Analgesic mechanisms of essential oil
components
2asayuki Takaishi
)octor of Philosophy
A nalgesic mechanisms of
essential oil components
M asayuk i T ak aishi
C ontents
S ummar y
1. I ntroduction
2. M ater ial and M ethods 2-1. Molecular cloning 2-2. R eagents
2-3. C ell C ulture 2-4. Human subjects
2-5. hT R PA 1, hT R PV 1 and hT R PM8 mutants 2-6. T ransfection
2-7. C alcium Imaging E xperiment 2-8. E lectrophysiology
2-9. S ensory irritation tests 2-10. D ata A nal ysis
3. R esults
3-1. S creening essential oils for the ability to activate hT R PM8 but not hT R PA 1 3-1-1. E ucal yptus oil shows hT R PM8-activating ability with little activation of
hT R PA 1
3-1-2. 1,8-cineole activates hT R PM8 but not hT R PA 1
3-1-3. 1,8-cineole acts on hT R PM8 and hT R PV 3, but not on hT R PV 1, hT R PV 2 or hT R PA 1
3-1-4. 1,4-cineole activates hT R PM8 and hT R PA 1 when expressed in HE K 293T cells
3-1-5. 1,4-cineole but not 1,8-cineole activates hT R PA 1
3-1-6. 1,8-cineole inhibits hT R PA 1 currents activated by different agonists 3-1-7. 1,8-cineole inhibits sensory irritation caused by hT R PA 1 agonists in vivo
3-2. S creening camphor analogs to find more effective T R PA 1 antagonists 3-2-1. S creening of naturally occurring compounds having effects on hT R PA 1
3-2-2. E ffects of borneol, 2-methylisoborneol, fenchyl alcohol, and norcamphor on hT R PA 1-mediated current responses
3-2-3. B orneol, 2-methylisoborneol, and fenchyl alcohol, but not norcamphor inhibit hT R PA 1 determined by a C a2+-imaging method
3-2-4. B orneol, 2-methylisoborneol, and fenchyl alcohol inhibit hT R PA 1 current in a dose-dependent manner
3-2-5. T he hydroxyl group of borneol contributes to inhibition of human T R PA 1
3-3. T he effects of menthol on human T R PV 1
3-3-1. Menthol inhibited sensory irritation caused by an hT R PV 1 agonist in vivo 3-3-2. Menthol inhibited hT R PV 1 activity induced by capsaicin.
3-3-3. Menthol inhibited the activation of hT R PV 1 induced by thermal stimulation. 3-3-4. C apsaicin inhibited hT R PM8 activity induced by menthol and cold
stimulation.
3-3-5. C apsaicin inhibited the activation of hT R PM8 induced by thermal stimulation.
3-3-6. T he binding site of menthol in T R PV 1 was distinguishable from that of capsaicin, whereas the binding site of capsaicin in T R PM8 modestly interacted with that of menthol.
4. D iscussion
4-1. S creening essential oils for the ability to activate hT R PM8 but not hT R PA 1 4-2. S creening camphor analogs to find more effective T R PA 1 antagonists
4-3. T he effects of menthol on human T R PV 1
5. A ck nowledgement 6. R eferences
S ummar y
T ransient receptor potential (T R P) channels respond to a wide variety of sensory stimuli, including temperature, nociceptive compounds, touch, osmolarity and pheromones. T R PV 1 can be activated by chemical ligands such as capsaicin, endocannabinoid, anandamide and protons. It is also activated by physical stimuli such as heat and acts as an integrator of multiple noxious stimuli. T R PA 1 is an ion channel targeted by pungent irritants such as those from mustard oil and garlic and is thought to function in diverse sensory processes, including cold nociception and inflammatory pain. T herefore, T R PV 1 and T R PA 1 ion channels are involved in acute nociception and inflammatory pain and are considered to be promising targets for the development of analgesic agents. Most of the published T R PV 1 and T R PA 1 antagonists lack optimal properties for clinical development such as selectivity, solubility, oral bioavailability, and/or acceptable pharmacokinetics. T hus, it is important to explore naturall y occurring antagonists of these channels, especially those that have an extensive safety profile based on widespread usage.
E ssential oils are often used in alternative medicine as analgesic and anti-inflammatory remedies. F or example, menthol, the main ingredient of peppermint, is commonly used for pain relief. T he analgesic effects of menthol are believed to occur through T R PM8 activation. However, the molecular mechanisms underlying the analgesic effects of these ingredients are largely unknown. In fact, high doses of menthol cause sensory irritation through T R PA 1 activation in humans. C amphor, another essential oil component, is now known to exert analgesic effects through inhibition of T R PA 1 and activation of T R PM8. However, camphor is not suitable for
use as an analgesic compound because it causes warm or hot sensations, probably through T R PV 1 and T R PV 3 activation. I concluded that it was important to identify other effective analgesic compounds that activated T R PM8 and inhibited T R PA 1 but did not activate T R PV 1.
T hrough the screening of essential oils, I found that eucalyptus oil exhibited a relativel y high hT R PM8-activating ability with less activation of hT R PA 1. F urthermore, 1,8-cineole, a major component of eucal yptus oil, inhibited hT R PA 1 activated by several agonists with a half-maximal inhibitory concentration ( IC50) of 3.4 mM for A IT C (20 μM)-evoked hT R PA 1 currents. In addition, sensory irritation tests in vivo showed that 1,8-cineole conferred an analgesic effect on the sensory irritation produced by menthol and a T R PA 1 agonist, octanol. Moreover, 1,4-cineole, another component of eucalyptus oil, activated hT R PA 1. S everal compounds with similar structures exhibited different effects on hT R PA 1. Menthol and 1,4-cineole activated hT R PA 1, whereas camphor and 1,8-cineole inhibited hT R PA 1. Given these variable effects on hT R PA 1, I hypothesized that more detailed analyses would lead to the identification of more effective compounds.
To identify more effective T R PA 1 antagonists, I screened camphor analogs among monoterpenes that comprise a group of naturall y occurring organic compounds derived from essential oils that have been used for anesthetic, analgesic, anti-inflammatory and anti-pruritic applications. I found that borneol, 2-methylisoborneol and fenchyl alcohol inhibited hT R PA 1 activity. T he hT R PA 1 currents induced by A IT C (20 µ M) were inhibited by borneol, 2-methylisoborneol or fenchyl alcohol, each in a dose-dependent manner. I obtained IC50 values of 0.20, 0.12 and 0.32 mM for borneol, 2-methylisoborneol and fenchyl alcohol, respectively, values that are much lower than
the IC50 values of 1,8-cineole (3.43 mM) and camphor (1.26 mM). In addition, sensory irritation tests in vivo showed that borneol conferred an analgesic effect on the sensory irritation produced by menthol. F urthermore, I found that the S 873, T 874, and Y 812 residues of hT R PA 1 were involved in the inhibitory effects, suggesting that the hydroxyl group in the cyclohexane of the inhibitors may interact with these amino acids.
To understand the analgesic mechanism of menthol, I examined the effects of menthol on hT R PV 1. T he hT R PV 1 currents induced by capsaicin were inhibited by menthol in a dose-dependent manner, with an IC50 value of 1.17 mM. In addition, an in vivo sensory irritation test showed that menthol conferred an analgesic effect on the sensory irritation produced by vanillyl butyl ether (V B E ), a T R PV 1 agonist. F urthermore, I found that Y 511, S 512 and T 550 of hT R PV 1, which are binding sites of capsaicin, were onl y slightly involved in the inhibitory effects of menthol. T hese data suggest that menthol interacts with sites different from those of capsaicin. T hese results show that an analgesic effect of high-dose of menthol is derived, at least in part, from its inhibitory effect on T R PV 1.
In this study, I elucidated the molecular mechanisms underl ying the analgesic effects of essential oil components. F urther anal ysis of these compounds could lead to better understanding of the structural determinants for the action of these compounds on T R PV 1 or T R PA 1 and the development of anti-nociceptive agents through T R PA 1 or T R PV 1 inhibition.
1 I ntroduction
Monoterpenes, like menthol and camphor, comprise a group of naturally occurring organic compounds derived from essential oils that have been used for anesthetic, analgesic [ X u et al., 2005; Galeotti et al., 2001; Galeotti et al., 2002] , anti-inflammatory [ X u et al., 2005; S antos et al., 2001] and antipruritic applications [ Umezu et al., 2001; A nand et al., 2003] . However, the molecular mechanisms by which essential oils and these compounds exert their effects are largely unknown.
T ransient receptor potential (T R P) channels respond to a wide variety of sensory stimuli, including temperature, nociceptive compounds, touch, osmolarity, and pheromones [ C hristensen et al., 2007; Minke et al., 2002; Z hang et al., 2008] . T R PV 1 is activated by capsaicin as well as heat, protons and some endogenous substances known to be associated with tissue inflammation [ C aterina et al., 1997; Tominaga et al., 1998; Tominaga and C aterina, 2004] . S ince T R PV 1 acts as an integrator of painful stimuli, T R PV 1 antagonists can be viewed as promising types of analgesics [ Gavva et al., 2008; Gunthorpe et al., 2009; Marcello et al., 2010; Marcello et al., 2013] . A number of potent, small T R PV 1 antagonists, including capsazepine, B C T C , C T PC , A MG9810 and S B -452533 have advanced into clinical trials for the evaluation of their analgesic activities [ B evan et al., 1992; V alenzano et al., 2003; R ami et al., 2004; Gavva et al., 2005; Weil et al., 2005] . A lthough some of these antagonists reduced noxious heat sensation, hyperthermia was a serious side effect that often led to their withdrawal from the clinical trials.
T R PA 1 is a receptor that responds to noxious cold temperatures in rodents and
pungent compounds, including all yl isothiocyanate (A IT C ), a component of mustard oil [ Story et al., 2003; B andell et al., 2004; J ordt et al., 2004; Obata et al., 2005; K wan et al., 2006] . A lthough the role of T R PA 1 in sensing noxious cold stimuli and somatic mechanosensations in vivo remains unsettled, especially in mammals [ Obata et al., 2005; K wan et al., 2006; Petrus et al., 2007] , T R PA 1 has been established as a chemical nocisensor for a wide variety of reactive compounds such as flufenamic acid (F FA ), 2-aminoethoxydiphenyl borate (2-A PB ), icilin, menthol, intracellular calcium, and zinc ions [ Story et al., 2003; B autista et al., 2005; Hinman et al., 2006; D oerner et al., 2007; Macpherson et al., 2007; Z urborg et al., 2007; Wang et al., 2008; A ndersson et al., 2009; Hu et al., 2009; Hu et al., 2010] . In addition, previous studies identified T R PA 1 as a receptor for parabens-mediated irritation of the skin [ F ujita et al., 2007] and for pain produced by alkaline pH [ F ujita et al., 2008] . On the other hand, menthol has different effects on T R PA 1 in humans and mice. Previous studies identified the bimodal action of several compounds such as menthol on mouse T R PA 1 (mT R PA 1) gating: submicromolar to low micromolar concentrations of menthol caused robust channel activation, whereas higher concentrations led to reversible channel blocking [ K arashima et al., 2007; X iao et al., 2008] . S uch bimodal action was not observed with human T R PA 1 (hT R PA 1) [ X iao et al., 2008] . T R PA 1 is reportedl y involved in inflammatory processes, including inflammation produced by several airway irritants that cause asthma [ B autista et al., 2006; C aceres et al., 2009; Z hou et al., 2013] and neuropathic pain [ Z hou et al., 2013] . T herefore, T R PA 1 is an excitatory ion channel targeted by acute nociception and inflammatory pain and is considered to be a promising target for the development of analgesic agents [ Nagata et al., 2005; X u et al., 2005; K lionsky et al., 2007; E id et al., 2008; McGaraughty et al., 2010; C hen et al., 2011] . T here are
several reported T R PA 1 antagonists: camphor obtained from the C innamonum camphora tree [ X u et al., 2005] , HC -030031 [ E id et al., 2008] , A Z 868 [ V allin et al., 2012] , A -967079 [C hen et al., 2011] , and C MP1, C MP2 and C MP3 that were identified as thioaminal-containing molecules [ C hen et al., 2008] . A mong these T R PA 1 antagonists, naturall y occurring analgesic compounds that inhibit hT R PA 1 and have a demonstrated safety profile from their lengthy usage would be desirable.
T R PM8 is a thermosensitive receptor that detects cool temperatures and menthol [ McK emy et al., 2002; Peier et al., 2002] , a natural non-reactive cooling compound that is also involved in anti-nociception to some extent [ D haka et al., 2007; Proudfoot et al., 2006] . Menthol (the main ingredient of peppermint) is used for pain relief in dail y life. It functions through T R PM8 activation [ Proudfoot et al., 2006; Galeotti et al., 2002] ; however, the molecular mechanisms underlying its analgesic effects are largel y unknown. Moreover, high doses of menthol cause sensory irritation [ Geen et al., 1992] because it acts as a T R PA 1 activator in humans [ K arashima et al., 2007] . C amphor, another essential oil component, is now known to exert analgesic effects, most likely through inhibition of T R PA 1 [ X u et al., 2005] and activation of T R PM8 [ Vogt-E isele et al., 2007] . However, camphor is not suited for use as an analgesic compound because it causes warm and hot sensations [ Green et al., 1990] , probably through T R PV 1 and T R PV 3 activation [ X u et al., 2005]. T herefore, I hypothesized that an effective analgesic compound would activate T R PM8 and inhibit T R PA 1, but not activate T R PV 1.
S everal T R P channels are known to be activated or inhibited by plant-derived substances, such as menthol and camphor, some of which are contained in essential oils. E ssential oils have long been used and their side effects are generall y considered to be
minimal. A ccordingl y, essential oils, especiall y ones acting on T R P channels, could be promising sources for the development of analgesic agents. T herefore, I have screened essential oils for their ability to activate human T R PM8 (hT R PM8) but not hT R PA 1, properties distinct from those of menthol. Moreover, I examined the effects of menthol on hT R PV 1 to clarify the mechanisms of its analgesic effect.
T hrough the screening, I found that eucal yptus oil exhibited relatively high hT R PM8-activating ability with comparativ ely less activation of hT R PA 1. F urthermore, 1,8-cineole, a main component of eucal yptus oil, was identified as a novel natural antagonist of hT R PA 1. I screened camphor analogs to find more effective T R PA 1 antagonists. F rom the screening, borneol, 2-methylisoborneol, and fenchyl alcohol were found to exhibit higher inhibitory effects than camphor and 1,8-cineole. In addition, the hT R PV 1-mediated currents induced by capsaicin were inhibited by menthol.
2 M ater ials and M ethods
2.1 M olecular cloning
F ull-length human genes T RPA1, T RPM8, T RPV 1, and T RPV 2 were obtained from L ife Technologies (C arlsbad, C A , US A ) and T RPV 3 was generousl y provided by D r. Hwang (K orea University) . cD NA s were cloned into the pcD NA 3.1 vector.
2.2 R eagents
C amphor, borneol, fenchyl alcohol, and 2-methylisoborneol were obtained from Wako Pure C hemical Industries L td. (Osaka, J apan). (-)-F enchone, 1,8-cineole, camphorquinone, norcamphor, α ,β-thujone, α -pinene oxide, (-)-limonene oxide, (+)-borneol, (-) -borneol and (±)-isobornyl methyl ether were obtained from S igma-A ldrich (St. L ouis, MO, US A ). B ornyl acetate, (±)-isoborneol and 3-methylene-2-norbornanone were obtained from Tokyo K asei C o. L td. (Tokyo, J apan). T he compounds were used as a mixture of (+) and (-) isomers unless otherwise stated.
2.3 C ell culture
Human embryonic kidney-derived 293T (HE K 293T ) cells were maintained in D ME M (WA K O Pure C hemical Industries, L td., Osaka, J apan) supplemented with 10% F B S (B iowest S A S , C aille, F rance), 100 units/mL penicillin ( L ife Technologies C orp., C arlsbad, C A , USA ), 100 µ g/mL streptomycin ( L ife Technologies C orp.) , and 2 mM L -glutamine (GlutaMA X , L ife Technologies C orp.) at 37°C in 5% C O2. F or C a
2+
-imaging, 1 µ g plasmid D NA carrying T RPA1, T RPV 1, T RPV 2, T RPV 3 or T RPM8 in pcD NA 3 in OPT I-ME M medium ( L ife Technologies C orp.) were transfected into HE K 293T cells using L ipofectamine Plus R eagent ( L ife Technologies C orp.). A fter incubating for 3 to 4 h, the cells were reseeded on coverslips and further incubated at 37°C in 5% C O2.
2.4 H uman subj ects
J apanese male subjects in their 20s and 30s were selected as participants to eliminate confounding factors that may have influenced the perception of skin sensitivity, including race, age, gender, and hormonal and psychosocial interactions. T o evaluate sensory irritation, I selected skin-sensitive male volunteers. F emale volunteers were excluded because of possible hormonal influences. E thics approval and informed consent was obtained from all participants.
2.5 T R PA 1, T R PV 1 and T R PM8 mutants
Two T RPA1 mutants (S873V /T 874L and Y 812A ) were constructed by double and single amino acids substitutions using a GeneTailor S ite-D irected Mutagenesis S ystem ( Invitrogen) [ X iao et al., 2008] . T hree T RPV 1 mutants (Y 511A , S 512Y and T 550I) were made using a modified QuickC hange S ite-D irected Mutagenesis method (A gilent Technologies Inc., S anta C lara, C A , US A ) [ J ordt et al., 2002; Gavva et al., 2004; S hintaku et al., 2012] . T hree T RPM8 mutants (Y 745H, Y 1005F and L 1009R ) were constructed by single amino acid substitutions using a GeneTailor S ite-D irected Mutagenesis S ystem ( Invitrogen) [ B andell et al., 2006; B enedikt et al., 2007; F ujita et al., 2013] . In each case, the entire sequence (including the desired substitutions at the site of mutation) was confirmed.
2.6 T r ansfection
C ells were transfected using one mg of T RPA1, T RPV 1, T RPV 2 or T RPM8 cD NA with L ipofectamine and OPT I-ME M I R educed S erum Medium ( Invitrogen, C arlsbad, C A ) as described previousl y (C aterina et al., 1997). F or patch clamp experiments, cells were co-transfected with these plasmids and green fluorescent protein (GF P) in pcD NA 3.1. G reen fluorescence from cells expressing G F P was detected with the aid of a Nikon (Tokyo, J apan) microscope equipped with a mercury lamp light source and a G F P filter ( emission wavelength, 510 nm).
2.7 C alcium imaging
C a
2+
-imaging was performed 1 day after transfection. HE K 293T cells on coverslips were mounted in an open chamber and superfused with standard bath solution (140 mM NaC l, 5 mM K C l, 2 mM MgC l2, 2 mM C aC l2, 10 mM HE PE S , 10 mM glucose, pH 7.4). C ytosolic-free C a
2+
concentrations in HE K 293T cells were measured by dual-wavelength fura-2 ( Molecular Probes, Invitrogen C orp.) microfluorometry with excitation at 340/380 nm and emission at 510 nm. T he fura-2 ratio image was calculated and acquired using the IP-L ab imaging processing system (Scanalytics Inc., F airfax, VA US A ).
2.8 E lectrophysiology
W hole-cell patch-clamp recordings were performed 1 day after transfection. T he standard bath solution was the same as that used in the C a
2+
-imaging experiments, but extracellular C a
2+
was omitted and 5 mM E GTA was added for the recording of A IT C -, menthol- and F FA -induced current responses. T he pipette solution contained 140 mM K C l, 5 mM E GTA , 10 mM HE PE S , pH 7.4 (adjusted with K OH). D ata from whole-cell voltage-clamp recordings were sampled at 10 kHz and filtered at 5 kHz for anal ysis (A xon 200B amplifier with pC L A MP software, A xon Instruments, S unnyvale, C A , USA ). Membrane potential was clamped at -60 mV with 5 sec voltage ramp-pulses from -100 to +100 mV (500 ms). A ll experiments were performed at room temperature.
2.9 S ensor y ir r itation tests
T he study was conducted at a temperature of 21–23°C and a relative humidity of 45-55%. I used the side of the neck, which is innervated by spinal nerves, for the skin irritation analysis because this region is known to be sensitive to various skin irritants. A reas of skin were cleaned with a wet towel and acclimatized for 10 min prior to testing. B lind randomized half-region ( left vs. right) trials were performed with two different samples applied to the neck region. A total of 200 µ L of 1,8-cineole or 80 µ L of menthol was applied. T he subjects evaluated pricking, stinging, burning and itching sensations after 1, 3, 5, 7 and 10 min of compound/chemical application in accordance with the criteria summarized in Table 1. T he total sensory irritation scores were calculated for the entire period.
2.10 D ata analysis
D ata in all figures are shown as means ± standard error of the mean (S E M) . S tatistical significance of the effects of 1,8-cineole on several T R P channels were evaluated using A NOV A followed by two-tailed multiple t-test with B onferroni correction. T he statistical significance of effects of menthol and capsaicin on hT R PV 1 and hT R PM8 mutants were evaluated using S tudent’s t test. D ose-dependent curves were fit with a Hill equation. S ensory irritation tests were evaluated using a W ilcoxon signed-rank test.
3 R esults
3-1. S creening essential oils for the ability to activate hT R PM 8 but not hT R PA 1 3-1-1. E ucalyptus oil possessed hT R PM 8-activating ability but little capacity to activate hT R PA 1
In order to find promising essential oils that could act as analgesics, I evaluated the ability of individual oils (0.01%) to activate hT R PM8 or hT R PA 1. T he results were compared with the response to 1 mM menthol. T his was accomplished by using a C a
2+
-imaging method with HE K 293T cells expressing hT R PM8 or hT R PA 1 proteins. F or example, the effect of peppermint oil is presented as the fura-2 ratio (corresponding to cytosolic C a
2+
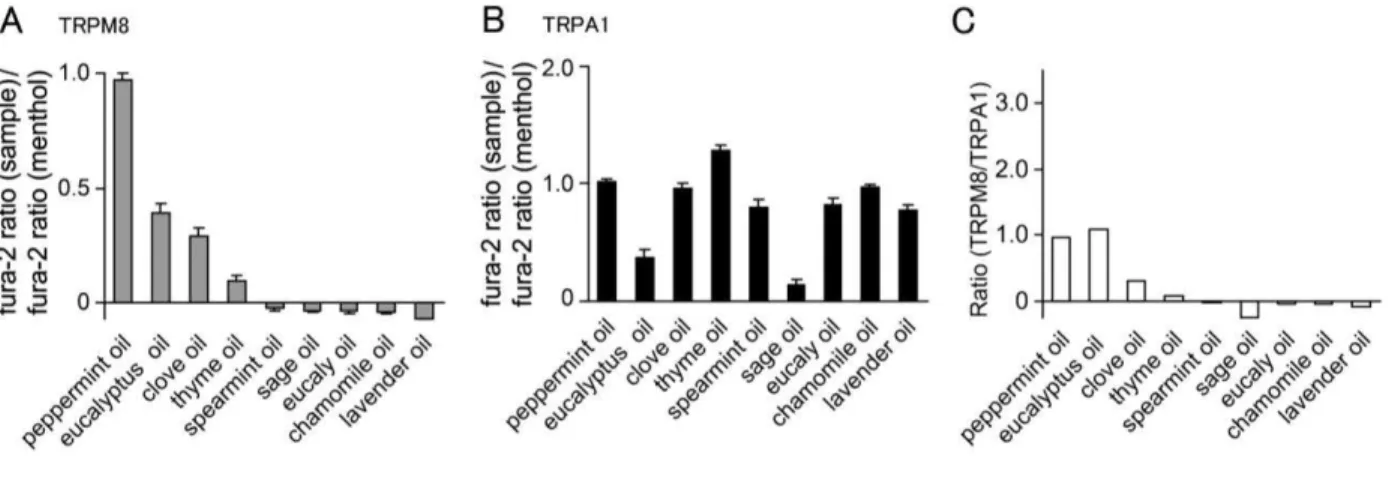
concentrations) compared to that caused by menthol, the main component of peppermint oil. I observed a value of nearl y 1.0. A mong the essential oils examined, clove oil and eucalyptus oil were found to exhibit some hT R PM8 activation ( F igure 1A ). A lthough many of the examined essential oils exhibited hT R PA 1 activation (like menthol) sage oil and eucalyptus oil showed less hT R PA 1 activation (F igure 1B ). W hen I calculated the ratio of hT R PM8-activating ability versus hT R PA 1-activating ability by simply dividing the values in F igure 1A by the values in F igure 1B , the ratio of eucalyptus oil was comparable to that of peppermint oil and much higher than any other oil examined ( F igure 1C ) .
3-1-2. 1,8-cineole activated hT R PM 8 but not hT R PA 1
Next, I examined the effects of fragrance chemicals, many of which are contained in the essential oils tested above. I compared the same parameters used for the essential oils. A s shown in F igure 2A and B , 1,8-cineole, menthone and eugenol showed relativel y
high responses compared with other chemicals as measured by the sample/menthol fura-2 ratio in HE K 293T cells expressing hT R PM8. T his is consistent with the observation that peppermint oil, clove oil and eucalyptus oil, which contain menthone, eugenol and 1,8-cineole, respectively, showed similar high ratios ( F igure 1A ). Interestingly, 1,8-cineole, but neither menthone nor eugenol, showed low changes in the fura-2 ratio in HE K 293T cells expressing hT R PA 1 (F igure 2C and D ). A lthough linalool was reported to produce anti-nociception [ Peana et al., 2003] , the ratio of hT R PM8-activating ability versus hT R PA 1-activating ability for linalool was found to be low, suggesting it was a poor candidate as an analgesic. A ccordingly, the ratio of 1,8-cineole was found to be very high as shown in F igure 2E . T hese data indicated that 1,8-cineole contained in eucal yptus oil can activate hT R PM8 without activating hT R PA 1.
3-1-3. 1,8-cineole acted on hT R PM 8 and hT R PV 3, but not on hT R PV 1, hT R PV 2 or hT R PA 1
In order to examine whether 1,8-cineole could activate other T R P channels expressed in sensory neurons, I performed C a
2+
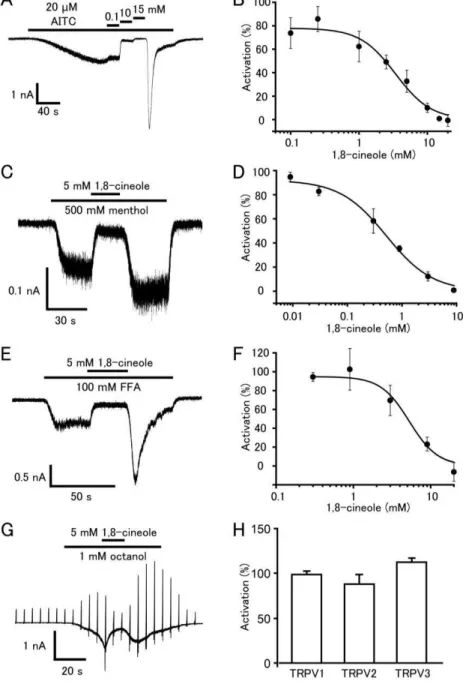
-imaging experiments using HE K 293T cells expressing hT R PV 1 or hT R PV 2 [ C aterina et al., 1997; C aterina et al., 1999; C aterina et al., 2000; McK emy et al., 2002; Peier et al., 2002; B ehrendt et al., 2004; B autista et al., 2006; K wan et al., 2006; Proudfoot et al., 2006; C olburn et al., 2007] . T reatment with 1,8-cineole increased the fura-2 ratio (340 nm/380 nm) in HE K 293T cells expressing hT R PM8, but not in cells expressing hT R PA 1, hT R PV 1 or hT R PV 2 ( F igure 3A , B and D ). T hese results were consistent with previous findings in rodent cell lines expressing these proteins. B ecause 1,8-cineole, like menthol, was reported to activate
mouse T R PV 3 in a X enopus oocyte expression system [ Vogt-E isele et al., 2007; S herkheli et al., 2009; S aito et al., 2011] , I checked the effect of 1,8-cineole on hT R PV 3. B asal fura-2 ratio levels were slightly higher for hT R PV 3-expressing HE K 293T cells compared to cells expressing hT R PM8, hT R PA 1, hT R PV 1 or hT R PV 2, probabl y because hT R PV 3 can be activated by the warm temperatures to which the cells were exposed during incubation (F igure 3C ) . 1,8-cineole caused a small but significant fura-2 ratio increase as expected (F igure 3C , D ).
3-1-4. 1,4-cineole activated hT R PM 8 and hT R PA 1 expressed in H E K 293T cells S ince eucal yptus oil contains not only 1,8-cineole but also 1,4-cineole, and because these chemicals have similar structures ( F igure 4A ), the actions of 1,4-cineole on hT R PM8 and hT R PA 1 were assessed using a C a
2+
-imaging method. S urprisingly, 1,4-cineole (5 mM) caused increases in the fura-2 ratio not only in cells expressing hT R PM8 (F igure 4B ), but also in cells expressing hT R PA 1 (F igure 4C ) . T hese results might explain the apparent difference in the effects between eucal yptus oil and 1,8-cineole regarding the ratio of hT R PM8-activating versus hT R PA 1-activating abilities (F igs. 1C and 2E ) in which the ratio for 1.8-cineole was very low while the ratio for eucal yptus oil was comparable to that of peppermint oil.
3-1-5. 1,4-cineole but not 1,8-cineole activated hT R PA 1
Next, I performed patch-clamp experiments to confirm the effects of 1,8-cineole and 1,4-cineole on hT R PM8 and hT R PA 1 expressed in HE K 293T cells. B oth 1,8- and 1,4-cineole (5 mM) evoked inward currents with outwardly rectifying current-voltage ( I-V ) relationships in cells expressing hT R PM8 (F igure 5A , C ). On the other hand,
1,8-cineole (5 mM) did not activate hT R PA 1 in cells responding to A IT C (a T R PA 1 agonist) whereas 1,4-cineole evoked an inward current with an outwardl y rectifying I-V relationship in cells expressing hT R PA 1 ( F igure 5B , D ). T hese results showed that chemicals with similar structures exhibited different effects on hT R PA 1. Menthol has bimodal action on mT R PA 1: lower concentrations of menthol activate mT R PA 1 whereas higher concentrations of menthol inhibit it [ X iao et la., 2008] . To confirm that low concentrations of 1,8-cineole activated hT R PA 1, I performed patch-clamp experiments. Ten mM and 100 mM 1,8-cineole did not activate hT R PA 1 in cells responding to A IT C (data not shown).
3-1-6. 1,8-cineole inhibited hT R PA 1 cur rents activated by different agonists
B ecause 1,8-cineole has analgesic and anti-inflammatory effects in vivo [ S antos et al., 2000; S ilva et al., 2003] , I hypothesized that 1,8-cineole would inhibit T R PA 1 [ B autista et al., 2006] . To test this hypothesis, I used A IT C (20 mM), which is a T R PA 1 agonist that acts through covalent cysteine modification [ Hinman et al., 2006; Macpherson et al., 2007] . I observed that induced hT R PA 1 currents were inhibited by 1,8-cineole in a dose-dependent manner with an IC50 value of 3.4 ± 0.6 mM (F igure 6A , B ). T he effects of 1,8-cineole on hT R PA 1 activated by other T R PA 1 agonists were also determined. S everal T R PA 1 agonists with different activation mechanisms were chosen: (1) menthol, which seems to interact specifically with residues within transmembrane domain 5 to gate T R PA 1 [ X iao et la., 2008] ; (2) flufenamic acid (F FA ), thought to be a cysteine-nonreactive compound [ Hu et al., 2010] ; and (3) octanol, the mechanism of action of which is largel y unknown [ K omatsu et al., 2012] . F or menthol- or F FA -evoked hT R PA 1 currents, I measured the current responses in the
absence of extracellular C a
2+
to minimize desensitization, similar to the experiment examining the effects on A IT C -evoked currents. F or examination of octanol-evoked hT R PA 1 currents, I performed patch-clamp recordings in the presence of extracellular C a
2+
because octanol-evoked responses were too small to analyze in the absence of extracellular C a
2+
(data not shown), thus leading to difficulties in plotting a dose dependency curve. S imilar to hT R PA 1 currents activated by A IT C , hT R PA 1 currents activated by menthol (500 mM) or F FA (100 mM) were inhibited by 1,8-cineole in a dose-dependent manner with IC50 values of approximately 0.5 ± 0.1 and 5.3 ± 0.1 mM, respectivel y ( F igure 6C , D , E , F ). A t 1 mM, octanol-evoked hT R PA 1 currents were inhibited reversibl y by 1,8-cineole (5 mM) ( F igure 6G). T hese results again suggested that 1,8-cineole was an antagonist of hT R PA 1. Interestingl y, currents evoked by A IT C , menthol or F FA were increased upon washout of 1,8-cineole (F igure 6A , C and E ) probably through the reversal of 1,8-cineole-mediated inhibition. I confirmed that 1,8-cineole did not inhibit hT R PV 1, hT R PV 2 or hT R PV 3 responses activated by capsaicin, 2-A PB and a cocktail (2-A PB + carvacrol), respectively (F igure 6H).
3-1-7. 1,8-cineole inhibited sensor y ir r itation caused by hT R PA 1 agonists in vivo To confirm the inhibitory effect of 1,8-cineole on hT R PA 1 in vivo, sensitive human subjects were recruited for sensory irritation tests. S ensory irritation caused by 1,8-cineole itself was comparable to that by vehicle alone (F igure 7A ,B ), indicating that 1,8-cineole did not cause sensory irritation. Next, I examined the inhibitory effect of 1,8-cineole with concomitant application of the T R PA 1 agonist octanol [ K omatsu et al., 2012] and 1,8-cineole. Octanol (0.2 wt%) caused sensory irritation for the 7 min , with
a gradual increase thereafter (F igure 7C ), as was previously reported [ V an T hriel et al., 2003] . T he octanol-induced sensory irritation was significantl y reduced by concomitant application of 1,8-cineole at the 7-min time point (F igure 7C ). A nalysis of the total sensory irritation score indicated that 1,8-cineole significantly inhibited sensory irritation caused by octanol ( F igure 7D ). T hese data further support the inhibitory action of 1,8-cineole on octanol-induced irritation, which could involve regulation of T R PA 1 activity. Menthol is known to activate both T R PM8 and T R PA 1, which can simultaneously cause comforting and irritating sensations. In order to examine whether 1,8-cineole can reduce the menthol-induced irritation, I applied menthol with or without 1,8-cineole. C oncomitant application of menthol with 1,8-cineole significantly reduced irritation, probably through inhibition of T R PA 1 by 1,8-cineole ( F igure 7E , F ) .
3-2. S creening camphor analogs to find more effective T R PA 1 antagonists 3-2-1. S creening of natur ally occur r ing compounds affecting hT R PA 1
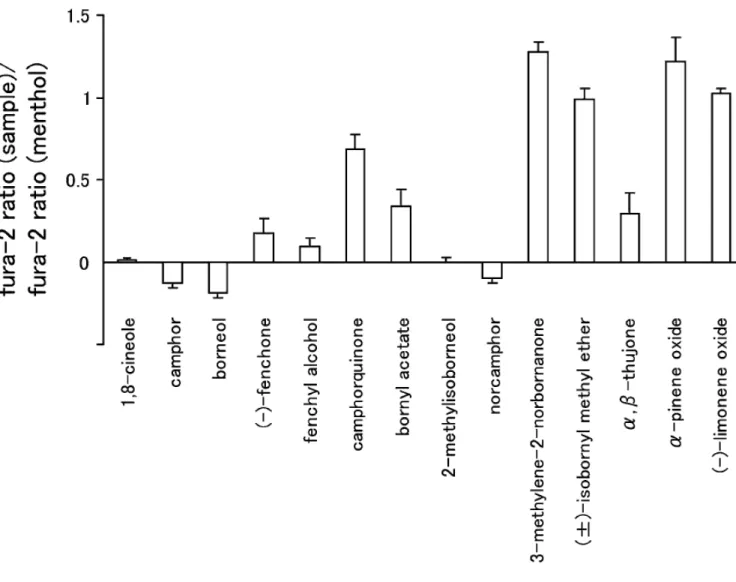
S everal compounds with similar structures exhibit different effects on hT R PA 1. F or example, menthol and 1,4-cineole activate hT R PA 1 while camphor and 1,8-cineole inhibit hT R PA 1 [ Takaishi et al., 2012] . Given these variable effects on hT R PA 1, more detailed anal yses could lead to a better understanding of the structural basis for the action of these compounds on T R PA 1. To identify more effective T R PA 1 antagonists, I next screened camphor analogs, many of which are present in essential oils (Table 2). Towards that end, I used a C a
2+
-imaging method with HE K 293T cells expressing hT R PA 1. In the experiments, changes in the fura-2 ratio induced by the test compounds and menthol were compared because menthol, which activates hT R PA 1,
and the test compounds are members of the monoterpene famil y. B orneol, 2-methylisoborneol, norcamphor, and fenchyl alcohol showed small changes in the fura-2 ratio similar to 1,8-cineole and camphor ( F igure 8). T hose data suggests that these compounds did not activate hT R PA 1.
3-2-2. E ffects of bor neol, 2-methylisobor neol, fenchyl alcohol and norcamphor on hT R PA 1-mediated cur rent responses
In order to confirm the effects of the above compounds, patch-clamp experiments with HE K 293T cells expressing hT R PA 1 were performed. A s shown in F igure 9, borneol, 2-methylisoborneol, fenchyl alcohol and norcamphor failed to activate hT R PA 1, whereas A IT C evoked a robust current activation with outward rectification.
3-2-3. B or neol, 2-methylisobor neol, and fenchyl alcohol (but not norcamphor ) inhibited hT R PA 1 in a C a
2+
-imaging method
I next investigated the possibility that the above four compounds inhibited hT R PA 1 using a C a
2+
-imaging method with HE K 293T cells expressing hT R PA 1. Increases in the fura-2 ratio caused by menthol were almost completely blocked in the presence of borneol (1 mM) , 2-methylisoborneol (1 mM) or fenchyl alcohol (1 mM) similar to that observed with 1,8-cineole and camphor (F igure 10A -C , E ). Washing out the three compounds after menthol exposure led to small increases in the fura-2 ratio, which could be due to the loss of inhibition and resumption of hT R PA 1 activity from residual menthol in the cell. On the other hand, norcamphor did not inhibit the menthol-induced increase in the fura-2 ratio ( F igure 10D , E ). It was confirmed that borneol, 2-methylisoborneol, fenchyl alcohol and norcamphor had no effect on
vector-transfected cells, while cells responded normally to ionomycin (5 mM) ( F igure 11). T hese results suggest that borneol, 2-methylisoborneol and fenchyl alcohol, but not norcamphor, inhibited hT R PA 1 activity.
3-2-4. B or neol, 2-methylisobor neol and fenchyl alcohol inhibited hT R PA 1 cur rent in a dose-dependent manner
To confirm the inhibition of hT R PA 1 activity by the three compounds, patch-clamp experiments were performed with HE K 293T cells expressing hT R PA 1. T he current response was measured in the absence of extracellular C a
2+
to minimize desensitization. One mM borneol, 2-methylisoborneol and fenchyl alcohol completely inhibited the hT R PA 1-mediated current activated by menthol (1 mM) or F FA (100 mM) (F igure 12). E nhancement of the hT R PA 1-current was observed upon washing out of the three compounds in response to F FA , but not menthol.
Next, the effective inhibitory concentrations of the three compounds were determined. F or this experiment, A IT C was chosen as the hT R PA 1 agonist because A IT C had a higher ability to activate hT R PA 1 than did menthol or F FA . One mM borneol, 2-methylisoborneol, and fenchyl alcohol completel y inhibited the A IT C -induced (20 mM) hT R PA 1 current, while 1 mM camphor partially inhibited the A IT C -induced response (F igure 13A -D ), suggesting that borneol, 2-methylisoborneol and fenchyl alcohol were more capable of inhibiting hT R PA 1 than was camphor. E nhancement of the hT R PA 1-current was again observed upon washing out of the three compounds.
T he dose-dependency of the inhibitory effects of the three compounds on hT R PA 1 was then examined using a patch-clamp method. S imilar to camphor and 1,8-cineole,
the hT R PA 1 current induced by A IT C (20 mM) was inhibited by borneol, 2-methylisoborneol and fenchyl alcohol in a dose-dependent manner with IC50 values of 0.20 ± 0.06 mM, 0.12 ± 0.03 mM and 0.32 ± 0.06 mM, respectively. T hose values were much lower than the IC50 of 1,8-cineole (3.43 ± 0.58 mM) and camphor (1.26 ± 0.32 mM) (F igure 13E ). T hese data suggested that borneol, 2-methylisoborneol and fenchyl alcohol have potential to be used as effective analgesic compounds. C ommercially available borneol contains both optical isomers. I confirmed that there was no difference in the inhibitory effects on hT R PA 1 activity between (+)- and (-)-borneol ( F igure 14).
3-2-5. T he hydroxyl group of bor neol contr ibuted to inhibition of human T R PA 1 X iao et al. showed that menthol acted as an activator of mT R PA 1 at low concentrations, and as a blocker at high concentrations [ X iao et al., 2008] . T his bimodal effect of menthol on T R PA 1 was observed in a mouse clone, but not in a human clone. T hat author studied a mouse T R PA 1 mutant in which serine and threonine residues located in the predicted inner side of T M5 were replaced with valine and leucine (S 876V /T 877L ), respectivel y. It was found that the mutant was neither activated nor inhibited by menthol [ X iao et al., 2008] . Moreover, the serine and threonine residues were found to be critical for the sensitivity of T R PA 1 to menthol in both mammalian T R PA 1 channels. It was suggested that T 877 of mT R PA 1 interacted with menthol through a hydrogen bond [ X iao et al., 2008] . T he fact that borneol, 2-methylisoborneol and fenchyl alcohol have similar hydroxyl groups in their structures led us to hypothesize that these compounds interacted with the same serine and/or threonine of T R PA 1 as did menthol. To test this hypothesis, I investigated whether the serine and threonine residues were
involved in the inhibitory effect of borneol. T he mutant channel hT R PA 1-S 873V /T 874L (corresponding to mT R PA 1-S 876V /T 877L ) exhibited significantl y lower inhibitory effects at three different concentrations of borneol than did hT R PA 1-W T when A IT C -activated current was examined (F igure 15A ). On the other hand, camphor and 1,8-cineole showed no significant changes in their inhibitory effects with cells expressing mutant hT R PA 1 compared with hT R PA 1-W T (F igure 15B , C ). I asked whether there was involvement of other amino acids in borneol activity. B ecause a tyrosine residue in T M2 of T R PM8 is known to be involved in interactions with menthol, I screened tyrosine mutants in T M2 and T M3 of hT R PA 1 using a C a
2+
-imaging method and found that the effect of borneol was reduced with a Y 812A mutant (data not shown) . T he mutant (hT R PA 1-Y 812A ) showed significantl y lower inhibitory effects of borneol on the A IT C -activated hT R PA 1 current at two different concentrations than did hT R PA 1-W T with a patch-clamp method, whereas such differences between W T hT R PA 1 and hT R PA 1-Y 812A were not observed for 1,8-cineole or camphor activity (F igure 15D -F ).
3-3. T he effects of menthol on human T R PV 1
3-3-1. M enthol inhibited sensor y ir r itation caused by an hT R PV 1 agonist in vivo Menthol has been used for its anti-nociceptive effects for more than one thousand years [ V an Hellemont et al., 1985] . Preparations containing menthol are used topically to relieve neuralgia in traditional C hinese and E uropean medicine [ Wright., 1870] . In addition, menthol demonstrated some anesthetic [ Galeotti et al., 2001; McK ay et al., 2006; Gaudioso et al., 2012] and k -opioid-mediated anti-nociceptive properties in mouse hot-plate tests [ Galeotti et al., 2002]. A t present, the anti-nociceptive
mechanisms of menthol are not fully understood. It has been shown that T R PM8 contributed to mediating the effects of cold analgesia in the rat [ Proudfoot et al., 2006] and that it had anti-nociceptive effects in T R PM8-deficient mice [ D haka et al., 2007] . S ince T R PV 1 acts as an integrator of painful stimuli, T R PV 1 antagonists can be used for promising therapeutics as novel types of analgesics [ Gavva et al., 2008; Gunthorpe et al., 2009; Marcello et al., 2010; Marcello et al., 2013] . T herefore, to understand the analgesic mechanism of menthol, I examined the effects of menthol on hT R PV 1. F irst, to examine the analgesic effects of menthol in vivo, sensitive human subjects were recruited for the sensory irritation test. In this study, I used V B E , which is structurally similar to capsaicin [ Mitsudome et al., 2012] . I observed that V B E activated hT R PV 1 ( F igure 16A ), and the activation was inhibited by capsazepine (1 µ M) (F igure 16B ). V B E did not cause any current activation, but rather decreased basal currents in the vector-transfected cells ( F igure 16C ). I confirmed that menthol (5 mM) inhibited the hT R PV 1 activity evoked by V B E (100 µ M) (F igure 16D and E ). T hus, V B E and menthol might allow us to examine the inhibitory effect of menthol on sensory irritation by V B E . V B E (0.1 wt %) caused sensory irritation, graduall y increasing with time after application (F igure 17A , C ). V B E -induced sensory irritation scores were not reduced by concomitant application of 0.1 wt% menthol. However, the scores were significantly reduced by 0.3 wt% menthol (F igure 17A - D ). In human subjects, inhibition of V B E -induced irritation by menthol was smaller than T R PV 1 inhibitory effects observed in vitro. I conjectured that V B E might activate hT R PA 1 as well. Indeed, V B E had a hT R PA 1-activating ability, although the effects were observed only at 1 mM whereas V B E activated hT R PV 1 at lower concentrations (F igure 18).
3-3-2. M enthol inhibited hT R PV 1 activity induced by capsaicin
Next, I examined the inhibitory effects of menthol on hT R PV 1 using a C a
2+
-imaging method at room temperature with HE K 293T cells expressing hT R PV 1. In these cells, the changes in the fura-2 ratios (i.e., cytosolic C a
2+
concentrations, [ C a
2+
]i) induced by capsaicin (0.1 µ M) in the presence of menthol (10 mM) were smaller than those in its absence ( F igure 19). A lthough there was a slight increase in [ C a
2+
]i upon application of menthol (10 mM) itself, I did not observe any current activation induced by 10 mM menthol in the cells expressing hT R PV 1 nor vector-transfected cells using a patch-clamp method (F igure 20A and B ), indicating that the slight [ C a
2+
]i increase could be a non-specific phenomenon. T hese results suggested that menthol inhibited hT R PV 1 activity.
To confirm these possibilities, I performed patch-clamp experiments using HE K 293T cells expressing hT R PV 1. C apsaicin (0.1 µ M) induced T R PV 1-mediated current activation with an outwardl y rectifying current-voltage ( I-V ) relationship revealed by 5 sec ramp-pulses from -150 to +100 mV. A ddition of menthol (5 mM) reduced the capsaicin-evoked currents at both positive and negative potenti als, and these were partl y recovered by menthol washout ( F igure 21A ). T he currents evoked by concomitant application of capsaicin with menthol were smaller than those activated by capsaicin alone. T hese results were consistent with the data in the C a
2+
-imaging experiments (F igure 19).
Next, I examined the dose-dependency of the inhibitory effects. T he current responses were measured in the absence of extracellular C a
2+
to minimize desensitization. Menthol inhibited 0.1 µ M capsaicin-activated hT R PV 1 currents in a
dose-dependent manner, with an IC50 value of approximately 1.2 ± 0.2 mM (F igure 21B and C ). In addition, I examined the effects of different concentrations of capsaicin on hT R PV 1 in the absence and presence of 1 mM menthol. I found an apparent shift of the capsaicin dose-dependency towards higher concentrations with menthol (F igure 22). T hese results indicated that menthol inhibited hT R PV 1 activity.
3-3-3. M enthol inhibited the activation of hT R PV 1 induced by heat stimulation Next, I examined the inhibitory effects of menthol on hT R PV 1 activities induced by heat stimulation using a C a
2+
-imaging method. W hen I examined the effects of menthol on heat-evoked (> 45°C ) T R PV 1-mediated [ C a
2+
]i changes, I compared the effects in different cells to avoid the effects of desensitization. T he increases in fura-2 ratios induced by heat stimulation in the absence of menthol (0.54 ± 0.03) were significantl y larger than the increases in fura-2 ratios induced by heat stimulation in the presence of menthol (0.35 ± 0.02, p < 0.01) ( F igure 23A , B ). T hese data suggested that menthol inhibited hT R PV 1 activity induced by heat stimulation. T he increases in fura-2 ratios induced by capsaicin after heat stimulation in the presence of menthol were slow, possibly because menthol effects were not completely abolished by washout. I more thoroughl y examined the inhibitory effects of menthol and capsaicin on the thermal responses of hT R PV 1. T hus, I performed patch-clamp experiments with HE K 293T cells expressing hT R PV 1. Menthol (5 mM) inhibited heat-evoked T R PV 1-mediated currents in a reversible manner (F igure 24A ). T he first heat-evoked currents in the presence of menthol (5 mM) were significantl y smaller than those in the absence of menthol (36.8 ± 7.1 and 123.3 ± 35.5 pA /pF in the presence and absence of
menthol, respectively, p < 0.05) ( F igure 24B ). T hus, both chemical and thermal responses of hT R PV 1 were inhibited by menthol.
I then asked whether the inhibition mediated by menthol affected temperature thresholds. T hus, I assessed the changes in temperature thresholds for hT R PV 1 activation through construction of A rrhenius plots for hT R PV 1-mediated current responses. T he temperature thresholds for hT R PV 1 activation were not significantl y different between the absence and presence of menthol (41.0 ± 1.3 and 41.2 ± 1.6 °C , respectivel y) (F igure 24C , D , E ). T hese data indicated that menthol affected hT R PV 1 current sizes without changes in the temperature thresholds for activation.
3-3-4. C apsaicin inhibited hT R PM 8 activity induced by menthol and cold stimulation
T R PV 1 is activated by elevated temperature ( > 42°C ) whereas cold temperature reportedl y decreases capsaicin-induced T R PV 1 activity. On the other hand, T R PM8 is activated by low temperatures and menthol, whereas heat stimulation suppresses menthol-evoked T R PM8 currents. T hese findings suggested that the effects of specific agents on the T R PV 1 channel and the T R PM8 channel are intricatel y related to one another. T herefore, I examined the effect of capsaicin on hT R PM8.
I first examined the effects of menthol on hT R PV 1 and the influence of capsaicin on hT R PM8 using a C a
2+
-imaging method at room temperature with HE K 293T cells expressing hT R PM8. T he changes in the fura-2 ratios induced by menthol (1 mM) in the presence of capsaicin (1 mM) were smaller than those in its absence in cells expressing hT R PM8 (F igure 25). F ura-2 ratios were slightl y reduced by capsaicin
alone in the cells expressing hT R PM8, probably because hT R PM8 was activated even at room temperature. I also confirmed that capsaicin (1 mM) did not cause current activation in the cells expressing hT R PM8 (F igure 26). T hese results suggested that capsaicin inhibited hT R PM8 activity.
To confirm these possibilities, I performed patch-clamp experiments using HE K 293T cells expressing hT R PM8. T he hT R PM8 currents induced by menthol (500 µ M) were similarly inhibited by capsaicin (100 µ M) in a reversible manner and the currents evoked by concomitant application of menthol with capsaicin were smaller than those activated by menthol alone (F igure 27A ). T hese results were consistent with the data in the C a2+-imaging experiments (F igure 25). Next, I examined the dose-dependency of the inhibitory effects. T he current responses were measured in the absence of extracellular C a
2+
to minimize desensitization. T he capsaicin-induced inhibition of menthol-activated hT R PM8 currents occurred in a dose-dependent manner, with an IC50 value of approximately 39.9 ± 6.4 µ M (F igure 27B , C ). T hese results indicated that capsaicin inhibited hT R PM8 activity.
3-3-5. C apsaicin inhibited the activation of hT R PM 8 induced by ther mal stimulation
Next, I examined the inhibitory effects of menthol and capsaicin on hT R PV 1 and hT R PM8 activities, respectively, by thermal stimulation using a C a
2+
-imaging method. In cells expressing hT R PM8, [C a
2+
]i increases caused by cold stimulation (< 20°C ) in the presence of capsaicin (1 mM) were significantly smaller than [ C a
2+
]i increases in the absence of capsaicin (0.19 ± 0.01 and 0.44 ± 0.01, respectively, p < 0.01) ( F igure 28). I more thoroughl y examined the inhibitory effects of capsaicin on the thermal
responses of hT R PM8. T hus, I performed patch-clamp experiments with HE K 293T cells expressing hT R PM8. C apsaicin (100 µ M) inhibited the cold-evoked T R PM8-mediated currents in a reversible manner (F igure 29A ). F urthermore, the first cold-evoked currents in the presence of capsaicin (100 µ M) were significantly smaller than those in the absence of capsaicin (3.4 ± 0.9 and 22.2 ± 3.4 pA /pF in the presence and absence of capsaicin, respectively, p < 0.01) (F igure 29B ). T hus, both chemical and thermal responses of hT R PM8 were inhibited by capsaicin. I then asked whether the inhibition mediated by capsaicin affected temperature thresholds. However, it was not possible to generate A rrhenius plots for hT R PM8 currents because capsaicin completely inhibited cold-induced hT R PM8-mediated currents ( F igure 29A ).
3-3-6. T he binding site of menthol in T R PV 1 was distinguishable from that of capsaicin, whereas the binding site of capsaicin in T R PM 8 modestly inter acted with that of menthol
Menthol activated T R PM8 and inhibited T R PV 1, whereas capsaicin activated T R PV 1 and inhibited T R PM8. T hese results led us to hypothesize that the two compounds interacted with similar sites on each channel in a competitive manner. T herefore, I examined whether menthol interacted with the same binding site as capsaicin on T R PV 1 and whether capsaicin interacted with the same binding site as menthol on T R PM8. Towards that end, I examined capsaicin and menthol binding sites that had been mutated. T hus, I investigated whether menthol affected the heat-evoked activities of mutant hT R PV 1 and whether capsaicin affected the cold-evoked activities of mutant hT R PM8. C apsaicin was reported to interact with Y 511 and S 512 in the cytosolic region linking
transmembrane domains 2 and 3 of rat T R PV 1 [ J ordt et al., 2002] and with T 550 in the transmembrane domain 4 of Oryctolagus cuniculus [ Gavva et al., 2004] , all of which are conserved in hT R PV 1 (F igure 30A ). T herefore, I examined the effects of menthol on the heat-evoked currents in the three hT R PV 1 mutants (Y 511A , S 512Y and T 550I) under naïve conditions. B ecause the heat-evoked inward currents of Y 511A and S 512Y mutants at -60 mV were very small (F igure 31B , C , E ), I examined the effects of menthol on the currents at both -60 and +100 mV. T he effects of menthol on the heat-evoked currents were larger at -60 mV compared with +100 mV in wild-type (W T ) and T 550I channels (F igure 31A , D , E , F ). T he very small heat-evoked currents in Y 511A and S 512Y channels at -60 mV made precise comparisons difficult. Nonetheless, menthol-induced inhibition of the mutant channels at +100 mV were large (F igure 31B , C , E , F ). T hus, the levels of inhibition were generall y similar between W T and mutants, suggesting that menthol might not share the three amino acids with capsaicin for its inhibitory effects.
In contrast, tyrosine 745 in the middle of transmembrane domain 2 was previously identified as a crucial residue for the menthol-sensitivity of mouse T R PM8 [ B andell et al., 2006] , a domain that is conserved in hT R PM8 (F igure 30B ). A s mentioned above, cold-induced inward currents at -60 mV were very small especiall y in the presence of capsaicin ( F igure 32A - E ), preventing us from comparing the effects at negative potentials. However, inhibition of cold-induced T R PM8-mediated currents by capsaicin appeared smaller at +100 mV in the Y 745H mutant (F igure 32D , F ), and ratios without and with capsaicin were significantly larger in the Y 745H mutant (p < 0.05) (F igure 32G ). C old-induced T R PM8-mediated currents decreased upon washout of capsaicin ( F igure 32A ), which probabl y reflected the inhibition of basal
T R PM8-mediated currents by residual capsaicin remaining in the cells. I examined the involvement of two other amino acids (Y 1005 and L 1009) that were reported to be involved in menthol action. However, neither of the mutants provided currents sufficient for analysis ( data not shown). T hese findings suggested that Y 745 was involved to some extent in the capsaicin-induced hT R PM8 inhibition.
4 D iscussion
4-1. S creening essential oils for the ability to activate hT R PM 8 but not hT R PA 1 T R PA 1 is an excitatory ion channel targeted by pungent irritants such as those from mustard oil and garlic. It is thought to function in diverse sensory processes, including cold nociception and inflammatory pain. T herefore, T R PA 1 is a promising target for the identification of analgesic drugs. A natural analgesic compound that does not accelerate pain signaling is desirable for pharmaceutical or cosmetic pain relief. T R PA 1 antagonists (ruthenium red, HC -030031, A MG5445, A 967079 and camphor) possess analgesic properties [ Nagata et al., 2005; X u et al., 2005; K lionsky et al., 2007; E id et al., 2008; McG araughty et al., 2010; C hen et al., 2011] . Of these, camphor is the only naturall y occurring compound and is often used in cosmetics because it possesses minimal adverse effects. However, camphor is not suited for use as an analgesic compound because it causes warm and hot sensations [ Green et al., 1990] . T hese sensations are mediated through activation of T R PV 1 [ X u et al., 2005; Vogt-E isele et al., 2007] . Moreover, T R PM8 contributes to sensing unpleasant cold stimuli or mediating the effects of cold analgesia [ D haka et al., 2007; Proudfoot et al., 2006] . A lthough menthol, the main ingredient of peppermint, is used for pain relief in daily life through T R PM8 activation [ Proudfoot et al., 2006] , its ability to activate hT R PA 1 restricts widespread use of menthol as an analgesic [ Galeotti et al., 2002]. T herefore, chemicals that activate T R PM8 and inhibit T R PA 1, but do not activate T R PV 1, would be ideal analgesic agents.
I found that the activation of hT R PA 1 (induced by several agonists with different
activation mechanisms) can be inhibited by 1,8-cineole. Moreover, 1,8-cineole activated hT R PM8 and hT R PV 3, but not hT R PA 1, hT R PV 1 or hT R PV 2. It was recentl y shown that both peripheral and central activation of T R PM8 could produce an analgesic effect that specifically reversed the sensitization of behavioral reflexes elicited by peripheral nerve injury [ G aleotti et al., 2002; Proudfoot et al., 2006; D haka et al., 2007] . F rom this point of view, 1,8-cineole appears to be an ideal natural analgesic that activates hT R PM8 and inhibits hT R PA 1.
1,8-cineole acts as an agonist of the T R PM8 channel with lower efficacy and potency (3.4 ± 0.4 mM) on T R PM8 than menthol [ McK emy et al., 2002; Vogt-E isele et al., 2007] . 1,8-cineole also activates the T R PV 3 channel in mice, but not the western clawed frog T R PV 3 [ S aito et al., 2011] . F urthermore, 1,8-cineole inhibits the chemical nociception produced by several irritants, and it has anti-inflammatory efficacy in patients with severe asthma [ J uergens et al., 2003] . T he present study suggests that the known analgesic and anti-inflammatory actions of 1,8-cineole can be attributed to its T R PM8-activating and T R PA 1-inhibiting abilities.
1,8-cineole has a fresh smell and elicits a cooling sensation when ingested or applied to the skin and is a common additive in flavorings, food, mouthwashes and cough suppressants. 1,8-cineole is often used in aromatherapy, as a stimulant in skin baths, by the pharmaceutical industry in drug formulations to enhance percutaneous penetration and as a decongestant and antitussive [ Williams et al., 1991; L aude et al., 1994; L evison et al., 1994] . E xperimental data have shown that 1,8-cineole is an analgesic and anti-inflammatory agent with beneficial effects for patients with severe asthma [ J uergens et al., 2003] . 1,8-cineole inhibits the in vitro formation of prostaglandins and cytokines by stimulated monocytes. However, the molecular
targets and mechanisms of the analgesic effect of 1,8-cineole remain unclear [ J uergens et al., 2004] .
In a human study, I examined sensitive volunteers to determine whether 1,8-cineole could inhibit sensory irritation caused by octanol and menthol. In the cosmetic research field, both menthol and octanol are well-known to cause skin irritation, and neither cinnamaldehyde nor allicin is used for human skin studies. 1,8-cineole activated T R PM8 less efficiently than did menthol, and it inhibited menthol-evoked skin irritation. T hose results clearly suggested that the inhibitory effects of 1,8-cineole are probably due to inhibition of T R PA 1 and not activation of T R PM8.
T he inhibitory effects of 1,8-cineole on menthol-induced hT R PA 1 activation was a little greater than those for A IT C - or F FA -induced hT R PA 1 activation ( F igure 6B , D , F ). Menthol has bimodal action through transmembrane domain 5 of T R PA 1 in some species [ X iao et al., 2008] . T herefore, the similarity between the molecular structures of menthol and 1,8-cineole (F igure 4A ) suggests that 1,8-cineole could act on the same domain of T R PA 1 as menthol, although the structural basis for menthol-evoked hT R PA 1 activation is not known. F our compounds with similar structures (F igure 4A ) exhibited different effects on hT R PM8 and hT R PA 1: i) menthol and 1,4-cineole activated both hT R PM8 and hT R PA 1 [ Macpherson et al., 2006] ; ii) camphor inhibited hT R PA 1 [ X u et al., 2005] ; iii) 1,8-cineole activated hT R PM8 and inhibited hT R PA 1 (F igs. 2-4). T he fact that the four compounds exhibited differing effects on hT R PM8 and hT R PA 1 suggests that more detailed anal yses would lead to a better understanding of the structural basis for the action of these compounds on T R PM8 and T R PA 1.
4-2. S creening camphor analogs to identify more effective T R PA 1 antagonists T R PA 1 is an excitatory ion channel targeted by pungent irritants such as those from mustard oil and garlic. It is thought to function in diverse sensory processes, including nociception and inflammatory pain. T herefore, T R PA 1 is a promising target for the development of analgesic agents. A s described above, I found that 1,8-cineole is a naturally occurring hT R PA 1 antagonist. However, this natural antagonist exhibited weaker inhibitory effects on T R PA 1 activity than other antagonists such as HC -030031, A -967079, and A Z 868. T herefore, identificati on of naturall y occurring compounds with greater inhibitory potency of hT R PA 1 activation has been sought.
B orneol is a bicyclic monoterpenoid alcohol that has been used in foods as an aromatic spice. It is a valuable medical and chemical material that has been used as a folk medicine in C hina and India [ A lmeida et al., 2013] . A dditionally, borneol is a fragrant ingredient used in decorative cosmetics, fine fragrances, shampoos, and other toiletries. Previous studies have shown that borneol has a vasorelaxant effect on the rat thoracic aorta [ S ilva-F ilho et al., 2011] and neuroprotective effects [ L iu et al., 2011] . A lthough borneol has been evaluated for anti-nociceptive and anti-inflammatory activities, the molecular targets and mechanisms of its analgesic effect remain unclear. T he fact that this monoterpene acts as an agonist of the T R PV 3 [ Vogt-E isele et al., 2007] and T R PV 1 [ X u et al., 2005] channels and specificall y inhibits nA C hR -mediated effects in a noncompetitive way [ Park et al., 2003] does not explain its anti-nociceptive effects. 2-methylisoborneol has been implicated as the cause of the muddy odor of fish from C edar L ake, Manitoba. A lgae produce 2-methylisoborneol and geosmin that are responsible for the musty odor [ Masakazu et al., 1988] . F enchyl alcohol, which is a component of several essential oils, is a fragrance ingredient used in decorative
cosmetics, fine fragrances, shampoos, and other toiletries. Moreover, this compound has an inhibitory effect on acetylcholinesterase activity [ Miyazawa et al., 2005]. A lthough borneol is the only terpene known to have anti-nociceptive effects, the two other terpenes identified in this study could have similar effects because all three monoterpenes inhibit hT R PA 1, and their derivatives might function as analgesics. A lpizar et al. reported that camphor exhibited a bimodal effect on mT R PA 1 [ A lpizar et al., 2013] . T he authors observed the inhibition of mouse T R PA 1-mediated basal current with 1 mM camphor, and the current increased as camphor was washed out, suggesting a bimodal action for camphor on mT R PA 1. In addition, 1-5 mM camphor increased the cytosolic C a
2+
concentration upon washing out without any effect during camphor application. I examined the effect of camphor (1 mM) in a C a
2+
-imaging method, but failed to observe changes in the cytosolic C a
2+
concentration (F igure 8). C amphor blocked the menthol-induced increase in cytosolic C a
2+
concentrations (1 mM, F igure 10E ). In addition, camphor inhibited the T R PA 1 current activated by 20 µ M A IT C in a dose-dependent manner ( F igure 13E ). A t present, I do not have an explanation for the apparent differences between the two studies, although a species difference mi ght have caused the different outcomes. T ransient enhancement of T R PA 1 current was observed after washing out of the three test compounds in F FA - or A IT C -evoked responses (F igs. 12 and 13), but not in the menthol-evoked response (F igure 12). S everal agonists have bimodal actions on mammalian T R PA 1, apparentl y involving non-covalent mechanisms. However, A lpizar et al. reported that cinnamaldehyde (a T R PA 1 activator with covalent modification) and camphor, which was thought to be a mammalian T R PA 1 antagonist, also exhibit bimodal actions on mT R PA 1, indicating that bimodal actions could be a more general phenomenon than
previously thought [ A lpizar et al., 2013] . T herefore, the transient enhancement of the hT R PA 1 current upon washing out of the antagonists (F igure 12, 13) might result from bimodal actions of the compounds. Interestingly, enhancement was not observed in the menthol-evoked T R PA 1 response ( F igure 12), possibly because menthol and hT R PA 1 inhibitors (borneol, 2-methyisoborneol and fenchyl alcohol) are all monoterpenes that act at similar sites, including ones that were identified in this study, through their cyclohexyl hydroxyl groups.
A nalogs of camphor have exhibited variable effects on hT R PA 1. B orneol, 2-methylisoborneol and fenchyl alcohol inhibited hT R PA 1. In contrast, norcamphor had no effect on hT R PA 1, and other related compounds activated hT R PA 1 (F igure 8). T he mechanisms of action with hT R PA 1 remain unclear although these monoterpenes have similar molecular structures. B orneol, which was synthesized by chemical reduction of camphor, Inhibited hT R PA 1, and the effects were greater than those of camphor. In addition, fenchyl alcohol, which was synthesized by chemical reduction of fenchone, inhibited hT R PA 1 activity, while fenchone activated hT R PA 1. C ommon structural differences between camphor and borneol and between fenchone and fenchyl alcohol are hydroxyl and carbonyl groups at the same position of their cyclohexane, which suggests that hydrogen bonding plays a pivotal role in the action of these compounds. T his idea is supported by the results from mutation of T 874, an amino acid thought to form a hydrogen bond with menthol, resulting in a reduction in the activity of borneol. In our study, S 873 and T 874 in T M5 and Y 812 in T M3 were found to be involved in the inhibitory effects of borneol. B ecause these two sites are somewhat distant from each other, borneol could fit separately into both sites.