博 士 学 位 論 文
Analysis of carbohydrates degrading enzymes and
development of efficient cultivation system in
ectomycorrhizal fungus Tricholoma matsutake
近 畿 大 学 大 学 院
農学研究科 応用生命化学専攻
大 沼 広 宜
Doctoral Dissertation
Analysis of carbohydrates degrading enzymes and
development of efficient cultivation system in
ectomycorrhizal fungus Tricholoma matsutake
Hiroki Onuma
Graduate school, Kindai University
Division of Agricultural Science
(Major: Applied Biological Chemistry)
Analysis of carbohydrates degrading enzymes and
development of efficient cultivation system in
ectomycorrhizal fungus Tricholoma matsutake
Hiroki Onuma
March, 2019
Graduate school, Kindai University
Division of Agricultural Science
Major: Applied Biological Chemistry
(Advisor: Prof. Norifumi Shirasaka)
外生菌根菌
Tricholoma matsutake の糖質分解酵素系の
解析と効率的培養法の開発
近 畿 大 学 大 学 院
農学研究科 応用生命化学専攻
大沼 広宜
(指導 : 白坂 憲章教授)
Submitted to the Graduate school, Kindai University, to fulfill the requirement for the Doctorate Degree.
CONTENTS
ABBREVIATIONS 1 INTRODUCTION 2 CHAPTER Ⅰ Production of starch utilization enzymes by ectomycorrhizal
fungus, Tricholoma matsutake
Section 1 Production of extracellular starch degradation enzymes by
T. matsutake 5 Section 2 Enzymatic characterization of an extracellular glucoamylase
from T. matsutake and its secretory expression in Pichia pastoris 21
CHAPTER Ⅱ Production of β-glucan hydrolytic enzymes by T. matsutake
Section 1 Purification and characterization of a glycoside hydrolase family 5 endoglucanase from T. matsutake in barley based
solid–state medium 41
Section 2 Functional analysis of a glycoside hydrolase family 7 protein
cellobiohydrolase gene from T. matsutake 63
Section 3 Substrate specificity of extracellular β-glucosidase from
T. matsutake, and its synergistic actions with β-glucan hydrolytic enzymes to produce glucose from β-glucan 75
CHAPTER Ⅲ Development of biomass quantification methods for mycelium solid-state medium cultivated T. matsutake 85
CONCLUSIONS 99 REFFRENCES 103 ACKNOWLEGMENTS PUBLICATION
1
ABBLEVIATIONS
BMGH medium Buffered glycerol-complex medium BMMH medium Buffered methanol-complex medium
bp Base pair(s)
cDNA Complementary DNA
DNA Deoxyribonucleic acid
GlcNAc N-acetylglucosamine
HPLC High performance liquid chromatography
kb Kilobase(s)
kDa Kilodalton
LB medium Luria-Bertani medium
MALDI-TOF/MS Matrix-assisted leaser desorption ionization - Time of flight/mass spectrometry
mRNA Messenger RNA
PCR Polymerase chain reaction
qRT-PCR Quantitative reverse transcription polymerase chain reaction
RNA Ribonucleic acid
RT-PCR Reverse transcription polymerase chain reaction
2
INTRODUCTION
Tricholoma matsutake (S. Ito et Imai) is an ectomycorrhizal fungus found in pine and
spruce forests in the Northern hemisphere [1, 2, 3]. This species forms soil mycelial aggregations termed “shiro”, which usually form a cycler arrangement of sporocarps known as fruiting fairy rings [4, 5]. The fungus produces commercially valuable mushrooms that have been revered in Japan for their flavor, medicinal properties and iconic significance for centuries. Over the past 50-60 years, this edible mushroom has become increasingly rare in Japan where the annual yield of the fungus has decreased from 12,000 tons in the 1940s to a few hundred tons today. One of the reasons for this reduction may be the introduction and spread of the pine nematode (Bursaphelencus lignicolus (Steiner and Buhrer) Nickle) in Japanese forests in addition to deforestation and modern forestry management practices that destabilize T. matsutake propagation [6, 7]. Nearly 3,000 tons of T. matsutake or closely related species are imported into Japan annually, with a retail value of approximately one billion US dollars [8]. Therefore, expectations have grown that pure artificial systems for cultivating T. matsutake will be developed, like those for currently commercially available mushrooms.
There are only three reports regarding the primordia or fruiting body formation of T. matsutake in a pure artificial culture system. First, Ogawa and Hamada reported that the formation of primordia using mineral soil with a mesh of <2 mm containing glucose and dried yeast and cultivated at 17-19°C [9]. Second, Kawai and Ogawa. also reported the formation of primordia of this fungus. Regarding the methods used for this, they emphasized the importance of selecting appropriately aged seeds for culture and the homogenization of mycelia. Then, they inoculated the homogenized seeds into solid medium containing vermiculite, glucose, and dried yeast. After 3 months at 17°C, primordia appeared [10]. Finally, Inaba et al. reported fruiting body formation of T. matsutake. When the mycelia from the latter’s “shiro” were mixed with sawdust medium containing several nutrients and sulfite pulp waste separated from softwood and inoculated in a plastic case at 24°C, the fruiting body formed 90 days after the mixing [11]. Unfortunately, other researchers have yet to be successful in repeating these methods.
3
However, artificial cultivation of ectomycorrhizal fungus, Lyophyllum shimeji (Kawamura) Hongo., had been succeeded in bottle cultivation using barley (Hordeum vulgare) grains and sawdust medium [12, 13, 14]. These reports indicated that the sufficient amount of barley starch to be used as a carbon source and can provide factors for production of the fruiting body in the medium. Ohta [15] investigated that the utilization ability of starch and amylose of ectomycorrhizal fungi (55 strains), shows that mycelial growth was good in barley grain medium for strains in which mycelial growth was also good in amylose as a carbon source. These reports indicate that some mycorrhizal fungi have the starch utilization ability.
Fig. 1. Comparison with the mycelial growth in T. matsutake NBRC 30605 strain between hamada’s agar plate and starch agar plate medium. A shows hamada’s agar medium (2% glucose as a carbon source) and B shows 2% soluble starch and 0.3% yeast extract agar medium. Cultivation was carried out at 24°C for 60 days.
The most serious problem in the artificial cultivation of the T. matsutake is its slow growth on artificial media (approximately 2 cm/m) [16], so that it is not easy to study on analysis of T. matsutake enzymes. Therefore, this species has been little studied regarding purification and characterization of carbohydrate degrading enzymes [16, 17, 18]. In practice, it is unclear which polysaccharides such as starch, cellulose, xylan, or others can be the best substrates for the fungal growth. Fig. 1 showed the comparison with soluble starch growth such as like glucose culture in 60 days cultivation. It is suggested that soluble starch can be good substrate for the mycelial growth as well as glucose which considered the most suitable carbon source. In this thesis, the author focused on
starch-4
degradation activities of the fungus and its related gene expression levels and mycelial growth of T. matsutake. Moreover, the author described to elucidate the enzymatic characterization and its relationships on β-glucan hydrolytic enzymes as endoglucanase, cellobiohydrolase and β-glucosidase. In order to develop the new method for artificial cultivation using starch and β-glucan substrate with T. matsutake.
Chapter Ⅰ describes the profiles of starch utilization and starch degrading enzyme in 35 strains of T. matsutake. The author also investigated the effect of various culture substrates for starch degrading enzyme activity and transcription levels of the starch-degrading enzyme related genes in T. matsutake strains. Moreover, the author describes the purification and enzymatic characteristics of glucoamylase from T. matsutake NBRC 30605 (TmGLA). Furthermore, the gene encoding TmGlu1 was expressed in the methylotrophic yeast Pichia pastoris to reveal that the translated gene has glucoamylase activity.
Chapter II describes the enzymatic characterization and some properties for its applications for β-glucan hydrolytic enzymes from T. matsutake. In the section 1, the author described purification and identify of an endoglucanase (TmEgl5A) produced by T. matsutake in barley based solid-state cultivation, and molecular cloning, heterologous expressed the gene encoding the enzyme in Pichia pastoris. In the section 2, heterologous expression in Pichia pastoris was attempted in order to analyze the function of CBH-like gene Tmcel7A derived from T. matsutake NBRC 30605 strain, and relationships in cellulose degradation was investigated. This section describes the molecular cloning and sequential analysis of the CBH-like gene, Tmcel7A from T. matsutake NBRC 30605 strain. After cloning, to understanding of the function of Tmcel7A that this gene was heterologous expression in Pichia pastoris. And in section 3, describes the isolation and identification of a primary structure of extracellular β-glucosidase and the substrate specificity of oligosaccharide degradation by the β-glucosidase. In addition, the synergy with β-glucosidase and cellulolytic enzymes toward β-glucans were described.
Chapter III describes that determined optimal enzymatic reaction conditions for chitin degradation in T. matsutake mycelia. The author also studied a solid-state culturing using barley and vermiculite to obtain a large amount of matsutake mycelia in a short period of time, and estimated T. matsutake fungal biomass in artificial cultivation medium.
5
CHAPTER Ⅰ
Production of starch utilization enzymes by ectomycorrhizal fungus,
Tricholoma matsutake
Section 1. Production of extracellular starch degradation enzymes by T. matsutake There have been studies on the productivity of carbohydrolases in modified artificial liquid medium with T. matsutake. The fungus showed low productivity of glucoamylase and endo-type cellulases, which are considered to have important roles in the artificial cultivation of mushrooms [19]. These results suggest that starch and cellulose are difficult to use as substrate for artificial cultivation on the fungus. However, Ohta investigated the utilization of starch-related substrate on 55 strains of ectomycorrhizal fungi including T. matsutake, and most of these fungi that grew well on amylose gave higher growth rates on solid-state barley medium [15]. Also, Terashita et al. examined extracellular amylase productions during the vegetative mycelial growth from T. matsutake, in a partly modified matsutake liquid medium (PMML) [20]. The α-amylase and glucoamylase activities in the culture filtrate were detected to have relatively high values at 40 days after inoculation. Using four kinds of starches purified from barley, corn, sweet potato and potato as the substrates, amylase activities in the culture filtrate (60 days after inoculation) of T. matsutake showed the highest activity toward the barley starches. Furthermore, Wan et al. reported that high glucoamylase activity was induced upon T. matsutake culture in medium containing amylose, compared with the levels with other saccharides such as glucose, rice starch, wheat starch, corn starch, and potato starch [21]. These results suggest that starch, especially amylose, affects the transcription of TmGlu1 and downstream glucoamylase activity, which is directly related to starch utilization. Therefore, in the artificial cultivation of T. matsutake, construction of a culture methods using starch-based medium is an effective culture substrate.
In this chapter, to clarify profiles of the starch utilization and starch degrading enzyme in 35 strains of T. matsutake. In addition, we investigated the effect of various culture substrates on starch degrading enzyme activity and transcription levels the starch-degrading enzyme related genes of T. matsutake strains.
6
MATERIALS AND METHODS
Materials. Pine-dex #1 was obtained from Matsutani Chemical Industry Co., Ltd., Hyogo, Japan. Barley starch was prepared from rolled barley from Hakubaku Co., Ltd., Yamanashi, Japan., and purified as described [22, 23].
Microorganisms and culture conditions. Tricholoma matsutake strains used in this section were shown in Table 1. The mycelia of T. matsutake strains were routinely grown on SY agar medium (2.0% soluble starch, 0.5% yeast extract 2.0% agar, pH 5.1). For screening, each strain was inoculated into 20mL of liquid medium containing 2.0% (w/v) glucose, 2.0% or 4.0% (w/v) soluble starch and Pine-dex #1 with 0.5% (w/v) yeast extract in a 100-mL of Erlenmeyer flask (pH 5.1).
For the qRT-PCR analysis, T. matsutake strains were inoculated to liquid medium containing 2% of carbohydrates such as glucose, maltose, soluble starch, Pine-dex #1, corn starch, and barley starch and 0.5% yeast extract. Hamada matsutake liquid medium [HML; 27], Kawai synthetic liquid medium [KSL; 28] and partially modified matsutake liquid medium [PMML; 20] were used as synthetic liquid medium. Hamada matsutake liquid medium consists of 2% (w/v) glucose, 10 g dried beer yeast (the extract from dried beer yeast was prepared in 100 ml tap water and boiled for 30 min), and 1.6 ml 1 N HCl per liter of distilled water. KSL medium comprised (per liter): glucose, 20 g; casamino acid, 1 g; yeast extract, 3 g; KH2PO4, 1 g; MgSO4・7H2O, 0.5 g; vitamin mixture, 1 mL;
metal solution, 1 mL. Vitamin mixture comprised (mg/liter): thiamine, 100; nicotinic acid, 100; folic acid, 100. Metal solution comprised (g/liter): CaCl2・2H2O 0.5: ZnSO4・7H2O,
0.5; MnSO4・4H2O, 0.5; ferric citrate, 0.5. A PMML medium was used in these
experiments. This medium was composed of 22.7 g glucose, 5.0 g yeast extract, 77.0 g potato extract, and 5.0 g Sunpearl CP (Nippon Seishi Industries; prepared from the sulfate pulp waste of softwood) per 1000 ml distilled water, with an initial pH of 5.1. These culture, containing 20 ml medium in a 100-ml Erlenmeyer flask. Following cultivations were done statically at 24℃ for 30 days.
7
Table 1. T. matsutake strains used in study.
strains Institution, description Host plant Isolation
No.101 The Institute of Iwaizumi Matsutake Mushroom, Iwate Prefecture,
Japan Pinus densiflora Iwate, Japan
No.103 〃 〃 〃 No.105 〃 〃 〃 No.107 〃 〃 〃 No.112 〃 〃 〃 No.114 〃 〃 〃 No.115 〃 〃 〃
KUKM-01 Isolated from commercially sold T. matsutake fruiting body 〃 Hyogo, Japan ATCC64715 American Type Culture Collection, USA [24] 〃 Hiroshima, Japan Z-1 Dr. K. Inaba, Kindai University [11] 〃 Niigata, Japan KMI-J-1 Dr. K. Yamanaka, Kyoto Mycological Institute, Kyoto, Japan [25, 26] Qurecus spp.,
(P. densata) Sichuan, China
KMI-N-5 〃 Lithocarpus, Castanopsis
(P. yunnanensis) Yunnan, China
KMI-W-1 〃 Lithocarpus, Castanopsis
(P. yunnanensis) Yunnan, China
KMI-Y-1 〃 Pinus densifrola,
Qurecus mongolica Jilin, China
KMI-103 〃 Pinus densifrola Shiga, Japan
NBRC6933 NITE Biological Resource Center, Chiba, Japan 〃
NBRC30604 〃 〃 NBRC30605 〃 〃 NBRC30606 〃 〃 NBRC30773 〃 〃 NBRC32806 〃 〃 NBRC32807 〃 〃 NBRC33137 〃 〃 NBRC101641 〃 〃 NBRC108253 〃 〃 NBRC108261 〃 〃 NBRC108720 〃 〃 NBRC108721 〃 〃 NBRC108722 〃 〃
KBM06 Kibun Co. Ltd., Tokyo, Japan 〃
TM-19 Hyogo Forestry Research and Technology Center, Hyogo, Japan 〃
Ichikawa 〃 〃
Tamba 〃 〃
Okayama Okayama Forestry Research and Technology Center, Okayama, Japan 〃 Okayama, Japan Hiroshima Isolated from the fruiting body 〃 Hiroshima, Japan
8
Enzyme assays and measurement of mycelial dry weight. After 30 days of incubation, vegetative mycelia and culture filtrate were separated. Culture filtrates were dialyzed in 50mM sodium acetate buffer (pH 5.0) for 24h. For measurement of the extracellular glucoamylase assayed with 1.0% soluble starch in 50mM sodium acetate (pH 5.0) at 37℃ for 60min. The enzymatic degradation of soluble starch was measured using the Glucose Colorimetric/Fluorometric Assay Kit (BioVision, Milpitas, CA, USA). One unit of glucoamylase activity was defined as that which produced 1 μmol glucose at 37°C for 1 min. α-Amylase activity was assayed with 1.0% soluble starch in 50mM sodium acetate (pH 5.0) at 37℃ for 60min. After the enzyme reaction, α-amylase was measured with the Somogyi method [29]. One unit of α-amylase activity was defined as the amount of enzyme that releases 1μmol of reducing end per min. Dry weight of T. matsutake lyophilized mycelium was measured using the precision electronic balance.
Analysis of the transcription levels of the starch degrading enzymes genes. The total RNA was extracted from the mycelia of T. matsutake NBRC 30605, Z-1 and KMI-J-1 using the FastGene™ RNA Basic Kit (NIPPON Genetics, Tokyo, Japan) and the
synthesis of the cDNA was carried out using the ReverTra Ace® qPCR RT Master Mix with gDNA Remover (Toyobo, Osaka, Japan). For quantitative analysis of gene expression, cDNA amplification was performed using FastStart SYBR Green Master (Roshe) and gene specific primers. Putative α-amylase gene (TmAmy1) was searched as based on Pholiota microspora PnAmy1 gene [30] using JGI Tricholoma matsutake 945 genome database (https://genome.jgi.doe.gov/Trima3/Trima3.home.html) and primers were designed (Forward primer, 5'-AACGGCAACTCCCTAAACATACA-3'; Reverse primer, 5'-CCGTCATCGGTATTAGAAGGAA-3'). The actin and glucoamylase gene (TmGlu1) primers were designed according to Wan et al. [21]. qRT-PCR analysis was used as the reference for mRNA level. The reactions were run on the LightCycler nano® system (Roshe). Relative expression levels were expressed as the 2-ΔCT calculated by the comparative CT value method: ΔCT = (CTtarget - CTactin) [31, 32].
9
RESULTS
Effect of the vegetative mycelial growth and production of extracellular starch degrading enzymes in T. matsutake.
(ⅰ) Vegetative mycelial growth. The effect of different starch substrates on vegetative mycelial growth of the T. matsutake 36 strains were showed in Table 2. The results of the vegetative mycelial growth of T. matsutake on a different medium at 24°C for 30 days are shown in Table 2. Cultivation in 2% soluble starch supplement medium culture average values was 61.1 mg/flask higher than glucose medium (53.6 mg/flask), especially, NBRC 30605 (109.9 mg/flask; 1.56 times higher with glucose medium), NBRC 30604 (109.1 mg/flask; 1.19 times) and NBRC 33137 (104.7mg/flask) showed the highest vegetative mycelial growth. Furthermore, in the 4% starch culture, there was a tendency to increased that the mycelial weights more than the average 76.2 mg / flask and glucose culture and 2% soluble starch. In T. matsutake 6 strains of NBRC 30604 (137.6 mg/flask), NBRC 33137 (131.4 mg/flask), TM-19 (130.4 mg/flask), Okayama (129.1 mg/flask), No. 103 (121.5 mg/flask) and NBRC 30605 (117.1 mg/flask), the mycelium dry weights were increased with the starch concentration. Also, in the Pine-dex#1 cultivation, vegetative mycelial dry weight were similar to 2% soluble starch cultivation, but in 4% Pine-dex#1 cultivation were comparable to 2% Pine-dex#1 cultivation expect for T. matsutake NBRC 33137 strain. In contrast, T. matsutake 17 strains values of mycelial dry weight showed that equally or lower than that of 2% glucose culture. Although, No. 112 was grown in 2% and 4% Pine-dex#1 medium (80.6 and 90.4 mg/flask) and No. 115 grown on 2% soluble starch and Pine-dex#1 medium (86.4 and 85.8 mg/flask) which grown compared with glucose medium (No.112; 63.5 mg/flask, No.115; 58.8 mg/flask).
(ⅱ) α-Amylase activity. Here, the effect of α-amylase production on different concentrations of medium were showed in Table 3. Average values of α-amylase activity were 48.5mU/flask (2% soluble starch, 2.4 times) and 48.6mU/flask (4% soluble starch, 2.4 times), 39.7mU/flask (2% Pine-dex#1, 1.9 times) and 35.7mU/flask (Pine-dex#1, 1.8 times) compared to that of glucose medium (20.5mU/flask).
10
(ⅲ) Glucoamylase activity. The glucoamylase activity in mycelia after incubation for 30 days in media containing several types of compounds was also investigated (Table 4). Average values of glucoamylase activity was 110mU/flask (2% soluble starch, 2.8 times) and 99.5mU/flask (4% soluble starch, 2.5 times) compared to that of glucose medium (39.7mU/flask). In contrast, there was no significance between glucoamylase activity in 2% Pine-dex#1 (29.9mU/flask, 0.8 times) and Pine-dex#1 (15.1mU/flask, 0.4 times). In the higher mycelial growth rate of strains that No. 103 (2% soluble starch; 216.4 mU/flask, 4% soluble starch; 251.3 mU/flask), NBRC 30605 (241.9 mU/flask; 232.3 mU/flask), No. 101 (145.9 mU/flask; 281.3 mU/flask) and Ichikawa (267.4 mU/flask; 245.3 mU/flask) showed high glucoamylase activity level with increasing the mycelial growth, as the case with 4% soluble starch medium. However, lower mycelial growth revels of strains that No. 112 (2% soluble starch; 272.4 mU/flask, 4% soluble starch; 233.5 mU/flask), NBRC 30773 (169.3 mU/flask; 219.9 mU/flask), NBRC 6933 (174.7 mU/flask; 209.8 mU/flask), No. 115 (306.6 mU/flask; 121.6 mU/flask), KMI-W-1 (145.9 mU/flask; 281.3 mU/flask) and No. 105 showed high levels of glucoamylase activity despite the low amount of the mycelial growth.
11
Table 2. The effect of different starch substrates on mycelial dry weight of Tricholoma matsutake.
Mycelial dry weight (mg/flask)
Glucose 2% Soluble starch 4% Soluble starch 2% Pine-dex #1 4% Pine-dex #1
KMI-J-1 63.7 ± 5.5 77.1 ± 10.4 146.3 ± 3.8 99.1 ± 13.1 94.3 ± 9.4 NBRC 30604 91.4 ± 5.0 109.1 ± 6.5 137.6 ± 6.2 111.7 ± 4.2 115.1 ± 5.6 NBRC 33137 86.2 ± 14.0 104.7 ± 5.1 131.4 ± 8.3 108.9 ± 3.8 140.6 ± 16.1 TM-19 91.4 ± 6.3 96.6 ± 9.7 130.4 ± 4.4 104.7 ± 9.8 117.0 ± 6.7 Okayama 73.5 ± 12.4 83.6 ± 3.1 129.1 ± 18.6 93.9 ± 8.6 96.9 ± 8.9 No. 103 61.7 ± 7.9 77.2 ± 4.8 121.5 ± 11.6 83.6 ± 8.0 71.0 ± 11.3 NBRC 30605 69.1 ± 7.7 109.9 ± 6.0 117.1 ± 4.9 97.1 ± 6.2 74.9 ± 8.4 ATCC 64715 83.5 ± 13.2 87.4 ± 7.1 114.8 ± 8.7 91.0 ± 10.4 86.1 ± 12.6 No. 101 69.1 ± 5.7 73.7 ± 9.2 106.3 ± 6.4 64.4 ± 9.8 97.8 ± 13.6 No. 107 56.3 ± 11.2 92.7 ± 2.6 104.1 ± 6.4 102.5 ± 4.8 104.8 ± 10.5 NBRC 108721 54.5 ± 6.9 64.5 ± 1.9 101.4 ± 8.2 71.8 ± 8.1 62.3 ± 16.9 NBRC 108722 37.3 ± 3.2 47.5 ± 5.3 95.1 ± 11.6 73.5 ± 13.6 47.2 ± 10.2 Ichikawa 48.7 ± 4.6 60.8 ± 5.4 94.1 ± 1.9 57.6 ± 7.4 59.2 ± 7.8 KMI-N-5 42.9 ± 1.7 75.7 ± 5.7 92.5 ± 11.8 91.8 ± 8.3 96.8 ± 10.0 NBRC 30606 38.5 ± 0.2 52.2 ± 5.4 75.8 ± 9.8 64.6 ± 5.6 76.3 ± 3.9 Z-1 58.0 ± 7.5 78.4 ± 1.7 74.4 ± 5.1 65.7 ± 11.0 52.9 ± 7.9 No. 114 47.6 ± 11.8 49.3 ± 11.5 71.5 ± 13.1 63.6 ± 8.2 67.0 ± 9.5 KBM06 45.2 ± 11.6 62.4 ± 13.6 68.4 ± 10.2 68.3 ± 7.3 32.9 ± 6.1 No. 112 63.5 ± 11.5 60.3 ± 11.2 67.8 ± 2.6 80.6 ± 4.4 90.4 ± 3.8 KUKM-03 54.2 ± 10.7 55.8 ± 7.8 64.5 ± 11.5 57.5 ± 1.1 76.0 ± 10.7 KMI-Y-1 48.2 ± 3.4 41.2 ± 2.6 63.2 ± 3.9 43.2 ± 6.7 56.3 ± 4.5 Hiroshima 70.0 ± 6.9 55.6 ± 7.8 62.0 ± 10.5 85.6 ± 8.7 79.0 ± 3.2 Tamba 46.1 ± 12.7 43.9 ± 2.6 61.9 ± 10.2 53.7 ± 7.4 45.3 ± 0.9 NBRC 30773 61.8 ± 3.1 62.3 ± 2.6 60.0 ± 3.7 71.4 ± 6.8 71.8 ± 5.2 NBRC 108720 88.8 ± 7.8 46.0 ± 4.7 56.9 ± 3.7 36.5 ± 2.3 47.5 ± 3.8 NBRC 6933 55.9 ± 6.3 53.2 ± 8.6 54.8 ± 3.5 63.3 ± 5.6 59.3 ± 6.1 NBRC 32807 22.6 ± 1.3 46.8 ± 5.6 48.5 ± 7.3 44.0 ± 2.5 22.7 ± 2.4 No. 115 58.8 ± 4.7 86.4 ± 5.4 44.2 ± 11.3 85.8 ± 12.7 55.7 ± 15.1 NBRC 108253 78.4 ± 8.5 39.5 ± 5.8 41.4 ± 5.2 74.8 ± 7.9 63.5 ± 10.4 KMI-W-1 19.8 ± 4.3 36.5 ± 5.2 39.8 ± 4.7 53.6 ± 2.8 33.9 ± 4.8 No. 105 22.9 ± 2.9 39.0 ± 6.0 31.9 ± 4.1 26.0 ± 1.3 37.3 ± 5.1 KMI-103 22.0 ± 0.8 32.1 ± 1.2 24.5 ± 2.0 26.2 ± 0.2 23.1 ± 4.2 NBRC 108261 17.3 ± 0.7 13.4 ± 0.8 14.9 ± 0.6 12.4 ± 0.4 5.5 ± 0.1 NBRC 101641 14.5 ± 0.5 10.1 ± 0.5 13.5 ± 0.3 5.4 ± 0.2 2.1 ± 0.3 NBRC 32806 12.4 ± 1.7 14.6 ± 1.7 7.7 ± 1.3 5.5 ± 0.6 7.9 ± 1.0
Above the dotted line indicates the group with high growth compared to the glucose culture. Below the dotted line shows the group with equivalent or lower growth than glucose culture. Each data value is
12
Table 3. The effect of different starch substrates on α-amylase activity from Tricholoma matsutake.
α-amylase activity (mU/flask)
Glucose 2% Soluble starch 4% Soluble starch 2% Pine-dex #1 4% Pine-dex #1
KMI-J-1 16.7 ± 0.7 50.3 ± 0.9 52.7 ± 0.9 45.3 ± 4.6 32.2 ± 3.2 NBRC30604 31.0 ± 0.4 63.7 ± 0.5 43.8 ± 1.7 51.2 ± 4.7 25.6 ± 2.5 NBRC33137 22.9 ± 2.4 22.9 ± 0.8 23.0 ± 0.5 22.9 ± 0.8 22.9 ± 0.4 TM-19 28.3 ± 3.6 53.0 ± 3.6 50.9 ± 0.3 48.0 ± 5.5 50.6 ± 6.5 Okayama 22.5 ± 1.4 94.1 ± 2.5 112.3 ± 11.2 86.4 ± 3.3 87.9 ± 2.9 No.103 14.6 ± 0.7 50.9 ± 2.1 63.7 ± 3.3 37.5 ± 2.8 28.6 ± 1.9 ATCC64715 24.1 ± 1.2 47.9 ± 2.6 50.0 ± 0.6 42.6 ± 2.1 45.6 ± 6.1 NBRC30605 33.4 ± 2.7 74.5 ± 6.5 63.4 ± 0.8 50.9 ± 6.9 41.4 ± 0.1 No.101 21.1 ± 1.9 62.2 ± 4.4 64.9 ± 2.4 37.2 ± 0.8 42.6 ± 4.8 No.107 20.1 ± 2.1 32.2 ± 0.9 39.0 ± 0.9 23.5 ± 0.3 26.8 ± 3.8 NBRC108721 24.1 ± 0.7 86.9 ± 4.8 97.1 ± 7.1 72.1 ± 3.8 40.2 ± 1.5 NBRC108722 9.8 ± 0.5 22.3 ± 0.9 23.8 ± 1.2 17.6 ± 1.1 13.7 ± 0.7 Ichikawa 30.4 ± 1.9 62.8 ± 2.2 81.3 ± 0.9 47.7 ± 3.5 45.3 ± 0.1 KMI-N-5 13.7 ± 1.4 35.1 ± 0.7 33.4 ± 0.4 45.6 ± 0.7 36.6 ± 4.3 NBRC30606 11.9 ± 0.3 33.4 ± 1.6 37.2 ± 0.4 29.5 ± 1.1 36.0 ± 1.3 Z-1 20.8 ± 2.7 45.2 ± 4.0 47.8 ± 1.4 35.7 ± 5.1 29.2 ± 1.2 No.114 22.9 ± 2.4 79.5 ± 10.2 76.5 ± 4.2 40.8 ± 1.5 44.1 ± 4.2 KBM06 33.4 ± 1.1 63.4 ± 4.3 61.3 ± 0.6 74.2 ± 4.8 54.5 ± 1.7 No.112 21.1 ± 2.0 62.3 ± 4.6 50.6 ± 0.5 64.9 ± 4.1 37.2 ± 1.5 KUKM-03 14.6 ± 0.1 53.6 ± 6.7 39.9 ± 3.8 31.6 ± 2.8 42.6 ± 2.7 KMI-Y-1 13.9 ± 0.2 23.5 ± 1.0 28.0 ± 1.9 27.6 ± 2.4 28.9 ± 3.4 Hiroshima 43.2 ± 3.2 100.7 ± 7.7 107.8 ± 8.4 101.9 ± 12.5 104.0 ± 10.8 Tamba 4.8 ± 0.3 22.3 ± 1.8 22.6 ± 0.2 25.3 ± 4.1 17.6 ± 0.1 NBRC30773 39.9 ± 3.2 54.8 ± 4.1 47.1 ± 0.7 17.6 ± 0.5 28.0 ± 2.7 NBRC108720 42.9 ± 0.7 88.2 ± 9.5 81.0 ± 2.7 79.5 ± 0.4 80.4 ± 7.1 NBRC6933 23.2 ± 0.8 43.8 ± 1.8 33.4 ± 0.3 33.1 ± 2.7 22.6 ± 0.2 NBRC32807 6.6 ± 0.9 9.8 ± 0.4 22.0 ± 3.5 22.9 ± 1.0 22.9 ± 1.1 No.115 16.7 ± 3.2 61.9 ± 3.7 44.9 ± 0.4 38.1 ± 3.9 19.1 ± 1.7 NBRC108253 15.8 ± 1.9 54.2 ± 6.5 49.4 ± 2.1 42.0 ± 0.5 34.6 ± 1.8 KMI-W-1 25.6 ± 0.6 32.2 ± 1.1 39.0 ± 1.0 29.5 ± 1.4 26.8 ± 4.5 No.105 17.9 ± 0.9 25.3 ± 1.4 19.9 ± 0.4 12.5 ± 0.3 17.0 ± 0.9 KMI-103 2.7 ± 0.1 11.0 ± 0.3 17.0 ± 1.5 15.8 ± 0.1 12.8 ± 0.1 NBRC108261 4.2 ± 0.1 8.3 ± 0.4 14.9 ± 0.1 12.8 ± 0.5 12.2 ± 0.1 NBRC101641 17.3 ± 0.6 51.2 ± 9.4 44.4 ± 4.3 15.8 ± 2.1 18.5 ± 1.1 NBRC32806 4.7 ± 0.2 16.7 ± 0.7 17.9 ± 0.9 11.3 ± 0.2 19.4 ± 2.4
α-amylase activity was measured by method of Somogyi (1952). Each data value is presented as the means ± SD (n = 3).
13
Table 4. The effect of different starch substrates on glucoamylase activities of Tricholoma matsutake. Glucoamylase activity (mU/flask)
Glucose 2% Soluble starch 4% Soluble starch 2% Pine-dex #1 4% Pine-dex #1
KMI-J-1 38.0 ± 4.3 71.7 ± 6.4 183.1 ± 14.5 82.8 ± 11.1 27.6 ± 4.7 NBRC30604 35.1 ± 0.8 59.2 ± 6.4 45.6 ± 1.1 13.0 ± 0.4 9.6 ± 0.1 NBRC33137 32.2 ± 0.6 63.1 ± 4.8 48.8 ± 4.1 15.6 ± 0.7 11.1 ± 0.3 TM-19 37.9 ± 2.3 101.8 ± 6.1 48.3 ± 1.5 23.3 ± 1.6 14.3 ± 1.8 Okayama 49.4 ± 3.2 99.4 ± 5.1 119.6 ± 7.6 29.8 ± 1.0 21.5 ± 0.4 No.103 87.1 ± 8.2 216.4 ± 14.2 251.3 ± 22.3 83.9 ± 6.2 15.1 ± 2.1 ATCC64715 41.6 ± 4.5 125.0 ± 8.4 82.8 ± 5.1 21.0 ± 2.6 17.1 ± 0.5 NBRC30605 63.6 ± 9.3 241.9 ± 15.4 232.3 ± 12.1 31.7 ± 1.8 19.7 ± 2.0 No.101 56.3 ± 1.5 145.9 ± 11.3 281.3 ± 18.6 8.8 ± 0.2 45.2 ± 3.9 No.107 30.7 ± 0.9 152.8 ± 6.9 57.8 ± 7.4 22.8 ± 1.0 14.1 ± 1.1 NBRC108721 19.4 ± 0.5 26.6 ± 2.9 18.7 ± 0.7 11.1 ± 1.5 11.3 ± 0.8 NBRC108722 25.6 ± 0.7 70.3 ± 6.5 66.3 ± 8.6 9.7 ± 0.9 9.0 ± 1.1 Ichikawa 111.5 ± 5.8 267.4 ± 21.9 245.3 ± 25.4 17.8 ± 0.7 20.0 ± 1.9 KMI-N-5 43.8 ± 2.1 68.8 ± 1.5 24.5 ± 0.7 20.2 ± 0.5 17.0 ± 0.9 NBRC30606 14.7 ± 0.4 26.3 ± 0.1 23.7 ± 0.4 9.9 ± 0.2 9.7 ± 0.5 Z-1 15.6 ± 3.3 71.5 ± 6.2 99.4 ± 7.8 18.3 ± 5.1 15.4 ± 1.2 No.114 14.4 ± 0.4 42.8 ± 1.8 27.1 ± 1.7 14.4 ± 0.8 11.8 ± 0.1 KBM06 28.0 ± 2.1 71.6 ± 9.3 77.2 ± 4.1 16.5 ± 0.8 9.7 ± 1.2 No.112 58.3 ± 4.2 272.4 ± 16.2 233.5 ± 17.6 47.7 ± 2.9 20.5 ± 0.2 KUKM-01 23.7 ± 1.4 124.2 ± 10.1 73.8 ± 15.4 17.6 ± 0.6 18.1 ± 0.5 KMI-Y-1 8.9 ± 0.2 22.8 ± 1.3 11.9 ± 0.9 10.6 ± 1.1 9.7 ± 0.3 Hiroshima 11.2 ± 0.8 56.1 ± 1.2 27.8 ± 2.1 11.9 ± 0.4 10.8 ± 0.2 Tamba 27.9 ± 4.3 88.2 ± 10.0 99.3 ± 5.5 15.9 ± 0.1 15.8 ± 0.5 NBRC30773 49.9 ± 3.1 169.3 ± 16.1 219.9 ± 18.9 14.7 ± 0.1 10.4 ± 1.2 NBRC108720 11.8 ± 1.2 32.8 ± 1.1 11.8 ± 0.2 10.7 ± 0.9 9.5 ± 0.8 NBRC6933 156.8 ± 12.4 174.7 ± 16.5 209.8 ± 14.2 55.1 ± 7.1 15.8 ± 0.3 NBRC32807 21.2 ± 0.7 76.3 ± 4.2 69.2 ± 3.7 15.9 ± 0.5 15.8 ± 0.3 No.115 53.3 ± 4.6 306.6 ± 24.2 121.6 ± 7.7 38.1 ± 0.9 12.3 ± 0.2 NBRC108253 11.1 ± 1.0 21.2 ± 0.5 23.2 ± 0.1 13.7 ± 1.1 21.2 ± 0.8 KMI-W-1 84.8 ± 10.5 273.4 ± 21.7 259.1 ± 19.3 261.2 ± 15.4 20.0 ± 1.1 No.105 76.3 ± 7.4 255.8 ± 13.8 138.3 ± 9.7 44.1 ± 4.3 10.7 ± 1.2 KMI-103 11.2 ± 0.4 29.8 ± 0.9 14.3 ± 0.4 9.8 ± 0.3 10.2 ± 0.1 NBRC108261 11.0 ± 0.1 9.6 ± 0.2 9.3 ± 0.8 9.1 ± 0.3 8.9 ± 0.6 NBRC101641 17.1 ± 1.3 35.8 ± 1.4 20.4 ± 1.7 8.8 ± 0.2 8.2 ± 0.1 NBRC32806 10.5 ± 1.1 12.2 ± 1.0 12.9 ± 0.3 9.6 ± 0.2 10.6 ± 0.3
Glucoamylase activity was measured by Glucose assay kit (Biovision). Each data value is presented as the means ± SD (n = 3).
14
Effect of medium composition on extracellular starch degrading enzyme productivities and gene expression profiles.
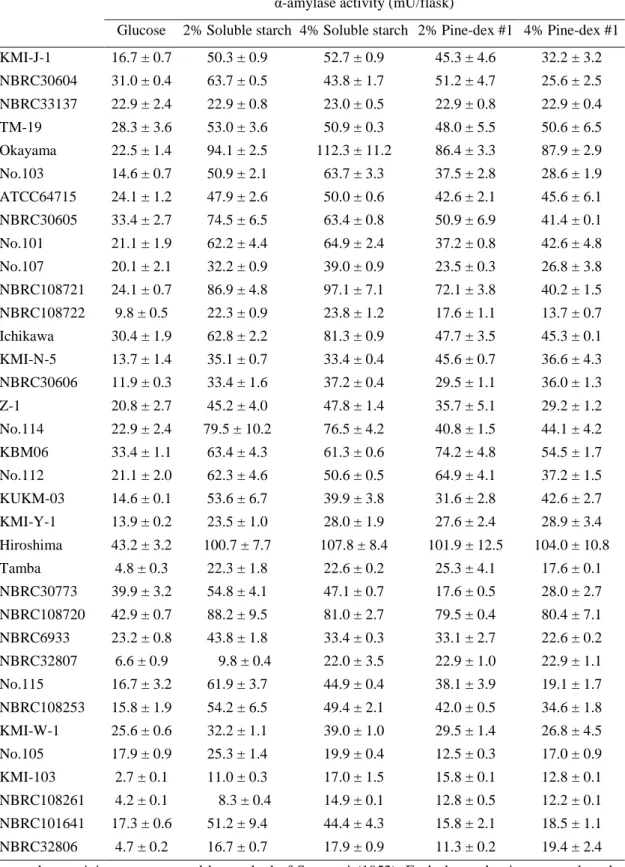
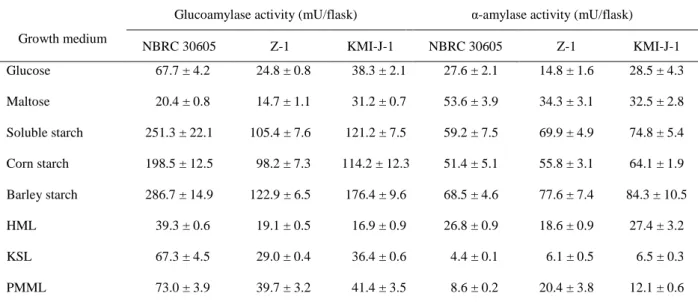
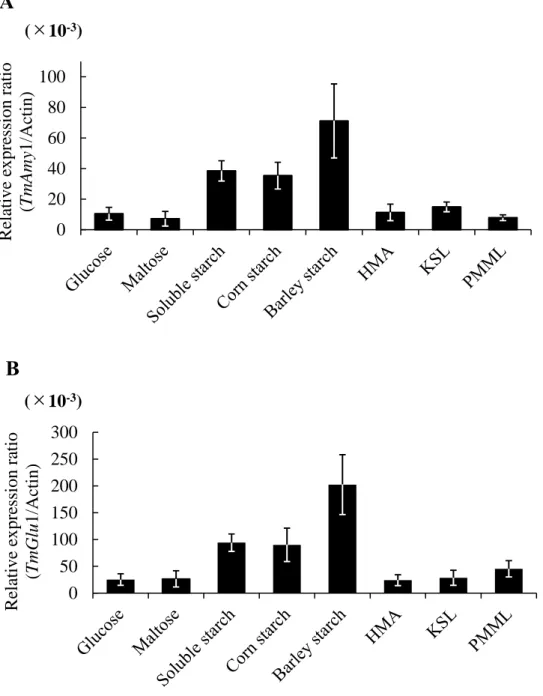
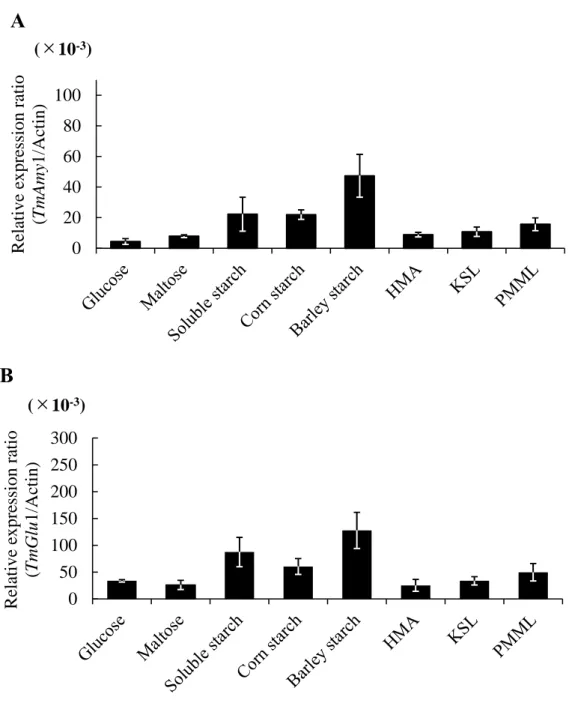
To investigate effect of medium composition and carbon source on mycelial growth and enzyme productivity of T. matsutake NBRC 30605, Z-1 and KMI-J-1, vegetative mycelial yield, α-amylase and glucoamylase activity was measured after cultivation for 30 days in listed liquid media (Table 5). The results showed that the mycelial dry weight in medium containing soluble starch (96.6, 81.1 and 75.8 mg/flask) and corn starch (74.4, 61.5 and 68.7 mg/flask), barley starch (108.4, 81.6 and 88.4 mg/flask) were higher than that media containing glucose (51.4, 58.1 and 60.8 mg/flask), maltose (41.7, 51.6 and 46.7 mg/flask) and synthetic media. The α-amylase and glucoamylase activity in the culture broth of 30 days cultivation were also measured (Table 6). The highest α-amylase and glucoamylase activity was observed in the barley starch medium (glucoamylase activity: 286.7 ± 14.9, 176.4 ± 9.6 and 122.9 ± 6.5 mU/flask; α-amylase activity: 68.5 ± 4.6, 84.3 ± 10.5 and 77.6 ± 7.4 mU/flask).
To determine the reasons for the increased α-amylase and glucoamylase activity in the media containing starch substrates as a carbon source, qRT-PCR was used to monitor the expression of TmAmy and TmGlu1 mRNA in mycelia which were cultured in different media. As shown in Fig. 1 to Fig. 3, transcription levels of TmAmy and TmGlu1 were increased remarkably in the starch media compared to glucose medium and synthetic media. The medium containing barley starch induced highest expression level among tested starches. However, there was no significance transcription levels between glucose, maltose, HMA, KSL and PMML. These results suggest that the media containing barley starch, corn starch, and soluble starch affect vegetative mycelial growth, α-amylase and glucoamylase production.
15
Table 5. Effects of medium compositions on mycelial growth of the T. matsutake.
Mycelial dry weight (mg/flask)
Growth medium NBRC 30605 Z-1 KMI-J-1
Glucose 51.4 ± 7.4 58.1 ± 4.8 60.8 ± 4.1
Maltose 41.7 ± 6.1 51.6 ± 1.0 46.7 ± 0.9
Soluble starch 96.6 ± 5.1 81.1 ± 7.1 75.8 ± 6.9
Corn starch 74.4 ± 4.2 61.5 ± 6.1 68.7 ± 4.8
Barley starch 108.4 ± 9.2 88.4 ± 9.6 81.6 ± 6.8
Hamada's liquid medium (HML) 29.8 ± 1.2 23.7 ± 3.0 21.6 ± 2.1
Kawai's liquid medium (KSL) 35.0 ± 5.0 34.1 ± 5.7 31.5 ± 3.3
Partial modified matsutake medium (PMML) 61.5 ± 6.0 56.0 ± 3.5 58.8 ± 7.2
Each data value is presented as the means ± SD (n = 3).
Table 6. Effects of medium compositions on starch degrading enzyme activities of the T. matsutake.
Glucoamylase activity (mU/flask) α-amylase activity (mU/flask)
Growth medium NBRC 30605 Z-1 KMI-J-1 NBRC 30605 Z-1 KMI-J-1 Glucose 67.7 ± 4.2 24.8 ± 0.8 38.3 ± 2.1 27.6 ± 2.1 14.8 ± 1.6 28.5 ± 4.3 Maltose 20.4 ± 0.8 14.7 ± 1.1 31.2 ± 0.7 53.6 ± 3.9 34.3 ± 3.1 32.5 ± 2.8 Soluble starch 251.3 ± 22.1 105.4 ± 7.6 121.2 ± 7.5 59.2 ± 7.5 69.9 ± 4.9 74.8 ± 5.4 Corn starch 198.5 ± 12.5 98.2 ± 7.3 114.2 ± 12.3 51.4 ± 5.1 55.8 ± 3.1 64.1 ± 1.9 Barley starch 286.7 ± 14.9 122.9 ± 6.5 176.4 ± 9.6 68.5 ± 4.6 77.6 ± 7.4 84.3 ± 10.5 HML 39.3 ± 0.6 19.1 ± 0.5 16.9 ± 0.9 26.8 ± 0.9 18.6 ± 0.9 27.4 ± 3.2 KSL 67.3 ± 4.5 29.0 ± 0.4 36.4 ± 0.6 4.4 ± 0.1 6.1 ± 0.5 6.5 ± 0.3 PMML 73.0 ± 3.9 39.7 ± 3.2 41.4 ± 3.5 8.6 ± 0.2 20.4 ± 3.8 12.1 ± 0.6
Glucoamylase activity was measured by Glucose assay kit (Biovision). α-Amylase activity was measured by method of Somogyi (1952). Each data value is presented as the means ± SD (n = 3).
16
Fig. 1. Effect of different media on the starch-degrading gene expression levels in T. matsutake NBRC 30605. A: Transcription levels TmAmy1 in T. matsutake mycelia. B: Transcription levels TmGlu1 in T. matsutake mycelia. The transcription levels are relative to the actin as housekeeping gene. Error bars indicate standard deviation (±SD) of experiments performed in three determinations.
0 20 40 60 80 100 R elative e x pre ssi on ra ti o (T mA my 1/Ac ti n) (×10-3)
A
0 50 100 150 200 250 300 R elative e x pre ssi on ra ti o (T mG lu 1/Ac ti n) (×10-3)B
17
Fig. 2. Effect of different media on the starch-degrading gene expression levels in T. matsutake Z-1 strain. A: Transcription levels TmAmy1 in T. matsutake mycelia. B: Transcription levels TmGlu1 in T. matsutake mycelia. The transcription levels are relative to the actin as housekeeping gene. Error bars indicate standard deviation (±SD) of experiments performed in three determinations.
0 20 40 60 80 100 R elative e x pre ssi on ra ti o (T mA my 1/Ac ti n) (×10-3) 0 50 100 150 200 250 300 R elative e x pre ssi on ra ti o (T mG lu 1/Ac ti n) (×10-3)
A
B
18
Fig. 3. Efficiency of different media on the starch-degrading gene expression levels in T. matsutake KMI-J-1 strain. A: Transcription levels TmAmy1 in T. matsutake mycelia. B: Transcription levels TmGlu1 in T. matsutake mycelia. The transcription levels are relative to the actin as housekeeping gene. Error bars indicate standard deviation (±SD) of experiments performed in three determinations.
0 20 40 60 80 100 R elative e x pre ssi on ra ti o (T mA my 1/Ac ti n) (×10-3)
A
0 50 100 150 200 250 300 R elative e x pre ssi on ra ti o (T mG lu 1/Ac ti n) (×10-3)B
19
DISCUSSION
In previous studies, it has been reported that T. matsutake has the low starch degradation activity, among the polysaccharide substrates [19, 20, 27]. L. shimeji, which has been able to obtain fruiting body artificially, shows good growth on starch substrate and apparent starch degradation ability compared to T. matsutake [12, 13, 14, 33, 34]. Therefore, it is important to select strains with high starch utilization availability not only by obtaining the mycelial yield of T. matsutake in a short period but also by adapting to culture with starch substrates. In this chapter, the author investigated the effect of medium composition of liquid medium and supplementation of starch and maltooligosaccharides on 35 kinds of T. matsutake strains.
Although, Pine-dex #1 which is a mixture of malto-oligosaccharides showed a comparatively vegetative mycelial growth and a higher activity of α-amylase than glucose medium. However, the productivity of glucoamylase activity was scarcely observed except for KMI-W-1 strain. Tricholoma spp. can grow slowly in a starch substrate when a small amount of glucose is added as a starter [35]. Kusuda et al. found that 5% glucose medium inhibited the mycelial growth of T. matsutake Z-1 and J-1 strains, whereas, T. matsutake Z-1 and J-1 strains were possible to growth rate of the soluble starch concentration in medium up to 10% and 15% [27].
In the most previous studies, the agar medium with glucose as the main carbon source were used for the inoculum of T. matsutake [33, 36, 37, 38].In this study, high yield of mycelium could be obtained in shorter period by using starch instead of glucose as the carbon source for inoculation medium. In addition, the glucoamylase activity was markedly increased by soluble starch culture.It is suggested that the use of glucose for inoculation medium of T. matsutake not only inhibits production of the starch-degrading activity, but also results in slowing mycelial growth. Therefore, in order to improve mycelial growth and starch utilization of T. matsutake, it is important to construct a culture system using soluble starch as a carbon source for inoculation medium.
I have showed that the mycelium and culture filtrate of various liquid media of three strains of T. matsutake (NBRC 30605, Z-1 and KMI-J-1) were used, and the effect on vegetative mycelia growth and starch-degrading enzymes activity were investigated.
20
As a result, it was shown that barley starch caused not only mycelial significantly higher weight yield compared to the glucose culture, but also the starch-degrading enzymes activity (α-amylase and glucoamylase), especially glucoamylase activity.
Intact normal barley starch, number (DPn) and weight (DPw) average degrees of polymerization, respectively (DP): 1,110 to 1,570 and 2,450 to 6,440 with 3.6-5.2 chains per molecules [39, 40, 41, 42, 43], whereas maize (corn starch) amyloses were 830-990 and 2500-2680 with 2.4-2.9 chains per molecules [44, 45]. In addition, barley amyloses had β-limit dextrin of 74-80% [43], which were slightly lower than those for maize (81-84%) [44, 45]. These values indicated that barley starch had higher amylose content and generally wider DP distribution than maize starch. Thus, cultivation with barley starch as a carbon source, it was suggested that T. matsutake had more affective the enzyme activity and related gene expression level of high amylose content, resulting in increase of vegetative mycelial growth.
These results suggested that starches, especially barley starch is an effective carbon source for mycelial growth and starch-degradation enzymes production of T. matsutake. However, since TmGlu1 and TmAmy1 genes are predicted and their functions as proteins were not characterized, it is necessary to clarify the functions of these genes in order to elucidate the starch utilization in T. matsutake.
SUMMARY
In this chapter, 35 strains of T. matsutake were compared regarding mycelium growth on several starch substrates, and starch-degrading enzyme activities. Mycelial weight and glucoamylase activity of almost strains were promoted by 2% soluble starch comparing with 2% glucose liquid culture. In addition, glucoamylase and α-amylase activities were significantly promoted by barley starch as a carbon source comparing with glucose culture. The results of gene expression levels of the putative gene of TmGlu1 and TmAmy also showed similar induction profiles.
21
Section 2. Enzymatic characterization of an extracellular glucoamylase from T. matsutake and its secretory expression in Pichia pastoris
In Lyophyllum shimeji (Kawamura) Hongo, an ectomycorrhizal basidiomycetous fungus, fruiting body formation has been successfully achieved by pure culture without a symbiotic relationship with a host plant [12, 13, 14]. The fruiting body of the fungus was cultivated in a barley-based solid medium containing synthetic liquid medium. One of the causes of this success is considered to be the decrease of osmotic pressure from glucose because of the slow degradation of barley starch by carbohydrolases from L. shimeji. Ohta investigated the utilization of starch-related substrate on 55 strains of ectomycorrhizal fungi including T. matsutake, and most of these fungi that grew well on amylose gave higher growth rates on solid-state barley medium [15]. In addition, Terashita et al. investigated the productivity of amylases in the cultivation from the vegetative mycelial stage to the fruiting body formation period in L. shimeji and found that the strain with the ability to undergo fruiting body formation showed higher glucoamylase activity than strains without this ability [33]. Moreover, Kusuda et al. found that the glucoamylase activity increased in the fruiting body formation phase of L. shimeji and purified the glucoamylase and characterized its properties [34]. Together, these reports suggest that starch is a suitable substrate for long-term cultivation of L. shimeji and that some amount of glucoamylase activity is necessary.
The most serious problem the artificial cultivation of the T. matsutake is its slow growth on artificial media (approximately 2 cm/m) [16], so that it is not easy to study on analysis of T. matsutake enzymes. Therefore, this species has been little studied regarding purification and characterization of carbohydrate degrading enzymes [16, 17, 18]. In practice, it is unclear which polysaccharides such as starch, cellulose, xylan, or others can be the best substrates for the fungal growth.
This chapter describes the purification and enzymatic characteristics of glucoamylase from T. matsutake NBRC 30605 (TmGLA). Furthermore, the gene encoding TmGlu1 was expressed in the methylotrophic yeast Pichia pastoris to reveal that the translated gene has glucoamylase activity.
22
MATERIALS AND METHODS
Strain and culture conditions. Tricholoma matsutake strain NBRC 30605 (NBRC; Biological Resource Center, NITE, Japan) was used in this study. Because the whole genome sequence of the strain was done and was released in DDBJ (Accession no. BDDP01000001-BDDP01088884). This strain was cultured on Hamada’s matsutake agar (HMA; 2% glucose, 0.5% dried yeast, 2.0% agar, pH 5.1) medium prepared with tap water [36]. For the purification of glucoamylase, a mycelial block (diameter 5 × 5 × 5 mm) was cut from a medium plate containing HMA for 60 days at 24°C in a petri dish (diameter, 90 mm). Then, it was inoculated into an Erlenmeyer flask (100 mL) containing 20 mL of synthetic liquid (SL) medium [28] comprised (liter−1): soluble starch, 20 g;
casamino acid, 1 g; yeast extract, 3 g; KH2PO4, 1 g; MgSO4・7H2O, 0.5 g; vitamin mixture,
1 mL; metal solution, 1 mL. Vitamin mixture comprised (mg/liter): thiamine, 100; nicotinic acid, 100; folic acid, 100. Metal solution comprised (g/liter): CaCl2・2H2O 0.5:
ZnSO4・7H2O, 0.5; MnSO4・4H2O, 0.5; Ferric citrate, 0.5. After inoculation, a static
culture was performed at 24°C and 70% humidity for 60 d in the dark.
Glucoamylase assay and protein determination. The standard glucoamylase assay was incubated for 60 min at 37°C. The enzymatic degradation of soluble starch was measured using the Glucose Colorimetric/Fluorometric Assay Kit (BioVision, Milpitas, CA, USA). One unit of enzyme activity was defined as that which produced 1 μmol glucose at 37°C for 1 min. Protein concentrations were determined using the Bio-Rad protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA) with bovine serum albumin as standard [46].
Purification of extracellular glucoamylase from T. matsutake NBRC 30605. After 60 d of incubation, crude TmGLA in the 2.3 liter of liquid culture medium from 154 flasks (100 mL erlenmeyer flask) were collected by filtration through No. 2 filter paper (Toyo Roshi, Tokyo, Japan) to remove mycelia. Proteins precipitating with 60% saturated ammonium sulfate were collected by centrifugation (12,000 × g, 30 min, 4°C), dissolved in a minimum volume of 20 mM sodium phosphate buffer (pH 6.5), and dialyzed against
23
the same buffer with 3 changes for 24 h. Then, the dialyzed solution was centrifuged and the supernatant was loaded onto a pre-equilibrated (with 20 mM sodium phosphate buffer, pH 6.5) TOYOPEARL NH2-750F column (φ2.5 × 30 cm). The active fraction appeared
as the unabsorbed fraction (450 mL), which was then brought to 30% ammonium sulfate saturation and loaded onto a TOYOPEARL Butyl-650M column (φ2.5 × 20 cm) that had been equilibrated with 20 mM sodium phosphate buffer containing 30% saturated ammonium sulfate. The bound proteins were eluted with a 300-mL gradient of ammonium sulfate at 30% to 0% saturation in 10 mM sodium phosphate buffer (pH 6.5). Active fractions from the TOYOPEARL Butyl-650M column were combined and finally concentrated with the Amicon centrifugal filtration device (Merck Millipore, Darmstadt, Germany). The protein samples obtained at the previous steps were applied to a column of TOYOPEARL HW-55F (φ1.0 × 100 cm) equilibrated with 20 mM sodium phosphate buffer containing 150 mM NaCl. The column was eluted at 0.1 mL/min and active fractions were collected.
SDS polyacrylamide gel electrophoresis (SDS-PAGE) analysis and estimation of apparent molecular mass. SDS-PAGE was performed by the method of Laemmli [47] using an electrophoresis unit supplied by Atto (Tokyo, Japan). Samples were boiled for 10 min in the presence of 10% SDS, 2-mercaptoehanol, and bromophenol blue. The gels were stained with Coomassie Brilliant Blue R-250. The following molecular mass marker proteins were used: Precision Plus Protein™ Standards (Bio-Rad, Tokyo, Japan). The molecular mass of native purified enzyme was analyzed using gel-permeation chromatography. The enzyme was subjected to HPLC with an LC-10A system (Shimadzu, Kyoto, Japan) on a TSKgel G3000SWXL column (7.8 mm I.D. × 30
cm length) at a flow rate of 1.0 mL/min, using 150 mM NaCl solution at room temperature. The absorbance of the effluent was recorded at 230 nm. The relative mobility of the enzyme was compared with the mobility of MW-Marker (Oriental Yeast, Kyoto, Japan): glutamate dehydrogenase (290,000), lactate dehydrogenase (142,000), enolase (67,000), myokinase (32,000), and cytochrome C (12,400).
24
Trypsin digestion and mass spectrometry. The purified TmGLA was digested with trypsin, and the resulting peptides were spotted on an MTP 384 target plate ground steel BC and matrixed using HCCA (α-cyano-4-hydroxycinnamic acid). The peptide mass spectrometric analysis was performed using Autoflex Speed KN2 (Bruker, MD, USA). Obtained mass spectra were accumulated until a satisfactory S/N ratio had been obtained. Parent mass peaks in the range from 1200 to 3000 m/z were picked out for MS/MS analysis. The peptide mass fingerprints of MS/MS data were acquired and processed using Flexanalysis software (Bruker) and were converted to LIFT files using Proteinscape 3.0.3 software (Bruker). The LIFT files were analyzed using the MASCOT search engine (Matrix Science, London, UK).
Enzymatic characterization of purified TmGLA. The effect of pH on the glucoamylase activity was assayed by varying the pH of the reaction mixture using various buffers: 50 mM acetate buffer (pH 2.0-5.0), 50 mM sodium phosphate buffer (pH 5.0-8.0), and 50 mM Tris-HCl (pH 8.0-10.0). The effect of temperature on the glucoamylase activity was assayed at pH 5.0 in sodium acetate buffer, in the range of 4-100°C. The effects of metallic ions and some enzyme inhibitors were determined by pre-incubating purified enzyme with various metal ions, ethylenediaminetetraacetic acid (EDTA), and dithiothreitol (DTT) at 37°C for 30 min before starting the enzyme reaction. The substrate specificity of glucoamylase was determined for a variety of natural substrates (soluble starch, corn starch, maltose, maltotriose, maltotetraose, maltopentaose, amylose A [MW, 2900], amylose B [MW, 16 000], amylopectin, pullulan, isomaltose, isomaltooligosaccharide, glycogen, α-cyclodextrin, and β-cyclodextrin). The Glucose Assay Kit (BioVision) was used to test these substrates.
Products analysis of the enzyme reaction mixture. The hydrolysis products released from soluble starch was individually treated with purified enzyme (TmGLA; 0.1 mg/mL) in 100 μL of 50 mM sodium acetate buffer (pH 5.0) at 37℃ for 0 h, 30 min, 1 h, 3 h, 6 h, 12 h and 24 h. The hydrolysis products were subjected to thin-layer chromatography (TLC) on silica gel 60 F254 plates (Merck, Darmstadt, Germany) in ethyl
25
De novo sequencing analysis of T. matsutake NBRC 30605 and determination of TmGlu1 gene sequence. The genomic DNA of dikaryotic T. matsutake NBRC30605 cells was extracted using ISOPLANT-Ⅱ (Wako Pure Chemical, Osaka, Japan). DNA libraries were prepared from 50 ng of genomic DNA using the Nextera DNA sample prep kit (Illumina, San Diego, CA, USA), obtaining fragments of approximately 700 bp. DNA libraries were sequenced using the Illumina NextSeq500 (Illumina) with 151 bp paired-end sequencing. Total read sequences (37,740,518) were assembled using CLC_Genomics_Workbench v. 8.0 (CLC bio, Aarhus, Denmark). Based on the internal amino acid sequences (7 peptides), the TmGlu1 gene sequence was used as a query for a local BLAST search in the draft whole-genome sequence of the fungus. The nucleotide sequence of the TmGLA gene has been registered DDBJ/EMBL/GenBank with accession no. BDDP00031826.
cDNA cloning of TmGlu1 gene. Total RNA was prepared from T. matsutake NBRC 30605 mycelia incubated at 24°C for 30 d on HMA plate medium using Sepasol-RNA I Super G (Nacalai Tesque, Kyoto, Japan), and first-strand cDNA was synthesized by ReverTra Ace (Toyobo, Osaka, Japan). Reverse transcription was performed at 42°C for 30 min and was stopped by exposure to 99°C for 5 min, followed by placing on ice. The TmGlu1 gene specific primer-pair was designed with reference the genome sequence database from T. matsutake NBRC 30605 and T. matsutake NBRC 30773 TmGlu1 gene (accession no. AB604354; Wan et al. 2012). The PCR mixture contained 10 μL of Emerald Amp® MAX PCR Master Mix (Takara Bio, Shiga, Japan), 1 μL of TmGlu1-F
ATGCGTCTGTCCTTTCTTTT-3′; 100 pmol/μL), 1 μL of TmGlu1-R (5′-CTACCTCCAAGAATCATTCA-3′; 100 pmol/μL), 1 μL of the first-strand cDNA (50-100 ng/μL), and 7 μL of sterilized water. Thirty cycles were performed, each consisting of a denaturation step at 98°C for 10 s, an annealing step at 56°C for 30 s, and an extension step at 72°C for 2 min. The amplified cDNA fragment was separated by 0.7% agarose gel electrophoresis, purified with an ISOSPIN Agarose Gel extraction kit (Nippon Gene, Tokyo, Japan), and then ligated in T-Vector pMD20 (Takara Bio), using Ligation high Ver.2 (Toyobo). Nucleotide sequence analysis of the ligated fragment was performed
26
using an Applied Biosystems 3730xl DNA Analyzer (Life Technologies, Carlsbad, CA, USA).
The nucleotide sequence data reported in this paper have been registered in the DDBJ/EMBL/GenBank nucleotide sequence database with accession no. LC203036. The deduced amino acid sequence of the enzyme was analyzed by BLAST searches in the National Center for Biotechnology and Information database (NCBI; https://www.ncbi.nlm.nih.gov/).
The signal sequence and N-glycosylation site were predicted using the SignalP 4.1 Server (http://www.cbs.dtu.dk/services/SignalP/) and NetNGlyc 1.0 Server (http://www.cbs.dtu.dk/services/NetNGlyc/), respectively.
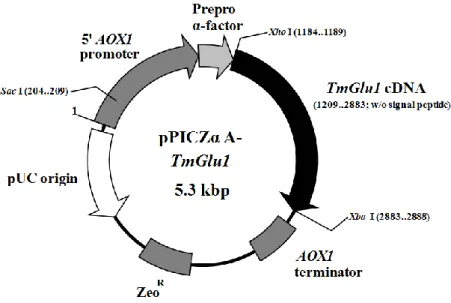
Heterologous expression of TmGlu1 gene in Pichia pastoris X-33 and purification of the recombinant enzyme. To express the TmGlu1 gene in Pichia pastoris, the plasmid pPICZα-A (Invitrogen, Carlsbad, USA) was used. For PCR amplification of the cDNA, TmGlu1-Xho Ⅰ F (5′-TATACTCGAGAAAAGAGAGGCTGAAGCT CAATCTGGTAC-3′; Xho Ⅰ site is underlined) and TmGlu1-Xba Ⅰ R (5′-TATATCTAGACTACCTCCAAGAATCATTCA-3′; XbaⅠ site is underlined and the stop codon is included) were used. The amplified DNA fragment digested with an appropriate amount of XhoⅠ and XbaⅠ (Takara Bio) enzyme was purified to the appropriate size (1,674 bp) from agarose gel, and was ligated into pPICZα-A to yield the expression vector. P. pastoris strain X-33 (Life Technologies) was used as a host strain. Transformation of P. pastoris X-33 with the expression vector pPICZα-A-TmGlu1 (Fig. 1) was performed using the Electro Gene Transfer Equipment GTE-10 (Shimadzu). The transformant was cultured as follows: Single colonies were grown in 3 mL of BMGH medium (100 mM potassium phosphate buffer at pH 6.0, 1.34% of yeast nitrogen base w/o amino acid, 0.4 µg/mL biotin, and 1.0% glycerol) containing 100 μg/mL zeocin (Life Technologies) with shaking (180 rpm) at 30°C. After 24 h of cultivation, 3 mL of the culture medium was added to 100 mL of BMGH medium in a 500-mL shaking flask, and cultured with shaking (130 rpm) at 26.5°C for 2 d. Cultured cells were collected by centrifugation (1,500 × g, 5 min, 4°C) into the same volume of BMMH (100 mM potassium phosphate buffer at pH 6.0, 1.34% yeast nitrogen base w/o amino acid, 0.4 µg/mL biotin, and 0.5% methanol)
27
containing 1.0% casamino acid (Nihon Pharmaceutical, Tokyo, Japan) and cultured with shaking (130 rpm) at 26.5°C. Methanol as an inductor was added to the culture medium every 24 h. The total cultivation time was 96 h from the start of induction, and the culture medium was centrifuged at 1,500 × g for 5 min at 4°C. The supernatant (2 L) of the culture medium was purified as native TmGLA.
Fig. 1. Recombinant P. pastoris expression plasmids pPICzαA-TmGlu1 constructed for the TmGlu1 gene from T. matsutake.
AOX1 is the alcohol oxidase gene from P. pastoris. ZeoR is a zeocin (phleomycin)
resistance gene expressed against E. coli and P. pastoris.
Endoglycosidase H treatment. Purified native TmGLA (0.1 mg/mL) and recombinant TmGlu1 (0.1 mg/mL) from the P. pastoris transformant were boiled for 5 min in 50 mM sodium phosphate buffer (pH 5.0). After cooling, the sample was mixed with an equal volume of 50 mM sodium phosphate buffer (pH 5.0) containing 0.01 units of endoglycosidase H or the same buffer without endoglycosidase H, for a sequential reaction at 37°C for 24 h. After incubation, the digested sample were analyzed by SDS-PAGE.
28
RESULTS AND DISCUSSION
Purification and internal amino acid sequence analysis of TmGLA.
Previously, it was reported that high glucoamylase activity was induced upon fungus culture in medium containing amylose, compared with the levels with other saccharides such as glucose, rice starch, wheat starch, corn starch, and potato starch [21]. However, the detailed enzymatic properties of glucoamylase from T. matsutake had not been elucidated. Here, TmGLA was purified from culture broth of soluble starch medium and its enzymatic characteristics were described.
TmGLA from T. matsutake NBRC 30605 was purified 33.2-fold with a 3.0 recovery yield (Table 1) after ammonium sulfate fractionation and three-column chromatography.
Table 1. Purification of glucoamylase from Tricholoma matsutake NBRC 30605.
Purification steps Total protein
(mg) Total activity (units) a Specific activity (units/mg) Yield (%) Purification (-fold) Crude enzyme 522.8 207.8 0.40 100 1
Ammonium sulfate fractionation
(0-60%) 307.1 181.8 0.59 87.5 1.5
TOYOPEARL NH2-750F 91.0 164.7 1.81 79.2 4.6
TOYOPEARL Butyl-650M 2.20 15.7 7.15 7.6 18.0
TOYOPEARL HW-55F 0.47 6.1 13.2 3.0 33.2
a One enzyme unit (U) was defined as the amount required to release 1 μmol of glucose from the substrate per
29
The purified enzyme hydrolyzed soluble starch into glucose at 13.2 units/mg under conditions. The purified TmGLA gave a single protein band on SDS-PAGE, with a molecular mass of approximately 61.5 kDa (Fig. 2). The molecular mass of the native enzyme as determined by high-performance gel chromatography on a TSKgel G3000SWXL column was 63.9 kDa, indicating that the native enzyme was active as a
monomer.
Fig. 2. SDS-PAGE of purified TmGLA.
Proteins were separated on 12.5% polyacrylamide gel in the presence of 0.01% SDS. Lane1, molecular weight standards; Lane 2, Purified TmGLA.
30
To identify the purified TmGLA, the internal amino acid sequences of the protein were analyzed. After SDS-PAGE of the purified TmGLA, it was digested by trypsin and then peptide fragments were obtained. The tryptic peptide fragments, G-1 to G-7, were sequenced by MALDI-TOF/MS analysis (Table 2).
Table 2. Tryptic fragments matching the theoretical digested fragments of glucoamylase (TmGLA) from Tricholoma matsutake a.
No. Measured mass (Da) Predicted mass (Da) Δmass (Da) MC b
Tryptic fragment sequence Position (start-end) G-1 2307.022 2306.206 -0.816 0 AGVVIASPSTTNPNYLFTWIR 55—75 G-2 1991.847 1990.965 -0.882 1 FNIDETAFTGPWGRPQR 130—146 G-3 2999.786 2999.049 -0.030 0 DANTVLASIHTFDAAAGCDATTFQPCSDR 269—297 G-4 1466.704 1465.793 -0.911 0 ALANLLTYVDAFR 298—310 G-5 1907.833 1906.826 -1.007 0 NIYAINSGIATNQGSATGR 311—329 G-6 1527.646 1526.737 -0.909 0 YTPIGGGLSEQYSR 415—428 G-7 1085.528 1084.520 -1.007 0 QGIAPASWGAK 454—464 a
Peptide mass spectrometric analysis was performed on a MALDI-TOF/MS system; autoflex speed KN2 and the MALDI-TOF/MS data were analyzed using Proteinscape.
b
MC, number of miscleavages in tryptic digestion.
NCBInr BLAST search of the protein database indicated that the peptide sequences of the enzyme were identical to that of glycoside hydrolase family 15 protein TmGlu1 from Tricholoma matsutake NBRC30773 [21]. Attempted analysis of the N-terminal amino acid sequence of TmGLA did not detect the sequence. Yoon et al. reported the purification and molecular cloning of GH family 15 glucoamylase of carbohydrate-active enzymes (CAZymes; CAZY Carbohydrate-Active enZYmes.: http://www.cazy.org/) from the brown-rot basidiomycete Fomitopsis palustris (FpGLA). The N-terminal glutamine of the native FpGLA might have been changed to pyroglutamate, leading to a failure of Edman degradation to function [48]. Because the amino acid sequence of FpGLA showed 62% homology with that of TmGLA and the N-terminal amino acid of TmGLA is glutamine, it is suggested that the glutamine changes to pyroglutamate in native TmGLA. Therefore, we detected the internal amino acid sequence of TmGLA by peptide mass fingerprinting using MALDI-TOF/MS.
31 Enzymatic properties of TmGLA.
The enzymatic properties of TmGLA were determined using soluble starch as a substrate. The enzyme activity was determined using the Glucose Assay Kit (BioVision). The effects of chemical compounds on TmGLA activity were assayed. The enzyme was slightly activated by the presence of the Ca2+ and Mn2+. Among the Ba2+, K+, Al3+, Mg2+,
Fe3+, Na+, Ni+, Li+, Cu2+, Co2+, EDTA and DTT had no effect on enzyme activity. The
activity of the enzyme was slightly inhibited by the presence of 5 mM Hg2+, Fe2+, Zn2+,
and Pb2+. In contrast, 5 mM Ag+ was strongly inhibited the glucoamylase activity. There
is a result indicating that this enzyme was not a divalent cation (Ca2+, Mg2+, Mn2+)
requiring protein, suggesting the stabilization of the higher structure of TmGLA. The effects of pH and temperature on enzyme activity are shown in Fig. 3. The enzyme was active between 4°C and 50°C, and was stable pH 4.0-6.0, with maximum activity at 60°C and pH 5.0.
Enzymatic characteristics of glucoamylase from basidiomycetes have been investigated and reported; however, few reports have been published on species such as Lyophyllum shimeji (LyGLA) [34], Lentinula edodes (LeGLA; gla1) [49], Schizophyllum commune (ScGLA) [50], and Hypocrea peltata (HyGLA) [51]. Except for LyGLA, the enzymatic properties of these enzymes are similar to TmGLA with regard to active temperature and pH range, the effect of metal ions, and molecular mass. LyGLA was purified from extracellular L. shimeji strain MH01721. The purified enzyme showed a molecular mass of approximately 25 kDa on SDS-PAGE, and was most active at around 40 °C and pH 5.0. LyGLA was remarkably activated by the presence of Ca2+ ions. These
32
Fig. 3. Effect of temperature and pH on purified TmGLA enzyme activity. A and B: Effect of temperature on enzyme stability and optimal activity, respectively. A: Purified enzyme was incubated in 50 mM sodium acetate buffer at pH 5.0 for 60 min; temperature range, 4°C-80°C. B: The effect of temperature (4°C-80°C) on enzyme activity. C and D: Effect of pH on enzyme stability and activity, respectively. C: Enzyme was incubated for 30 min at 30°C in 50 mM sodium acetate buffer (pH 2.0-5.0, black square), 50 mM sodium phosphate buffer (pH 5.0-8.0, black circle), or 50 mM Tris-HCl buffer (pH 8.0-10.0, black triangle). D: The activity of purified enzyme on soluble starch in 50 mM sodium acetate buffer (pH 2.0-5.0, black square), 50 mM sodium phosphate buffer (pH 5.0-8.0, black circle), or 50 mM Tris-HCl buffer (pH 8.0-10.0, black triangle). Data are presented as the mean of three trials ± SD.
33
The substrate specificity of TmGLA was assayed using carbohydrates with different polysaccharides (Table 3). TmGLA readily hydrolyzed amylose A (MW, 2,900; 146%) and showed equal activity against pullulan (95%), amylopectin (92%), and amylose B (MW, 16,000; 90%). Activity against glycogen and corn starch was weak (49% and 35%, respectively). In addition, the enzyme also acted against malto-oligosaccharides (maltose, 15%; maltotriose, 52%; maltotetraose, 65%; and maltopentaose, 74%) and isomaltose (7%). No enzyme activity was observed with the substrates trehalose (α, α-1,1-linkage), kojibiose (α-1,2-linkage), nigerose (α-1,3-linkage), α-cyclodextrin, and β-cyclodextrin. These results indicate that TmGLA hydrolyzes α-1,4 and α-1,6 linkages of glycosidic saccharides. Soluble starch was hydrolyzed by TmGLA, and the final products were analyzed by TLC (Fig. 4). It was shown that the purified glucoamylase gradually hydrolyzed to glucose units during 24 hours after the start of the reaction.
Glucoamylase (α-1,4-glucan glucohydrolase, amyloglucosidase, EC 3.2.1.3) belongs to GH family 15; it is an exo-acting enzyme that removes the glucose units from the non-reducing ends of amylose, amylopectin, and glycogen by hydrolyzing α-1,4-linkages in a consecutive manner, producing D-glucose as the sole product. It also hydrolyzes α-1,6-linkages, although at a much slower rate [52]. TmGLA showed broad specific activity toward and α-1,6-glycosidic substrates. Amylose A with α-1,4-glycosidic polysaccharide could be the best substrate of the enzyme, and low-molecular-weight amylose A (average M.W. 2,900) was better than the high-molecular-low-molecular-weight form (amylose B, average M.W. 16,000) in this regard. Hydrolyzing activity toward several malto-oligosaccharides was also examined. The enzyme could hydrolyze maltopentaose, maltotetraose, maltotriose, maltose, and isomaltose, although the activity was lower toward smaller oligosaccharides.
However, the enzyme did not hydrolyze the cyclic polysaccharides. When the enzyme hydrolyzed low-molecular-weight amylose A, the produced saccharide was glucose. From these results, we conclude that TmGLA is a common glucoamylase belonging to GH family 15.
34
Fig. 4. Hydrolysis product of soluble starch by the purified enzyme.
TmGLA was incubated in 50mM sodium acetate buffer (pH 5.0) at 37 ℃ for 0 h, 0.5 h, 1 h, 3 h, 6 h, 12 h and 24 h. Hydrolysates were subject with TLC in ethyl acetate/methanol/water (8:4:3, v/v) and stained with 50% H2SO4.
Table 3. Substrate specificity of TmGLA.
Substrate a Relative activity (%) (units/mg protein) Specific activity Polysaccharides (1.0%) Soluble starch 100 13.9 Amylose A (MW, 2,900) 139 19.3 Amylose B (MW, 16,000) 90 12.5 Amylopectin 92 12.8 Pullulan 95 13.2 Glycogen 49 6.81 Corn starch 35 4.86 Oligosaccharides (10 mM) Maltopentaose 74 10.3 Maltotetraose 65 9.02 Maltotriose 52 7.22 Maltose 16 2.22 Isomaltose 7 1.04 β-cyclodextrin ND ND α-cyclodextrin ND ND Trehalose ND ND Kojibiose ND ND Nigerose ND ND
a Assays were performed at 37°C and pH 5.0. The activities on
polysaccharides and oligosaccharides are based on released D-glucose as determined using the Glucose assay kit.
35 Cloning and sequencing of TmGlu1 gene.
The de novo genome sequence of the genomic DNA T. matsutake NBRC 30605 was determined using Illumina NextSeq 500 paired-end technology provided in Kindai University, Faculty of Agriculture. The final assembled data contained 88,884 contigs of total length 131,718,925 bp, with an N50 contig length of 2909 bp. The gene prediction and functional annotation were carried out by AUGUSTUS 2.5.5 [53] and the Microbial Genome Annotation Pipeline [MiGAP; 54]. It predicted a total of 23,546 protein coding genes. The deduced amino acid sequences of seven trypsin-digested peptides from native TmGLA of TmGlu1 from T. matsutake NBRC30773 [accession no. AB604354; 21] were mapped. A 2,186 bp DNA fragment starting at ATG and terminating at the TAG codon was found in the nucleotide sequences encoding TmGlu1 on the whole genome sequence data of T. matsutake NBRC 30605. Alignment with the TmGlu1 cDNA indicated that this genomic sequence contained the complete protein-coding region. The coding region of the TmGlu1 gene (1,731 bp) was interrupted by eight introns (58, 59, 59, 59, 56, 52, 56, and 56 bp). The open reading frame encoded a polypeptide of 576 amino acids (Fig. 5). The first 19 amino acid residues in the N-terminal region were assigned as a signal peptide using the online program SignalP 4.1 server. The result of NCBI Conserved Domain Search suggested that this protein has a two-domain structure consisting of an N-terminal domain with the GH family 15 signature and a C-terminal carbohydrate-binding module (CBM) 20 region. Six N-glycosylation sites were predicted as an Asn-Xaa-Thr/Ser region [55]. The alignment of the TmGlu1 protein sequence compared with the T. matsutake NBRC 30773 [21], Pleurotus ostreatus PC15 [56], Phanerochaete carnosa HHB-10118-sp [57], Pholiota microHHB-10118-spora [58], Lentinula edodes L-54 [59], F. palustris FFPRI 0507 [48] and glucoamylase gene encoding sequences, deduced from the individual cDNAs were shown in Table 4. The primary structure of TmGlu1 was found to have 100%, 73%, 70%, 69%, 67%, 66%, and 62% sequence homology with the sequences of these glucoamylase genes encoding amino acid respectively.