はじめに わが国を始めとする先進国では,医療技術の進歩, 高齢化,癌,エイズなどによって免疫能の低下した患 者の増加により,医療現場において真菌症は重大な問 題となっている.また,ヒト,物質の国境を越えた移 動の急激な増加による輸入感染症としても増加してい る(http://www.pf.chiba-u.ac.jp/clinical/mycosis. html). 千葉大学真菌医学研究センターは,国内唯一の真菌 症とその原因菌を専門に研究する公的機関である.そ の研究活動の一環として,病原真菌・放線菌を収集 し,安定的に性状を保った状態で保存し,それらを必 要とする研究者(利用者)に提供(分譲)を行ってい る.当初,この菌株保存・提供業務は,センター長付 の非公式なグループで行っていたが,2010 年 4 月に バイオリソース管理室として正式な組織となった.そ の使命は,病原真菌・放線菌の「収集・保存・提供」 体制を整備し,最新情報が付加された信頼できる菌株 の提供を通じて,真菌症ならびにその原因菌の研究・ 教育の基盤を支援することである.その運営は室長, 助教,技術職員各 1 名,技術補佐員 2 名の計 5 名体制 であったが,2020 年 3 月に定年を迎える助教との引 継ぎ期間を設けて本年 7 月に助教が採用され,日々の 業務のなかで技術,知識の習得,継承を行っている. 筆者は,2003 年,千葉大学真菌医学研究センターに赴 任して以来,病原真菌・放線菌の収集・保存・提供を 行うとともに,病原真菌および関連菌において形態的 な知見,系統分類上重要な有性型と無性型との関連, 薬剤感受性など生理的特徴および複数の遺伝子による 分子系統的な知見を勘案した多相的な(polyphasic) 分類研究を実施してきた.この知見の活用により,効 率的な菌株の分離,収集が可能となり,迅速診断や治 療などの応用研究に発展させることができる.さら に,これら情報およびゲノム情報を付与することによ り菌株に付加価値が生まれ,研究の推進につながって いる. 臨床菌株コレクションの整備状況 1)菌株の収集・保存・提供 当センターでは,主要な深在性,表在性真菌症原因 菌とそれら関連菌種における臨床分離株および基準株 を網羅的に収集,保存している.特に病原真菌として 重 要 な 属 で あ る Candida,Aspergillus,Cryptococ-cus,白癬菌の臨床株を世界的に収集し,その数はそれ ぞれ約 6,100 株,4,600 株,1,700 株,2,300 株,他の菌 種との総計約 22,000 株に上る.保存は,胞子を形成す る株は L- 乾燥法で,菌糸のみの菌株などは−80℃で の凍結で行っている.従来は形態を主に菌株の同定を 行っていたが,現在では分類の基準となる rDNA-ITS 領域の塩基配列,Aspergillus,Penicillium において は β-tubulin もしくは calmodulin 遺伝子の塩基配列を 決定し,正確な同定を実施している. 患者由来の喀痰,皮膚などの臨床検体の採取は医療 機関のみ可能である.そのため,医療機関との連携を
病原真菌の分類学的研究と臨床菌株コレクションの充実
(平成 30 年度日本微生物資源学会賞受賞)
矢口貴志
千葉大学真菌医学研究センター 〒260-8673 千葉市中央区亥鼻 1-8-1Studies on classification of pathogenic fungi and enhancement of collection of
clinical isolates
Takashi Yaguchi
Medical Mycology Research Center, Chiba University, 1-8-1, Inohana, Chuou-ku, Chiba 260-8673, Japan
受 賞 総 説
密にし,臨床現場へのフィードバック,たとえば起因 菌の同定,薬剤感受性試験,学会・論文発表のサポー トなどを通じて,新鮮な臨床株,臨床情報(患者の性 別,年齢,基礎疾患,菌株の分離源,薬剤の処方歴, 渡航歴など)の提供を受けている(図 1).病原微生物 を使用した研究においては,この臨床情報がたいへん 重要である. 当センターの保存菌株のうち約 7 割が真菌症原因菌 であり,真菌全般を保存している世界的な保存機関で あ る NBRC,JCM( と も に 日 本 ),ATCC( ア メ リ カ),CBS(オランダ)より臨床由来株ははるかに多 く,臨床情報も充実している.遺伝子の塩基配列,薬 剤感受性などの性状,臨床情報など研究者の求める情 報をデータベースとして整備,公開し(https://daph ne.pf.chiba-u.jp/distribution/catalog),利用者に提供 (分譲)できるシステムを構築している.菌株カタログ ページでは,塩基配列が DDBJ などに登録されている 場合は,そのアクセッション・ナンバーとリンクし, 関連文献が表示され,菌名,菌株ナンバーによる検索 だけではなく,DNA 塩基配列による相同性検索機能 もあり,利用者の利便性を図っている. 生の病原微生物を取り扱えない機関には DNA で提 供するなど,ユーザーの拡大を図るとともに安全かつ 効率的な運営を実施している. 2)高度病原性微生物の提供体制の整備 病原微生物においては,「生物多様性条約」「名古屋 議定書」に基づく「遺伝資源の取得の機会及びその利 用から生ずる利益の公正かつ衡平な配分に関する指 針」のほか,病原菌株を収集・保存にはさまざまな規 制がある.「感染症の予防及び感染症の患者に対する 医療に関する法律(感染症法)」およびこの法律に関連 した政令・省令に基づき,国立大学法人千葉大学病原 体等安全管理規程が制定されている.さらに本学内規 程に基づき,千葉大学真菌医学研究センター微生物取 扱管理規程が定められている.千葉大学では,感染症 法等の一部を改正する法律に基づき,三種病原体等と して Coccidioides immitis 所持の届出を行った.ま た,国立感染症研究所の定める病原体等の分類に基づ き, 学 内 規 程 に お い て 表 1 に 示 す よ う に 真 菌 を BSL1-3 に分類している.センター内規程では,現場 に即した病原体の取り扱い施設と運営,実験室の日常 安全管理実験室等の運営,緊急時対策,病原体等を取 り扱う職員などの資格と健康管理などを定めている. 特に緊急時の対策では,三種病原体等を取り扱う職員 には病原体の逸出を防止する社会的責任がある.具体 的には,災害などの緊急時には,三種病原体および使 用中の実験器具をオートクレーブに入れ密封(電源は 入れない),もしくは準備した密閉容器に入れ,ドア, 排気系を閉鎖し退去すると定めている.3 年間に 1 度 の割合で,厚生労働省による第三種病原体の管理状況 確認の立入検査がある. 高度病原菌の取り扱う際は HEPA フィルターを介 した給排気設備を備えた室内を陰圧にすることで外部 とは遮断,病原体を遺漏させず封じ込めが可能な施設 で,研究者は防護服を着用して行っている.さらに, その施設内でグローブボックスと呼ばれる陰圧の装置 内でグローブ越しに作業する新たな設備を 2016 年に 導入した(図 2). バイオテロの手段となりうる高度病原性微生物の提 供体制の整備は,安全な社会の構築にとってきわめて 重要である.そのため,輸入真菌症原因菌 5 種の保存 菌株のリストは非公開とし,提供は設備の整った機関 に,抽出した DNA のみに限っている. 図 1 医療機関との連携による菌株の収集と提供
3)ナショナルバイオリソースプロジェクト(NBRP) の支援 文部科学省では 2002 年度からナショナルバイオリ ソースプロジェクト(NBRP)を開始し,国が戦略的 に整備することが重要な生物資源について体系的に収 集,保存,提供などを行うための体制を整備してきた. その後 5 年ごとに対象生物種の見直しを行い,2017 年度より第 4 期が開始された. 第 3 期までは病原細菌グループ(岐阜大学,大阪大 学)と病原真菌・原虫グループ(千葉大学,長崎大学) は NBRP「病原微生物」として活動してきたが,第 4 期より別々に活動することとなり,NBRP「病原真核 微生物」では千葉大学真菌医学研究センター(病原真 菌・放線菌,中核機関)と長崎大学熱帯医学研究所 (病原性原虫)は,相互の機関の連携を図り,これらの 病原微生物株の収集・保存・提供体制を整備して,高 度情報を賦与した信頼できる病原微生物株として提供 し,感染症と病原体の教育・研究をする人々を支援し ている.究極の目標として本プロジェクトは,今後い かなる感染症が発生しても対応できる病原真核微生物 コレクションを目指している.当初,NBRP では,リ ソースの提供は無償で行っていたが,受益者負担,本 事業の安定的な運営のため,現在では提供にかかわる 費用の実費を利用者に負担していただいている. また,本事業の一環として,ホームページに病原真 菌・放線菌のギャラリーを開設し,主要な菌種の形態 を電子情報としてネット上で公開している(http:// www.pf.chiba-u.ac.jp/gallery.html). 4)病原真菌の利用目的 教育目的以外に以下のような目的のため,病原真菌 を提供している. (1)原因菌の同定のためのレファレンス株 (2)薬剤感受性試験 (3)感染実験 (4)交配試験 (5)分類学的研究 (6)生物資源,遺伝子資源としての有効利用 特に直近 5 年以内に分離された臨床株は,薬剤評 価,感染症動向調査のため需要が高い.新規な薬剤を 開発し,その有効性を評価する場合,多くの菌種,同 図 2 クラス 3 レベル無菌実験室内 表 1 千葉大学における真菌の BSL 分類 BSL1 BSL2 および BSL3 に属さない真菌 BSL2
Aspergillus fumigatus Fonsecaea pedrosoi
Candida albicans Microsporum canis
Cladosporium carrionii Sporothrix schenckii Cladosporium trichoides Trichophyton (C. bantianum) T. mentagrophytes Cryptococcus neoformans T. verrucosum Exophiala dermatitidis
BSL3
Blastomyces dermatitidis Histoplasma farciminosum Coccidioides immitis(三種) Paracoccidioides brasiliensis Histoplasma caspsulatum1) Penicillium marneffei
1)H. capsulatum var capsulatum と H. capsulatum var duboisii の両 variant を含む.
註:Aspergillus spp., Chaetomium spp., Fusarium spp., Myrothecium spp., Penicillium spp. の毒素産生株は,BSL2 扱いとする.
また,上記 BSL3 の真菌で Coccidioides immitis 以外のものは二形性真菌であり,こ れらが酵母形の場合においては,BSL2 扱いとする.
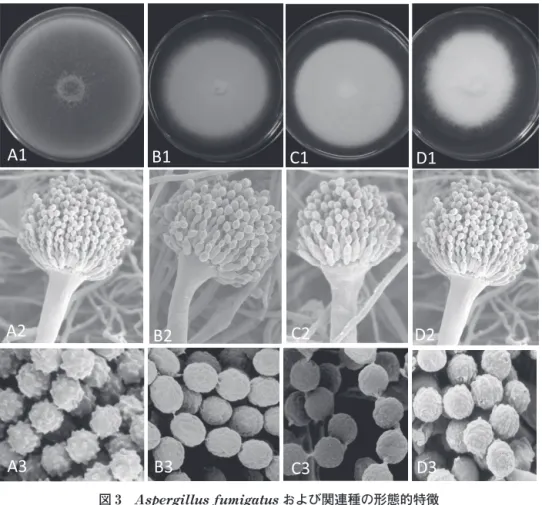
一菌種でも複数株を試験することになる.その利用例 として,新規外用爪白癬治療薬 ME1111 が,白癬菌に 対して強い抗真菌活性を有することを当センター所有 の直近 5 年間で臨床から分離された新鮮な株を使用す ることで実証し(Tabata et al., 2016),これまでに開 発されてきた抗白癬薬と異なる作用メカニズムである ことが解明された.現在,爪白癬外用治療薬として臨 床第 II 相試験が米国において進められている.同様 に,真菌選択的ミトコンドリア阻害薬 T-2307 の前臨 床試験に 114 株が使用され(Mitsuyama et al., 2008; Nishikawa et al., 2017),現在,米国において臨床第Ⅰ 相試験中である.また,新規外用抗真菌薬ルリコナ ゾールの白癬菌に対する既存外用抗真菌薬との比較試 験に 28 菌株を提供した(前田ら,2016). 病原真菌の分類学的研究 1)病原性 Aspergillus および関連菌における形態およ び系統解析による分類学的研究 深在性真菌症は増加傾向であるが,なかでも Asper-gillus 属を原因とするアスペルギルス症は致死率が高 く,治療に有効な抗真菌薬の品揃えは不十分で,その 開発が求められている.アスペルギルス症の主原因菌 は A. fumigatus である.臨床検体から分離され,ア ゾールなどの抗真菌剤に対して耐性示す A. fumigatus 関連菌種が新種 A. lentulus として報告されて以来 (Balajee et al., 2005),この菌群の分類研究が進展し (Hong et al., 2005),これまで形態的に A. fumigatus と同定された菌株の一部は,A. lentulus,A. udaga-wae などと分子系統的に別種として認識され,その薬 剤感受性の違いから原因菌の正確な同定が必要とされ ている(Balajee et al., 2004).そこで,千葉大学真菌 医学研究センターに A. fumigatus と同定された保存 菌株約 300 株を,3 遺伝子による分子系統解析,形態, 生理性状を詳細に検討した.その結果,A. fumigatus および関連菌種は 4 つの分類群(I.A. fumigatus が 属する菌群,II.A. lentulus が属する菌群,III.A. fumigatiaffinis,A. novofumigatus が 属 す る 菌 群, IV.A. viridinutans 種複合体が属する菌群)に分かれ た(Yaguchi et al., 2007a).典型的な A. fumigatus と 比較し,その 3 関連種は形態的には非常に類似し,一 般に分生子の形成が悪く,かつては分生子を形成しな い白色の A. fumigatus と考えられていたが(図 3), 分生子の表面微細構造が異なった.すなわち,A. fu-migatus の分生子が球形,刺状突起を有するのに対し て,A. lentulus は だ 円 形, ほ ぼ 滑 面,A.
viridinu-tans,A. udagawae は球形,ほぼ滑面であった(図 3).一方,生育温度は,A. fumigatus が 50℃でもよく 生育するのに対して,A. lentulus は 45℃まで,III, IV に属する菌種は 42℃まで生育し,明らかな違いが 見られた.A. viridinutans は,A. viridinutans s.s.(狭 義)のほか,A. udagawae,A. felis などに細分化され ている(Sugui et al., 2014).各種抗真菌剤に対する感受 性は,関連種においてアゾールに対して耐性を示すこと を明らかにした(図 4.Yaguchi et al., 2007a; Kikuchi et al., 2014; Tamiya et al., 2015; Lyskova et al., 2018). A. lentulus,A. udagawae,A. viridinutans の臨床 株と環境株から選択した薬剤感受性株と耐性株におい て全ゲノム配列を解析し,データベース上で公開した (https://daphne.pf.chiba-u.jp/distribution/wholege nome/WholeGenomeDownload.html).このうち,A. lentulus,A. udagawae の基準株においては,ドラフ トゲノム解析を行った(Kusuya et al., 2015, 2016).今 回得られたゲノム情報は A. fumigatus のアゾール耐 性株における既存のゲノムデータと比較することによ り,Aspergillus 菌種における薬剤耐性の機能解明に つなげることが可能である. 関連種においてアゾールに対する薬剤感受性の違い があるため,臨床上,正確な種の同定が求められてい る.そのため,リボソームタンパク質の分子量変異パ ターンから A. fumigatus を迅速かつ正確に同定・分 類する新しい手法の開発を行った.全ゲノム解読され た A. fumigatus 2 株および類縁の Neosartorya fisch-eri 1 株の計 3 株について,リボソームタンパク質の 質量分析を行うことによって,分類の基準となるリボ ソームタンパク質の正しい分子量リストを作成した. ゲノム解読株間で,一部のリボソームタンパク質に変 異が生じていることを実証し,A. fumigatus および関 連種の同定が本法によって可能であることを見出した (Nakamura et al., 2016, 2017). また,形態的には区別の困難な A. fumigatus,A. lentulus,A. udagawae の 3 菌 種 を PCR 法 を 用 い て それぞれ迅速に同定できる方法(図 4,特開 2010-115122),さらに A. fumigatus とその関連有性型で耐 熱性のある Neosartorya 属を迅速に識別できる方法 (Yaguchi et al., 2012,特開 2010-4879)を開発した. 長年にわたり多くの研究者とともに Aspergillus section fumigati における分類学的な研究を進め,本 section に属する新種 10 種を報告してきた(Yaguchi et al., 1994, 2010; Someya et al, 1999; Hubka et al., 2013, 2017; Matsuzawa et al., 2014, 2015; Novakova et
図 4 と関連種の系統関係,最高生育温度,薬剤耐性の関連と PCR 法に よる迅速同定法.レーン 1-3:Aspergillus fumigatus,レーン 4-8:A. lentulus(A. fumi-synnematus 含む),レーン 9-11:A. udagawae,レーン 12-28:その他の菌種
図 3 および関連種の形態的特徴
A: A. fumigatus.1:集落(PDA,25℃,7 日培養),2:アスペルジラ,3:分生子 B: A. lentulus.1:集落(PDA,25℃,7 日培養),2:アスペルジラ,3:分生子 C:A. udagawae.1:集落(PDA,25℃,7 日培養),2:アスペルジラ,3:分生子 D:A. viridinutans.1:集落(PDA,25℃,7 日培養),2:アスペルジラ,3:分生子
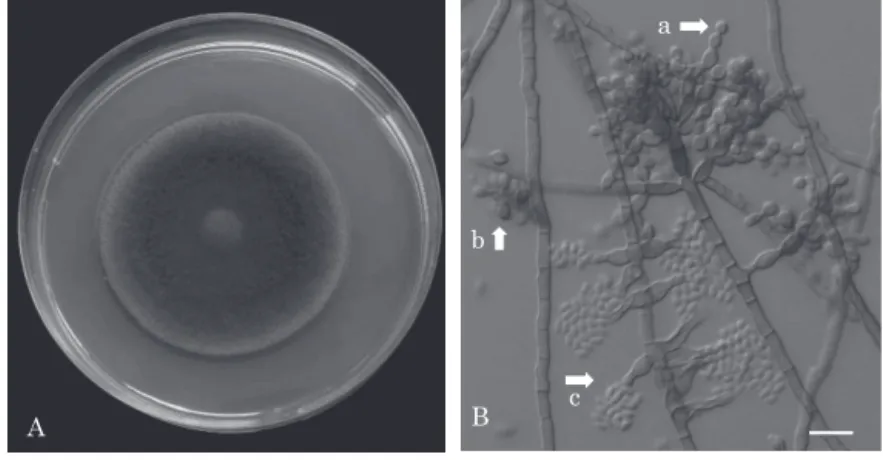
al., 2014).分類が混乱している A. viridinutans 種複 合 体 に お い て, 交 配 遺 伝 子 で あ る MAT1-1-1 と MAT1-2-1 を有する菌株を種内,種間で交配し,形成 された有性胞子を詳細に検討することにより,A. parafelis,A. pseudofelis を A. felis のシノニムとした (Hubka et al., 2018). A. fumigatus において,これまで多くの代謝産物が 報告されてきている.分類学的な知見を活用し,その 関連種においても新規で有用物質の探索を実施した. なかでも A. novofumigatus の培養液から既知化合物 とは立体化学の異なる数種のアミノ酸由来第二次代謝 産物を単離した.これらの化合物は他の Aspergillus 属からの分離例はなく,A. novofumigatus に特徴的 な化合物であったことから A. novofumigatus が他の Aspergillus 属とは異なった生合成経路を有している 可能性を示唆している(Ishikawa et al., 2010, 2011, 2016). 2)スポロトリクス症の起因菌 Sporothrix schenkii の 分類学的研究 本菌は主に皮膚およびリンパ管を侵す.小児や初老 から老人の顔面,上肢に結節や潰瘍,軽微な外傷の後 に発症する.自然界では土壌,朽木より分離されてい る.ポテト・ブドウ糖寒天(PAD)培地,25℃培養で は菌糸形(図 5A,C),1%デキストロース添加ブレイ ン・ハート・インフージョン(BHI)寒天培地,37℃ 培養では酵母形(図 5B)の 2 つの形状を取る. 代表的な深在性皮膚真菌症スポロトリクス症の起因 菌は Sporothrix schenkii 1 種と考えられていたが,分 子系統学の発展により,新たに S. brasiliensis,S. glo-bosa,S. luriei が新種として報告され,現在では S. schenckii s.s. と併せて S. schenckii 種複合体と認識さ れている(Zhang et al., 2015).当研究室に保存されて いる国内におけるスポロトリクス症起因菌 300 株の再 同定の結果,S. schenckii s.s. は 9 株,S. globosa 291 株(subgroup I 241 株,II 50 株)で(図 6),S. brasil-iensis,S. luriei は 見 出 さ れ な か っ た.S. schenckii s.s.,S. globosa の抗真菌剤に対する薬剤感受性および 生育速度の検討を実施した.S. schenckii s.s. は 37℃ で生育したが,S. globosa は 37℃での生育は認められ なかった.本邦ではスポロトリクス症に対し局所温熱 療法が有効なことが多いが,発育速度の差がかかわっ ていることが示唆された.S. globosa の一部の菌株の みイトラコナゾールに耐性が見られ,アムホテリシン B, ボ リ コ ナ ゾ ー ル に 対 し て は S. globosa,S. schenckii ともにすべての株で低感受性・耐性を示し た.S. globosa では臨床で使用されるイトラコナゾー ルに対して低感受性を示したが,subgroup I と II で は相違は認められなかった. 世界的にはスポロトリコーシスの流行が見られるた め,今後,輸入・再興感染症として注意すべきである. 3)黒色真菌症起因菌 Fonscecaea の分類学的研究 Fonsecaea pedrosoi は,クロモミコーシスの原因菌 の一つで,顔面,四肢,特に下肢に難治性の肉芽腫性 病変を起こすほか,脳を含む内臓諸臓器の病変を惹起 する.通常の培地で黒色ないし黒緑色の集落(図 7A) を形成し,Cladosporium 様の出芽型(図 7B 矢印 a), シンポジオ型(図 7B 矢印 b)およびまれにフィアロ 型分生子(図 7B 矢印 c)が見られる多形性真菌であ る.本種を含む Fonsecaea 属は,病原性黒色真菌のな かで最も重要な属である.rDNA-ITS 領域の系統解 析および RFLP 分析で本属の検討結果より,F. pe-drosoi と新規提唱の F. monophora に分け,F.
pedro-図 5
A:PAD 培地.25℃,21 日培養
B: 1%デキストロース添加 BHI 寒天培地.37℃,14 日 培養
図 7 A:PAD 培地.25℃,21 日培養
B: 顕微鏡像.矢印 a:出芽型分生子,矢印 b:シンポジオ型分生子,矢 印 c:フィアロ型分生子,スケールバー;10 μm
soi と F. compacta とに分ける伝統的な分類とは異な り,F. pedrosoi と F. compacta は同一種であるとさ れた(de Hoog et al., 2004).F. monophora は,1936 年,南アフリカにおけるクロモミコーシス原因菌 Botrytoides monophora として報告され,その後 F. pedrosoi のシノニムとされたが,形態的には F. pedro-soi にきわめて類似している. そこで当センター保存の日本産の臨床菌株を解析し たところ,すべて F. monophora であった(図 8,Ya-guchi et al., 2007b).その後,日本国内においても F. pedrosoi に属する菌株が報告された.また,上記 2 種 と形態的には類似するが,分子系統の違いによって F. nubica,F. multimorphosa が 新 種 報 告 さ れ た(Na-jafzadeh et al., 2010, 2011). 病原性の違いとして,F. pedrosoi は深在性の皮膚 のみに感染を起こし,F. monophora は脳への移行性 が高いとされているが,必ずしも明確でない. 謝 辞 この度,研究課題「病原真菌の分類学的研究と臨床 菌株コレクションの充実」で平成 30 年度日本微生物 資源学会賞を授賞いただき,大変光栄に存じます.こ れまで多くの先生方のご指導,ご鞭撻の賜物であり, また,ご推薦いただいた先生方に厚く御礼申し上げま す.中でも,千葉大学真菌医学研究センターにて病原 真菌の基礎をご教授いただいた西村和子先生(千葉大 学名誉教授),明治製菓株式会社入社以来,土壌生子嚢 菌類の分類,同定をご指導いただいた宇田川俊一先生 (元国立衛生試験所微生物部,元日本食品分析セン ター)に深く感謝申し上げます.これまで,日々の菌 株収集,保存,提供業務を通じ,常に本研究をサポー トいただいた千葉大学真菌医学研究センターバイオリ ソース管理室の皆様,共同研究を通じて本研究をご推 進いただいた先生方に心より御礼申し上げます. 文 献
Balajee, S.A., Weaver, M., Imhof, A., Gribskov, J. & Marr, K.A. 2004. Aspergillus fumigatus variant with decreased susceptibility to multiple antifungals. Antimicrob. Agents Chemother. 48: 1197-1203.
Balajee, S.A., Gribskov, J.L., Hanley, E., Nickle, D. & Marr, K. 2005. Aspergillus lentulus sp. nov., a new sibling species of A. fumigatus. Eukaryot. Cell 4: 625-632.
de Hoog, G.S., Attili-Angelis, D., Vicente, V.A., Van Den Ende, A.H. & Queiroz-Telles, F. 2004. Molecular ecology and pathogenic potential of Fonsecaea species. Med. Mycol. 42: 405-416.
Hong, S.-B., Go, S.-J., Frisvad, J.C. & Samson, R. 2005. Polyphasic taxonomy of Aspergillus fumigatus and related species. Mycologia 97: 1316-1329.
Hubka, V., Peterson, S.W., Frisvad, J.C., Yaguchi, T., Kubátová, A. & Kolarík, M. 2013. Aspergillus waksmanii sp. nov. and Aspergillus marvanovae sp. nov., two closely related species in section Fumigati described using polyphasic approach. Int. J. Syst. Evol. Microbiol. 63: 783-789.
Hubka, V., Dudová, Z., Kubátová, A., Frisvad, J.C., Yaguchi, T., Horie, Y., Jurjević, Ž., Hong, S.-B. & Kolařík, M. 2017. Taxonomic novelties in Aspergillus section Fumigati: A. tasmanicus sp. nov., induction of sexual state in A. turcosus and overview of related species. Plant. Syst. Evol. 303: 787-806.
Hubka, V., Barrs, V., Dudová, Z., Sklenář, F., Kubátová, A., Matsuzawa, A., Yaguchi, T., Horie, Y., Nováková, A., Frisvad, J.C., Talbot, J.J. & Kolařík, M. 2018. Unravelling species boundaries in the Aspergillus viridinutans complex (section Fumigati): opportunistic human and animal pathogens capable of interspecific hybridization.
Persoonia 41: 142-174.
Ishikawa, K., Hosoe, T., Itabashi, T., Wakana, D., Takizawa, K., Yaguchi, T. & Kawai, K. 2010. Novoamauromine and ent-Cycloechinulin: two new diketopiperazine derivatives from Aspergillus novofumigatus. Chem. Pharm. Bull. 58: 717-719. Ishikawa, K., Hosoe, T., Itabashi, T., Sato, F., Wachi,
H., Nagase, H., Yaguchi, T., & Kawai, K. 2011. Q u i n a z o l i n o b e n z o d i a z e p i n e d e r i v a t i v e s , novobenzomalvins A-C, fibronectin expression regulators from Aspergillus novofumigatus. Sci. Pharm. 79: 937-950.
Ishikawa, K., Sato, F., Itabashi, T., Wachi, H., Takeda, H., Wakana, D., Yaguchi, T., Kawai, K. & Hosoe, T. 2016. Asnovolins A-G, spiromeroterpenoids isolated from the fungus Aspergillus novo f umigatus, and suppression of fibronectin expression by asnovolin E. J. Nat. Prod. 79: 2167-2174.
Kikuchi, K., Watanabe, A., Ito, J., Oku, Y., Wuren, T., Taguchi, H., Yarita, K., Muraosa, Y., Yahiro, M., Yaguchi, T. & Kamei, K. 2014. Antifungal susceptibility of Aspergillus fumigatus clinical isolates collected from various areas in Japan. J. Infect. Chemother. 20: 336-338. 公開特許公報.特開 2010-115122.アスペルギルスフ ミガタス類縁菌の検出方法. 公開特許公報.特開 2010-4879.ネオサルトリア属に 属する菌類及びアスペルギルスフミガタスの検出方 法.
Kusuya, Y., Takahashi-Nakaguchi, A., Takahashi, H. & Yaguchi, T. 2015. Draft genome sequence of the pathogenic filamentous fungus Aspergillus udagawae strain IFM 46973T. Genome Announc. 3: pii: e00834-15.
Kusuya, Y., Sakai, K., Kamei, K., Takahashi, H. & Yaguchi, T. 2016. Draft genome sequence of the pathogenic filamentous fungus Aspergillus lentulus IFM 54703T. Genome Announc. 4: e01568-15. Lyskova, P., Hubka, V., Svobodova, L., Barrs, V.,
Dhand, N., Yaguchi, T., Matsuzawa, T., Horie, Y., Kolarik, M., Dobias, R. & Hamal, P. 2018. Antifungal susceptibility of the Aspergillus viridinutans complex: comparison of two in vitro methods. Antimicrob. Agents Chemother. 62: e01927-17. 前田 潤,南條育子,古賀裕康,栂 哲夫,槇村浩一, 坪井良治 2016.ルリコナゾールの白癬菌に対する in vitro 抗真菌活性─ MIC および MFC の同時測定 による既存外用抗真菌薬との比較試験─.日本医真 菌学会雑誌 57:J1-J6.
Matsuzawa, T., Horie, Y., Abliz, P., Gonoi, T. & Yaguchi, T. 2014. Aspergillus huiyaniae sp. nov., a teleomorphic species in sect. Fumigati isolated from desert soil in China. Mycoscience 55: 213-220.
Matsuzawa, T., Takaki, G.M.C., Yaguchi, T., Okada, K., Abliz, P., Gonoi, T. & Horie, Y. 2015. Aspergillus arcoverdensis, a new species of Aspergillus section Fumigati isolated from caatinga soil in State of Pernambuco, Brazil. Mycoscience 56: 123-131. Mitsuyama, J., Nomura, N., Hashimoto, K., Yamada,
E., Nishikawa, H., Kaeriyama, M., Kimura, A., Todo, T. & Narita, H. 2008. In vitro and in vivo antifungal activity of T-2307: a novel arylamidine. Antimicrob. Agents Chemother. 52: 1318-1324. Najafzadeh, M.J., Sun, J., Vicente, V., Xi, L., van den
Ende, A.H. & de Hoog, G.S. 2010. Fonsecaea nubica sp. nov, a new agent of human chromoblastomycosis revealed using molecular data. Med. Mycol. 48: 800-806.
Najafzadeh, M.J., Vicente, V.A., Sun, J., Meis, J.F. & de Hoog, G.S. 2011. Fonsecaea multimorphosa sp. nov, a new species of Chaetothyriales isolated from a feline cerebral abscess. Fun. Biol. 115: 1066-1076. Nakamura, S., Sato, H., Tanaka, R. & Yaguchi, T.
2016. Verification of ribosomal proteins of Aspergillus fumigatus for the use of biomarkers in MALDI-TOF MS identification. Mass Spectromet. 5: A0049.
Nakamura, S., Sato, H., Tanaka, R. Kusuya, Y., Takahashi, H. & Yaguchi, T. 2017. Ribosomal subunit protein typing using matrix-assisted laser desorption ionization time-of-flight mass s p e c t r o m e t r y ( MA LD I - T O F M S ) f o r t h e identification and discrimination of Aspergillus species. BMC Microbiol. 17: 100.
Nishikawa, H., Fukuda, Y., Mitsuyama, J., Tashiro, M., Tanaka, A., Takazono, T., Saijo, T., Yamamoto, K., Nakamura, S., Imamura, Y., Miyazaki, T., Kakeya, H., Yamamoto, Y., Yanagihara, K., Mukae,
H., Kohno, S. & Izumikawa, K. 2017. In vitro and in vivo antifungal activities of T-2307, a novel arylamidine, against Cryptococcus gattii: an emerging fungal pathogen. J. Antimicrob. Chemother. 72: 1709-1713.
Novakova, A., Hubka, V., Dudova, Z., Matsuzawa, T., Kubatova, A., Yaguchi, T. & Kolarik, M. 2014. New species in Aspergillus section Fumigati from reclamation sites in Wyoming (U.S.A.) and revision of A. viridinutans complex. Fung. Divers. 64: 253-274.
Someya, A., Yaguchi, T. & Udagawa, S. 1999. Neosartorya sublevispora, a new species soil-borne Eurotiales. Mycoscience 40: 405-409.
Sugui, J.A., Peterson, S.W., Figat, A., Hansen, B., Samson, R.A., Mellado, E., Cuenca-Estrella, M. & Kwon-Chung, K.J. 2014. Genetic relatedness versus biological compatibility between Aspergillus fumigatus and related species. J. Clin. Microbiol. 52: 3707-3721.
Suzuki, R., Yikelamu, A., Tanaka, R., Igawa, K., Yokozeki, H. & Yaguchi, T. 2016. Studies in phylogeny, development of rapid identification, antifungal susceptibility and growth rates on clinical strains of Sporothrix schenckii complex in Japan. Med. Mycol. J. 57: E47-E57.
Tabata, Y., Takei-Masuda, N., Kubota, N., Takahata, S., Ohyama, M., Kaneda, K., Iida, M. & Maebashi, K. 2016. Characterization of antifungal activity and nail penetration of ME1111, a new antifungal agent for topical treatment of onychomycosis. Antimicrob. Agents Chemother. 60: 1035-1039. Tamiya, H., Ochiai, E., Kikuchi, K., Yahiro, M.,
Toyotome, T., Watanabe, A., Yaguchi, T. & Kamei,
K. 2015. Secondary metabolite profiles and antifungal drug susceptibility of Aspergillus fumigatus and closely related species, A. lentulus, A. udagawae, and A. viridinutans. J. Infect. Chemother. 21: 385-391.
Yaguchi, T., Someya, A., Miyadoh, S. & Udagawa, S. 1994. A new species of Neosartorya from Taiwan soil. Mycoscience 35: 309-313.
Yaguchi, T., Horie, H., Tanaka, R., Matsuzawa, T., Ito, J. & Nishimura, K. 2007a. Molecular phylogenetics of multiple genes on Aspergillus section Fumigati isolated from clinical specimens in Japan. Jpn. J. Med. Mycol. 48: 37-46.
Yaguchi, T., Tanaka, R., Nishimura, K. & Udagawa, S. 2007b. Molecular phylogenetics of strains morphologically identified as Fonsecaea pedrosoi from clinical specimens. Mycoses 49: 255-260. Yaguchi, T., Matsuzawa, T., Tanaka, R., Abliz, P.,
Hui, Y. & Horie, Y. 2010. Two new species of Neosartorya isolated from soil in Xinjiang, China. Mycoscience 51: 253-262.
Yaguchi, T., Imanishi, Y., Matsuzawa, T., Hosoya, K., Hitomi, J. & Nakayama, M. 2012. Method for identifying heat-resistant fungi of the genus Neosartorya. J. Food Prot. 75: 1806-1813.
Zhang, Y., Hagen, F., Stielow, B., Rodrigues, A.M., Samerpitak, K., Zhou, X., Feng, P., Yang, L., Chen, M., Deng, S., Li, S., Liao, W., Li, R., Li, F., Meis, J.F., Guarro, J., Teixeira, M., Al-Zahrani, H.S., Pires de Camargo, Z., Zhang, L. & de Hoog, G.S. 2015. Phylogeography and evolutionary patterns in Sporothrix spanning more than 14 000 human and animal case reports. Persoonia 35: 1-20.