浄に要する水道代など の光熱水費は網生簀 1 面を管理するに当たり便宜上 4,000円/月として加算した。人件費は飼育作業に要す る必要人数,業務時間,日数および雇用単価から算出 した。雇用単価は,正規職員2,800円/時間および臨時 職員750円/時間とした。 結 果 仔稚魚の飼育成績 Table 1に 各 飼 育 群 を 累 計 し た 育 成 結 果 を 示 す。 2003年度の12,13 mm 半粗放群と2004年度の17 mm 半粗放群は,試験開始時にはそれぞれ網生簀 4 ,3 お よび 3 面であったが沖出し後の減耗が顕著であった ため,飼育開始22,20および 7 日目にそれぞれの群を 2 面に併合して網生簀毎の生残率を求めた(Fig. 3)。 2003年度における12,13 mm 半粗放群では27 ∼ 29日 間の飼育により平均全長27.6 ∼ 29.1 mm の稚魚20.48 万 尾 を 取 り 揚 げ た( 平 均 生 残 率46.4 ∼ 48.7%)。 一 方,対照区の13 mm 粗放群では21日間の飼育により 平均全長26.5 mm の稚魚1,178尾を取り揚げた(生残 率58.9%)。12 mm 半 粗 放 群 は13 mm 半 粗 放 群 よ り も沖出し後 4 日間の減耗が顕著に多かった(Fig. 3)。 2004年度における19 mm 半粗放群 1 区では15日間の 飼育により平均全長30.6 mm の稚魚7.36万尾(生残率 78.9%),17,19( 2 区 ),20 mm 半 粗 放 群 で は33 ∼ 41日間の飼育により平均全長45.3 ∼ 50.2 mm の稚魚 18.37万尾を取り揚げた(平均生残率51.4 ∼ 76.5%)。 一方,対照区の17 mm 粗放群では37日間の飼育によ り平均全長45.0 mm の稚魚1,865尾を取り揚げた(生 残率93.3%)。17,19,20 mm 半粗放群では小型サイ ズほど沖出し後 4 日間の減耗が大きく,特に17 mm
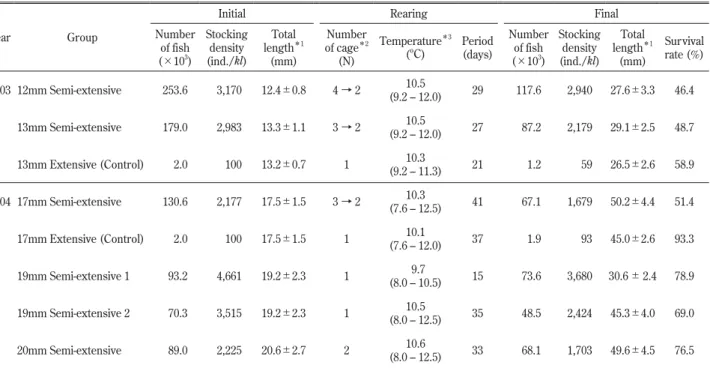
Table 1. Results of semi-extensive larviculture trials of Pacific cod Gadus macrocephalus in the sea net-cages
Year Group
Initial Rearing Final Number of fish (×103 ) Stocking density (ind./kl) Total length*1 (mm) Number of cage*2 (N) Temperature*3 (ºC) Period (days) Number of fish (×103 ) Stocking density (ind./kl) Total length*1 (mm) Survival rate (%) 2003 12mm Semi-extensive 253.6 3,170 12.4±0.8 4 → 2 (9.2 – 12.0)10.5 29 117.6 2,940 27.6±3.3 46.4 13mm Semi-extensive 179.0 2,983 13.3±1.1 3 → 2 (9.2 – 12.0)10.5 27 87.2 2,179 29.1±2.5 48.7 13mm Extensive (Control) 2.0 100 13.2±0.7 1 10.3 (9.2 – 11.3) 21 1.2 59 26.5±2.6 58.9 2004 17mm Semi-extensive 130.6 2,177 17.5±1.5 3 → 2 (7.6 – 12.5)10.3 41 67.1 1,679 50.2±4.4 51.4 17mm Extensive (Control) 2.0 100 17.5±1.5 1 (7.6 – 12.0)10.1 37 1.9 93 45.0±2.6 93.3 19mm Semi-extensive 1 93.2 4,661 19.2±2.3 1 (8.0 – 10.5)9.7 15 73.6 3,680 30.6 ± 2.4 78.9 19mm Semi-extensive 2 70.3 3,515 19.2±2.3 1 10.5 (8.0 – 12.5) 35 48.5 2,424 45.3±4.0 69.0 20mm Semi-extensive 89.0 2,225 20.6±2.7 2 10.6 (8.0 – 12.5) 33 68.1 1,703 49.6±4.5 76.5 *1 Mean±S.D. (n = 30 – 121)
*2 Net-cage ; Scale 3.0 × 3.0 × 2.5 m (volume 20 kl) *3
Fig. 3. Survival curves of Pacific cod Gadus macrocephalus larvae and juveniles during larviculture trials in sea
net-cages. +, 12mm Semi-extensive A; ×, 12mm Semi-extensive B; +, 12mm Semi-extensive C; ×, 12mm Semi-extensive D; ◇, 13mm Extensive; ◆, 13mm Semi-extensive A; ◆, 13mm Semi-extensive B; ◆, 13mm Semi-extensive C; △, 17mm Extensive; ▲, 17mm Semi-extensive A; ▲, 17mm Semi-extensive B; ▲, 17mm Semi-extensive C; ●, 19mm Semi-extensive 1; ●, 19mm Semi-extensive 2; ■, 20mm Semi-extensive 1; ■, 20mm Semi-extensive 2.
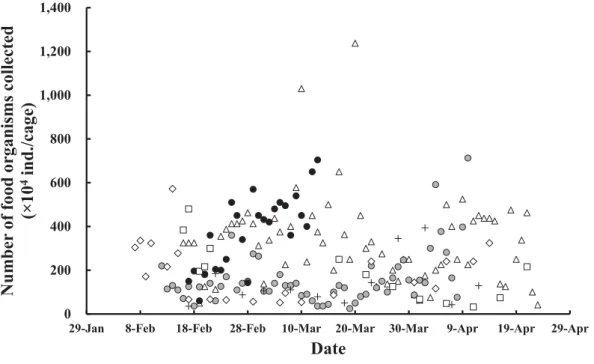
Fig. 4. Daily changes of number of food organisms collected from the sea net-cage. +, 2000 (Morioka 2002);
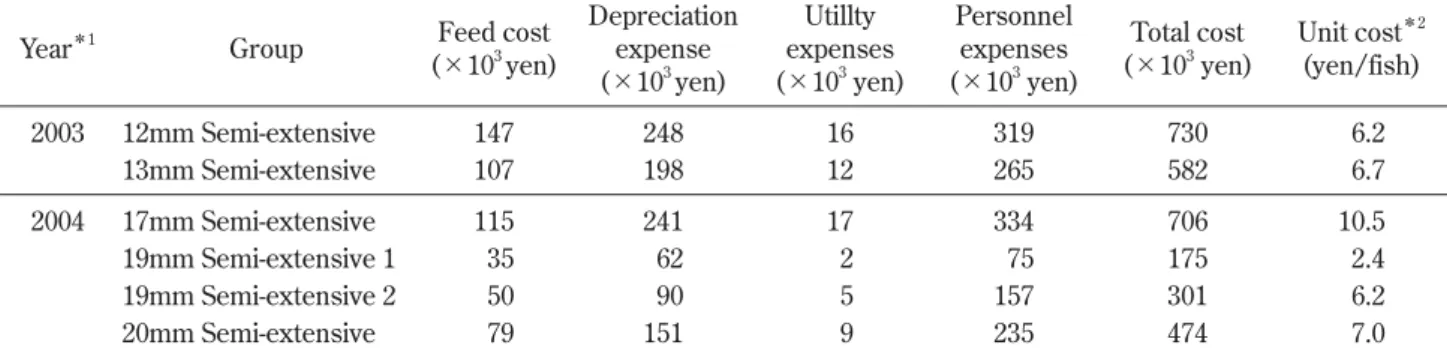
半粗放群では沖出し後翌日のまでの減耗が顕著であっ た(Fig. 3)。なお,17 mm 粗放群と17 mm 半粗放群 の間で生残率に顕著な差が見られたことは沖出し時に おけるハンドリング負荷(実数計数とポンプ移送計 数,陸上輸送と船舶輸送,コンクリート水槽→輸送容 器→網生簀間の収容方法)の違いによるものであり, 17 mm 粗放群では比較的丁寧な取り扱いがなされた 結果である。 餌生物の出現状況 Fig. 4 に2000年から2006年度にわたり調査した網生 簀内の餌生物採集量を示した。餌生物の出現状況に は年変動と日変動が大きかった。2003年度は平均163 万 個 体/面・ 日(25 ∼ 713万 個 体 ) で あ り,2000年 度(平均142万個体),2005年度(平均198万個体)お よび2006年度(平均193万個体)と同等であった。一 方,2004年度は平均380万個体/面・日(60 ∼ 704万 個体)であり,2002年度(平均331万個体)と同等で あった。なお,両年度とも飼育開始時に観察した餌 生物の個体数組成では枝角目 Cladocera とカイアシ亜 綱 Copepoda が全体の 7 ∼ 8 割を占めており,前述の 採集量とあわせて既報(森岡 2002; 荒井ら 2006, 2009) と同様の傾向であった。 生産コスト Table 2 に海上網生簀での半粗放的飼育に要した餌 料,主な飼育用具と実働時間を示した。本集計結果と 前述の飼育結果(Table 1)をもとに,海上網生簀で の半粗放的飼育に要した経費を試算したところ,総経 費は2003年度では1,313千円,2004年度では1,655千円 となった(Table 3)。両年度の全体経費に占める割合 は,人件費46.9%,減価償却費(備品・資材消耗品) 33.2%,餌料費18.1%,光熱水費1.9%の順となった。 マダラ種苗 1 尾当たりの生産コストは全長12,13 mm から30 mm サイズまでの飼育では平均6.4円/尾(そ れぞれ6.2,6.7円/尾),全長17 mm から50 mm サイズ までの飼育では10.5円/尾,全長19,20 mm から45, 50 mm サイズまでの飼育ではそれぞれ6.2,7.0円/尾 となった。なお,全長19 mm から30 mm サイズまで の短期飼育(15日間)の場合は2.4円/尾と低くなった。
Table 2. Total amount of food, rearing gear and total hours spent at work required in semi-expensive larviculture of Pacific
cod Gadus macrocephalus in the sea net-cage
Year*1 Group Artemia nauplii (×108 ind.) Commercial frozen copepoda (kg) Commercial dry feed (kg)
Major rearing gear (piece) Total hours spent at work(man・hour)
Net-cage Sinker Automatic feeder controllerFeed Staff Part-timer
2003 12mm Semi-extensive 6.2 28.2 12.8 12 40 4 1 79.5 128.7 13mm Semi-extensive 4.3 20.0 9.9 10 32 3 1 65.7 108.0 2004 17mm Semi-extensive 2.1 15.5 38.2 15 32 3 1 87.5 118.0 19mm Semi-extensive 1 0.2 8.1 7.6 3 12 1 1 19.5 27.8 19mm Semi-extensive 2 0.2 8.1 18.2 6 12 1 1 42.7 49.5 20mm Semi-extensive 0.0 9.2 37.8 10 24 2 1 63.3 76.3 *1 Cod larvae were raised until approximately 30 mm TL in 2003, and larvae and juveniles were raised until 45 to 50 mm TL except for 19 mm Semi-extensive 1 in 2004.
Table 3. Estimate of expenses in semi-expensive larviculture of Pacific cod Gadus macrocephalus in the sea net-cage
Year*1
Group Feed cost (×103 yen) Depreciation expense (×103 yen) Utillty expenses (×103 yen) Personnel expenses (×103 yen) Total cost (×103 yen) Unit cost*2 (yen/fish) 2003 12mm Semi-extensive 147 248 16 319 730 6.2 13mm Semi-extensive 107 198 12 265 582 6.7 2004 17mm Semi-extensive 115 241 17 334 706 10.5 19mm Semi-extensive 1 35 62 2 75 175 2.4 19mm Semi-extensive 2 50 90 5 157 301 6.2 20mm Semi-extensive 79 151 9 235 474 7.0
*1 Cod larvae were raised until approximately 30 mm TL in 2003, and larvae and juveniles were raised until 45 to 50 mm TL except for 19mm Semi-extensive 1 in 2004.
考 察 本試験により,変態期から変態完了後の早期沖出し によるマダラ半粗放的種苗生産が量産規模で可能であ ることが実証された(Table 1)。本生産手法の有効性 は,粗放的種苗生産事例(荒井ら 2006, 2009; Tomoda et al. 2009)のみならず,類似する飼育方法によるハ タハタ量産事例(日本栽培漁業協会 2002; 友田 2008) からも支持されるものと考える。海上網生簀では,変 態初期の発育段階から天然魚本来の餌生物であるカイ アシ亜綱や枝角目など(Takatsu et al. 1995; 森岡・桑 田 2002; 荒井ら 2006)を捕食させることができる。こ のため,海上網生簀で飼育された稚魚は天然海域での 索餌能力を備え,飢餓耐性能や形態発育に加えて外敵 からの逃避能力が優れている可能性があることも期待 されている(荒井ら 2009)。 本試験において,海上網生簀における生残率は沖出 しサイズに伴い高くなる傾向であった(Fig. 3)。一方 で,17 mm 半粗放群の顕著な減耗は沖出し時にかかっ た過大なハンドリング負荷と沖出し当日から 4 日間に わたる暴風波浪の影響が大きいものと推測された。こ れまでに,沖出し後の主な減耗要因として,① 沖出 し時のハンドリング負荷(取り揚げ,計数,船舶輸 送,網生簀収容),② 網替え時の擦れ,および ③ 成長 差に伴う噛み合いの三つが観察されている。これらに 暴風波浪などの自然要因が加わることにより,減耗の 度合いはさらに顕著となる傾向であった。特に,主 要因となる ① については,海上筏への輸送条件(海 況,沖出しサイズ,輸送密度,水温,溶存酸素濃度な ど)および網生簀への収容方法を考慮する必要があ る。② については,網替え作業時のハンドリングミ スのみならず,仔稚魚の活力不足,運動能力の不備や 外部形態の未発達さに起因するものと考えられ,海上 飼育を開始する至適サイズを選定する必要がある。な お,19,20 mm 半粗放群の生残率が70%前後と比較 的に高かった背景として,水槽や網生簀などの障害物 に対する感覚器官の発達,行動様式の変化,さらには 少々網擦れしても生残に影響しない程度までの外部形 態(鰭,皮膚など)の発達が考えられる。行動様式の 変化については全長14 mm 前後で夜間灯火に対する パッチ形成が顕著となり,18 mm 前後で電灯直下を 成群回遊し始めること,外部形態の発達については 全長15 mm 前後で各鰭条が定数に達し,18 mm 前後 で下顎先端にアンテナの役割を持つ髭が出現,20 mm 時には体側背部に色素胞が発達し体色が飴色を呈する など仔魚期から稚魚期への移行に伴う頑強性の向上が 確認されている(久門,未発表)。なお,感覚器官の 発達と行動,外部形態との関連についは現時点では不 明である。③ については,定期的なモジ網選別や密 度調整を検討する必要がある。 海上網生簀における配合飼料の摂餌状況につい て,給餌開始当初は頻繁に微粒子飼料(粒径230 ∼ 420μm)が自動給餌器に詰まり設定量が落下しな かったため,全長17 mm の給餌開始から20 mm 時ま で餌付きが遅れた。この問題は,海面からの湿気上昇 によるものと考えられ,自動給餌器に配合飼料と乾燥 剤 2 ∼ 3 袋を充填することで改善された。配合飼料の 餌付け方法については,配合飼料の種類(製造メー カー)により嗜好性や成長・生残が大きく異なり,全 長15 mm からの餌付け開始が実用的であり,19 mm から開始すればより短期間で餌付き,生残率も高くな ることが確認されている(渡辺 2003)。すなわち,全 長15 mm から陸上水槽で餌付けを開始して,変態完 了する18 ∼ 19 mm までに餌付かせることで基礎体力 の強化を図り,沖出しする生産スタイルがその後の減 耗を防除するうえでも適当と考える。 陸上水槽で一貫飼育する従来の集約的生産手法で は,ふ化から全長30 mm サイズ(平均76.7日齢)まで の平均生残率は24.9%である(久門ら,未発表)。一 方,現状の生産手法において,陸上水槽での省力化 型集約的生産では全長20 mm サイズ(平均54.3日齢) までの平均生残率は33.4%である(友田ら,未発表)。 続く,海上網生簀での半粗放的生産では全長19 mm から30 mm サイズ(75日齢)までの生残率は78.9% と高いため(Table 1),ふ化からの累積生残率は約 26.4%と試算される。このことから,種苗期後期から の半粗放的生産への移行は飼育成績の面で遜色ないと 言える。 餌生物の出現状況について,網生簀内の採集量は既 報(森岡 2002; 荒井ら 2006, 2009)の年平均142,198, 193万個体/面・日と同等であり,個体数組成もマダ ラ仔稚魚の食性(Takatsu et al. 1995; 森岡・桑田 2002; 荒井ら 2006)と同様の傾向を示したことから,主要 餌生物として十分に寄与していたものと判断される。 マダラ仔魚は環境中に優占する餌生物を捕食する日和 見的捕食者であるとされる(高津 2003)。本試験で補 助的に用いたアルテミア幼生,冷凍コペポーダや配合 飼料は給餌後,直ちに仔稚魚に利用される分以外は潮 流・風雨または沈降により網生簀内に長時間滞留せず 流出していたことから,餌料としての遭遇機会,すな わち摂餌可能時間は常に網生簀内に流入する天然プラ ンクトンよりも少なかったものと推測される。現状で は本生産手法を支える天然プランクトンの貢献度は不 明であり,今後は仔稚魚の成長に伴う摂餌量や摂餌選 択性を調査する必要がある。
粗放的種苗生産では,網生簀内の収容尾数増加に伴 いマダラ仔稚魚 1 尾が摂餌できる餌生物数が必然的に 制限されるため,成長速度と無給餌生残指数の低下が 生じる(荒井ら 2009)。すなわち,天然プランクトン のみに依存して無給餌条件下で飼育できる収容密度に は限度があるため,本飼育手法のような市販飼餌料の 補助的給餌は必須である。天然プランクトンを利用し た半粗放的飼育(大野 2001)は,他の魚種において も省コスト・省力化および種苗性強化(日本栽培漁業 協会 1996)を図る中間育成の手段として活用できる。 現状ではマダラの種苗性を厳密かつ科学的に評価する までには至っていないため,今後はさらなる栄養学・ 生化学・行動学的調査と合わせて,生産種苗の健全性 と比較放流試験の効果を検証する必要がある。 ハタハタの半粗放的生産は秋田県単県で年間500万 尾を生産可能な技術水準にまで達している(日本栽培 漁業協会 2002)。能登島庁舎では放流試験用として年 間30万尾前後の生産規模に抑えているものの,2003年 以降は85%前後の高生残率を維持し続けている。本種 の種苗生産ではワムシ・アルテミア幼生の培養と給餌 が全く不要であり,労働人員数を要する沖出しと取 り揚げ計数以外のすべての飼育管理を正規職員 1 人と 臨時職員 1 ∼ 2 人でまかなえる。このため,ふ化から 全長30 mm サイズまでの生産コスト(減価償却費と 光熱水費を含まない)は年平均2.5 ∼ 5.1円/尾である (友田 2008)。マダラの半粗放的生産も同様に変態期 からの飼育管理を海上網生簀で行うため,陸上での種 苗生産に見られるコンクリート水槽,送水用ポンプ, 通気用ブロワー,加温用ボイラー,濾過・殺菌装置な どの動力電源を要する施設やライフラインとなる大型 機器を使用せずに済む(Fig. 1,Table 2)。このため, ほとんど光熱水費が掛からず,人件費に次いで比較的 安価な備品費と資材消耗費,および餌料費が生産コス トの97%以上を占める(Table 3)。また,飼育管理も 拘束時間の短いルーチンワーク(死亡魚除去,冷凍コ ペポーダや配合飼料の補充)と不定期の非ルーチン ワーク(網生簀側面のブラシ掛け,6 ∼ 12日間隔の網 替え)に限定されるため,計画的に行えば大幅な省コ スト・省力化が実現可能である。陸上水槽で一貫飼育 する従来の集約的生産手法では,ふ化から全長30 mm サイズまでの生産コスト(減価償却費と光熱水費を 含まない)は年平均12.7円/尾である(荒井ら,未発 表)。一方,種苗期後期から海上網生簀で飼育する現 状の種苗生産手法(Fig. 1)では大幅なコスト軽減が 可能となる。まず,陸上水槽における省力化型の集約 的生産では全長20 mm サイズまでの生産コストは年 平均3.6円/尾である(友田ら,未発表)。続く,海上 網生簀における半粗放的生産では全長12,13,19 mm から30 mm サイズまでの生産コストはそれぞれ6.2, 6.7,2.4円/尾であり(Table 3),単純に積算しても6.0 ∼ 10.3円/尾に抑えられる。なお,全長19 mm から 30 mm(19 mm 半粗放群 1 区)または45 mm(19 mm 半粗放群 2 区)まで育成する場合,飼育日数,生残率 と生産コストから勘案すると30 mm サイズでの取り 揚げが効率的と考えられる(Table 1,3)。 ワムシ・アルテミア幼生など生物餌料の培養・栄 養強化技術や配合飼料開発は従来よりも進展したも のの,天然プランクトンの餌料価値には依然として 及ばない面も多いと考えられる(荒井ら 2006, 2009; Tomoda et al. 2009)。そして,そのような現状が今の 種苗生産技術レベル,すなわち飼育成績や放流効果な どを反映している可能性も考えられる(北島 1993; 日 本栽培漁業協会 1996)。一般に,ふ化から放流までの 長い過程において,上記のような培養可能な生物餌料 に依存する期間は非常に短く,これら餌料に起因する 飼育初期の成長不良や形態異常は本試験のような天然 プランクトンの代替利用により十分に克服可能な場合 もある(友田 2008; 荒井ら 2009)。さらに,海上網生 簀における中間育成では放流までの長期間にわたり海 上生活を経験させることができるため,種苗性(日本 栽培漁業協会 1996)の付与ならびに放流海域への馴 致効果も期待できる。我々は本手法を様々な栽培漁業 対象魚種に適用可能な種苗生産手法として提案した い。一方,天然プランクトンの発生量は年度・立地条 件に左右されるものの,能登島地先のような北部日本 海のみならず,気象・海況・陸水からの栄養塩流入な どの環境条件が良好な西日本の内湾海域でも省コス ト・省力化型の健苗生産モデルとして成立しうると考 える。このような点を踏まえ,今後は未利用生物資源 である天然プランクトンを活用した半粗放的飼育(大 野 2001; Shields 2001; 友田 2008)のさらなる技術開発 を進めつつ,新たな天然餌料の探索とその培養研究に も取り組む必要があると考える。 要 約 マダラ稚魚の種苗量産技術を開発するため,海上 網生簀の明かりに蝟集する天然プランクトンを利用 した半粗放的飼育を検討した。変態中期のマダラ仔 魚(全長12,13 mm 群)を約30 mm サイズまで育成 したところ,生残率は46.4 ∼ 48.7%となった。変態 後期と変態完了後の仔稚魚(17,19,20 mm 群)を 45 ∼ 50 mm サイズまで育成したところ,生残率51.4 ∼ 76.5%となった。この結果,それぞれの飼育事例で 204.8千尾および183.7千尾の稚魚を生産することがで きた。本飼育方法による種苗 1 尾当たりの生産コスト
は30 mm サイズで平均6.4円,45 ∼ 50 mm サイズで は6.2 ∼ 10.5円と見積もられた。海上網生簀で天然プ ランクトンを利用する生産手法は高成長,高生残のみ ならずコスト軽減も期待できることが示された。我々 は本手法を様々な栽培漁業対象魚種に適用可能な種苗 生産手法として提案したい。 謝 辞 本研究を行うにあたり,日本海区水産研究所能登島 庁舎の臨時職員の皆様には餌料培養や飼育作業に多大 なるご協力をいただいた。ここに記して深謝の意を表 します。 文 献 荒井大介(2005)マダラの種苗生産技術の現状について∼ 大量生産と放流が可能になり,地先にとどまるマダラ の生態∼.豊かな海,7,14-17. 荒井大介・友田 努・森岡泰三(2006)天然プランクトン を 利 用 し た 海 上 網 生 簀 に よ る マ ダ ラ Gadus macrocephalus 仔稚魚の飼育.水産増殖,54,409-410.
[Arai, D., T. Tomoda and T. Morioka (2006) Larval cul-ture of Pacific cod using net cages in the sea with zoo-plankton. Aquacult. Sci., 54, 409-410 (in Japanese with English abstract).]
荒井大介・友田 努・手塚信弘・堀田和夫(2009)海上網 生簀で飼育したマダラ仔稚魚の飢餓耐性.水産増殖,
57,61-69. [Arai, D., T. Tomoda, N. Tezuka and K. Hotta
(2009) Starvation resistance of Pacific cod (Gadus
macrocephalus) larvae and juveniles reared in the sea
net-cage. Aquacult. Sci., 57, 61-69 (in Japanese with English abstract).]
Brown, J. A., G. Minkoff and V. Puvanendran (2003) Larviculture of Atlantic cod (Gadus morhua): progress, protocols and problems. Aquaculture, 227, 357-372. 藤田大介・堀田和夫(1998)最近の富山県マダラ漁獲量に
つ い て. 富 山 水 試 研 報,10,27-40. [Fujita, D. and K. Hotta (1998) On the recent catch of Pacific cod Gadus
macrocephalus in Toyama prefecture. Bull. Toyama Pref. Fish. Res. Inst., 10, 27-40 (in Japanese with English
abstract).]
岩本明雄・山崎英樹・藤本 宏・奥村重信・山本義久・小 畑泰弘(2006)サワラの種苗生産単価の試算.栽培技 研,33,61-65. [Iwamoto, A., H. Yamazaki, H. Fujimoto, S. Okumura, Y. Yamamoto and Y. Obata (2006) Estimation of seed production costs for the Japanese Spanish mackerel, Scomberomorus niphonius. Saibai
Giken, 33, 61-65 (in Japanese with English abstract).]
北島 力 編(1993)放流魚の健苗性と育成技術.水産学 シリーズ,93,恒星社厚生閣,東京,119 pp. [Ed. by Kitajima, C. (1993) Healthy Fry for Release , Kouseisha-kouseikaku, Tokyo, 119 pp (in Japanese).]
久門一紀・渡辺研一・友田 努(2003)Ⅱ-1.4.3マダラ種苗 生産技術の開発.平成15年度日本栽培漁業協会事業年 報,51-52.
町 田 洋 一(1985) マ ダ ラ の 種 苗 生 産. 水 産 の 研 究,4, 46-51.
Matsunari, H., D. Arai, M. Koiso, H. Kuwada, T. Takahashi and T. Takeuchi (2005) Effect of feeding rotifers enriched with taurine on growth performance and body composition of Pacific cod larvae Gadus macrocephalus.
Aquacult. Sci., 53, 297-304.
森岡泰三・桑田 博(2002)七尾湾北湾とその沖における マダラ稚魚の生息上限水温と食性.日水誌,68,345-350. [Morioka, T. and H. Kuwada (2002) The upper limit of inhabiting temperature and the diet of juvenile Pacific cod Gadus macrocephalus in the northern part of Nanao Bay and its vicinity, Japan. Nippon Suisan Gakkaishi, 68, 345-350 (in Japanese with English abstract).]
森岡泰三(2002)プランクトン螺集ランプを設置した海面 網生簀におけるハタハタ仔稚魚の食性.日水誌,68, 526-533. [Morioka, T. (2002) The diet of japanese sandfish Arctoscopus japonicus larvae and juveniles reared in a net-cage with a plankton-alluring lamp. Nippon
Suisan Gakkaishi, 68, 526-533 (in Japanese with English
abstract).] 中川雅弘・大河内裕之・有瀧真人(2006)クロソイの種苗 単価の試算.栽培漁業センター技報,(独)水産総合研 究センター,神奈川,5,28-33. 中野昌次(1986)マダラの種苗生産.さいばい,39,23-27. 日本栽培漁業協会(1996)生産力応用技術開発報告[Ⅴ]− 放流用マダイの種苗性強化技術開発−.日本栽培漁業 協会,東京,130 pp. 日本栽培漁業協会(2002)栽培漁業技術シリーズ No. 8「ハ タハタの生物特性と種苗生産技術」.日本栽培漁業協会, 東京,113 pp. 大野 淳(1992)マダイの粗放的種苗生産に関する研究. 特別研究報告 2 号,日本栽培漁業協会,東京,110 pp. [Ohno, A. (1992) Fundamental study on the extensive seed production of the red seabream, Pagrus major.
Special Research Report No. 2 , Japan Sea-Farming Association, Tokyo, 110 pp (in Japanese with English abstract).]
大野 淳(2001)コペポーダを利用する種苗生産方法のす すめ.月刊アクアネット,湊文社,東京,12,29-35. 小關信章(1947)富山湾に於けるタラの人工孵化放流事業
現状.農林時報,6,24-27.
Rosenlund, G. and O. Halldórsson (2007) Cod juvenile production: Research and commercial developments.
Aquaculture, 268, 188-194.
Shields, R. J (2001) Larviculture of marine finfish in Europe.
Aquaculture, 200, 55-88.
Takatsu, T., T. Nakatani, T. Mutoh and T. Takahashi (1995) Feeding habits of pacific cod larvae and juveniles in Mutsu Bay, Japan. Fish. Sci., 61, 415-422.
高津哲也(2003)底生魚類仔稚魚の時空間分布と餌料環境 に関する研究.日水誌,69,543-546. [Takatsu, T. (2003)
Field research on spatial and temporal distributions of demersal fish larvae and juveniles and their prey abun-dance. Nippon Suisan Gakkaishi, 69, 543-546 (in Japanese).] 竹内俊郎・鄭 鋒・與世田兼三・廣川 潤・渡邉 武(1994)
DHA 強化ワムシのマダラ仔魚に対する栄養価.日水誌,
60, 641-652. [Takeuchi, T., F. Zheng, K. Yoseda, J.
Hirokawa and T. Watanabe (1994) Nutritive value of DHA-enriched rotifer for larval cod. Nippon Suisan
Gakkaishi, 60, 641-652 (in Japanese with English abstract).]
手塚信弘(2006)第 6 章:親魚飼育の具体例と将来構想. 親魚飼育の具体例−マダラ.水産重要魚種の生殖機構 と水温による影響.水研センター研報,別冊4,147-149. [Tezuka, N. (2006) Session 6: Broodstock rearing of resource fishes. A concrete example for rearing broodstock of Pacific cod (Gadus macrocephalus). In
Reproductive properties and environmental effects on reproduction in resource fishes . Bull. Fish. Res. Agen.
Supplement No.4, 147-149 (in Japanese with English abstract).]
友田 努(2008)近年のハタハタ種苗生産技術開発.栽培 漁業センター技報,8,9-13.
友田 努・團 重樹(2014)低温馴致したワムシを用いた マ ダ ラ 仔 魚 の 止 水 式 飼 育. 水 産 増 殖,62,307-318. [Tomoda, T. and S. Dan (2014) Stagnant water larvicul-ture using the rotifer Brachionus plicatilis acclimated at low temperature in Pacific cod Gadus macrocephalus.
Aquacult. Sci., 62, 307-318 (in Japanese with English
abstract).]
Tomoda, T., D. Arai, N. Tezuka and K. Hotta (2009) Towards the improvement in seed quality of reared Pacific cod (Gadus macrocephalus). In Larvi ’09–Fish & Shellfish
Larviculture Symposium (ed. by C. I. Hendry, G. Van
Stappen, M. Wille and P. Sorgeloos), European Aquaculture Society Special Publication no. 38, pp. 440-443.
友田 努・手塚信弘・小磯雅彦・荒井大介・島 康洋・榮 健次(2009)省力化型マダラ種苗生産手法の検討. 栽培漁業センター技報,9,15-19. 渡辺研一(2003)3. 冷水性魚類の早期種苗生産技術開発(マ ダラ)2 )マダラ種苗生産における天然プランクトンに 替わる餌料の探索と配合飼料化に向けた飼育試験.平 成14年度日本栽培漁業協会事業年報,110-112. 渡辺研一・堀田和夫・桑田 博(2005)富山県水産試験場 で海洋深層水を用いて飼育したマダラ親魚の日長処理 による採卵時期の早期化.栽培漁業センター技報,3, 4-8. 吉松隆夫(1999)やさしくできるアルテミア耐久卵脱殻処 理のすすめ方.月刊養殖,緑書房,東京,36 (2),106-112. 與世田兼三(1993)マダラ種苗生産技術開発の現状.日本 海ブロック増養殖研究推進連絡会議報告,27,33-44. 鄭 鋒・竹内俊郎・與世田兼三・廣川 潤・渡邉 武(1995) 種苗生産過程におけるマダラ仔稚魚の体脂肪酸組成の 変化.日水誌,61,756-761. [Zheng, F., T. Takeuchi, K. Yoseda, J. Hirokawa and T. Watanabe (1995) Changes in fatty acid composition of cod larvae in mass production.
Nippon Suisan Gakkaishi, 61, 756-761 (in Japanese with
English abstract).]
鄭 鋒・竹内俊郎・與世田兼三・小林真人・廣川 潤・渡 邉 武(1996)アルテミア幼生摂餌期のマダラ仔稚魚 のアラキドン酸,EPA および DHA 要求.日水誌,62, 669-676.[Zheng, F., T. Takeuchi, K. Yoseda, M. Kobayashi, J. Hirokawa and T. Watanabe (1996) Requirement of larval cod for arachidonic acid, eicosapentaenoic acid, and docosahexaenoic acid using by their enriched
Artemia nauplii. Nippon Suisan Gakkaishi, 62, 669-676
(in Japanese with English abstract).]
鄭 鋒・竹内俊郎・小林真人・廣川 潤・渡邉 武(1997) 形態の異なるビタミン E のワムシおよびマダラ仔魚へ の取り込みとワムシ中 DHA 含量の影響.日水誌,63, 77-84. [Zheng, F., T. Takeuchi, M. Kobayashi, J. Hirokawa and T. Watanabe (1997) A study of vitamin E absorption in rotifer and cod larvae and the effect of DHA content in rotifer for cod larvae. Nippon Suisan Gakkaishi, 63, 77-84 (in Japanese with English abstract).]