昆虫の宇ノ木‐水波学習モデルの2次条件付けを用
いた検証
著者
森 康博
平成19 年度 修士論文
昆虫の宇ノ木‐水波学習モデルの
2 次条件付けを用いた検証
専攻:生命機能科学専攻
学籍番号:
A6BM2020
氏名:森 康博
目次 Ⅰ 要旨...2 Ⅱ 序論...3 Ⅲ 材料と方法...6 1 材料 ...6 2 行動実験 ...6 2-1 2 次条件付け ...6 2-3 視覚嗜好性テスト ...6 3 オクトパミン受容体阻害剤(エピナスチン)の投与 ...7 4 実験スケジュール...7 4-1 2 次条件付け実験...7 4-2 第 1 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 ...7 4-3 第 2 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 ...7 4-4 記憶の読み出し段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 ...8 5 統計解析 ...8 Ⅳ 結果...9 1 2 次条件付け実験...9 2 第 1 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験...9 3 第 2 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験...9 4 記憶の読み出し段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 ...9 Ⅴ 考察... 11 1 2 次条件付けの成立 ... 11 2 2 次条件付けの各段階におけるオクトパミン受容体阻害剤の効果... 11 3 宇ノ木-水波モデル... 11 Ⅵ 引用文献... 13 Ⅶ 図の説明... 15
Ⅰ 要旨
古典的条件付けとは、水などの無条件刺激(unconditioned stimulus; US)と中性的な刺 激(条件刺激、conditioning stimulus; CS)とが対提示されることで、CS が条件反応 (conditioned response; CR)を誘発するようになる現象である。2 次条件付けとは、古典 的条件付けにより条件反応を誘発するようになった条件刺激(CS1)を別の中性的な刺激 (CS2)と対提示すると、CS2 も条件反応を誘発するようになる現象である。 動物の学習にとってアミン作動性ニューロンの働きは重要である。いくつかの種の昆虫 において、オクトパミン作動性ニューロンは報酬学習の、ドーパミン作動性ニューロンは 罰学習の成立にそれぞれ必要であることが知られている。 宇ノ木と水波は、フタホシコオロギにおいて、オクトパミン受容体阻害剤によって報酬 記憶の読み出しが、ドーパミン受容体阻害剤によって罰記憶の読み出しが阻害される事を 発見した。この現象は従来の学習モデルでは説明できないので、宇ノ木と水波は新しい学 習モデルを提案した。このモデルから、報酬条件付けの際にオクトパミン受容体阻害剤の 影響下にあったとしても CS ニューロンとオクトパミン作動性ニューロンの同時活性化に よってCS ニューロンからオクトパミン作動性ニューロンへのシナプスは強化され、条件付 け後、条件刺激の提示によってオクトパミン作動性ニューロンを活性化させることが予測 できる。つまり、報酬2 次条件付けにおいて、2 次条件付けの第 1 段階で、オクトパミン受 容体阻害剤で報酬学習の成立を阻害しても 2 次条件付けが成立することが予想される。そ こで、報酬2 次条件付けの系を開発し、宇ノ木-水波モデルを検証した。 2 次条件付けの第 1 段階では、嗅覚刺激(パイナップルの匂い、CS1)の提示開始 2 秒後に 水(US)を与えた。第 2 段階では、嗅覚刺激(CS1)と視覚刺激(視覚パターン、CS2)を同時に 提示した。条件付けの前と後で視覚パターンに対する嗜好性のテストを行った。2 次条件付 け後、第 2 段階で使用した視覚パターンに対する嗜好性は訓練前に比べて有意に高くなっ た。第1 段階あるいは第 2 段階で刺激の対提示を行わなかった対照群では嗜好性に変化は なかった。この結果から、2 次条件付けがコオロギで起こることが確認された。このような 嗅覚-視覚連合型の 2 次条件付けは無脊椎動物では初めてである。 次に2 次条件付けの第 1 段階、第 2 段階、または記憶の読み出し段階でオクトパミン受 容体阻害剤であるエピナスチンをコオロギの頭部の血リンパ中に投与し、その影響を調べ た。第1 段階でエピナスチンを投与しても、予想通り 2 次条件付けは成立した。第 2 段階 でエピナスチンを投与すると 2 次条件付けは成立せず、エピナスチンの代わりに生理食塩 水を投与した対照群では 2 次条件付けは成立した。また、オクトパミン受容体阻害剤の投 与により記憶の読み出しが阻害された。これらの結果は宇ノ木-水波モデルを支持している。 宇ノ木-水波モデルは、記憶の獲得には CS と US の関係の内的表現の形成が伴い、記憶 の読み出しにはUS の想起が伴うことを示唆している。これは哺乳類における古典的条件付 けの認知的な説明と合致している。宇ノ木-水波モデルは、古典的条件付けを裏打ちするシ ステムが、今まで考えられてきた以上に哺乳類と昆虫とで似ていることを示唆している。
Ⅱ 序論 学習とは、動物が経験によって行動を変化させることである。地球上で動物は学習によ ってさまざまな環境に適応して生活している。その学習を裏打ちするシステムとはどのよ うなものだろうか。昆虫の脳のニューロンは10 万から 100 万個と、哺乳類と比べて非常に 少ない。そのため、神経回路が比較的少数のニューロンで構成されており、神経回路の働 きが個々のニューロンの性質、関係性まで還元して調べることが比較的容易である。また、 昆虫と哺乳類の神経系では似ている部分も多く、たとえば、昆虫と哺乳類の両方で、アミ ン作動性ニューロンは学習に重要な働きをしている(Hammer, 1993; Hammer and Menzel, 1998; Farooqui et al.,2003; Schwaerzel et al., 2003; Riemensperger et al., 2005; Schroll et al., 2006; Vergoz et al., 2007; Wise, 2004; Schultz, 2006)。コオロギではオクトパミン (OA)作動性ニューロンが報酬学習に、ドーパミン(DA)作動性ニューロンが罰学習に関与し ている (Unoki et al., 2005, 2006)。 宇ノ木がオクトパミン受容体阻害剤またはドーパミン受容体阻害剤の報酬学習または罰 学習の記憶の読み出しに対する効果をしらべたところ、オクトパミン受容体阻害剤は視覚 学習、嗅覚学習ともに報酬学習でのみ記憶の読み出しを阻害し、ドーパミン受容体阻害剤 は視覚学習、嗅覚学習ともに罰学習でのみ記憶の読み出しを阻害した(Unoki 博士論文, 2007)。つまりオクトパミン作動性ニューロンまたはドーパミン作動性ニューロンは報酬ま たは罰記憶の読み出しにも必要であることがわかった。 この現象はシュワルツェルらによる学習モデル(Schwaerzel et al., 2003)などの従来の昆 虫の学習モデルでは説明できない(Fig.1A)。シュワルツェルらのモデルにおいて、無条件刺 激(unconditioned stimulus; US)のシグナルを伝達するオクトパミン作動性ニューロンま たはドーパミン作動性ニューロンと、条件刺激(conditioning stimulus; CS)のシグナルを伝 達するCS ニューロンは条件反応(conditioned response; CR)を引き起こす CR ニューロン にシナプス結合している。条件付け前、オクトパミン作動性ニューロンまたはドーパミン 作動性ニューロンからCS ニューロンのシナプスへのシナプスは強く、CS ニューロンから CR ニューロンへのシナプスは沈黙しているか、弱い。条件刺激と無条件刺激の対提示時に おける CS ニューロンとオクトパミン作動性ニューロンまたはドーパミン作動性ニューロ ンの同時活性化によってCS ニューロンから CR ニューロンへのシナプスは強化される。条 件付け後の条件刺激の提示はCS ニューロンの活性化を引き起こし、続いて CR ニューロン を活性化させる。このモデルの場合オクトパミン受容体またはドーパミン受容体が阻害さ れていても記憶の読み出しには影響は出ないはずである。 そこで、宇ノ木と水波はこの新しい発見を説明できる宇ノ木-水波モデルを提案した (Fig.1B)。宇ノ木-水波モデルの骨子は CS ニューロンからオクトパミン作動性ニューロンま たはドーパミン作動性ニューロンへ接続するシナプスと、CR ニューロンを活性化させるの に CS ニューロンとオクトパミン作動性ニューロンまたはドーパミン作動性ニューロンの 両方の活性化を必要とするAND ゲートの仮定である。報酬学習の場合、条件付け前、オク
トパミン作動性ニューロンからCS ニューロンへのシナプスは強く、CS ニューロンから CR ニューロンへのシナプス、CS ニューロンからオクトパミン作動性ニューロンへのシナプス は沈黙しているか、弱い。条件刺激と無条件刺激の対提示時におけるCS ニューロンとオク トパミン作動性ニューロンの同時活性化によってCS ニューロンから CR ニューロンへのシ ナプスとCS ニューロンからオクトパミン作動性ニューロンへのシナプスは強化される。条 件付け後の条件刺激の提示はCS ニューロンの活性化を引き起こし、続いてオクトパミン作 動性ニューロンの活性化を引き起こす。この2 つのニューロンの活性化によって AND ゲー トが開き、CR ニューロンが活性化する。罰学習の場合も同様に、条件付けによって、CS ニューロンからCR ニューロンへのシナプスと CS ニューロンからドーパミン作動性ニュー ロンへのシナプスが強化される。条件付け後の条件刺激の提示はCS ニューロンの活性化を 引き起こし、続いてドーパミン作動性ニューロンの活性化を引き起こし、この 2 つのニュ ーロンの活性化によってAND ゲートが開き、CR ニューロンが活性化する。つまり、宇ノ 木-水波モデルでは、報酬条件付けによって CS ニューロンからオクトパミン作動性ニュー ロンまたはドーパミン作動性ニューロンへの経路と、CS ニューロンから CR ニューロンへ の経路という 2 つのシナプス経路が増強され、この両経路の活性化が条件反応の発生に必 要である。すなわち、宇ノ木-水波モデルの場合オクトパミン受容体またはドーパミン受容 体が阻害されていると、AND ゲートが開かず、記憶の読み出しができなくなる。 宇ノ木-水波モデルを検証するにはどうすればよいだろうか。このモデルからある現象が 予測できる。2 次条件付けにおいて、第 1 段階の際に、オクトパミン受容体阻害剤によって 報酬学習の、ドーパミン受容体阻害剤によって罰学習の成立を阻害しても、2 次条件付けが 成立するという現象である(Fig.1C)。報酬 2 次条件付けの場合、第 1 段階において条件刺激 (CS1)と無条件刺激の対提示時における CS1 ニューロンとオクトパミン作動性ニューロン の同時活性化によってCS1 ニューロンからオクトパミン作動性ニューロンへのシナプスは 強化される。しかし、オクトパミン受容体阻害剤によって、オクトパミン作動性ニューロ ンからのシナプス伝達は阻害されるので、CS1 ニューロンから CR ニューロンへのシナプ スは強化されない。この状態でCS1 を提示しても CR は起きない。続いて第 2 段階におい て、新しい条件刺激(CS2)と CS1 の対提示時に CS1 ニューロンの活性化によって、オクト パミン作動性ニューロンが活性化する。CS2 ニューロンと CR ニューロンの同時活性化に よってCS2 ニューロンから CR ニューロンへのシナプスと CS2 ニューロンからオクトパミ ン作動性ニューロンへのシナプスは強化される。条件付け後のCS2 の提示は CS2 ニューロ ンの活性化を引き起こし、続いてオクトパミン作動性ニューロンの活性化を引き起こす。 この2 つのニューロンの活性化によって AND ゲートが開き、CR ニューロンが活性化し、 条件反応が起きる。また、罰2 次条件付けの場合も同様に第一段階において CS1 ニューロ ンからドーパミン作動性ニューロンへのシナプスは強化されるため、第2 段階において CS2 ニューロンからCR ニューロンへのシナプスと CS2 ニューロンからドーパミン作動性ニュ ーロンへのシナプスは強化され、2 次条件付けは成立する。
この現象が実際に確認できるかを確かめるために、コオロギで 2 次条件付けの方法を確 立した。昆虫の2 次条件付けは、ミツバチ(Bitterman and Menzel, 1983; Hussainin et al., 2007)やショウジョウバエ(Brembs and Heisenberg, 2001)で報告されている。それらは嗅 覚のみ、あるいは視覚のみを用いた 2 次条件付けである。このように刺激の様相が同じ 2 次条件付けの場合、刺激の汎化(generalization)が問題となる。フタホシコオロギは嗅覚学 習と視覚学習ができることがわかっている(Unoki et al., 2005, 2006)。そこで、汎化の問題 を回避するために、条件刺激として嗅覚刺激と視覚刺激を用いた2 次条件付けを行った。 本研究では、第 1 に、無脊椎動物で初めて条件刺激のモダリティの異なる 2 次条件付け を成立させ、第2 に、2 次条件付けと薬理学的手法を用いて宇ノ木-水波学習モデルを検証 した。
Ⅲ 材料と方法 1 材料 本研究では成虫脱皮後1 週間程度のフタホシコオロギ Gryllus bimaculatus のオスを 用いた。室温26~28℃、明暗周期(明期:暗期=12 時間:12 時間)下で飼育した。水へ のモティベーションを高めるため、実験前の3 日間水を与えなかった。 2 行動実験
実験方法はこれまでの研究を参考に設定した(Unoki et al., 2005, 2006; Matsumoto et al., 2006)。
2-1 2 次条件付け
条件刺激1(conditioning stimuli 1; CS1)として匂いを、条件刺激 2(conditioning stimuli 2; CS2)として視覚パターンを、無条件刺激(unconditioned stimuli; US)として水を使った。 渇水状態のコオロギにとって水は報酬となる。 2 次条件付けの第 1 段階で、パイナップルの匂い(CS1)と水(US)を対提示する嗅覚条件付 けを行った(Fig.2A)。パイナップルエッセンスを 3mm 四方のろ紙片に 1 滴たらし、ろ紙片 を水の入った1ml 注射器の針の先端から 1cm の位置に取り付けた(Fig.2B)。コオロギの触 角にろ紙を近づけて匂いをかがせ始めた2 秒後にコオロギの口に水を 1 滴与えた。条件付 けの後に、匂いを掃除機で吸い取った。 2 次条件付けの第 2 段階で、視覚パターン(CS2)と匂い(CS1)を対提示した(Fig.2A)。視覚 パターンとして白中心黒周辺、あるいは黒中心白周辺パターンを用いた(Fig.3A)。視覚パタ ーンを注射器の針の根元に取り付け、パイナップルエッセンスをしみこませたろ紙片を針 の先端から 1cm の位置に取り付けた(Fig.2B)。注射針の先で触角を触り、上に注意を向け させてから視覚パターンと匂いを同時に 5 秒間提示した。条件付けの後に、匂いを掃除機 で吸い取った。 2-3 視覚嗜好性テスト 視覚嗜好性テスト装置は、2 つの待機室、スライド式扉、テスト室、スライド式の壁に載 った3 つの視覚パターンからなる(Fig.3B)。2 つの白中心パターンと 1 つの黒中心パターン が灰色のスライド式の壁に載っていて、同時に 2 つが提示される。視覚パターンの中心に は小さな穴が開いていて、ぬれたろ紙の入ったプラスチック容器に繋がっている。コオロ ギは空気中の水分を感じ取ることができ、視覚パターンを探索するモティベーションとな る。 嗜好性テストではコオロギにテスト室の中を 4 分間自由に探索させて行った。テストの 前に各コオロギを待機室へ移し、4 分間周囲の環境に慣れさせた。その後、待機室をテスト 室への入り口までずらし、スライド式扉を上げた。コオロギがテスト室に入ったら扉を下
ろし、テストを開始した。テスト開始から 2 分後スライド式の壁をずらし、視覚パターン の位置を逆転させた。コオロギが視覚パターンに口器をつけ、動かしている時間をパター ンへの探索時間として計った。総探索時間が10 秒未満の場合、パターン探索へのモティベ ーションが低いとみなし、データ解析からははずした。その割合は 30%未満だった。嗜好 性テストは条件付けの前後1 回ずつ行った。 3 オクトパミン受容体阻害剤(エピナスチン)の投与 報酬 2 次条件付けにおいてオクトパミン作動性ニューロンがどのように関与しているの かを調べるために、エピナスチン(Epinastine)をコオロギの血リンパ中に投与した。コオロ ギの頭部(中央単眼)に注射針で穴を開け、クチクラと脳の間の血リンパ中に 1μM のエピナ スチンを含む生理食塩水(NaCl 150mM, KCl 9mM, CaCl2-H2O 5mM, NaHCO3 2mM,
glucose 40mM, NaOH で pH7.2 に調整)を 3μl 投与した。エピナスチンの投与量は 3pmol、 コオロギの体重1g あたり約 1.01ng となる。対照実験では、生理食塩水を 3μl 投与した。 エピナスチンは日本ベーリンガーインゲルハイム株式会社から提供を受けた。 4 実験スケジュール 4-1 2 次条件付け実験 2 次条件付けは第 1 段階を訓練回数 4 回、訓練間隔 5 分間で行った 1 日後に第 2 段階を 訓練回数4 回、訓練間隔 5 分間で行った。訓練前と訓練 1 時間後に嗜好性テストを行った (Fig.2D)。対照群では 2 次条件付けの第 1 段階または第 2 段階の訓練を不対合試行(unpaired trials)にした。不対合対照群では US、CS1 または CS2 を 150 秒間隔で偽ランダム化して 与 え た 。 実 験 群 を P/P(paired/paired) 群 、 第 1 段 階 を 不 対 合 試 行 に し た 対 照 群 を UP/P(unpaired/paired)群、第 2 段階を不対合試行にした対照群を P/UP(paired/unpaired) 群と呼ぶ。 4-2 第 1 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 第1 段階の開始 30 分前に上記の方法でエピナスチンを投与した。2 次条件付けは第 1 段 階を訓練回数4 回、訓練間隔 5 分間で行った 1 日後に第 2 段階を訓練回数 4 回、訓練間隔 5 分間で行った。訓練前と訓練 1 時間後に視覚嗜好性テストを行った。 4-3 第 2 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 第2 段階の開始 30 分前にエピナスチンを投与した。対照実験では生理食塩水を投与した。 2 次条件付けは第 1 段階を訓練回数 4 回、訓練間隔 5 分間で行った 1 時間後に第 2 段階を 訓練回数6 回、訓練間隔 5 分間で行った。訓練前と訓練 1 日後に視覚嗜好性テストを行っ た。また、非投与群でも同様のスケジュールで2 次条件付けを行った。
4-4 記憶の読み出し段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 訓練後の視覚嗜好性テスト30 分前にエピナスチンを投与した。対照実験では生理食塩水 を投与した。また、エピナスチン投与後、効果が切れた後に視覚嗜好性テストを行う対照 実験も行った。2 次条件付けは第 1 段階を訓練回数 4 回、訓練間隔 5 分間で行った 1 時間 後に第2 段階を訓練回数 6 回、訓練間隔 5 分間で行った。訓練前と訓練 1 時間後または 1 日後に視覚嗜好性テストを行った。 5 統計解析 パターンに口器をつけている時間を探索時間とし、総探索時間のうち条件付けした方の パターンへの探索時間の割合を算出し、100 をかけた値を Preference Index(PI)とした (Fig.3C)。PI が 50 のとき、パターンに対する嗜好性に差はない。同 1 個体群内の比較には ウィルコクソン検定(WCX)、異なる個体群間の比較にはマンホイットニーの U 検定(M-W) を用いた。
Ⅳ 結果
1 2 次条件付け実験
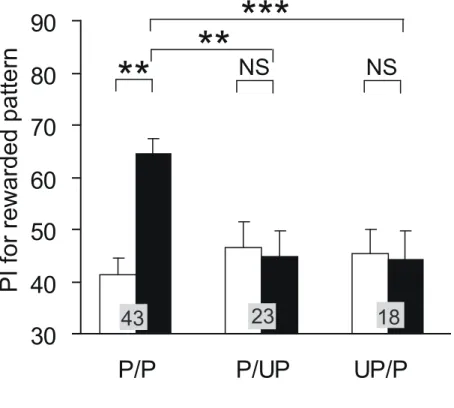
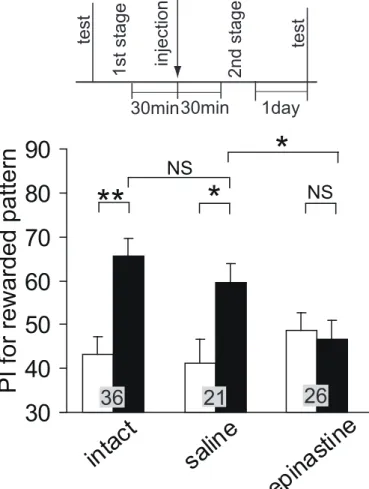
P/P 群において、視覚嗜好性は訓練前と訓練後とで有意な差があり、2 次条件付けの効果 が見られた(Fig.4)。不対合対照群では UP/P 群、P/UP 群ともに条件付け効果は見られなか った。訓練前と比べてP/P 群では WCX p<0.001、UP/P 群、P/UP 群では WCX p≧0.05 だ った。また、P/P 群の訓練後の視覚嗜好性は UP/P 群あるいは P/UP 群に比べそれぞれ有意 に高かった。P/P 群と P/UP 群で M-W p<0.01、P/P 群 と UP/P 群で M-W p<0.001 だった。 2 第 1 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 第 1 段階においてエピナスチン投与によってオクトパミン受容体を阻害したコオロギで は、視覚嗜好性は訓練前と訓練後とで有意な差があり、条件付け効果が見られた(Fig.5)。 訓練前と比べてWCX p<0.001 だった。 3 第 2 段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 第 2 段階においてエピナスチン投与によってオクトパミン受容体を阻害した実験群では 訓練後の視覚嗜好性には訓練前に比べて有意な差はなく、条件付け効果は見られなかった (Fig.6)。生理食塩水を投与した対照群及び非投与群では条件付け効果が見られた。訓練前 と比べて実験群ではWCX p≧0.05、対照群では WCX p<0.05、非投与群では WCX p<0.01 だった。実験群の訓練後の条件付けをした視覚パターンへの嗜好性は対照群に比べ有意に 低く、また、対照群の訓練後の視覚嗜好性は非投与群のものと有意な差はなく、条件付け 効果に差はなかった。実験群と対照群でM-W p<0.05、対照群と非投与群で M-W p≧0.05 だった。 4 記憶の読み出し段階におけるオクトパミン受容体阻害剤(エピナスチン)投与実験 訓練後の視覚嗜好性テストの30 分前にエピナスチンを投与してオクトパミン受容体を阻 害した実験群の視覚嗜好性は訓練前後で有意差がなく、条件付け効果は見られなかった (Fig.7)。生理食塩水を投与した対照群では条件付け効果が見られた。訓練後の実験群の条 件付けをした視覚パターンへの嗜好性は生理食塩水を投与した対照群に比べ有意に低かっ た。この結果だけでは記憶の読み出しが阻害されているのか、記憶が消えているのか判断 できないので、訓練後、エピナスチンを投与し、エピナスチンの効果が切れた 1 日後に視 覚嗜好性テストをする対照実験を行った。エピナスチン投与から 1 日後の条件付けをした 視覚パターンへの嗜好性は訓練前より有意に高く、投与の30 分後にテストをした実験群と 比べても有意に高かった。このことから、実験群では条件付けの記憶の読み出しが阻害さ れているだけで、記憶が消えているわけではないといえる。訓練前と比べて実験群では WCX p≧0.05、生理食塩水対照群では WCX p<0.01、1 日後対照群では WCX p<0.01 だっ た。訓練後の視覚嗜好性は実験群と生理食塩水対照群でM-W p<0.01、実験群と 1 日後対照
Ⅴ 考察
1 2 次条件付けの成立
昆虫の2 次条件付けはミツバチ(Bitterman and Menzel, 1983; Hussaini et al., 2007)と ショウジョウバエ(Brembs and Heisenberg, 2001)において報告されている。しかし、とも に嗅覚刺激と嗅覚刺激、視覚刺激と視覚刺激など同種の刺激を使ったものだった。コオロ ギは嗅覚学習と視覚学習のどちらもできることがわかっているのでCS1 として匂い刺激を、 CS2 として視覚パターンを利用した。この条件付けでは CS1 と CS2 で刺激の種類が違うこ とから、汎化の問題が退けられる。 UP/P 群つまり、第 1 段階においてパイナップルの匂いと水の不対提示を受けたコオロギ は第 2 段階で視覚パターンとパイナップルの匂いの対提示を受けても訓練前後で視覚パタ ーンの嗜好性に有意差がなかった。したがって、P/P 群で見られる嗜好性の変化はパイナッ プルの匂いに対する生得的嗜好性が視覚パターンの嗜好性に移行したということでは説明 できない。したがって、この嗜好性の変化は2 次条件付けの効果によるものと判断できる。 このような、条件刺激のモダリティが異なる2 次条件付けは無脊椎動物では初めてである。 2 2 次条件付けの各段階におけるオクトパミン受容体阻害剤の効果 第 1 段階でオクトパミン受容体阻害剤を投与したコオロギで 2 次条件付けが成立した。 これは宇ノ木-水波モデルの予測と一致する。つまり、第 1 段階でオクトパミン受容体阻害 剤が投与されても CS 経路からオクトパミン作動性ニューロンに入力するシナプスは増強 されるので、第2 段階においてオクトパミン作動性ニューロンを活性化させることができ、 2 次条件付けが成立する。 第 2 段階でオクトパミン受容体阻害剤を投与したコオロギでは 2 次条件付けは成立しな かった。これは、CS2 ニューロンから CR ニューロンへのシナプスが増強されなかったた めだと説明できる。このことから、2 次条件付けもオクトパミン作動性報酬系の働きにより 起こっているとのである。また、この場合もCS2 経路からオクトパミン作動性ニューロン に入力するシナプスは増強されていると考えられるため、この状態からCS2 と別の条件刺 激(CS3)を対提示すれば、3 次条件付けが成立する可能性がある。 オクトパミン受容体阻害剤によって記憶の読み出しが阻害された。このことから、2 次条 件付けの記憶の読み出しも宇ノ木-水波モデルで説明できる。 これらの結果から、報酬 2 次条件付けは宇ノ木-水波モデルに従ったオクトパミン作動性 報酬系の働きで起こっていることが示唆される。今後、罰2 次条件付けが宇ノ木-水波モデ ルに従ってドーパミン作動性罰系の働きによって起こるかについての研究が必要である。 3 宇ノ木-水波モデル 宇ノ木-水波モデルにおいて、CS ニューロンから CR ニューロンへのシナプス以外にも、 CS ニューロンからオクトパミン作動性ニューロンへのシナプスも条件付けの際に増強され
る。これは昆虫の条件付けには条件刺激と無条件刺激の関係、つまり、感覚刺激と感覚刺 激の関係の内的表現の形成が伴うことを意味する。また、記憶の読み出しの際、条件刺激 はCS 経路の活性化とともにオクトパミン作動性ニューロンの活性化を引き起こし、それに よって条件反応が起きる。これは昆虫が学習した記憶の読み出しには無条件刺激の想起が 伴うことを意味している。これらは哺乳類における古典的条件付けの認知的な説明に合致 する(Rescorla, 1988; Holland, 1993)。宇ノ木-水波モデルは古典的条件付けを裏打ちするシ ステムが今まで考えられてきた以上に哺乳類と昆虫とでよく似ていることを示唆する。 宇ノ木-水波モデルによれば 2 次条件付けにおける CS1 と CS2 の連合にはオクトパミン 作動性ニューロンまたはドーパミン作動性ニューロンの活性化が必要である。では、強化 されていないCS1 と CS2 の間で連合は起こるのであろうか。ショウジョウバエや、ミツバ チでは、CS1 と CS2 を対提示した後 CS1 と無条件刺激を対提示すると、CS2 によって条 件反応が引き起こされるという、感性予備条件付けが起こることが知られている(Müller et al., 2000; Brembs and Heisenberg,2001)。もし、コオロギでも感性予備条件付けが成立す るなら、CS1 ニューロンと CS2 ニューロンで互いにシナプス結合している可能性がある。 つまり、条件刺激と無条件刺激の間の関係が内的に表現されるように条件刺激間の関係を 内的に表現するようなシナプスも存在するということである。このように、今後、条件付 けのシステムを明らかにするための様々な実験を行うことで、宇ノ木-水波モデルが更に改 良されることが期待できる。
Ⅵ 引用文献
Bitterman, M. E., Menzel, R., Classical conditioning of proboscis extension in honeybees (Apis mellifera)., J. Comp. Phychol. 97, 107-119 (1983).
Brembs, B., Heisenberg, M., Conditioning with compound stimuli in Drosophila melanogaster in the flight simulator., J. Exp. Biol. 204, 2849-2859. (2001)
Farooqui, T., Robinson, K., Vaessin, H. & Smith, B. H., Modulation of early olfactory processing by an octopaminergic reinforcement pathway in the honeybee., J. Neurosci. 23, 5370-5380 (2003).
Hammer, M., An identified neuron mediates the unconditioned stimulus in associative olfactory learning in honeybees., Nature 366, 59-63 (1993).
Hammer, M. & Menzel, R., Multiple sites of associative odor learning as revealed by local brain microinjections of octopamine in honeybees., Learn. Mem. 5, 146-156 (1998). Heisenberg, M. Mushroom body memoir: from maps to models. Nat. Rev. Neurosci. 4, 266-275 (2003).
Holland, P. C., Cognitive aspects of classical conditioning., Curr. Opinion Neurobiol. 3, 230-236 (1993).
Hussainin S.A., Komischke, B, Menzel, R., Lachnit, H., Forwarad and backward second-order Pavlovian conditioning in honeybees., Learn. Mem. 14, 678-683 (2007). Matsumoto, Y., Unoki, S., Aonuma, H., Mizunami, M., Nitric oxide-cGMP cascade is critical for cAMP-dependent long-term memory formation., Learn. Mem. 13, 35-44 (2006).
Menzel, R. & Giurfa, M., Dimensions of cognition in an insect, the honeybee., Behav. Cogn. Neurosci. Rev. 5, 24-40 (2006).
Müller, D., Gerber, B., Hellstern, F., Hammer, M., Menzel, R., Sensory preconditioning in honeybees., J. Exp. Biol. 203, 1351-1364 (2000).
Rescorla, R. A., Pavlovian conditioning., Am. Psychologist 40, 151-160 (1988).
Riemensperger, T., Völler, T., Stock, P., Buchner, E. & Fiala, A., Punishment prediction by dopaminergic neurons in Drosophila., Curr. Biol. 15, 1953-1960 (2005).
Schroll, C., Riemensperger, T., Bucher, D., Ehmer, J., Voller, T., Erbguth, K., Gerber, B., Hendel, T., Nagel, G., Buchner, E. & Fiala, A., Light-induced activation of distinct modulatory neurons triggers appetitive or aversive learning in Drosophila larvae., Curr Biol. 16, 1741-1747 (2006).
Schultz, W., Behavioral theories and the neurophysiology of reward., Annu. Rev. Psychol. 57, 87-115 (2006).
Schwaerzel, M., Monastirioti, M., Scholz, H., Friggi-Grelin, F., Birman, S. & Heisenberg, M., Dopamine and octopamine differentiate between aversive and appetitive olfactory memories in Drosophila., J. Neurosci. 23, 10495-10502 (2003).
Unoki, S., Matsumoto, Y., & Mizunami, M., Participation of octopaminergic reward system and dopaminergic punishment system in insect olfactory learning revealed by pharmacological study., Eur. J. Neurosci. 22, 1409-1416 (2005).
Unoki, S., Matsumoto, Y., & Mizunami, M., Roles of octopaminergic and dopaminergic neurons in mediating reward and punishment signals in insect visual learning., Eur. J. Neurosci. 24, 2031-2038 (2006).
Vergoz, V., Roussel E., Sandoz J.-C., Giurfa M., Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex., PLoS one 2, e288 (2007). Wise, R. A., Dopamine, learning and motivation., Nat. Rev. Neurosci. 5, 483-494 (2004).
Ⅶ 図の説明
Fig. 1. 昆虫における古典的条件付けのモデル OA/DA:報酬情報をもたらすオクトパミン作動性ニューロンまたは罰情報をもたらすドー パミン作動性ニューロン。US:無条件刺激。CS:条件刺激。UR:無条件反応。CR:条件 反応。US:無条件刺激を表現するニューロン。CS:条件刺激を表現するニューロン。 (A)ショウジョウバエにおける古典的条件付けを説明するモデル(Schwaerzel et al., 2003)。 CS ニューロンとオクトパミン作動性ニューロンまたはドーパミン作動性ニューロンからの シナプスが CR ニューロン上へ収束している。 (B)宇ノ木-水波モデルと名づけた新しい古 典的条件付けのモデル。条件付けの際CS ニューロンからオクトパミン作動性ニューロンま たはドーパミン作動性ニューロンへのシナプスが強化され、また、記憶の読み出しの際の CR ニューロンの活性化には CS ニューロンからの入力とともにオクトパミン作動性ニュー ロンまたはドーパミン作動性ニューロンからの入力が必要である(AND ゲート)と仮定した。 (C)宇ノ木-水波モデルによる二次条件付けの説明。 Fig. 2. 2 次条件付けの手順 (A)2 次条件付けの手順。第 1 段階で、匂い(CS1)と水(US)を対提示する嗅覚条件付けを行っ た。第2 段階で、視覚パターン(CS2)と匂い(CS1)を対提示した。(B)2 次条件付けの様子。 第1 段階では、パイナップルエッセンスを含ませた 3mm 四方のろ紙片を水の入った 1ml 注射器の針の先端から1cm の位置に取り付け、コオロギの触角にろ紙を近づけて匂いをか がせた2 秒後にコオロギの口に水を 1 滴与えた。第 2 段階では、視覚パターンを注射器の 針の根元に取り付け、パイナップルエッセンスを含ませたろ紙片を針の先端から1cm の位 置に取り付け、視覚パターンと匂いを同時に5 秒間提示した。視覚パターンとして白中心、 あるいは黒中心パターンを用いた。(C)2 次条件付けのスケジュール。訓練前テストの後、 第1 段階の 1 日後に第 2 段階を行い、その 1 時間後にテストを行った。(D)2 次条件付けと 対照実験の刺激スケジュール。第1 段階では CS1 を提示した 2 秒後に US を提示し、第 2 段階ではCS2 と CS1 を同時に 5 秒間提示した。これらをそれぞれ 5 分間隔で与えた。不対 合試行ではUS、CS1 または CS2 を 150 秒間隔で偽似ランダム化して提示した。 Fig. 3. 視覚嗜好性テスト (A)視覚パターン。黒中心パターンと白中心パターンを用いた。(B)視覚嗜好性テスト装置。 WCH :待機室。SD:スライド式扉。TCH:テスト室。SW:スライド式の壁。P:視覚パ ターン。(C)Preference Index(PI)の計算式。TA:パターンA を探索した時間。TB:パター ンB を探索した時間。 Fig. 4. 2 次条件付けの効果 2 次条件付けは、第 1 段階を訓練回数 4 回、訓練間隔 5 分間で行った 1 日後に、第 2 段階を訓練回数4 回、訓練間隔 5 分間で行った。訓練前と訓練 1 時間後に嗜好性テストを行っ た。対照実験は2 次条件付けの第 1 段階または第 2 段階の訓練を不対合試行にした。UP/P 群では第1 段階を不対合試行にし、P/UP 群では第 2 段階を不対合試行にした。条件付け前 (白のグラフ)後(黒のグラフ)(WCX test)と個体群間(M-W test)の統計解析の結果はグラフ上 のアステリスクで示した(*p<0.05; **p<0.01; ***p<0.001, NS p>0.05)。グラフ上の 数字は被験個体数を示す。 Fig. 5. 2 次条件付け第 1 段階でのオクトパミン受容体阻害剤の影響 プロトコルを上に示す。第1 段階の開始 30 分前に 1μM エピナスチンを含む生理食塩水を 3μl 投与した。2 次条件付けは、第 1 段階を訓練回数 4 回、訓練間隔 5 分間で行った 1 日 後に、第2 段階を訓練回数 4 回、訓練間隔 5 分間で行った。訓練前と訓練 1 時間後に視覚 嗜好性テストを行った。条件付け前(白のグラフ)後(黒のグラフ)(WCX test)の統計解析の結 果はアステリスクで示した(*p<0.05; **p<0.01; ***p<0.001, NS p>0.05)。グラフ上 の数字は被験個体数を示す。 Fig. 6. 2 次条件付け第 2 段階でのオクトパミン受容体阻害剤の影響 プロトコルを上に示す。第2 段階の開始 30 分前に 1μM エピナスチンを含む生理食塩水を 3μl 投与した。対照実験では生理食塩水を投与した。また、非投与群でも同じスケジュー ルで2 次条件付けを行った。2 次条件付けは、第 1 段階を訓練回数 4 回、訓練間隔 5 分間 で行った1 時間後に、第 2 段階を訓練回数 6 回、訓練間隔 5 分間で行った。訓練前と訓練 1 日後に視覚嗜好性テストを行った。条件付け前(白のグラフ)後(黒のグラフ) (WCX test)と個 体群間(M-W test)の統計解析の結果はアステリスクで示した(*p<0.05; **p<0.01; ** *p<0.001, NS p>0.05)。グラフ上の数字は被験個体数を示す。 Fig. 7. 記憶の読み出し段階でのオクトパミン受容体阻害剤の影響 プロトコルを上に示す。訓練後の視覚嗜好性テスト30 分前に 1μM エピナスチンを含む生 理食塩水を3μl 投与した。対照実験では生理食塩水を投与した。また、エピナスチン投与 後、効果が切れた後に視覚嗜好性テストを行う対照実験も行った。2 次条件付けは第 1 段階 を訓練回数4 回、訓練間隔 5 分間で行った 1 時間後に第 2 段階を訓練回数 6 回、訓練間隔 5 分間で行った。訓練前と訓練 1 時間後または 1 日後に視覚嗜好性テストを行った。条件付 け前(白のグラフ)後(黒のグラフ)(WCX test)と個体群間(M-W test)の統計解析の結果はアス テリスクで示した(*p<0.05; **p<0.01; ***p<0.001, NS p>0.05)。グラフ上の数字は 被験個体数を示す。
B
CS
AND gate
modifiable
CR
modifiable
CR
OA/DAUR
Appetitive/
aversive US
Olfactory/visual CS
Olfactory CS
CR
Appetitive/
aversive US
A
modifiable
CR
UR
CS
OA/DAC
Olfactory CS1
CR
CS1Visual CS2
CS2AND gate
modifiable
modifiable
OA/DAAppetitive/
aversive US
CR
UR
2nd stage
1st stage
test
test
1day
1 h
water odor source (pineapple)1st stage
odor source (pineapple) balack-center or white-center pattern2nd stage
B
A
C
1st stage
2nd stage
test
CS1→US CS2→CS1
CS2
CS1 : pineapple odor US : water CS2 : black-center or white-center pattern 5min 2s 2s paired 1st stage 5min 5s paired 2nd stage2.5min 2.5min 2.5min
unpaired 2nd stage unpaired 1st stage
2.5min 2.5min 2.5min
D
Fig. 2. 2 次条件付けの手順
B
WCH SD TCH P SWA
black-center pattern white-center pattern